Introduction
The aryl hydrocarbon receptor (AHR) is a nuclear
transcription factor known as a dioxin receptor and mediates the
toxic effects of industrial contaminants such as polycyclic
aromatic hydrocarbons, the forefather being
2,3,7,8-tetrachlorodibenzo-p-dioxin or TCDD. The effect of
TCDD upon AHR activation on cancer stages are well known (1,2). Prior
to ligand binding AHR is found in the cytosol in the form of a
multiprotein complex with a sedimentation coefficient of 9 Svedberg
(AHRC-9S). The complex consists of five different polypeptides and
six proteins including AHR, heat shock protein 90-homodimer (two
HSP-90 proteins), P23 (a prostaglandin E synthase with
co-chaperonin activity vs. HSP-90), SRC kinase (a tyrosin kinase
homologous to viral oncogenes of the Rous sarcoma) and AHR
interacting protein (AIP) (3). Upon
activation by a wide range of environmental toxins, AHR
translocates to the nucleus and heterodimerizes with the AHR
nuclear translocator protein (ARNT). Finally, the AHR-ARNT complex
binds DNA to the xenobiotics responsive element (XRE) of the
promoters, resulting in the transcriptional activation of genes,
such as cytochrome P450 (CYP450). The human CYP superfamily (CYP1,
CYP2 and CYP3) contains 57 functional genes, and play an important
role in the metabolism of therapeutic drugs, other xenobiotics, and
various endogenous compounds. Several of these CYPs (e.g., CYP1A1
and 1A2) are involved in the metabolism and bioactivation of
carcinogens. Cytochrome P450 1A1, a well-known aryl hydrocarbon
hydroxylase, is one of the most important enzymes responsible for
the metabolism of environmental pollutants and many aromatic
compounds involved in tumorigenesis (4).
Since AhR activation and subsequent induction of
AhR-regulated P450 enzymes (CYP1A1, CYP1B1) have been implicated in
oral squamous cell carcinoma (OSCC) (5), we investigated the role of AHR and
HSP-90 in OSCC tumorigenesis and progression. The expression of AHR
by immunohistochemical staining of OSCC formalin-fixed,
paraffin-embedded (FFPE) tissues was determined and its correlation
with different tumor grades was assessed. As control of the
transcriptional activation of AHR downstream stimulated genes we
also investigated CYP1A1 (6).
Polydatin (trans-piceid
3,5,4′-trihydroxystilbene-3-O-β-D-glucopyranoside) is a widespread
natural substance contained in red wine and a direct precursor of
resveratrol, a well-known AHR inhibitor (7,8). The
glucopyranoside ring of polydatin influences the pharmocodynamics
(9) and pharmacokinetics (10,11)
resulting in better biodisponibility in respect to resveratrol.
Polydatin was also demonstrated to act as an
antitumoral substance by stimulation of apoptosis in leukemia, lung
cancer, colorectal cancer cell lines, and ovarian cancer (12–17).
In the present study, we confirmed the effect of
polydatin in the CAL27 cell line, a model of oral carcinoma to
establish the effects on AHR expression and monitoring the AHR
transcriptional factor activity using CYP1A1 as an AHR reporter
gene. Moreover, we monitored the effect of polydatin on HSP-90, a
component of the AHR complex, essential for Ah receptor signaling
(18,19).
In a previous study by computational analysis, the
binding site for ligands on the human AHR surface was proposed
(20). With a similar computational
approach, we present our results relative to docking studies on the
direct interaction between polydatin and human AHR.
Materials and methods
Patients and tissue samples
Twenty-five OSCC patients were enrolled in the
present study. Tumor tissues were collected at the time of
resection, and informed written consent for the use of sample
tissues was obtained from all patients. We did not seek ethics
committee approval, as all data analyzed were collected as part of
routine diagnosis and treatment, and all patients were diagnosed
and treated according to institutional guidelines and
agreements.
Moreover, the present study does not report on the
use of experimental therapies. The baseline characteristics of the
OSCC patients investigated in the study are reported in Table I. Twenty-five samples of hOSCC from
various sites of oral regions were collected. Each tissue sample
was fixed in 10% neutral-buffered formalin, embedded in paraffin,
routinely processed and stained in hematoxylin and eosin. Each
sample was independently graded by two observers: grade 1 (11/25),
grade 2 (8/25) and grade 3 (6/25) OSCC, according to the
Histological Grading System proposed by Bryne et al
(21).
 | Table I.Baseline characteristics of the OSCC
patients investigated in the study. |
Table I.
Baseline characteristics of the OSCC
patients investigated in the study.
|
| Patients |
|---|
|
|
|
|---|
| Characteristics | No. | % |
|---|
| Age, years |
| ≤60 | 6 | 24 |
|
>60 | 19 | 76 |
| Sex |
|
Male | 15 | 60 |
|
Female | 10 | 40 |
| N-regional lymph
node status |
|
Negative | 9 | 36 |
|
Positive | 16 | 64 |
| Histological
grade |
| Grade
1 | 11 | 44 |
| Grade
2 | 8 | 32 |
| Grade
3 | 6 | 24 |
| Smoking
history |
|
Yes | 18 | 72 |
| No | 7 | 28 |
Immunoperoxidase staining
Sections were de-paraffinized in xylene, dehydrated
in graded alcohols and washed in 0.01 M phosphate-buffered saline
(PBS). Endogenous peroxidase was blocked with hydrogen peroxide
0.3% in absolute methanol for 30 min. The
streptavidin-biotin-peroxidase method (LSAB kit; Dako, Glostrup,
Denmark) was used. Antigen enhancement was performed by
pretreatment with microwave heating in citrate buffer, pH 6.0,
twice for 5 min at 750 W. As primary antibodies, polyclonal rabbit
anti-human AHR receptor (clone H-211) (Santa Cruz Biotechnology,
Inc., Dallas, TX, USA), polyclonal rabbit anti-human cytochrome
P450 (clone A1A) and HSP-90 (AB 13492) (both from Abcam, Cambridge,
UK) were used; they were diluted 1:100 and 1:200 in PBS and applied
overnight at 4°C. The immunolabeling procedure included negative
control sections incubated with normal serum, instead of the
primary antibody. The MACH1 Universal HPR-Polymer Detection kit
(Biocare Medical LLC, Concord, CA, USA) was used as the secondary
antibody. In order to visualize immunolabeling,
3′3′-diaminobenzidine tetrahydrochloride was used as a chromogen,
with hematoxylin as a counterstain.
Scoring of immunoreactivity and
statistical analysis
The intensity of immunolabeling in each specimen,
for each antibody, was scored by two independent observers, under
blinded conditions, as performed in a previous study (22). For each tumor, 20 fields were
examined at a magnification of ×200 (20× objective, 10× ocular),
and the immunosignal was scored from absent to strong, as follows:
n.a., not assessed; -, negative staining; +/−, weak immunolabeling;
+, moderate immunolabeling; ++, strong immunolabeling.
Moreover, the mean percentage of positive cells in
10 high-power microscopic fields was determined using a scale of
1–4 as follows: 1 indicated the staining of 0–25% of cells, and 2,
3 and 4 indicated 25–50, 50–75 and 75–100% staining,
respectively.
Cell culture
Human tongue squamous cell carcinoma cell line CAL27
was obtained from the American Tissue Culture Collection (ATCC;
Rockville, MD, USA). The cells were grown in DMEM with 10% fetal
bovine serum (FBS) (both from Gibco; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), 100 U/ml penicillin and 100 µg/ml
streptomycin in a humidified 37°C incubator with 5% CO2.
After incubation for 4 h in Dulbecco's modified Eagle's medium
(DMEM) with 10% FBS, the cells were washed with 1%
phosphate-buffered saline (PBS) to remove unattached dead cells,
and were incubated for 24 h with different concentrations (range
0–150 µM) of polydatin
(trans-5,4′-dihydroxystilbene-3-O-β-D-glucopy-ranoside).
Cell viability
We assessed the sensitivity of the cell lines tested
to polydatin using a microplate colorimetric assay that measures
the ability of viable cells to transform a soluble tetrazolium salt
(MTT) to an insoluble purple formazan precipitate. Cells were
plated at the appropriate density (5×103 Cal-27 cells)
in 96-well microtitre plates. After 4 h, cells were exposed to
definite concentrations of polydatin for 24 h. Then, 50 µl of MTT
(1 mg/ml) and 200 µl of medium were added to the cells in each
well. After a 4 h incubation at 37°C, the medium was removed, and
the formazan crystals were solubilized by adding 150 µl of DMSO and
by mixing it in an orbital shaker for 5 min. The absorbance at 570
nm was measured by Bio-Rad 550 microplate reader (Bio-Rad
Laboratories, Milan, Italy). Experiments were performed in
triplicate. As a control, 0.5% DMSO was added to untreated
cells.
Western blot analysis
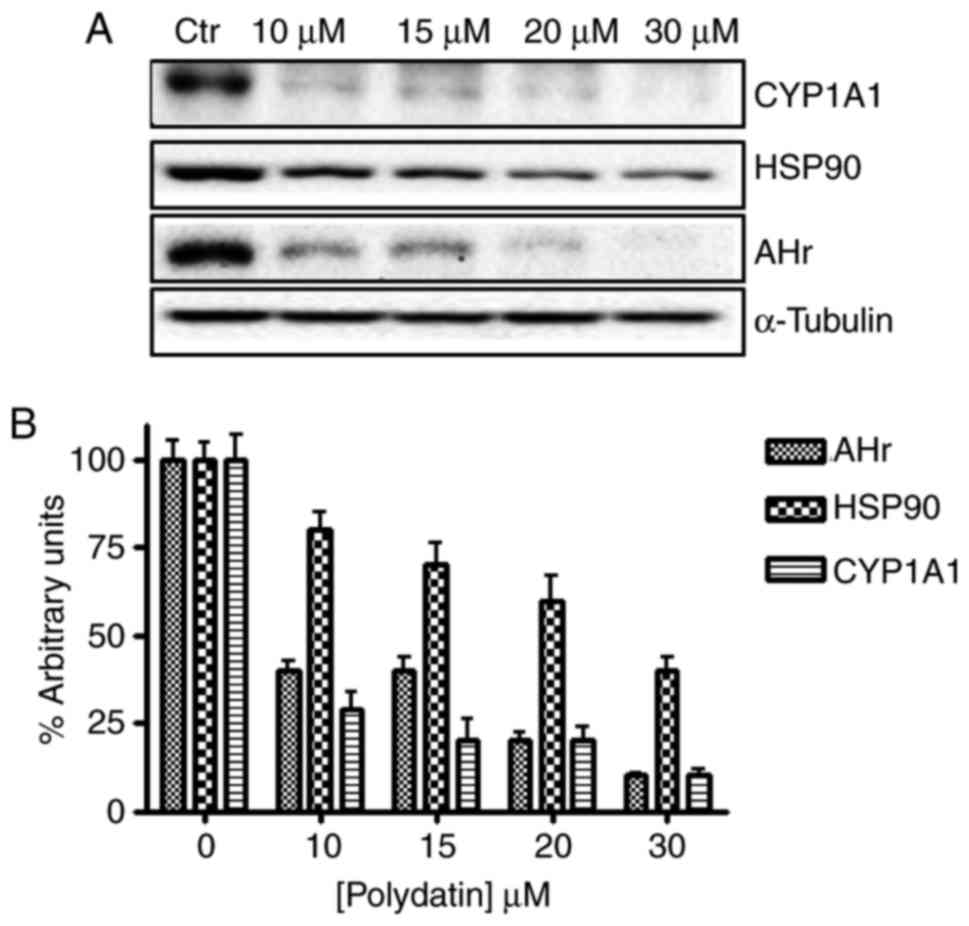
Cal-27 cells were grown for 24 h with or without
polydatin at the concentrations reported in Fig. 5 legend. The cells were washed twice
with ice-cold PBS/BSA, scraped and centrifuged for 30 min at 4°C in
1 ml of lysis buffer (1% Triton, 0.5% sodium deoxycholate, 0.1 M
NaCl, 1 mM EDTA, pH 7.5, 10 mM Na2HPO4, pH
7.4, 10 mM PMSF, 25 mM benzamidin, 1 mM leupeptin, 0.025 U/ml
aprotinin). Equal amounts of cell proteins were separated by
SDS-PAGE. The proteins on the gels were electrotransferred to
nitrocellulose and reacted with polyclonal rabbit anti-human AHR
receptor (clone H-211) (Santa Cruz Biotechnology), polyclonal
rabbit anti-human cytochrome P450 (clone A1A) and HSP-90 (AB 13492)
(both from Abcam) as recommended by the manufacturer. This step was
followed by incubation with corresponding horseradish peroxidase
(HRP)-conjugated secondary antibody. Protein bands were detected by
chemiluminescence detection reagents (Millipore Corporation,
Billerica, MA, USA). The protein bands were quantified using the
NIH ImageJ system (National Institute of Health, Bethesda, MD,
USA). All the western blot analyses were repeated three times.
Western blotting for tubulin was used as an internal control. To
quantify the results, the relative amount of each protein was
determined.
Molecular simulations
Molecular docking approach was used to simulate
interaction of polydatin and the human AHR ligand binding domain.
The structure of polydatin was obtained from PubChem database
(https://pubchem.ncbi.nlm.nih.gov/).
The structure of human AHR ligand binding domain used in the
simulations was that described in a previous study (20). In brief, the amino acid sequence of
human Ah receptor obtained from Uniprot (Accession no. P35869) was
restricted to the region between residue 290 and 390. Secondary
structures were calculated with the DSSP program and
three-dimensional models of human AHR-polydatin were elaborated
using MODELLER 9v7 version and evaluated for stereo-chemical with
PRO-CHECK program. The energy profiles were elaborated applying
PROSAII program. Docking procedure was performed using AutoDock 4.2
and ADTools for preparation of molecules and parameters files,
according to the procedures used in a previous study (20). At least 100 docking determination
using Lamarckian genetic algorithm with the default parameters were
performed, with a maximum of 2.5×106 energy evaluations. Only the
conformational clusters most populated and with lower binding
energy (Eb) were utilized in docking analysis and clustered using
an root-mean-square distance (RMSD) value of 2.0 Å. Molecular
images were generated by BIOVIA Discovery Studio (Dassault
Systèmes).
Results
Immunohistochemistry
We examined the intensity of immunolabeling, the
percentage of positive cells and the localization of AHR, CYP1A1
and HSP-90 in 25 OSCCs, and we report the results in Table II.
| Table II.IHC of Ah receptor, cytochrome P450
and HSP-90 in 25 OSCC tissue samples. |
Table II.
IHC of Ah receptor, cytochrome P450
and HSP-90 in 25 OSCC tissue samples.
|
|
| AhR | CYP1A1 | HSP-90 |
|---|
|
|
|
|
|
|
|---|
| Samplea | Grade | Intensity
IHCb | % Positive
cellsc | Intensity
IHCb | % Positive
cellsb | Intensity
IHCb | % Positive
cellsc |
|---|
| T1 | 1 | +/− | 2 | +/− | 2 | +/− | 2 |
| T2 | 1 | +/− | 2 | + | 2 | ++ | 2 |
| T3 | 1 | +/− | 2 | + | 2 | +/− | 2 |
| T4 | 1 | +/− | 2 | + | 2 | + | 2 |
| T5 | 1 | +/− | 2 | +/− | 2 | +/− | 2 |
| T6 | 1 | + | 2 | ++ | 2 | +/− | 2 |
| T7 | 1 | +/− | 2 | +/− | 2 | + | 2 |
| T8 | 1 | +/− | 1 | +/− | 2 | + | 2 |
| T9 | 1 | +/− | 2 | + | 2 | +/− | 2 |
| T10 | 1 | +/− | 2 | + | 2 | + | 2 |
| T11 | 1 | +/− | 2 | +/− | 2 | + | 2 |
| T12 | 2 | + | 3 | + | 2 | + | 2 |
| T13 | 2 | + | 3 | ++ | 3 | + | 3 |
| T14 | 2 | ++ | 3 | ++ | 3 | + | 3 |
| T15 | 2 | + | 2 | + | 2 | + | 2 |
| T16 | 2 | + | 3 | ++ | 3 | + | 3 |
| T17 | 2 | + | 3 | ++ | 3 | + | 3 |
| T18 | 2 | + | 3 | + | 3 | + | 3 |
| T19 | 2 | + | 3 | + | 3 | + | 3 |
| T20 | 3 | ++ | 4 | + | 4 | ++ | 4 |
| T21 | 3 | ++ | 3 | ++ | 4 | ++ | 3 |
| T22 | 3 | + | 4 | ++ | 4 | ++ | 4 |
| T23 | 3 | ++ | 4 | + | 3 | ++ | 4 |
| T24 | 3 | + | 4 | ++ | 4 | ++ | 4 |
| T25 | 3 | ++ | 4 | ++ | 4 | ++ | 4 |
AHR expression
In all neoplastic samples, AHR expression was
presented by a cytoplasmic pattern; however, 50% of grade 2 (4/8)
and 80% of grade 3 (4/6) OSCCs showed nuclear immunostaining
Moreover, the intensity of immunostaining and the percentage of
positive epithelial neoplastic cells were increased according to
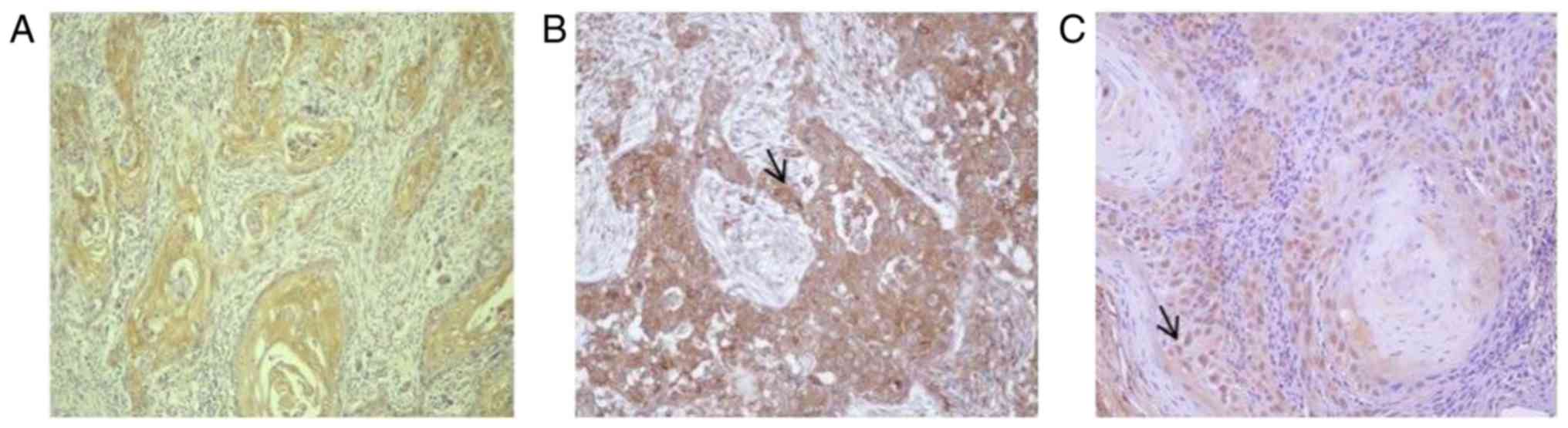
the differentiation grade of the tumor. Particularly, in almost all
G1 tumors (10/11), the percentage of positive epithelial neoplastic
cells ranged from 25 to 50% and the intensity of immunostaining was
weak (Fig. 1). The majority of
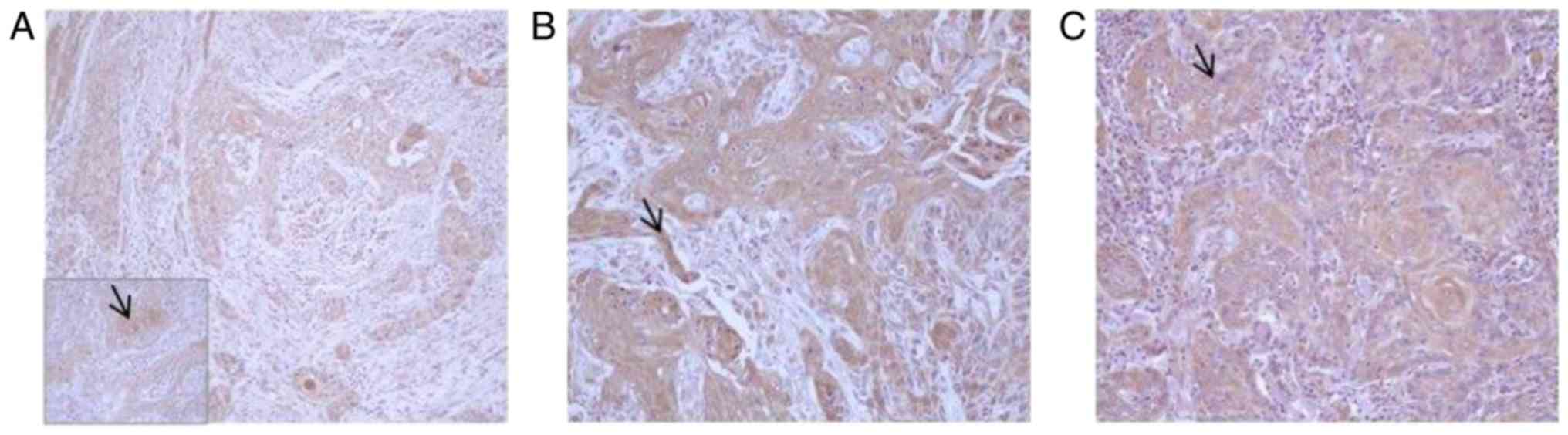
grade 2 tumors (7/8) showed a percentage of positive epithelial
neoplastic cells ranging from 50 to 75% and a moderate intensity of
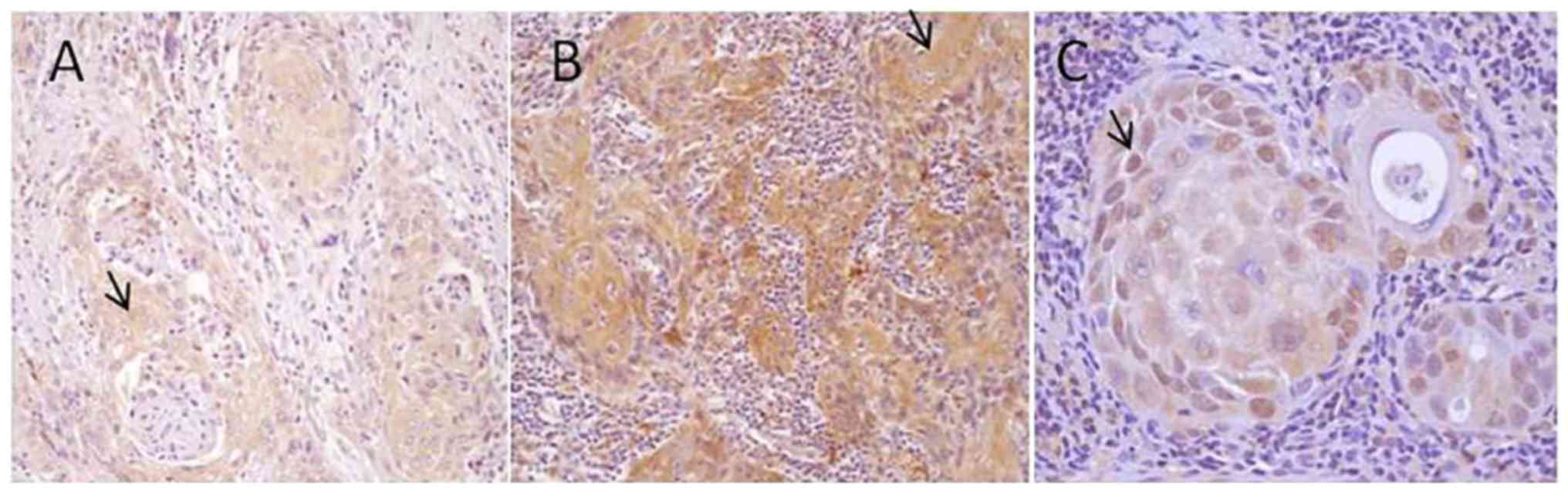
immunostaining (Fig. 2). Grade 3
tumors showed a moderate (2/6) to strong immunoreactivity (4/6) and
80–100% of epithelial neoplastic cells were positive in almost all
samples (5/6) (Fig. 3).
CYP1A1 and HSP-90 expression
In all neoplastic samples, CYP1A1 and HSP-90
expression showed a cytoplasmic pattern, particularly in grade 1
and 2, while 50% of grade 3 OSCCs (3/6) showed a nuclear
localization of HSP-90. The intensity of immunostaining and the
percentage of epithelial positive neoplastic cells were increased
proceeding from grade 1 OSCC, in which only 30% of cells were
weakly positive for both CYP1A1 (Fig.
2A) and HSP-90 (Fig. 3A), to
grade 2 (Figs. 2B and 3B) in which almost all samples (6/8)
showed a moderate immunolabeling in 50–75% neoplastic cells,
proceeding to grade 3 OSCC, in which 75–100% of neoplastic cells
were strongly positive in 5/6 samples (Figs. 2C and 3C).
In vitro effect of polydatin on AHR,
CYP1A1 and HSP-90 expression in human tongue squamous cell
carcinoma CAL27 cells
The effects of trans-polydatin on the proliferation
of human tongue squamous cell carcinoma CAL27 cells was evaluated
by MTT assay (see section reported in ‘Materials and Methods’).
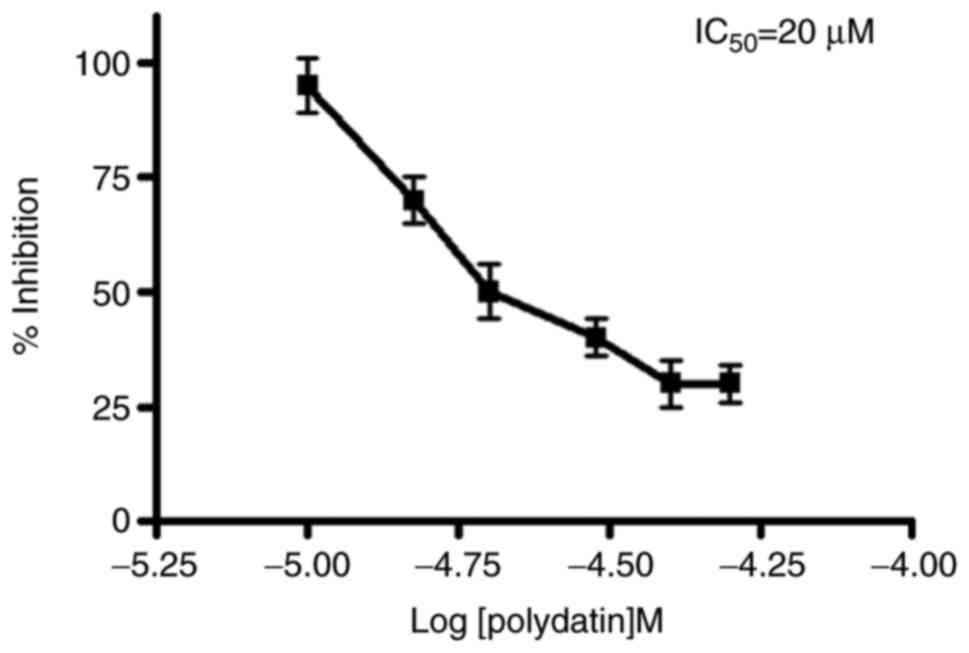
Polydatin induced a dose-dependent growth inhibition after 24 h and
the IC50 (inhibition of 50% of cell growth) was reached at the 20
µM concentration (Fig. 4).
The expression and densitometric quantification of
AHR and related proteins CYP1A1 and HSP-90 in the CAl27 cells
before and after treatment with different concentrations of
polydatin are shown (Fig. 5). The
data showed that following polydatin treatment for 24 h, a
significant decrease in AHR, Cyp and HSP-90 expression in CAL27
cells was noted compared to that in the control (cells without
treatment).
Molecular simulations
We simulated the interaction of polydatin with human
AHR ligand binding domain, similarly to a previous study (20) in order to investigate if and how the
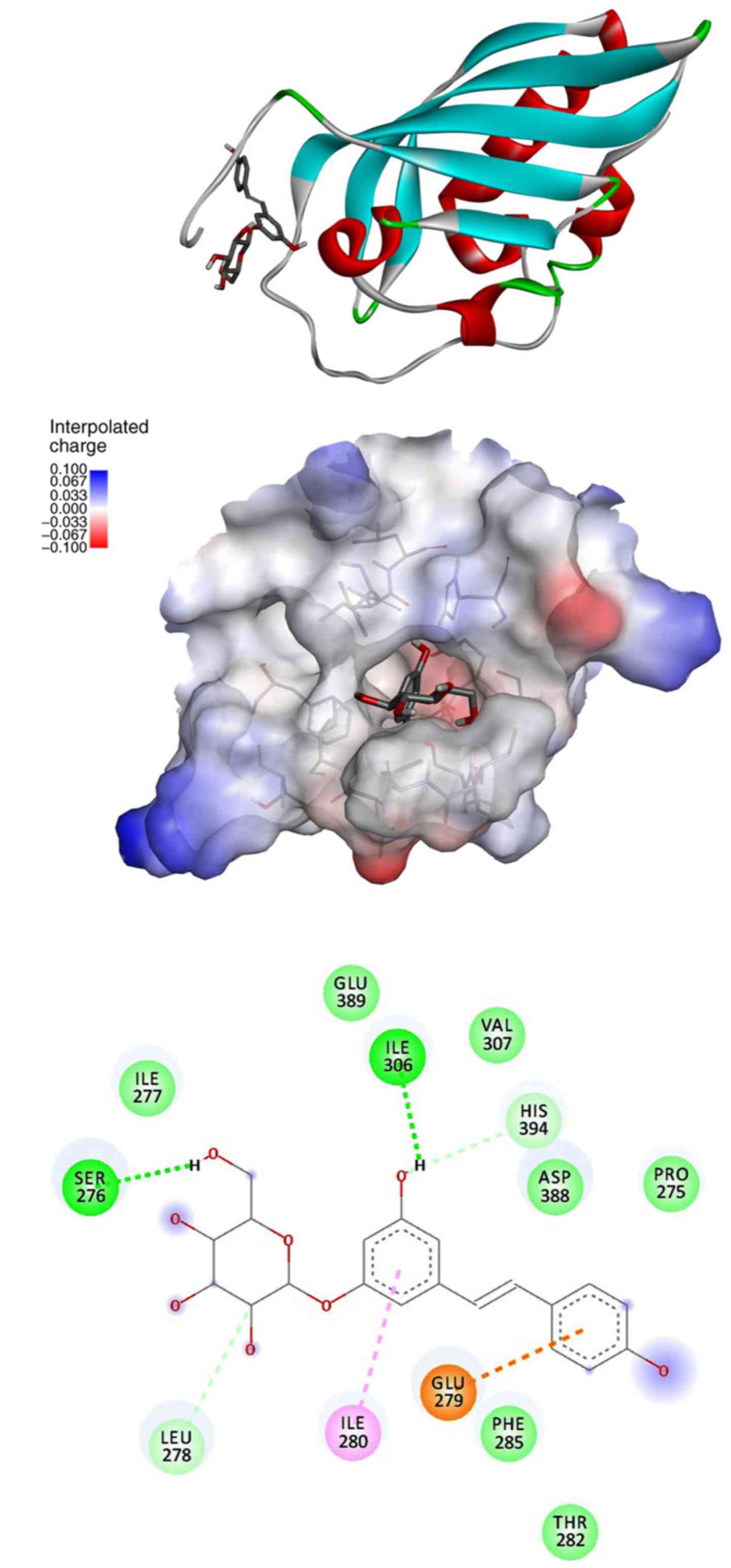
molecules may interact. In Fig. 6
we report images to describe the results. The structure of the
ligand binding domain consists of a globular domain having the
typical PAS domain architecture, which consists of an anti-parallel
five-strand β sheet and an α-helix, with a few other short
secondary structure elements. Polydatin finds a suitable pocket for
binding, similarly to a previous study with other ligands that are
evidenced by four different possible sites (20). The preferred site is that indicated
as B site in that article, and polydatin shows a mean energy of
interaction of −6.51, similar to that reported in the previous
study for TCDD (dioxin), i.e, −6.2, quercetin, i.e., −6.5, and
pentachlorobyphenyl, i.e., 6.54.
Discussion
In the present study we found that AHR expression
was extremely high in human oral squamous cancer (hOSC) tissues
(Table I), suggesting that IHC
evaluation of AHR may be considered as a prognostic marker. The
influence of AHR in tumor development of hOSC is supported by a
published study investigating AHR expression in cells derived from
head and neck cancers and demonstrating the ability of AHR
inhibitors to induce transcriptional repression of the
pro-inflammatory cytokine Il-6, to counteract the drug resistance
induced by ABCG2 and to repress cell migration and invasion
(23). More recently, the same
authors identifed three growth factors as a target of AHR,
amphiregulin (AREG), epiregulin (EREG) and platelet-derived growth
factor (PDGF) associated with metastatic phenotype and displaying a
common DRE-like promoter motif (24). These findings are consistent with
the increased AHR expression observed for other types of cancer
such as gastric and non-small cell lung cancer, and mammary gland
tumors (25–27). Increased AHR expression is likely to
increase basal AHR activity within tumors, particularly given the
systemic omnipresence of AHR agonist ligands derived from
xenobiotic, dietary and microbial sources. Moreover, a correlation
was found between increasing nuclear AHR protein expression and
increasing tumor grade, suggesting at least for these tumors that
nuclear AHR levels are predictive of a higher tumor grade.
AHR was predominantly expressed in the cytoplasm in
all grade 1 and 50% of grade 2 OSCC tissues samples; on the
contrary in 50% of grade 2 OSCC and all grade 3 OSCC the AHR
expression was prevalently nuclear. Since AHR acts as a
transcriptional regulator with nuclear translocation, the presence
of nuclear AHR in grade 2 and grade 3 samples suggest a strong
transcriptional activity of AHR in these high-grade tumors.
Research of malignant tumors showed increased levels of AHR and its
constitutive localization in the nucleus, suggesting that the AHR
is chronically activated in tumors, facilitating tumor progression
(28).
A large body of evidence has demonstrated that AHR
is the major regulatory element for CYP1A1 expression as confirmed
by our results due to high expression of AHR and CYP1A1 in high
grade OSCC (5,29,30).
In addition, we showed that HSP-90, a related cytosolic AHR-complex
protein, increased its expression from grade 1 proceeding to higher
grade OSCC, and that it had prevalently a cytoplasmic localization
in grade 1, grade 2 and 50% of grade 3 OSCC. In the other 50% of
grade 3 OSCC, HSP-90 showed a nuclear localization, confirming what
has already been demonstrated by other studies, that AHR
translocates to the nucleus while in complex with HSP-90 (19).
We studied the antitumoral effect of polydatin in
vitro, on the human tongue squamous cell carcinoma cell line
CAL27. Cytotoxic study of polydatin was evaluated by MTT assay and
showed a IC50 value of 20 µM. Our western results clearly indicated
a comparable inhibition of AHR, CYP1A1 and HSP-90 at all
concentrations utilized ranging between 10 and 30 µM. Yet, while
the AHR and CYP1A1 expression tended to disappear, the HSP-90
expression did not fully shutdown. This experimental evidence is
ascribed to the fact that the molar ratio AHR/HSP-90 is 1/2 since
HSP-90 is present as a homodimer, in double copy, in the
AHR-complex and HSP-90 is not exclusively part of the AHR-complex.
Thus, polydatin did not act as an HSP-90 inhibitor and its decrease
was AHR-dependent.
Concerning the mechanism of action of polydatin on
AHR inhibition, molecular simulations suggest that this molecule
may interact directly with human AHR ligand binding domain (middle
panel, Fig. 6), at the same site
and with similar mean energy of binding reported for dioxin,
pentachlorobyphenyl, and quercetin (20). These findings suggest a direct
interaction of polydatin in the active site of the human AHR where
the glycosidic ring does not compromise the bond of the
resveratrolic counterpart of polydatin, but that it is stabilized
by Ser 276 and Leu 278 (bottom panel, Fig. 6). Polydatin is not the only
glycoside derivative that enhances its inhibitory action on the AHR
receptor, quercetin-6-C-β-D-glucopyranoside, a natural glycosidic
analog of quercitin, was shown to interact with the active site of
the human aryl receptor (31).
Moreover, in this study the authors, utilizing the PC-3 cell line
as a model of prostate cancer, pointed out the involvement of the
Akt/mTOR pathway according to our published experiment on Caco2
gastrointestinal cancer cells showing the inhibitory effect of
polydatin on AKT expression (16).
The involvment of the AKT/mTOR pathway also leads to a autophagic
input (32) which may explain the
lack of expression of AHR that we registered in western analysis
and that was maximum at 30 µM polydatin.
In conclusion, oral squamous cell carcinoma is
increasingly associated with exposure to numerous environmental
carcinogens, including mostly tobacco smoke and its carcinogenic
components, such as polycyclic aromatic hydrocarbons (PAHs).
These carcinogens are transformed in mutagenic
compounds by catabolic activity of AHR-regulated P450 oxygenases
(29). Therefore, the modulation of
P450 1A1 activity has been considered as a potential target for
cancer chemoprevention.
The present study is defined as a ‘pilot study’
since it was performed with 25 clinical cases. The association of
AHR expression with tumor grade, although it was strong, needs to
be confirmed in a further full-scale experiment. In conclusion, we
demonstrated that: i) the majority of hOSCC tissues expressed
elevated levels of AHR, CYP1A1 and HSP-90; ii) the intensity and
the percentage of positive neoplastic cells increased proceeding
from low grade to high grade hOSCCs; iii) in vitro cytotoxic
polydatin concentration induced a decrease in AHR, CYP1A1 and
HSP-90 protein expression. Finally, molecular simulation studies
provided a new rationale use of polydatin and encourage us to
continue in vitro studies to support the use of polydatin in
oral cancer prevention and/or as alimentary support associated with
antitumoral therapy.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Poland A, Palen D and Glover E: Tumour
promotion by TCDD in skin of HRS/J hairless mice. Nature.
300:271–273. 1982. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Murray IA, Patterson AD and Perdew GH:
Aryl hydrocarbon receptor ligands in cancer: Friend and foe. Nat
Rev Cancer. 14:801–814. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Meyer BK, Pray-Grant MG, Vanden Heuvel JP
and Perdew GH: Hepatitis B virus X-associated protein 2 is a
subunit of the unliganded aryl hydrocarbon receptor core complex
and exhibits transcriptional enhancer activity. Mol Cell Biol.
18:978–88. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhou S, Liu J and Chowbay B: Polymorphism
of human cytochrome P450 enzymes and its clinical impact. Drug
Metab Rev. 41:89–295. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stanford EA, Ramirez-Cardenas A, Wang Z,
Novikov O, Alamoud K, Koutrakis P, Mizgerd JP, Genco CA,
Kukuruzinska M, Monti S, et al: Role for the Aryl hydrocarbon
receptor and diverse ligands in oral squamous cell carcinoma
migration and tumorigenesis. Mol Cancer Res. 14:696–706. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Quattrochi LC and Tukey RH: Nuclear uptake
of the Ah (dioxin) receptor in response to omeprazole:
Transcriptional activation of the human CYP1A1 gene. Mol Pharmacol.
43:504–508. 1993.PubMed/NCBI
|
|
7
|
Singh SU, Casper RF, Fritz PC, Sukhu B,
Ganss B, Girard B Jr, Savouret JF and Tenenbaum HC: Inhibition of
dioxin effects on bone formation in vitro by a newly described aryl
hydrocarbon receptor antagonist, resveratrol. J Endocrinol.
167:183–195. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lee JE and Safe S: Involvement of a
post-transcriptional mechanism in the inhibition of CYP1A1
expression by resveratrol in breast cancer cells. Biochem
Pharmacol. 62:1113–1124. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
He H, Zhao Y, Chen X, Zheng Y, Wu X, Wang
R, Li T, Yu Q, Jing J, Ma L, et al: Quantitative determination of
trans-polydatin, a natural strong anti-oxidative compound, in rat
plasma and cellular environment of a human colon adenocarcinoma
cell line for pharmacokinetic studies. J Chromatogr B Analyt
Technol Biomed Life Sci. 855:145–151. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Henry-Vitrac C, Desmoulière A, Girard D,
Mérillon JM and Krisa S: Transport, deglycosylation, and metabolism
of trans-piceid by small intestinal epithelial cells. Eur J Nutr.
45:376–382. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Rotches-Ribalta M, Andres-Lacueva C,
Estruch R, Escribano E and Urpi-Sarda M: Pharmacokinetics of
resveratrol metabolic profile in healthy humans after moderate
consumption of red wine and grape extract tablets. Pharmacol Res.
66:375–382. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ravagnan GP: Extracts from spermatophyte
plants with antitumor activity Euoropean Patent EP1292319B1. Filed
May 29, 2001; issued Aug 15. 2002
|
|
13
|
Wang C, Luo Y, Lu J, Wang Y and Sheng G:
Polydatin induces apoptosis and inhibits growth of acute monocytic
leukemia cells. J Biochem Mol Toxicol. 30:200–205. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cao WJ, Wu K, Wang C and Wan DM:
Polydatin-induced cell apoptosis and cell cycle arrest are
potentiated by Janus kinase 2 inhibition in leukemia cells. Mol Med
Rep. 13:3297–3302. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang Y, Zhuang Z, Meng Q, Jiao Y, Xu J
and Fan S: Polydatin inhibits growth of lung cancer cells by
inducing apoptosis and causing cell cycle arrest. Oncol Lett.
7:295–301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
De Maria S, Scognamiglio I, Lombardi A,
Amodio N, Caraglia M, Cartenì M, Ravagnan G and Stiuso P:
Polydatin, a natural precursor of resveratrol, induces cell cycle
arrest and differentiation of human colorectal Caco-2 cell. J
Transl Med. 11:2642013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hogg SJ, Chitcholtan K, Hassan W, Sykes PH
and Garrill A: Resveratrol, acetyl-resveratrol, and polydatin
exhibit antigrowth activity against 3D cell aggregates of the
SKOV-3 and OVCAR-8 ovarian cancer cell lines. Obstet Gynecol Int.
2015:2795912015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hughes D, Guttenplan JB, Marcus CB,
Subbaramaiah K and Dannenberg AJ: Heat shock protein 90 inhibitors
suppress aryl hydrocarbon receptor-mediated activation of CYP1A1
and CYP1B1 transcription and DNA adduct formation. Cancer Prev Res.
1:485–493. 2008. View Article : Google Scholar
|
|
19
|
Tsuji N, Fukuda N, Nagata Y, Okada H, Haga
A, Hatakeyama S, Yoshida S, Okamoto T, Hosaka M, Sekine K, et al:
The activation mechanism of the aryl hydrocarbon receptor (AhR) by
molecular chaperone HSP-90. FEBS Open Bio. 4:796–803. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Salzano M, Marabotti A, Milanesi L and
Facchiano A: Human aryl-hydrocarbon receptor and its interaction
with dioxin and physiological ligands investigated by molecular
modelling and docking simulations. Biochem Biophys Res Commun.
413:176–181. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bryne M, Koppang HS, Lilleng R, Stene T,
Bang G and Dabelsteen E: New malignancy grading is a better
prognostic indicator than Broder's grading in oral squamous cell
carcinomas. J Oral Pathol Med. 18:432–437. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martano M, Carella F, Squillacioti C,
Restucci B, Mazzotta M, Lo Muzio L and Maiolino P: Metallothionein
expression in canine cutaneous apocrine gland tumors. Anticancer
Res. 32:747–752. 2012.PubMed/NCBI
|
|
23
|
John K, Lahoti TS, Wagner K, Hughes JM and
Perdew GH: The Ah receptor regulates growth factor expression in
head and neck squamous cell carcinoma cell lines. Mol Carcinog.
53:765–76. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
DiNatale BC, Smith K, John K, Krishnegowda
G, Amin SG and Perdew GH: Ah receptor antagonism represses head and
neck tumor cell aggressive phenotype. Mol Cancer Res. 10:1369–1379.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yin XF, Chen J, Mao W, Wang YH and Chen
MH: Downregulation of aryl hydrocarbon receptor expression
decreases gastric cancer cell growth and invasion. Oncol Rep.
30:364–370. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Tsai CH, Li CH, Cheng YW, Lee CC, Liao PL,
Lin CH, Huang SH and Kang JJ: The inhibition of lung cancer cell
migration by AhR-regulated autophagy. Sci Rep. 7:419272017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brooks J and Eltom SE: Malignant
transformation of mammary epithelial cells by ectopic
overexpression of the aryl hydrocarbon receptor. Curr Cancer Drug
Targets. 11:654–669. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Al-Dhfyan A, Alhoshani A and Korashy HM:
Aryl hydrocarbon receptor/Cytochrome P450 1A1 pathway mediates
breast cancer stem cells expansion through PTEN inhibition and
β-Catenin and Akt activation. Mol Cancer. 16:692017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Androutsopoulos VP, Tsatsakis AM and
Spandidos DA: Cytochrome P450 CYP1A1: Wider roles Androutsopoulos
in cancer progression and prevention. BMC Cancer. 9:1872009.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Moyer BJ, Rojas IY, Murray IA, Lee S,
Hazlett HF, Perdew GH and Tomlinson CR: Indoleamine 2,3-dioxygenase
1 (IDO1) inhibitors activate the aryl hydrocarbon receptor. Toxicol
Appl Pharmacol. 323:74–80. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hamidullah, Kumar R, Saini KS, Kumar A,
Kumar S, Ramakrishna E, Maurya R, Konwar R and Chattopadhyay N:
Quercetin-6-C-β-D-glucopyranoside, natural analog of quercetin
exhibits anti-prostate cancer activity by inhibiting Akt-mTOR
pathway via aryl hydrocarbon receptor. Biochimie. 119:68–79. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chang CH, Lee CY, Lu CC, Tsai FJ, Hsu YM,
Tsao JW, Juan YN, Chiu HY, Yang JS and Wang CC: Resveratrol-induced
autophagy and apoptosis in cisplatin-resistant human oral cancer
CAR cells: A key role of AMPK and Akt/mTOR signaling. Int J Oncol.
50:873–882. 2017. View Article : Google Scholar : PubMed/NCBI
|