Introduction
Bladder cancer (BC) is the most common malignant
disease in elderly patients throughout the world. BC typically
arises from urinary bladder tissue, where it predominates across
North America and Western Europe. The incidence of BC is more
common in males than in females (1,2). The
primary risk factors for BC consist of aging, smoking, chronic
irritation, family genetic history and chemical exposure (3,4). There
are sophisticated techniques for the treatment of BC, however
previous literature has stated that the treatment of BC remains a
challenge (5,6). Developing economically sustainable and
available agents for the treatment of BC is urgently needed within
the scientific community.
Anthocyanidins are antioxidant polyphenolic
compounds that are found in fruits and vegetables, where they are
refined into anthocyanins (7).
Purple sweet potato (PSP) obtains its unique color due to the
presence of anthocyanins, and therefore it is recognized as a
healthy food and used as a food colorant. Recent studies have
revealed that the purple sweet potato anthocyanin (PSPA) has
various health benefits such as antioxidant, antihypertensive, as
well as liver and retina protective (8–10).
Studies in humans and rats revealed that PSPA is absorbed into the
body, where it is rapidly detected in the blood (11,12).
The antitumor effect of the PSPA has gained attention, since it has
been demonstrated to have an effect in rectal (13), colon (14) and stomach cancer (15). However, the potential function that
PSPA plays in BC remains undetermined.
Molecular-targeted therapy is a hot topic in cancer
prevention, including BC, where it is thought to disrupt the
molecular events associated with tumor proliferation (6). The phosphatidylinositol 3-kinase
(PI3K) pathway is an important signaling pathway within living
cells. This pathway is responsible for regulating cell survival and
cell proliferation (16). The
abnormal activation of the PI3K pathway has been identified in
multiple cancers (17,18). The components involved in this
pathway, including Akt, could serve as possible therapeutic targets
during cancer development (19,20).
The PI3K/Akt is a promising molecular target among the treatments
of BC.
It appears plausible that PSPA could be involved in
the physiological development of BC. The present study aimed to
investigate whether PSPA exerted antitumor effects on BC by
detecting its effect on cell apoptosis and cell cycle arrest of BC
cells in vitro. The underlying molecular mechanisms were
also explored. This study aimed to contribute to the treatment of
BC and enhance the understanding of the possible role that PSPA
plays in cancer prevention.
Materials and methods
PSPA preparation
The PSP cultivars Zi A1, Anhui Zi, Chuanshan Zi,
Jishu 18 and Zi A0 were obtained from the Academy of Agricultural
Sciences in Xuzhou (Jiangsu, China). Based on previously described
methods (21), 5 to 8 mm PSP slices
were dried under the condition of 50°C (moisture content in dry
products <12%). Subsequently, the dry PSP powder was mixed with
acidizing solution (95% ethanol:1 M HCL=9:1, v/v). After 3 h, the
mixture was centrifuged for 20 min at 8000 × g/min. The supernatant
was decreased using petroleum ether (extract:petroleum ether=1:2).
The concentrate was absorbed by a macroporous resin (AB-8) and
eluted by an ethanol solution (containing 0.01% HCL) at a flow rate
of 1 ml/min. The anthocyanin effluent was harvested and
concentrated. The solutions were diluted to perform the spectral
scanning using the UV-vis spectrophotometer model Genesys 10S
(Thermo Fisher Scientific, Inc., Waltham, MA, USA). The absorbance
was assessed at the maximum absorption peak. The total content of
PSPA was calculated according to the following formula: Fw =
(ExV)/(98.2×M) × 100 [E, optical density; V, final volume (ml), M,
sample weight (g), 98.2, the molar extinction coefficient of
anthocyanins].
Cell culture
Bladder cell lines, 5637 and T24, were purchased
from the American Type Culture Collection (ATCC, Manassas, VA,
USA). The cells were cultured in RPM-1640 and McCoy's 5a (ATCC)
medium with 10% FBS, at 37°C in an incubator containing 5%
CO2 atmosphere. The non-cancerous bladder cells,
SV-HUC-1 (ATCC) were cultured in Ham's F-12K medium. When the cells
reached ~80% confluency, PSPA was added for the treatment of BC.
The concentrations and incubation time were stated in the following
assays.
CCK-8 assay
Cell viability was estimated by a CCK-8 kit
(Beyotime Institute of Biotechnology, Haimen, China) according to
the manufacturer's instructions. The 5637 and T24 cells
(1×103 cells/well) were seeded into 96-well plates, and
incubated for 24 h. According to a previous study (2), different doses of PSPA (100, 300, 500,
800 and 1000 µg/ml) were added to the culture medium. The cell
viability was assessed after 24, 48 and 72 h. The optical density
(at 450 nm) was recorded with a microplate reader (Bio-Rad
Laboratories, Hercules, CA, USA).
Assessement of mitochondrial membrane
potential (MMP)
The cells were plated at a density of
1×104 cells/well into 6-well plates, where they were
cultured for 24 h. The cells were treated with various doses of
PSPA (100, 300 and 500 µg/ml) for 48 h. A flow cytometry assay was
employed to assess the MMP with Rhodamine123 fluorescent dye
(Rho123; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). The
deteriorated Rho123 accumulation in the mitochondria, indicated the
collapse of the MMP. The cells were incubated with a Rho123 working
solution in the dark, for 30 min, at 37°C. The cells were then
loaded and the fluorescent signals with an emission of 525 nm were
applied on a FACSCalibur (Becton-Dickinson, Franklin Lakes, NJ,
USA).
Reactive oxygen species (ROS) content
detection
The cells in each group were treated as follows: the
cells were washed and then stained with fluorescent dye
2,7-dichlorofluorescein diacetate (DCFH-DA) (10 µM) (Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 15 min at 37°C. The
DCFH-DA was transformed into the fluorescent DCF via ROS. The
fluorescence intensity denoted the ROS level. The fluorescence was
observed with a fluorescence plate reader (EnVision; PerkinElmer,
Inc., Waltham, MA, USA).
Apoptosis detection
An Annexin V-fluorescein isothiocyanate
(FITC)/propidium iodide (PI) kit (Sigma-Aldrich) was used to
evaluate the apoptosis level in the BC cells. The indicated
treatment with the PSPA followed. The cells were collected and
washed with a PBS working buffer. The collected cells were
incubated with Annexin V-FITC for 15 min, followed by another 10
min with PI in the dark. The apoptosis rate was analyzed on a
FACSCalibur, according to the manufacturer's protocol.
Cell-cycle distribution analysis
The cells were stained with a Cycletest Plus DNA
Reagent kit (Becton-Dickinson). Ethanol was used to fix the cells.
The cells were then incubated with RNase for 30 min at 37°C. The
DNA content was assessed with a FACSCalibur, followed by PI
staining for 15 min at room temperature. The data from at least
10,000 events in each group were acquired with CellQuest software
(Becton-Dickinson). The percentage of the cells in each phase were
presented in the results.
Real-time PCR
The cells were collected after the PSPA treatment.
The total RNA was prepared from the cell cultures, using an RNA
isolation kit (Takara Bio, Inc., Shiga, Japan), according to the
manufacturer's instructions. The concentration of RNA was detected
by NanoDrop (NanoDrop Technologies, Wilmington, DE, USA). The cDNA
Synthesis kit (Takara Bio) was used to synthesize the cDNA from 2
µg of RNA, according to the manufacturer's protocol. A SYBR Green
PCR Master Mix (Takara Bio) was used to quantify the target gene
expression level. The amplification assay was performed on Applied
Biosystems ABI 7500 (Thermo Fisher Scientific, Inc). The primers
used were as follows: Fas sense, 5′-GTGCTTTGCTTAGGGTTCCC-3′ and
antisense, 5′-AACTTGCACTTCTGGCCATG-3′; Fasl sense,
5′-GTCCAACTCAAGGTCCATGC-3′ and antisense,
5′-TTGTTGCAAGATTGACCCCG-3′; Bcl-2-associated X proteins (Bax)
sense, 5′-GTGCCGGAACTGATCAGAAC-3′ and antisense,
5′-CCAAAGTAGGAGAGGAGGCC-3′; Bcl-2 sense, 5′-GCCTTCTTTGAGTTCGGTGG-3′
and antisense, 5′-GAAATCAAACAGAGGCCGCA-3′; cyclin B1 sense, 5′
-TTGTGTGCCCA AGAAGATGC-3′ and antisense,
5′-GAAGTGCAAAGGTAGAGGCC-3′; Cdc2 sense, 5′-GGAAGCTAGGGTAGTCTGGTC-3′
and antisense, 5′-TCTGCAGAGTGGTTTGGTAGA-3′; GAPDH sense,
5′-CACAGTCCATGCCATCACTG-3′ and antisense,
5′-ATCTCGCTCCTGGAAGATGG-3′.
Western blotting
The proteins of the collected cells were isolated
with a lysis buffer containing protease inhibitors (Roche
Diagnostics, Indianapolis, IN, USA). The concentrations of the
extracted proteins were assessed with a BCA protein kit (Bio-Rad
Laboratories). The proteins were denatured, separated via SDS-PAGE,
and then transferred onto the PVDF membrane. The membranes were
blocked with not-fatty milk and then incubated overnight with
primary antibodies at 4°C. The primary antibodies were as follows:
anti-Bcl-2 (1:1,000; cat. no. ab196495; Abcam, Cambridge, MA, USA),
anti-Bax (1:2,000; cat. no. ab182733; Abcam), anti-Fas (1:1,000;
cat. no. ab82419; Abcam), anti-Fasl (1:200; cat. no. ab15285;
Abcam), anti-cyclin B1 (1:3,000; cat. no. ab32053; Abcam),
anti-Cdc2 (1:10,000; cat. no. ab133327; Abcam), anti-PI3K (1:2,000;
cat. no. ab40755; Abcam), anti-phospho-PI3K (1:1,000; cat. no.
ab191606; Abcam), anti-cleaved caspase-3 (1:200; cat. no. 9664;
Cell Signaling Technology, Danvers, MA, USA), anti-phospho-AKT
(1:2,000; cat. no. 4060; Cell Signaling Technology), anti-AKT
(1:1,000; cat. no. 4685; Cell Signaling Technology) and anti-GAPDH
(1:1,000; cat. no. 5174; Cell Signaling Technology). Then, the
secondary antibodies (horseradish peroxidase-conjugated; 1:2,000;
cat. no. ab97051) were added onto the membrane and incubated for 1
h at room temperature. The membrane was washed and moved to
ChemiDoc XRS (Bio-Rad in order to develop the protein band with an
enhanced chemiluminescence reagent (GE Healthcare, Buckinghamshire,
UK).
Statistical analysis
One-way analysis of variance (ANOVA) following
Turkey's multiple comparison test was performed to compare the
differences between the groups. Data were presented as the mean ±
standard deviation (SD). P<0.05 was considered to indicate a
statistically significant difference.
Results
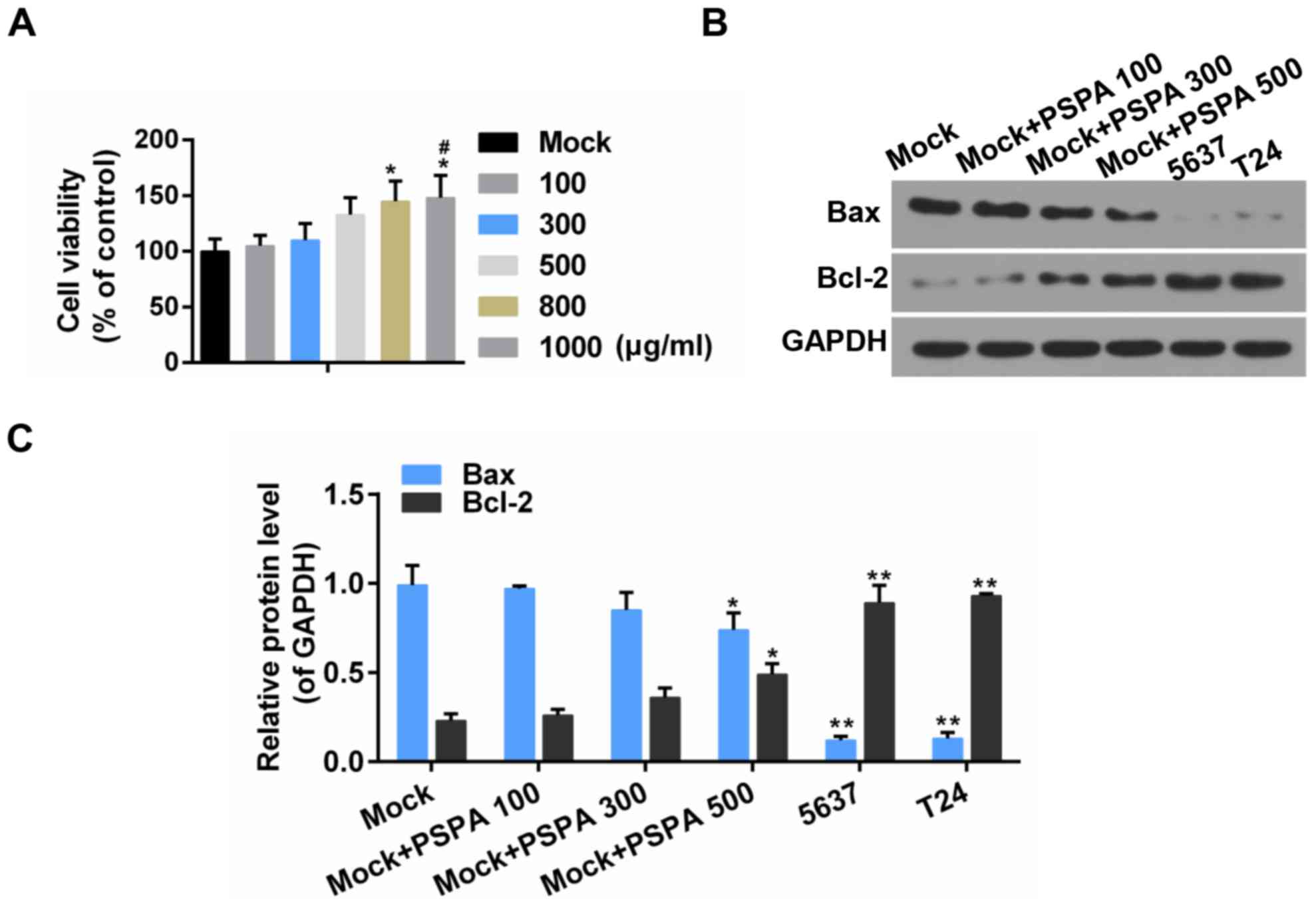
The effect of PSPA on the viability of
BC cells
The acyl-anthocyanin structure in PSP caused the
extracts to have a strong absorption peak approximately 330 nm
(Fig. 1A). The absorbance value in
the visible region (approximately 530 nm) ranged from 0.2–0.4
contingent on the different PSPA types. The PSPA from the Zi A1
extract had the highest concentration of anthocyanins (76.63 mg/100
g Fw) and the highest absorbance value. Therefore, the PSPA from Zi
A1 was selected for the subsequent experiments. The effect of PSPA
on BC cell lines, 5637and T24, was determined. The results revealed
that the viability of the BC cells decreased as the PSPA treatment
concentrations increased. The inhibition rate of the PSPA at 800
µg/ml was >60% after a 72-h incubation (Fig. 1B). PSPAs at concentrations of 100,
300 and 500 µg/ml were chosen to explore the function of PSPA in BC
cells.
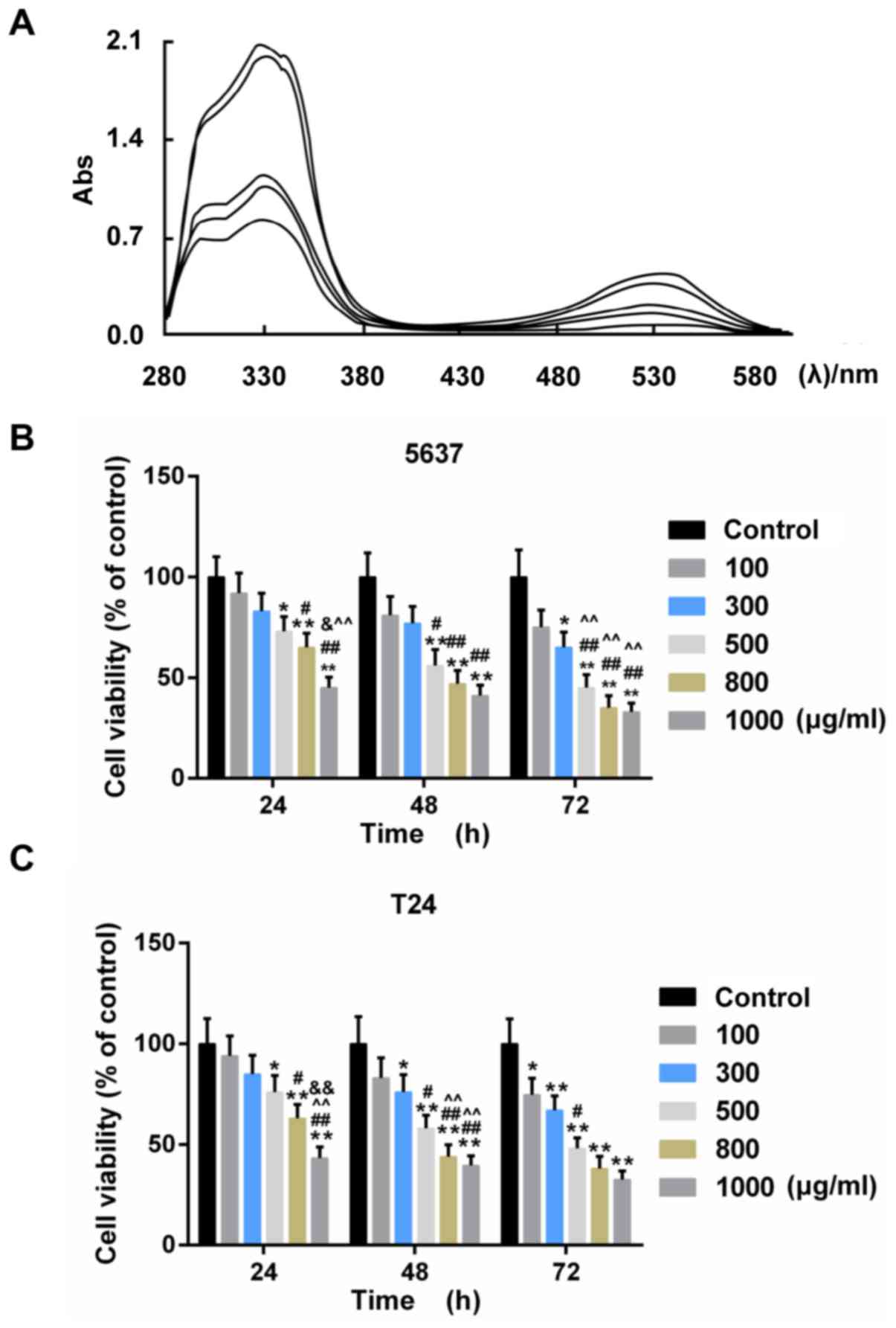 | Figure 1.(A) UV-vis spectra of anthocyanins
from different PSPs at 200–600 nm. The curves in the graph denote
the following PSPA extracts (from the upper to the lowest): Zi A1,
Anhui Zi, Chuanshan Zi, Jishu 18 and Zi A0 at 330 nm. Abs,
absorption rate. (B and C) The cell viability of 5637 (B) and T24
(C) cells under treatment with PSPA at different concentrations:
100, 300, 500, 800 and 1,000 µg/ml; *P<0.05, **P<0.01 vs.
control (BC cells); #P<0.05, ##P<0.01
vs. PSPA 100 µg/ml; ^P<0.05, ^^P<0.01
vs. PSPA 300 µg/ml; &P<0.05,
&&P<0.01 vs. PSPA 500 µg/ml.. |
The effect of PSPA on cell viability
and apoptosis in non-cancerous bladder cells
To estimate the effect of PSPA on non-cancerous
bladder cells, the cell viability and apoptosis was assessed. It
was revealed that, after 48-h incubation with PSPA, the viability
of non-cancerous cells was improved and the non-cancerous cells
exhibited an elevated Bcl-2/Bax ratio. Furthermore, the Bcl-2/Bax
ratio was more obvious in BC cells (Fig. 2). It was indicated that the PSPA
treatment did not decrease the cell proliferation of non-cancerous
cells and it presented proliferation promotion effects to some
extent.
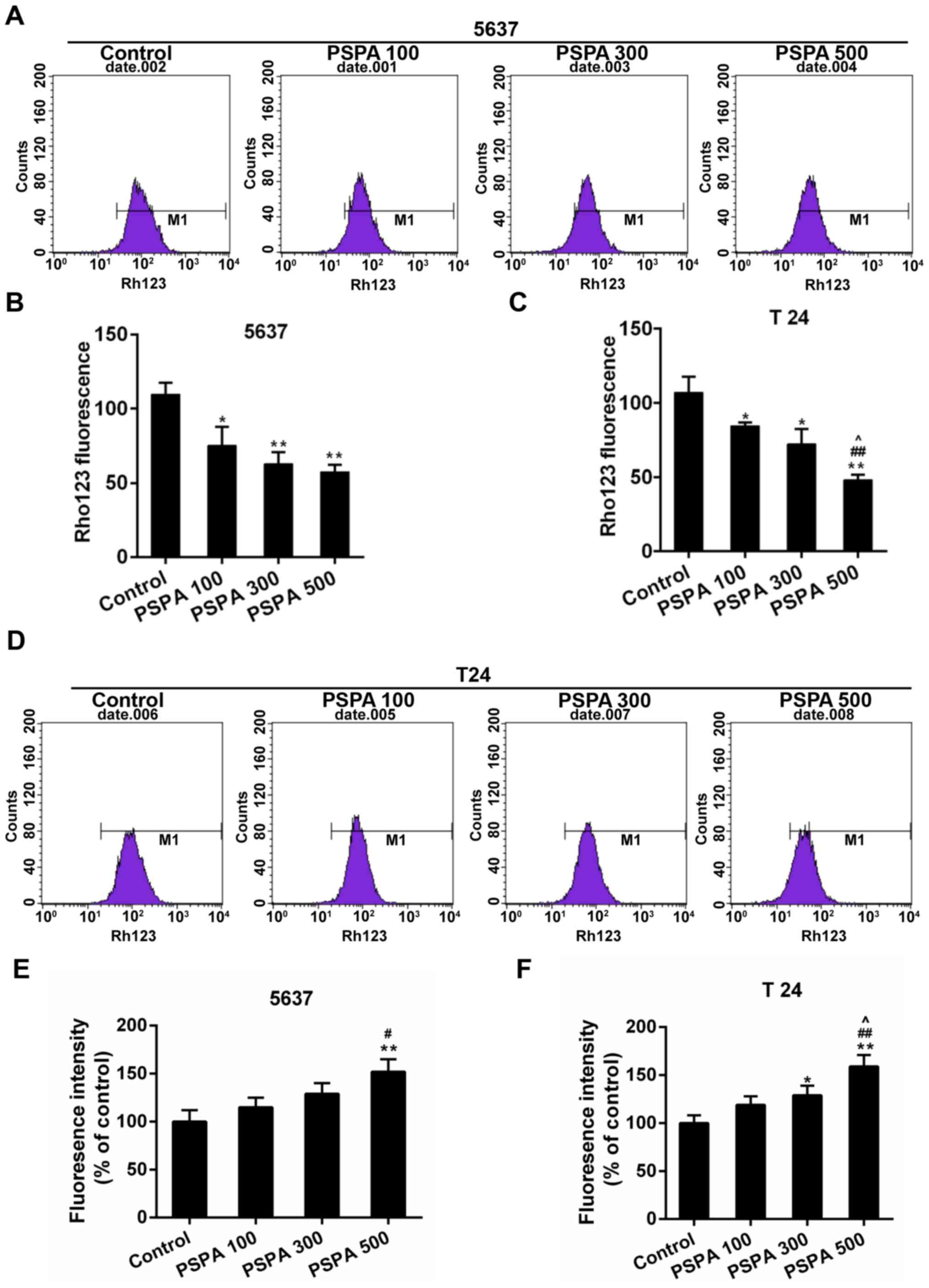
PSPA enhanced the collapse of MMP and
ROS accumulation of BC cells
Mitochondria are deeply involved in cell death. The
collapse of mitochondrial membrane potential (MMP) is the hallmark
step in the progression of cell apoptosis. The loss of the MMP
induces the ROS-mediated oxidative stress (22). The MMP and the ROS content were
assessed to detect whether the PSPA caused oxidative stress in the
BC cells. The results displayed in Fig.
3A-D demonstrated that the Rho123 staining fluorescence was
reduced by the PSPA in a dose-dependent manner. The ROS content in
5637 and T24 cells gradually increased after the PSPA treatment
(Fig. 3E-F).
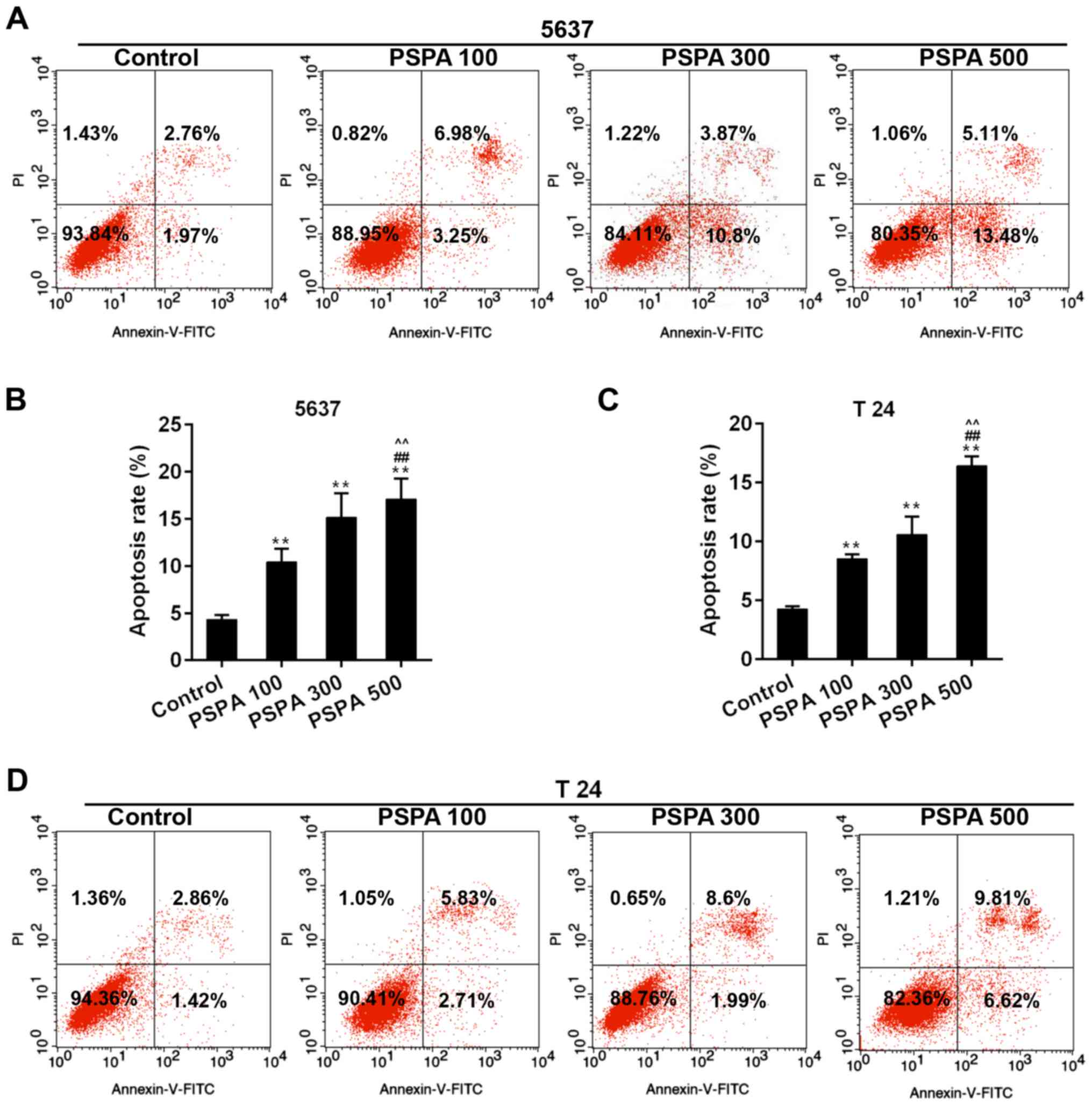
PSPA triggered apoptosis and cell
cycle arrest of BC cells
The apoptosis rate and cell cycle progression of BC
cells were assessed in order to confirm the effect of PSPA on BC
cells. The results from flow cytometry revealed that the marked
apoptosis of 5637 and T24 cells was triggered by the dose-dependent
PSPA treatment (Fig. 4). The cell
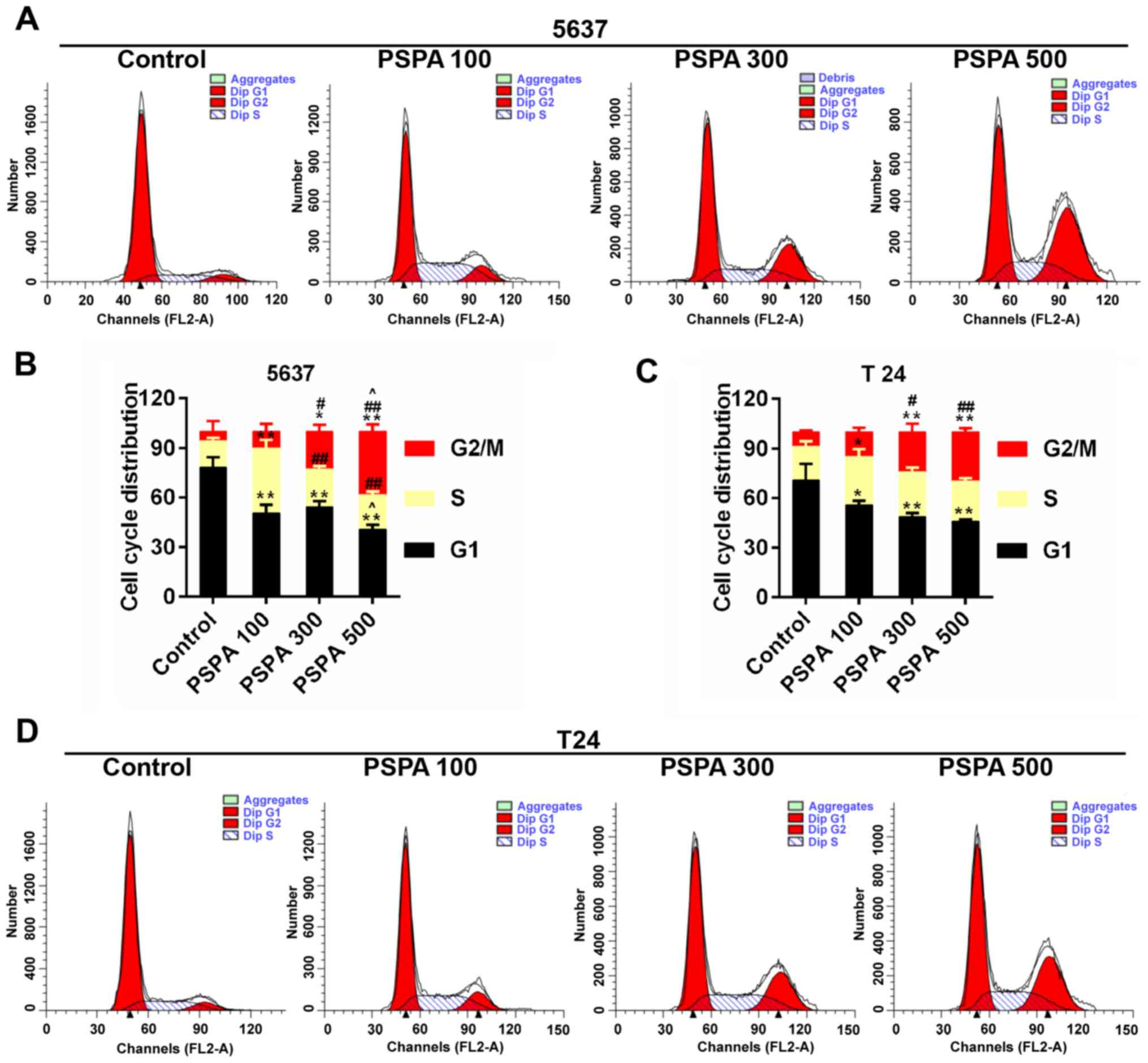
cycle checkpoint has been widely accepted to be linked to cancer.
The cell cycle distribution after the treatment of PSPA was
determined. It can be noticed that a climb in the proportion of
S-phase cells, a decrease in G1-phase cells and an increase in the
G2/M phase. Among these changes, the proportions of cells in the
G2/M phase were increased in an almost linear manner (Fig. 5).
The effect of PSPA on the expression
of apoptotic and cell cycle related proteins
The expression of the apoptosis-related genes was
estimated in order to investigate the mechanisms by which PSPA
enhanced the sensitivity to apoptosis of the BC cells. The results
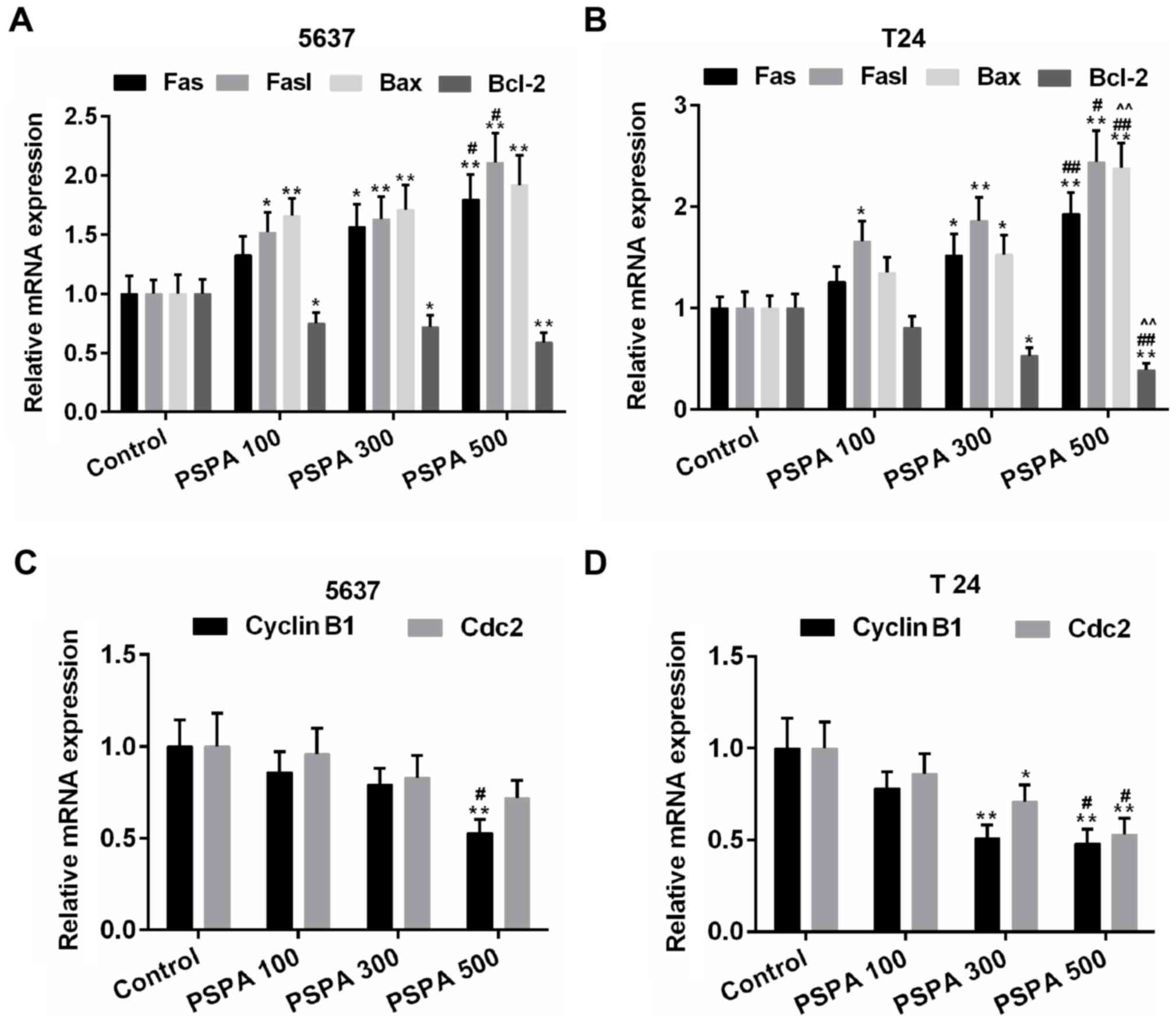
revealed that the expression of the pro-apoptotic genes (Fas, Fasl
and Bax) and the anti-apoptotic gene (Bcl-2) was elevated and then
weakened by the PSPA in the transcriptional level (Fig. 6A and B). Cyclin B1 and Cdc2 are
important cyclin regulatory proteins in the G2/M checkpoint
(23). The mRNA expression of these
two proteins was downregulated in the PSPA groups compared to the
control group both in the transcriptional and translational levels
(Fig. 6C and D). The expression of
these apoptosis- and cell cycle-associated proteins was consistent
within the mRNA level. The expression of cleaved caspase-3 was
higher in the PSPA treatment groups (Fig. 7).
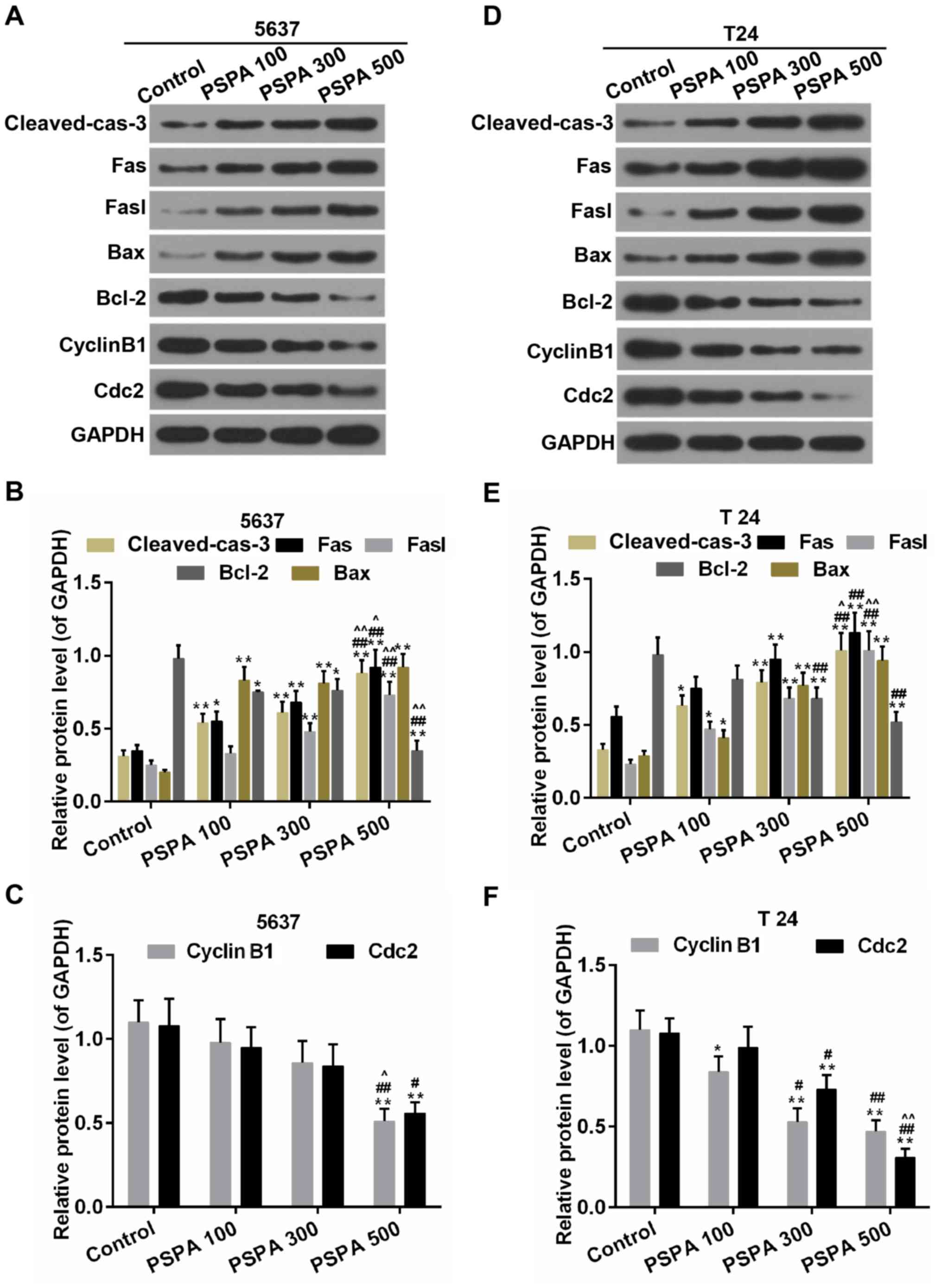 | Figure 7.Western blot analysis and
determination of the expression of cleaved caspase-3, Fas, Fasl,
Bax, Bcl-2, cyclin B1and Cdc2 in 5637 (A-C) and T24 (D-F) cells.
Cleaved-cas-3, cleaved caspase-3; Control, BC cells; PSPA
100/300/500 indicated cells treated with 100/300/500 µg/ml PSPA,
respectively; *P<0.05, **P<0.01 vs. control;
#P<0.05, ##P<0.01 vs. PSPA 100;
^P< 0.05, ^^P< 0.01 vs. PSPA 300. |
PSPA inhibited the PI3K/Akt signaling
pathway in BC cells
PI3K/Akt signaling is an important signaling pathway
in the cell cycle regulation, where it is directly related to
cancer (16). As displayed in
Fig. 8, the PSPA treatment
activated the expression of p-PI3K and p-Akt in the non-cancerous
bladder cells. Furthermore, the activity of PI3K/Akt was elevated
in the BC cells compared to the non-cancerous bladder cells. By
contrast, the phosphorylation of PI3K was suppressed by the PSPA
pre-treatment. The expression of the total PI3K was not influenced.
The phosphorylation of its downstream substrate, Akt, was inhibited
by the PSPA. The expression of the total Akt was not altered.
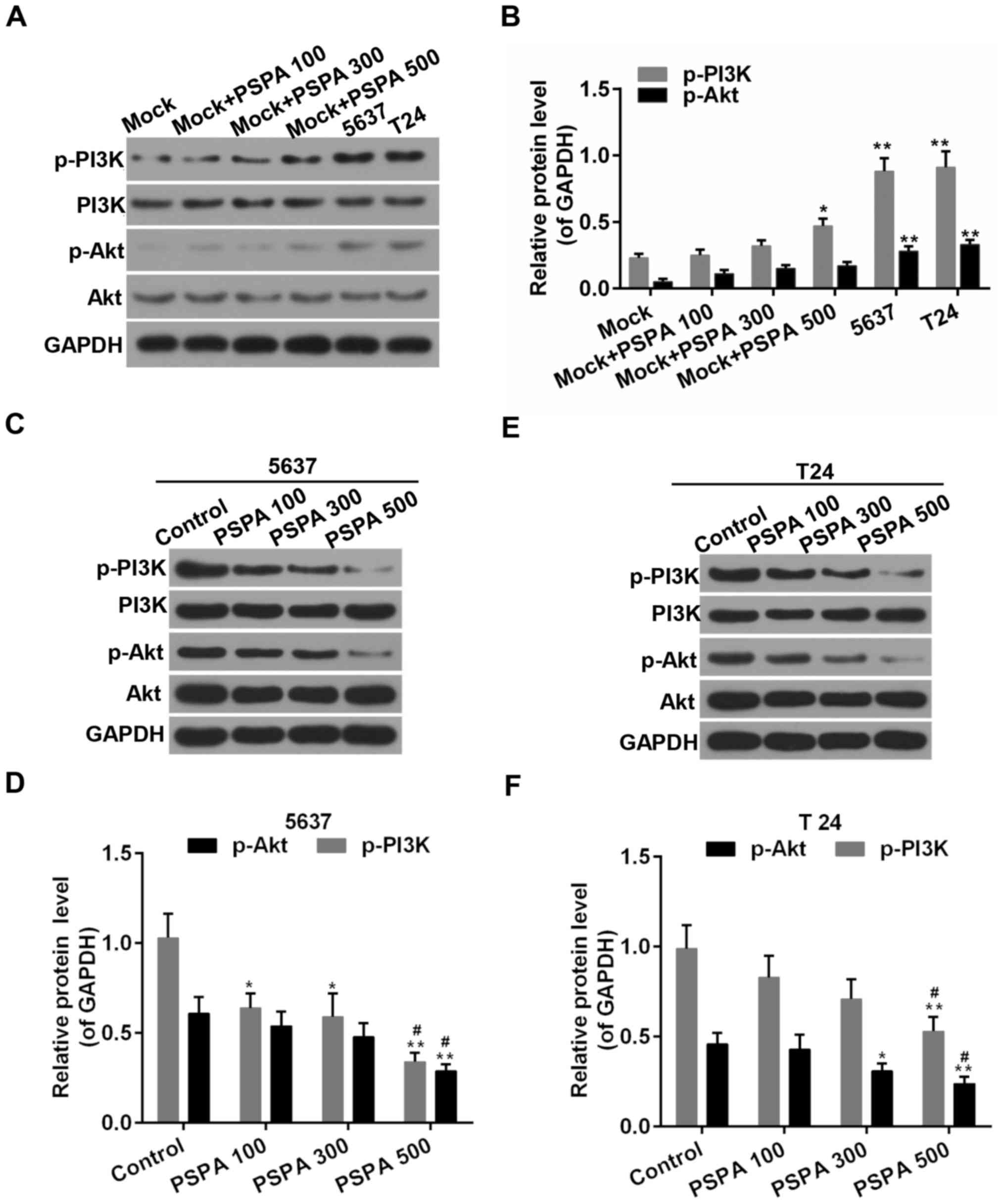 | Figure 8.(A and B) Western blot analysis and
determination of the expression of PI3K, p-PI3K, Akt, and p-Akt in
non-cancerous bladder cells; Mock, non-cancerous bladder cells;
*P<0.05, **P<0.01 vs. mock. (C-F) Western blot analysis and
determination of the expression of PI3K, p-PI3K, Akt, and p-Akt in
5637 (A and B) and T24 (C and D) cells. Control, BC cells; PSPA
100/300/500 indicated cells treated with 100/300/500 µg/ml PSPA,
respectively; *P<0.05, **P<0.01 vs. control;
#P<0.05 vs. PSPA 100. |
Discussion
In the present study, we observed a strong
absorption peak approximately 330 nm that indicated the profound
distribution of acylated anthocyanins in PSP, especially PSPA from
Zi A1. The results in the present study proved that PSPA inhibited
the cell viability of BC cells in a dose-dependent manner. By
contrast, PSPA exhibited an opposite effect in non-cancerous cells
by enhancing cell viability and resistance to apoptosis.
Furthermore, according to previous studies, PSPA was capable of
inhibiting the proliferation in multiple cell types (24), while the proliferation and
anti-apoptotic effect of PSPA has also been reported (25,26).
The reason for this appeared contradictory effect was not very
clear. It may have been influenced by the intracellular
microenvironments.
The present study focused on the antitumor effect of
PSPA. However, the effect of the anthocyanins in these studies was
attributed to the non-acylated anthocyanins. There was a strong
absorption peak approximately 330 nm, which indicated the
distribution of the acylated anthocyanins in the PSPA, particularly
the PSPA from Zi A1. We observed that the PSPA inhibited the BC
cell viability in a dose-dependent manner. This effect was largely
attributed to the acylated anthocyanins in PSPA, which was
supported by a previous study (27). Caffeic and ferulic acid also
demonstrated anti-proliferative abilities in cancer cells (28,29),
therefore, coffeoyl and feruloyl in PSPA provided the beneficial
antitumor effect. Anthocyanins are first metabolized by the gut
microflora, leading to anthocyanidin metabolites (30,31).
It was uncertain whether the anthocyanins were metabolized into
anthocyanidins or other metabolites within this study model. The
exact molecules (anthocyanins, anthocyanidins or their combined
effect) that exerted this cancer prevention effect were unclear in
the present study.
Furthermore, the collapse of the MMP and the ROS
content were enhanced by the PSPA treatment. Oxidative stress was
subsequently triggered, which was closely related to cell survival
(32). The growth inhibitory effect
of PSPA could be attributed to the overload of ROS in BC cells.
This was in agreement with some cancer cell studies (13,14).
There were studies that reported a promotion effect of PSPA on cell
viability (25,33). The anthocyanins appeared to have a
paradoxical role in the generation of the ROS (34–36).
These contradictory results could be caused by the different cell
redox status, the cell types and the concentration employed, which
would lead to different cell fates.
The anti-proliferation ability could be modulated by
the induction of apoptosis. Mitochondria have been implicated in
apoptosis (37). Mitochondria
dysfunction triggers the dissipation of MMP, the release of
caspase-activating factors and finally leads to the activation of
apoptosis cascades (38). The
apoptosis of BC cells was aggravated by PSPA. On the contrary, cell
cycle arrest is another common cancer prevention strategy. The
effect of PSPA on cell cycle arrest was further evaluated. The data
in the present study indicated that PSPA induced the G2/M arrest of
the BC cells. The G2/M checkpoint is a DNA-damage checkpoint in
cell cycle regulation, which suppresses the progression of the cell
cycle (23). This checkpoint was
used to prevent tumor growth and to induce the apoptosis of tumor
cells. Thus, PSPA was able to suppress the growth of BC cells by
inducing cell cycle arrest.
Many molecules have been identified as participants
in the apoptosis cascades, which either promote or inhibit
apoptosis. For instance, Bcl-2 is an anti-apoptotic molecule, while
the Fas/Fasl and Bcl-2-associated X protein (Bax) are pro-apoptotic
molecules (39,40). The activation of caspase-3 is the
convergence of multiple apoptosis signaling (41). The expression of the pro-apoptotic
genes (Fas, Fasl, and Bax) and the anti-apoptotic gene (Bcl-2) was
induced and suppressed by the PSPA. The PSPA enhanced the activity
of caspase-3 and increased the BC cell apoptosis. This effect was
partly due to the inhibitory effect of PSPA on cell viability. In
addition, the expression of regulatory proteins, cyclin B1 and Cdc2
that were involved in the G2/M checkpoint was evaluated to confirm
the effect of PSPA. The degradation of cyclin B1 and Cdc2 would
promote the G2/M arrest, which could be a treatment option for
tumor suppression (42). The
results revealed that the expression of cyclin B1 and Cdc2 was
downregulated by PSPA. These results indicated that PSPA inhibited
tumor growth, enhanced the sensitivity to apoptosis and induced the
G2/M arrest of the BC cells.
PI3K/Akt is an important mechanism for controlling
cell growth and proliferation (16). The PSPA was suggested to activate
the PI3K/Akt signaling in some study models (43,44).
The data in the present study revealed that the activity of PI3K
and Akt was mitigated in the PSPA treatment group. This result
seemed to be conflicting to the effect of the PSPA. However,
hyperactivation of the PI3K/Akt signaling is common in cancer
(45,46). Therefore, the decreased activity of
the PI3K/Akt signaling could be favorable for cancer prevention.
This could depend on the cell types and cell context (47). These observations support the
beneficial effect of PSPA. Collectively, the antitumor effect of
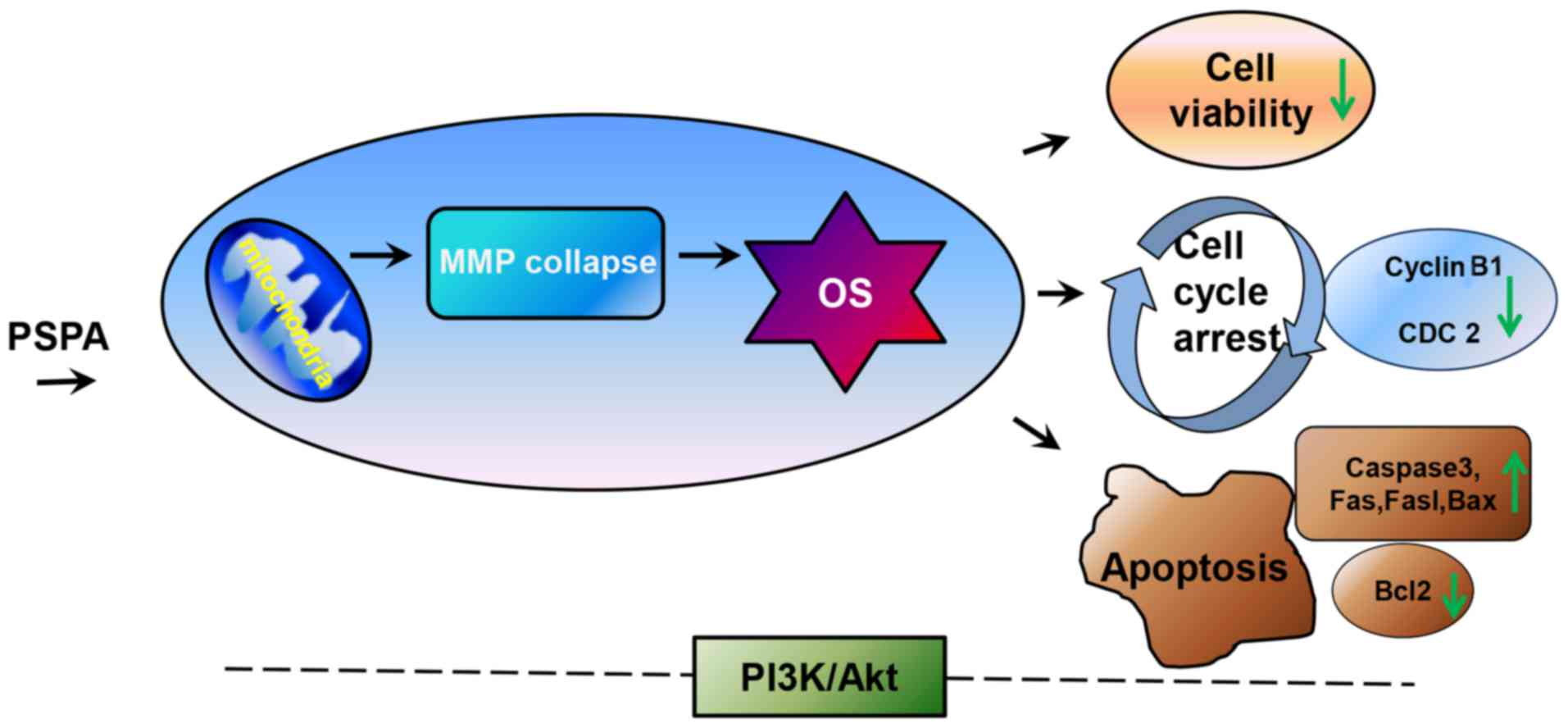
PSPA was proved in BC cells. According to the work model scheme
displayed in Fig. 9, after the
treatment of PSPA, the mitochondria in BC cells were dysregulated
and then serious oxidative stress was triggered. Under this
excessive oxidative stress, the cell viability was reduced, the
cell cycle was arrested and apoptosis was induced in BC cells.
Although the exact regulation mechanism needed further
investigation, our results revealed that the inhibition of PI3K/Akt
was associated with the protective effect of PSPA. Furthermore, the
antitumor effect of PSPA was gathered from the in vitro
data. This was another limitation of the present study. It is
necessary to confirm the antitumor effect within further in
vivo studies. The combination administration of the current
chemotherapeutic drug with the PSPA could bring an intriguing
effect in protection against BC.
The current study demonstrated that PSPA exerted an
antitumor effect in BC cells through the suppression of cell
viability, the aggravation of MMP collapse, the promotion of
apoptosis and the induction of cell cycle arrest, which could be
related to the suppression of the PI3K/Akt signaling. The data in
the present study were an in vitro analysis and this study
seems to be the first preclinical study that illustrated the effect
of PSPA in BC. The present study shed light on a possible antitumor
agent as a treatment option for BC.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Zhejiang
Provincial Health Department (grant no. 2014ZB135).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
WLL and TH conceived and designed the study. HYY,
XJZ and MK performed the experiments. WLL wrote the study. WLL,
HYY, XJZ and TH reviewed and edited the manuscript. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jemal A, Siegel R, Ward E, Hao Y, Xu J,
Murray T and Thun MJ: Cancer statistics, 2008. CA Cancer J Clin.
58:71–96. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CA Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wu X, Ros MM, Gu J and Kiemeney L:
Epidemiology and genetic susceptibility to bladder cancer. BJU Int.
102:1207–1215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeegers MP, Tan FE, Dorant E and van Den
Brandt PA: The impact of characteristics of cigarette smoking on
urinary tract cancer risk: A meta-analysis of epidemiologic
studies. Cancer. 89:630–639. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rödel C, Grabenbauer GG, Kühn R,
Papadopoulos T, Dunst J, Meyer M, Schrott KM and Sauer R:
Combined-modality treatment and selective organ preservation in
invasive bladder cancer: Long-term results. J Clin Oncol.
20:3061–3071. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kaufman DS, Shipley WU and Feldman AS:
Bladder cancer. Lancet. 374:239–249. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hanneken A, Lin FF, Johnson J and Maher P:
Flavonoids protect human retinal pigment epithelial cells from
oxidative-stress-induced death. Invest Ophthalmol Vis Sci.
47:3164–3177. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang ZF, Fan SH, Zheng YL, Lu J, Wu DM,
Shan Q and Hu B: Purple sweet potato color attenuates oxidative
stress and inflammatory response induced by d-galactose in mouse
liver. Food Chem Toxicol. 47:496–501. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shindo M, Kasai T, Abe A and Kondo Y:
Effects of dietary administration of plant-derived anthocyanin-rich
colors to spontaneously hypertensive rats. J Nutr Sci Vitaminol
(Tokyo). 53:90–93. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhao JG, Yan QQ, Lu LZ and Zhang YQ: In
vivo antioxidant, hypoglycemic, and anti-tumor activities of
anthocyanin extracts from purple sweet potato. Nutr Res Pract.
7:359–365. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Suda I, Oki T, Masuda M, Nishiba Y, Furuta
S, Matsugano K, Sugita K and Terahara N: Direct absorption of
acylated anthocyanin in purple-fleshed sweet potato into rats. J
Agric Food Chem. 50:1672–1676. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Harada K, Kano M, Takayanagi T, Yamakawa O
and Ishikawa F: Absorption of acylated anthocyanins in rats and
humans after ingesting an extract of Ipomoea batatas purple
sweet potato tuber. Biosci Biotechnol Biochem. 68:1500–1507. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hagiwara A, Yoshino H, Ichihara T, Kawabe
M, Tamano S, Aoki H, Koda T, Nakamura M, Imaida K, Ito N, et al:
Prevention by natural food anthocyanins, purple sweet potato color
and red cabbage color, of
2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP)-associated
colorectal carcinogenesis in rats initiated with
1,2-dimethylhydrazine. J Toxicol Sci. 27:57–68. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lim S, Xu J, Kim J, Chen TY, Su X,
Standard J, Carey E, Griffin J, Herndon B, Katz B, et al: Role of
anthocyanin-enriched purple-fleshed sweet potato p40 in colorectal
cancer prevention. Mol Nutr Food Res. 57:1908–1917. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hayashi K, Hibasami H, Murakami T,
Terahara N, Mori M and Tsukui A: Induction of apoptosis in cultured
human stomach cancer cells by potato anthocyanins and its
inhibitory effects on growth of stomach cancer in mice. Food Sci
Technol Res. 12:22–26. 2006. View Article : Google Scholar
|
|
16
|
Cantley LC: The phosphoinositide 3-kinase
pathway. Science. 296:1655–1657. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Luo J, Manning BD and Cantley LC:
Targeting the PI3K-Akt pathway in human cancer: Rationale and
promise. Cancer Cell. 4:257–262. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shaw RJ and Cantley LC: Ras, PI(3)K and
mTOR signalling controls tumour cell growth. Nature. 441:424–430.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Philp AJ, Campbell IG, Leet C, Vincan E,
Rockman SP, Whitehead RH, Thomas RJ and Phillips WA: The
phosphatidylinositol 3′-kinase p85alpha gene is an oncogene in
human ovarian and colon tumors. Cancer Res. 61:7426–7429.
2001.PubMed/NCBI
|
|
20
|
Parsons DW, Wang TL, Samuels Y, Bardelli
A, Cummins JM, DeLong L, Silliman N, Ptak J, Szabo S, Willson JK,
et al: Colorectal cancer: Mutations in a signalling pathway.
Nature. 436:7922005. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu X, Mu T, Sun H, Zhang M and Chen J:
Optimisation of aqueous two-phase extraction of anthocyanins from
purple sweet potatoes by response surface methodology. Food Chem.
141:3034–3041. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang LJ, Zhu LX, Dong WR, Cao YL, Wang K,
Huang HQ and Rong ZJ: Department of Orthopaedics, Zhujiang
Hospital, Southern Medical University: Changes of ROS and
mitochondrial transmembrane potential in oxidative stress-induced
apoptosis in nucleus pulposus cells of rats. J Pract Med.
2:2014.(In Chinese). http://en.cnki.com.cn/Article_en/CJFDTOTAL-SYYZ201402011.htm
|
|
23
|
Yao YB, Peng ZG, Liu ZF, Yang J and Luo J:
Effects of mangiferin on cell cycle status and CDC2/Cyclin B1
expression of HL-60 cells. Zhong Yao Cai. 33:81–85. 2010.(In
Chinese). PubMed/NCBI
|
|
24
|
Matsunaga N, Tsuruma K, Shimazawa M,
Yokota S and Hara H: Inhibitory actions of bilberry anthocyanidins
on angiogenesis. Phytother Res. 24 Suppl 1:S42–S47. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sun M, Lu X, Hao L, Wu T, Zhao H and Wang
C: The influences of purple sweet potato anthocyanin on the growth
characteristics of human retinal pigment epithelial cells. Food
Nutr Res. 59:278302015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shih PH, Yeh CT and Yen GC: Anthocyanins
induce the activation of phase II enzymes through the antioxidant
response element pathway against oxidative stress-induced
apoptosis. J Agric Food Chem. 55:9427–9435. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu X, Sun M, Hao L, Wu T, Zhao H and Wang
C: Purple sweet potato anthocyanin inhibits the proliferation of
human retinal pigment epithelial cell by blocking cell cycle and
inducing apoptosis. Adv J Food Sci Technol. 11:561–569. 2016.
View Article : Google Scholar
|
|
28
|
Serafim TL, Carvalho FS, Marques MP,
Calheiros R, Silva T, Garrido J, Milhazes N, Borges F, Roleira F,
Silva ET, et al: Lipophilic caffeic and ferulic acid derivatives
presenting cytotoxicity against human breast cancer cells. Chem Res
Toxicol. 24:763–774. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Eitsuka T, Tatewaki N, Nishida H, Kurata
T, Nakagawa K and Miyazawa T: Synergistic inhibition of cancer cell
proliferation with a combination of δ-tocotrienol and ferulic acid.
Biochem Biophys Res Commun. 453:606–611. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Felgines C, Texier O, Besson C, Fraisse D,
Lamaison JL and Rémésy C: Blackberry anthocyanins are slightly
bioavailable in rats. J Nutr. 132:1249–1253. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tsuda T, Horio F and Osawa T: Absorption
and metabolism of cyanidin 3-O-beta-D-glucoside in rats. FEBS Lett.
449:179–182. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Brieger K, Schiavone S, Miller FJ Jr and
Krause KH: Reactive oxygen species: From health to disease. Swiss
Med Wkly. 142:w136592012.PubMed/NCBI
|
|
33
|
Ye J, Meng X, Yan C and Wang C: Effect of
purple sweet potato anthocyanins on beta-amyloid-mediated PC-12
cells death by inhibition of oxidative stress. Neurochem Res.
35:357–365. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Cvorovic J, Tramer F, Granzotto M,
Candussio L, Decorti G and Passamonti S: Oxidative stress-based
cytotoxicity of delphinidin and cyanidin in colon cancer cells.
Arch Biochem Biophys. 501:151–157. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Feng R, Ni HM, Wang SY, Tourkova IL,
Shurin MR, Harada H and Yin XM: Cyanidin-3-rutinoside, a natural
polyphenol antioxidant, selectively kills leukemic cells by
induction of oxidative stress. J Biol Chem. 282:13468–13476. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schumacker PT: Reactive oxygen species in
cancer cells: Live by the sword, die by the sword. Cancer Cell.
10:175–176. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kaul M, Garden GA and Lipton SA: Pathways
to neuronal injury and apoptosis in HIV-associated dementia.
Nature. 410:988–994. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
38
|
Jeong SY and Seol DW: The role of
mitochondria in apoptosis. BMB Rep. 41:11–22. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Villa-Morales M and Fernández-Piqueras J:
Targeting the Fas/FasL signaling pathway in cancer therapy. Expert
Opin Ther Targets. 16:85–101. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gil-Gómez G, Berns A and Brady HJ: A link
between cell cycle and cell death: Bax and Bcl-2 modulate Cdk2
activation during thymocyte apoptosis. EMBO J. 17:7209–7218. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Harada J and Sugimoto M: Activation of
caspase-3 in beta-amyloid-induced apoptosis of cultured rat
cortical neurons. Brain Res. 842:311–323. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang HC, Pao J, Lin SY and Sheen LY:
Molecular mechanisms of garlic-derived allyl sulfides in the
inhibition of skin cancer progression. Ann NY Acad Sci. 1271:44–52.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lu J, Wu DM, Zheng YL, Hu B and Zhang ZF:
Purple sweet potato color alleviates D-galactose-induced brain
aging in old mice by promoting survival of neurons via PI3K pathway
and inhibiting cytochrome C-mediated apoptosis. Brain Pathol.
20:598–612. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhang ZF, Lu J, Zheng YL, Hu B, Fan SH, Wu
DM, Zheng ZH, Shan Q and Liu CM: Purple sweet potato color protects
mouse liver against d-galactose-induced apoptosis via inhibiting
caspase-3 activation and enhancing PI3K/Akt pathway. Food Chem
Toxicol. 48:2500–2507. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shukla S, Maclennan GT, Hartman DJ, Fu P,
Resnick MI and Gupta S: Activation of PI3K-Akt signaling pathway
promotes prostate cancer cell invasion. Int J Cancer.
121:1424–1432. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tokunaga E, Kimura Y, Mashino K, Oki E,
Kataoka A, Ohno S, Morita M, Kakeji Y, Baba H and Maehara Y:
Activation of PI3K/Akt signaling and hormone resistance in breast
cancer. Breast Cancer. 13:137–144. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Powis G, Ihle N and Kirkpatrick DL:
Practicalities of drugging the phosphatidylinositol-3-kinase/Akt
cell survival signaling pathway. Clin Cancer Res. 12:2964–2966.
2006. View Article : Google Scholar : PubMed/NCBI
|