Introduction
Lung cancer is the leading cause of
cancer-associated mortality worldwide, and non-small cell lung
cancer (NSCLC) accounts for 80–85% all lung cancer cases (1). Tumors harboring somatic activating
mutations in the exon encoding the kinase domain of epidermal
growth factor receptor (EGFR), account for 10–15% (2,3) and
40% (4) of NSCLC cases in the
Western and Asian populations, respectively. EGFR tyrosine kinase
inhibitors (TKIs) present a novel paradigm for molecular targeted
therapeutics for patients with NSCLC and are recommended as
first-line treatments for patients with advanced NSCLC harboring an
EGFR-TKI-sensitizing mutation (EGFRm) (5,6).
Despite reports of high response rates with first-line EGFR-TKI
therapy, the majority of responsive patients ultimately develop
disease progression following 9–14 months of treatment, and ~50%
cases are caused by the acquisition of the T790M mutation (7–9).
Osimertinib (TAGRISSO™, AZD9291) is an oral, irreversible, third
generation EGFR-TKI that targets EGFRm and T790M (10,11).
It has been approved by the US Food and Drug Administration for the
management of T790M-positive NSCLC that has progressed following
the introduction of first-line EGFR-TKI treatment (12). Furthermore, a recently published
study (FLAURA) demonstrated that osimertinib as a first-line
treatment for patients with EGFRm and locally advanced or
metastatic NSCLC, significantly extended progression-free survival
when compared with patients treated with first generation EGFR-TKIs
(18.9 vs. 10.2 months) (13).
Radiotherapy is an important treatment modality for
lung cancer, particularly for patients who are ineligible for
surgery. Approximately 2/3 of patients with lung cancer receive
radiotherapy during the course of treatment, with either definitive
or palliative intent (14).
However, the effectiveness of radiotherapy is often limited by
intrinsic radio-resistance. The expression and activity of EGFR are
determinants of the radioresponse in patients with NSCLC, and
anti-EGFR therapy has been shown to enhance the radiosensitivity of
tumor cells (15–17). Recent data demonstrated that
patients with EGFR mutations have improved outcomes when treated
with a combination of first-generation EGFR-TKIs and radiotherapy
(18–21). However, to the best of our
knowledge, no previous studies have discussed the effectiveness of
osimertinib combined with ionizing radiation (IR).
In the present study, the effects of osimertinib on
the radiosensitivity of NSCLC cells with T790M/L858R mutations were
evaluated in vitro and in vivo.
Materials and methods
Cell lines and reagents
The human lung cancer cell line, NCI-H1975
(L858R/T790M), was obtained from the American Type Culture
Collection (Manassas, VA, USA) and authenticated by short-tandem
repeat analysis in December 2016. The cells were maintained in
RPMI-1640 (cat. no. 22400; Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA), containing 10% fetal bovine serum
(Hyclone; GE Healthcare Life Sciences, Logan, UT, USA; cat. no.
SV30087) in a humidified incubator at 37°C with 5% CO2.
Osimertinib (AZD9291) was provided by AstraZeneca (Shanghai,
China), and was dissolved in dimethyl sulfoxide (DMSO;
Sigma-Aldrich; Merck KGaA, Darmstadt, Germany; cat. no. SHBG6226v)
for in vitro study and 0.5% Hydroxy propyl methyl cellulose
(HPMC; Sigma-Aldrich; Merck KGaA; cat. no. 9004-67-5)/0.1% Tween-80
(cat. no. P1754; Sigma-Aldrich; Merck KGaA) for the in vivo
study.
Irradiation
The cells and xenografts were irradiated using the
X-cell 160 Irradiator (137Cs; Kubtec, Milford, CT, USA)
at a dose rate of 140 cGy/min at room temperature with the
following protocol: 1.42 min at 2 Gy, 2.85 min at 4 Gy, 4.26 min at
6 Gy, 5.68 min at 8 Gy, 7.10 min at 10 Gy and 14.2 min at 20 Gy,
respectively. Cells were treated with Osimertinib at 37°C 1 h prior
to irradiation in the combination groups involved in the in
vitro study.
In vitro cell proliferation assays
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetra
zolium, inner salt (MTS) assay]
Cells were divided into 6 groups: i) Osimertinib
alone [at dosages of 0 (control), 0.0001, 0.001, 0.003, 0.01, 0.03,
0.1, 0.3, 1 and 3 µM, respectively); ii) osimertinib with 2 Gy
irradiation; iii) osimertinib with 4 Gy irradiation; iv)
osimertinib with 6 Gy irradiation; v) osimertinib with 8 Gy
irradiation; and vi) osimertinib with 10 Gy irradiation. The
proliferation analysis was performed using a tetrazolium-based Cell
Titer 96® Aqueous One Solution Assay (cat. no. G3581;
Promega Corporation, Madison, WI, USA), according to the
manufacturer's instructions. Briefly, exponentially growing cells
were diluted to 2×104/ml and seeded at 100 µl/well into
96-well plates. Following 24 h, cells were treated with irradiation
and increasing concentrations of osimertinib. Then, an MTS assay
was performed following 3 days. Relative cell viability was
expressed as the percentage of untreated control.
In vitro cell clone formation assay
(CFA)
Cells in the log phase were plated into 6-well
plates with the desired cell density (300 cells/well for 0 Gy, 500
cells/well for 2 Gy, 1,000 cells/well for 4 Gy, 3,000 cells/well
for 6 Gy and 5,000 cells/well for 8 Gy, respectively) and
pretreated with osimertinib at 10, 30 and 100 nM, or DMSO,
respectively. Irradiation was delivered 1 h later. Cells were then
maintained for 14 days with osimertinib in RPMI-1640 and stained
with 0.1% crystal violet (Sigma-Aldrich; Merck KGaA; cat. no.
5K219R5). Colonies of >50 cells were defined as surviving
colonies and the number of colonies was normalized to that of
non-irradiated controls. The sensitizer enhancement ratio (SER) for
osimertinib treatment was calculated as the ratio of the mean
inactivation dose of control cells / the mean inactivation dose of
osimertinib-treated cells at the 0.01 survival fraction.
Flow assisted cell sorting (FACS)
assay
Cells were trypsinized with 0.25% trypsin-EDTA (cat.
no. 25200-114; Invitrogen; Thermo Fisher Scientific, Inc.) and the
suspended cell pellet was incubated with 70% ethanol (cat. no.
1000927; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) at
4°C. Following the thorough removal of ethanol, the cells were
suspended in a proprium iodide (PI) staining solution (cat. no.
P3566; Invitrogen; Thermo Fisher Scientific, Inc.) in the dark, at
room temperature for 30 min. Flow cytometry was performed using a
FACSCanto Cell Analyzer (V07300617; BD Biosciences, Franklin Lakes,
NJ, USA).
Immunofluorescence assay
Cells were cultured on chamber slides and the
samples were collected at 2, 24 and 48 h, respectively. Cells were
washed with ice-cold Ca2+/Mg2+-free phosphate
buffered saline (PBS), and fixed in 4% formaldehyde (cat. no.
SZBF0690v; Sigma-Aldrich; Merck KGaA) for 60 min at room
temperature. Following permeabilization in 1% Triton X-100 (cat.
no. 057K00161; Sigma-Aldrich; Merck KGaA) and blocking with 5%
bovine serum albumin (BSA; cat. no. 12575v; Sigma Aldrich; Merck
KGaA)/0.3% Triton™ X-100 in PBS at room temperature for 1 h, the
cells were incubated with a fluorescein isothiocyanate-conjugated
anti-phospho histone γ-H2A histone family member X (H2AX) primary
antibody (dilution, 1:800; cat. no. 2577s; CST Biological Reagents
Co., Ltd., Shanghai, China) overnight at 4°C, then incubated with
an Alexa 647-conjugated anti-rabbit secondary antibody (dilution,
1:1,000; cat. no. 4412S; CST Biological Reagents Co., Ltd.) for 1 h
at room temperature in the dark. Coverslips were mounted using
Mounting Medium with DAPI (H-1200; Vector Laboratories, Inc.,
Burlingame, CA, USA) overnight at 4°C. Images were acquired with an
Olympus BX61 laser scanning confocal microscope (7E18988; Olympus
Corporation, Tokyo, Japan) using ×60 magnification. Using ≥150
cells from each experiment, the cells were counted and the
percentage of cells positive for γ-H2AX was calculated. A positive
cell was defined by >5 discrete dots in the nucleus.
Western blotting
Cells were lysed in 2X SDS buffer containing
protease and phosphatase inhibitors (cat. no. 1861282; Thermo
Fisher Scientific, Inc.), then the protein concentration was
determined via a BCA protein assay (cat. no. 34076; Thermo Fisher
Scientific, Inc.). Equal amounts of protein (20 µg/well) were
loaded for SDS-PAGE using 4–12%-gradient Bis-Tris precast gels
(cat. no. 345-0124; Bio-Rad Laboratories, Inc., Hercules, CA, USA),
followed by transfer to polyvinylidene difluoride membranes using
the iBlot dry transfer system (cat. no. IB21001; Novex; Thermo
Fisher Scientific, Inc.). Membranes were blocked in 5% fat-free
milk in Tris-buffered saline with 0.1% Tween-20 (TBS-T) for 30 min
at room temperature, then incubated at 4°C overnight with the
following primary antibodies: Total EGFR (dilution, 1:1,000; cat.
no. 4267S; CST Biological Reagents Co., Ltd.), phospho (p)-EGFR
(dilution, 1:1,000; Tyr1068l, cat. no. 3777S; or Tyr1173, cat. no.
4407S; CST Biological Reagents Co., Ltd.), total protein kinase B
(AKT; dilution, 1:1,000; cat. no. 4691S; CST Biological Reagents
Co., Ltd.), phospho-AKT (Ser473; dilution, 1:1,000; cat. no.
12694S; CST Biological Reagents Co., Ltd.), total extracellular
signal-regulated kinase (ERK; dilution, 1:1,000; cat. no. 4695S;
CST Biological Reagents Co., Ltd.), phospho-ERK (dilution, 1:1,000;
Thr202/Tyr204; cat. no. 4370S; CST Biological Reagents Co., Ltd.)
and GAPDH (dilution, 1:10,000; cat. no. 2118L; CST Biological
Reagents Co., Ltd.). Membranes were then incubated for 1 h at room
temperature with a horseradish peroxidase (HRP)-conjugated
secondary antibody (dilution, 1:2,000; cat. no. 7074 or 7076; CST
Biological Reagents Co., Ltd.). Membranes were visualized using a
mixed detection solution (Super Signal West Dura Extended Duration
Substrate; cat. no. 34076; Thermo Fisher Scientific, Inc.) for 5
min at room temperature, protected from light. Densitometry of
western blots was conducted using a Fujifilm Image Reader LAS-4000
2.1 (Fujifilm Corp., Tokyo, Japan).
Mice and tumor model
Specific pathogen-free immunodeficient female nude
mice (n=170; age, 6–8 weeks; weight, 20–25 g) were purchased from
Beijing Vital River Laboratory Animal Technology (Beijing, China).
Animals had free access to food and water, and were maintained at
21–23°C with 40–70% humidity and a 12-h light/dark cycle. All of
the animal studies were approved by the Institutional Animal Care
and Use Committee of AstraZeneca (Shanghai, China). NCI-H1975 tumor
cells (5×106 cells/0.1 ml) were injected subcutaneously
into the left flank of the nude mice to establish the tumor
model.
In vivo efficacy study
Tumor growth was monitored twice weekly by caliper
measurements. When the tumors grew to 0.2–0.4 cm3, the
mice were randomized into 6 groups: i) vehicle (no treatment); ii)
osimertinib alone (5 mg/kg/day); iii) 2 Gy × 10 F irradiation alone
(5 fractions/week from the first day); iv) 2 Gy × 10 F irradiation
and osimertinib (5 mg/kg/day); v) 20 Gy × 1 F irradiation alone on
the first day; and vi) 20 Gy × 1 F irradiation and osimertinib (5
mg/kg/day). Osimertinib was administered at 5 mg/kg once daily by
oral gavage for 31 days. Mice were euthanatized (by excessive
CO2 inhalation) 25 days following the termination of
osimertinib treatment to evaluate the persistent inhibitory effect,
or when tumor volume ≥1,500 mm3 according to the Animal
Welfare of the Institutional Animal Care and Use Committee of
AstraZeneca (10). Power analysis
was performed whereby group sizes were calculated to enable
statistically robust detection of tumor growth inhibition. Tumor
growth inhibition from the start of treatment was assessed by
comparing the mean change in tumor volume of the control and
treatment groups.
In vivo drug metabolism and
pharmacokinetics (DMPK/PD) assay
For pharmacodynamic studies, mice were randomized
when the tumor volumes reached 0.5–0.8 cm3: Mice were
treated with either a single dose of osimertinib, IR alone (20 Gy ×
1 F), or osimertinib plus IR. Following a single dose, the blood
plasma was collected, except for that of the IR 20 Gy × 1 F group
due to the absence of osimertinib in the plasma, and the tumors of
all groups were harvested 0, 0.5, 1, 2, 4, 7, 16 and 24 h later.
The total osimertinib concentration in plasma was detected using
the ACQUITY SM Method, as previously described (22). Sections were fixed with 4% formalin
for 24 h at room temperature, embedded in paraffin and were then
immunohistochemically stained for the phosphorylated forms of EGFR
(Tyr1068) and EGFR (Tyr1173).
Immunohistochemistry (IHC)
IHC was performed on 3-µm sections using a Lab
Vision autostainer. The tumors were excised as aforementioned for
the DMPK/PD assay. The paraffin slides were incubated at 56°C for
30 min then dewaxed and rehydrated in a Leica Autostainer XL and
subjected to antigen retrieval (cat. no. S1699; Dako; Agilent
Technologies, Inc., Santa Clara, CA, USA) for 15 min at room
temperature followed by washing under running tap water for 5 min.
Then the sections were rinsed in TBS-T, and assessed on a LabVision
autostainer. Following incubation with an endogenous peroxidase
blocker (3% hydrogen peroxide; cat. no. GTX30967; GeneTex, Inc.,
Irvine, CA, USA) for 10 min at room temperature, slides were washed
twice in TBS-T and blocked with 5% BSA (cat. no. 12575v; Sigma
Aldrich; Merck KGaA) at room temperature for 10 min. Sections were
then incubated with the following primary antibodies for 60 min at
room temperature: p-EGFR Tyr1068 (dilution, 1:200; cat. no. 2234;
CST Biological Reagents Co., Ltd.), p-EGFR Tyr1173 (dilution,
1:200; cat. no. 4407; CST Biological Reagents Co., Ltd.), Histone
H2AX phospho (ser139; dilution, 1:150; cat. no. 2577; CST
Biological Reagents Co., Ltd.) and cleaved caspase-3 (CC3) (Asp175;
dilution, 1:200; cat. no. 9661; CST Biological Reagents Co., Ltd.).
The sections were then washed twice in TBS-T. The p-EGFR
(Tyr1068/Tyr1073)/CC3 slides were incubated with a biotinylated
goat anti-rabbit immunoglobulin secondary antibody (cat. no. E0432;
dilution, 1:100; Dako; Agilent Technologies, Inc.) and
Streptavidin-Peroxidase for 30 min at room temperature. The Histone
H2AX phospho slides were incubated with EnVision system-HRP Labeled
Polymer Anti-Rabbit for 30 min at room temperature and washed twice
in TBS-T. All sections were incubated in diaminobenzidine substrate
for 5 min at room temperature and rinsed with tap water. The
sections were then counter stained with Mayer's hematoxylin for 5
min at room temperature, dehydrated and cleared with xylene in a
Leica XL autostainer, and finally sealed in the ClearVue automated
cover slipper. Images were acquired with a laser scanning confocal
microscope (Olympus BX61 microscope; Olympus Corporation).
Statistical analysis
Experiments were performed ≥3 times and data are
presented as the mean ± standard error of the mean. Statistical
analysis was performed using SPSS 19.0 (IBM Corp., Armonk, NY,
USA). One-way analysis of variance (ANOVA) with a Bonferroni post
hoc test were conducted for comparisons of the percentage of H2AX
positive cells in the immunofluorescence assay, percentage of cells
in S and G2/M phase in the flow cytometry assay, the tumor volume
25 days following the final day of osimertinib treatment and the
quantitative changes of protein phosphorylation in the western
blotting and IHC. Two-way ANOVA with a Bonferroni post hoc test was
applied for the comparison of tumor volume in the tumor growth
curves. Pearson coefficient-parametric analysis was performed to
determine whether there was a correlation between the
phosphorylated forms of EGFR (Tyr1068/Tyr1173) and the
concentration of osimertinib in the blood plasma. P<0.05 was
considered to indicate a statistically significant difference.
Results
Osimertinib promotes the inhibition of
cell proliferation in NSCLC following irradiation
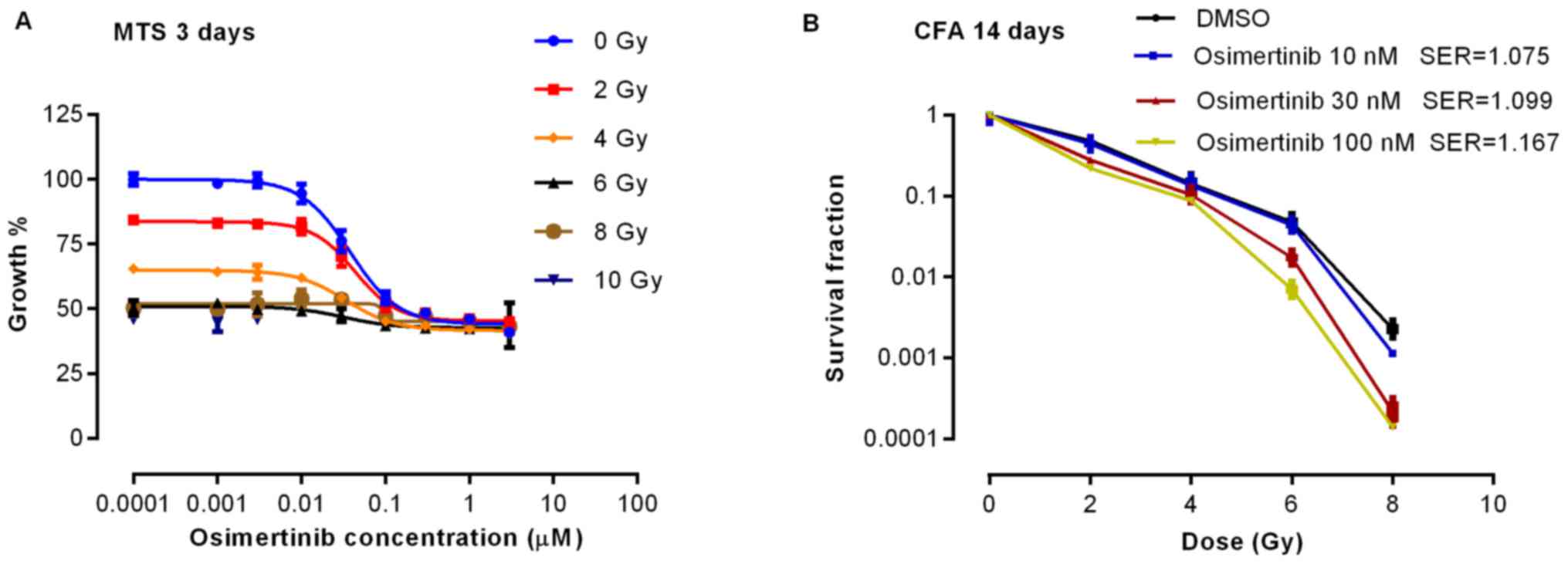
To determine the effect of osimertinib combined with
irradiation on cell proliferation, the present study performed a
cell viability assay 3 days following treatment. As shown in
Fig. 1A, the rate of cell
proliferation decreased markedly when osimertinib was administrated
prior to 2 or 4 Gy irradiation. Proliferation was demonstrated to
be inhibited in a dose-dependent manner by CFA in which the SER was
>1, indicating that the effect on proliferation inhibition was
synergistic when irradiation was combined with osimertinib
treatment (Fig. 1B).
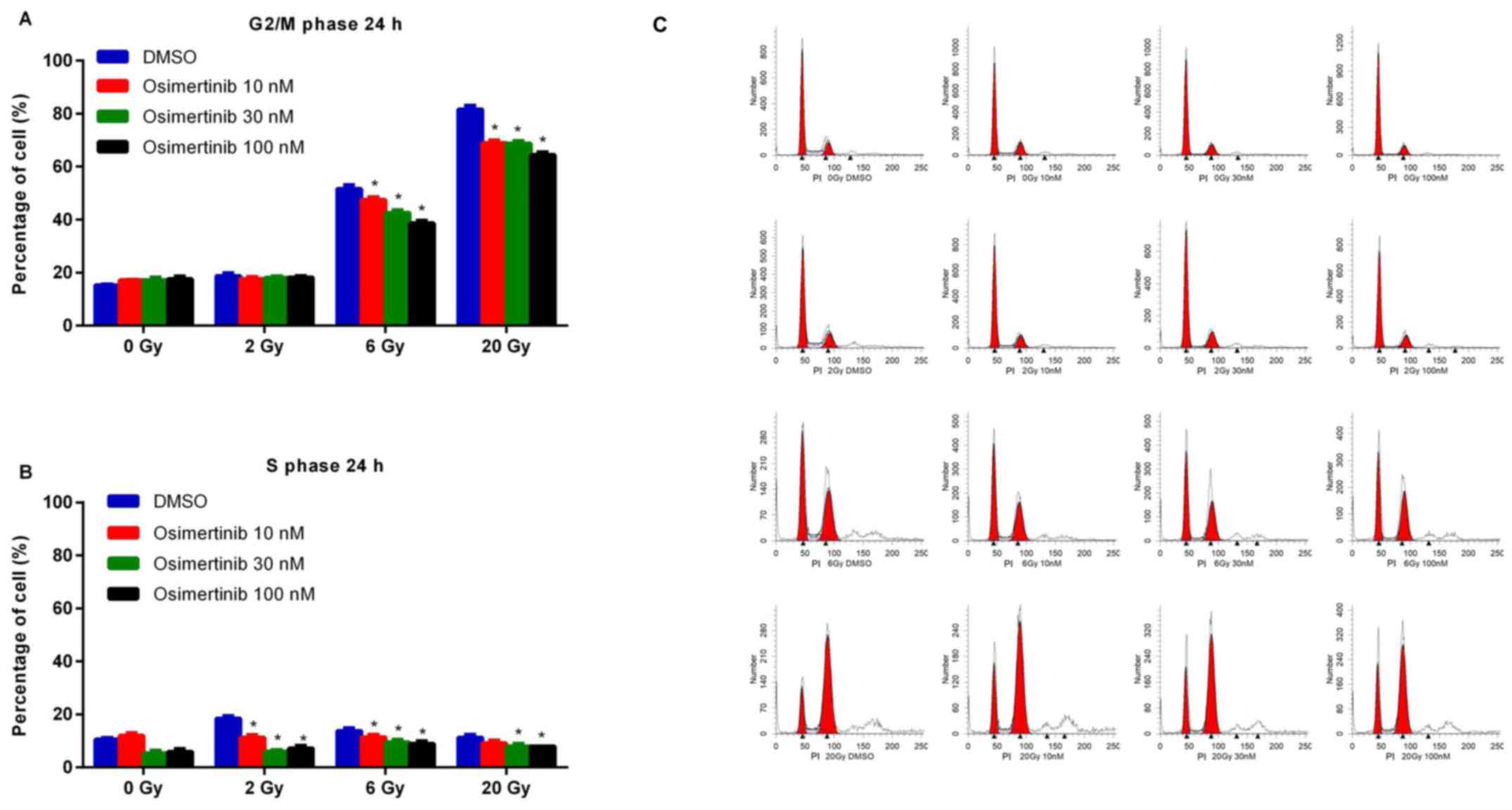
Osimertinib reduces G2/M-phase cell
cycle arrest following irradiation
To assess the effects of osimertinib on cell cycle
arrest following irradiation, cells were treated with 10, 30 or 100
nM osimertinib for 24 h and then irradiated with dose of 2, 6 or 20
Gy due to their different role in clinical practice: 2 Gy was
recognized as the conventional fraction; 6 Gy was chosen due to the
best inhibiting effect as determined by CFA in Fig. 1; and 20 Gy represented
hypofractionated radiotherapy. As shown in Fig. 2, dose-dependent reductions in the
G2/M and S phases were demonstrated in the combination treatment
group when compared with irradiation alone (DMSO group).
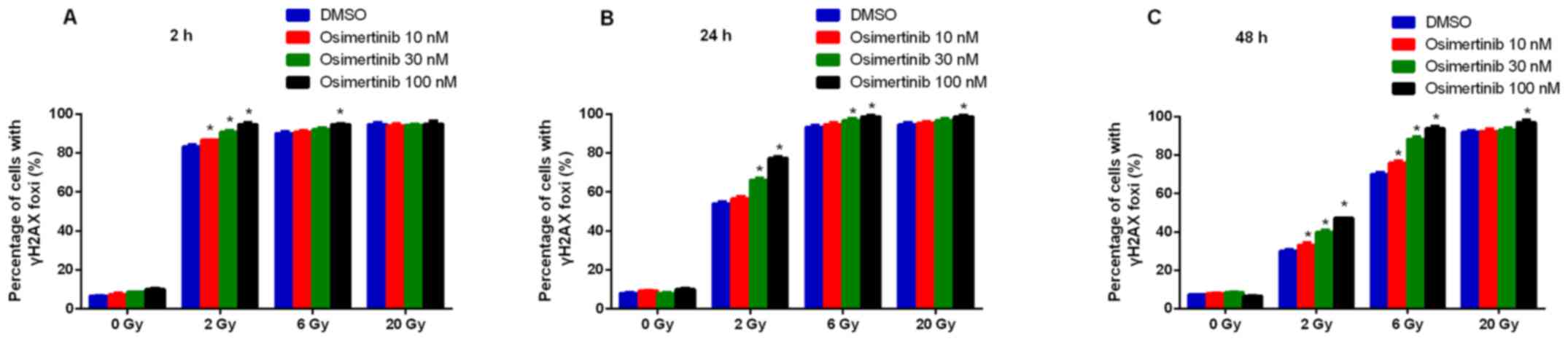
Osimertinib inhibits IR-induced DNA
double-strand breaks (DSB) repair in NCI-H1975 cells
To determine whether the increased radio-sensitivity
of cell lines following osimertinib treatment was a product of
compromised DNA-break repair, the present study conducted
immunofluorescence staining for γ-H2AX in NCI-H1975 cells. It was
revealed that the formation of γ-H2AX began to increase in the 2
and 6 Gy groups at 2 (Fig. 3A) and
24 h (Fig. 3B); whilst the
formation of γ-H2AX only began to increase at 24 h in the 20 Gy
group (Fig. 3C). In addition,
osimertinib significantly increased the number of γ-H2AX foci per
cell at 48 h following 2 and 6 Gy radiation in a dose-dependent
manner (Fig. 3C and D).
Osimertinib reduces the
phosphorylation of EGFR involved in the AKT/ERK signaling pathway
following IR
Osimertinib was revealed to be a potent inhibitor of
EGFR and downstream signaling substrate (p-AKT and p-ERK)
phosphorylation in cells with mutant EGFR (10). Therefore, the present study
investigated whether AKT/ERK were the main downstream targets of
EGFR proteins following treatment with osimertinib combined with
radiation. The western blotting results demonstrated that
osimertinib inhibits p-EGFR (1068)/p-EGFR (1173)/p-AKT/p-ERK
protein expression when treated alone or in combination with IR in
a concentration-dependent manner (Fig.
4).
 | Figure 4.Western blot analysis of the effect
of osimertinib combined with IR on EGFR phosphorylation levels
in vitro. Cells were treated with osimertinib 1 h prior to
IR, and cell proteins were collected 2 h following IR treatments.
GAPDH was included as a loading control. (A) Western blot analysis.
(B-E) Quantitative analysis of changes in (B) p-EGFR (1068), (C)
p-EGFR (1173), (D) p-AKT and (E) p-ERK, respectively. *P<0.05
vs. IR alone; #P<0.05 vs. DMSO control. IR,
irradiation; DMSO, dimethyl sulfoxide; p-, phosphorylated; EGFR,
epidermal growth factor receptor; AKT, protein kinase B; ERK,
extracellular signal-regulated kinase. |
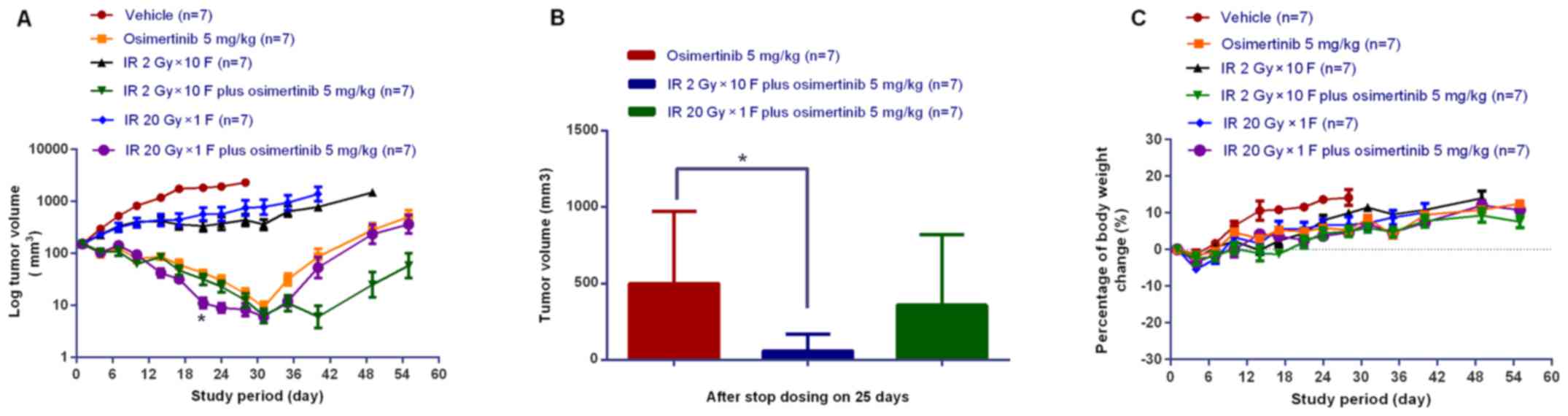
Osimertinib combined with IR induces
tumor regression in the NCI-H1975 ×enograft model
To explore the in vivo activity of
osimertinib combined with IR, the present study administered
osimertinib during the irradiation of nude mice bearing NCI-H1975
subcutaneous xenografts. As shown in Fig. 5A, the combination treatment
exhibited more potent antitumor efficacy when compared with IR or
osimertinib alone. At the end of the treatments, the mean residual
tumor volume in the IR 2 Gy × 10 F plus osimertinib 5 mg/kg group
was lower than that of osimertinib alone or osimertinib plus IR 20
Gy × 1 F groups (Fig. 5A and B). In
addition, the tumor complete response rates were 1/7, 3/7 or 4/7
for the osimertinib alone, osimertinib plus IR 20 Gy × 1 F or
osimertinib plus IR 2 Gy × 10 F groups, respectively, at the time
of treatment termination (day 31). Inhibition of tumor growth was
observed for an additional 25 days following treatment termination
(Fig. 5A and B). It was
demonstrated that radiation of conventional fraction may be more
powerful than hypofractionated radiotherapy when combined with
osimertinib administration; however, this requires further
investigation in future clinical observations. The present study
also measured mouse body weight in order to assess the treatment
tolerability, and no evident body weight changes were observed
(<5% of starting body weight), as shown in Fig. 5C. These results suggested that
treatment with IR combined with osimertinib was tolerated well.
DMPK/PD correlation and osimertinib
target-inhibition confirmation in combination with IR in NCI-H1975
subcutaneous xenograft models
To confirm the target and pathway of osimertinib
activity when in combination with IR, the present study examined
the total osimertinib concentration in the plasma and the
phosphorylation level of EGFR in tumor tissues following
treatments. In the osimertinib and osimertinib plus IR groups, the
expression of p-EGFR (Tyr1068)/p-EGFR (Tyr1173) was negatively
associated with the plasma concentration of osimertinib (Fig. 6A and B). As displayed by the
representative IHC quantification of p-EGFR (Tyr1068)/p-EGFR
(Tyr1173)/γ-H2AX and cleaved caspase-3 (Fig. 6C-G), osimertinib suppressed the
activity of p-EGFR (Tyr1068)/p-EGFR (Tyr1173), particularly when
administrated combined with IR; while the phosphorylation of EGFR
maintained high levels in the IR only group. However, the
expression levels of γ-H2AX and cleaved caspase-3 significantly
increased in the osimertinib plus IR combination group.
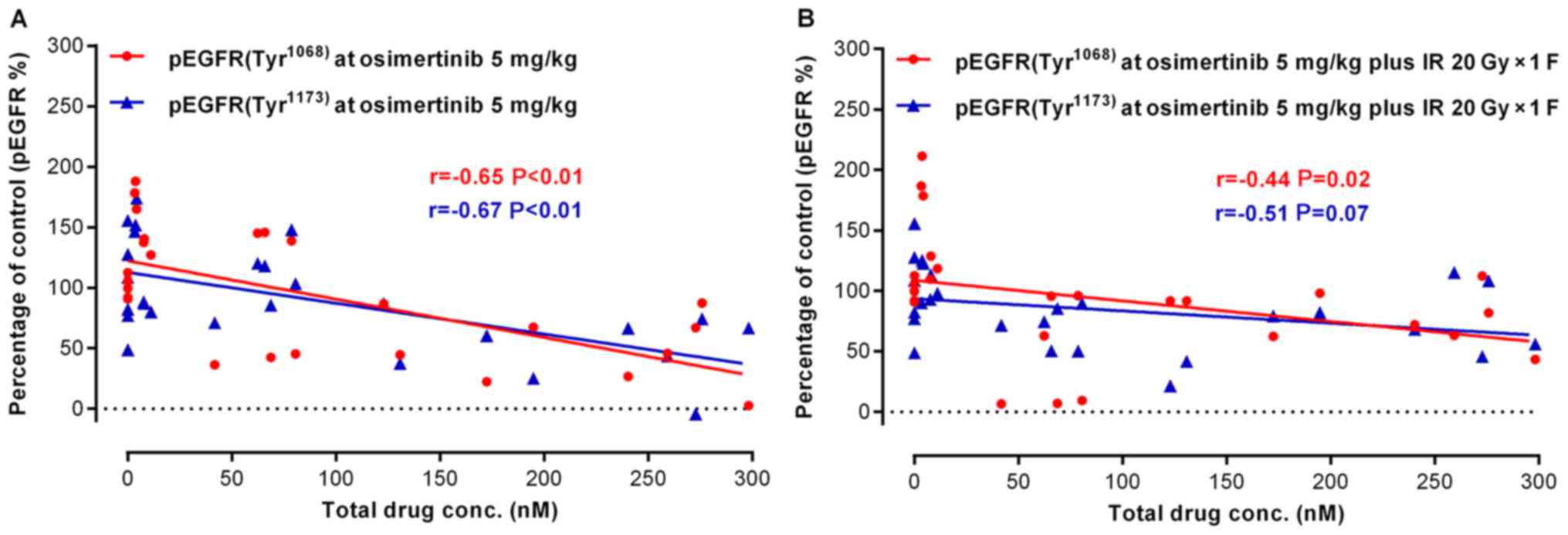 | Figure 6.Association between drug metabolism
and pharmacokinetics in the NCI-H1975 ×enograft model. (A and B)
The association between osimertinib concentration in plasma and
p-EGFR (Tyr1068)/p-EGFR (Tyr1173) levels in tumor tissues, for (A)
osimertinib alone or (B) in combination with IR. (C) The
representative immunohistochemical images of p-EGFR (Tyr1068),
p-EGFR (Tyr1173), γH2AX and CC3. Scale bars, 100 µm. (D-G)
Quantitative analysis of the expression levels of (D) p-EGFR
(Tyr1068), (E) p-EGFR (Tyr1173), (F) γH2AX and (G) CC3,
respectively. *P<0.05, as indicated. p-, phosphorylated; EGFR,
epidermal growth factor receptor; γH2AX, H2A histone family member
X; CC3, cleaved caspase-3. |
Discussion
Osimertinib is an irreversible third generation
EGFR-TKI that has demonstrated significant potency in patients with
NSCLC with EGFR-sensitizing mutations and the T790M-resistance
mutation (10). First generation
EGFR-TKI therapy coupled with radiotherapy was revealed to have the
potential to improve outcomes for patients with NSCLC exhibiting
EGFR-sensitive mutations (9,23–25).
However, the role of osimertinib in the effect of IR remains
elusive. In the present study, it was demonstrated that osimertinib
combined with IR could significantly decrease the proliferation of
NSCLC cells harboring the T790M/L858R mutation in vitro and
in vivo, reduce G2/M-phase cell cycle arrest and block
IR-induced DNA DSB repair, demonstrating its role in
radio-sensitivity. This may provide a rationale for clinically
combining osimertinib with IR to treat patients with NSCLC
exhibiting EGFR mutations.
Synergistic inhibition of cell proliferation has
been reported to be the predominant mechanism underlying the
effectiveness of the combination treatment of IR and gefitinib
(26,27). In the present study, proliferation
and clone formation were significantly inhibited when treated with
osimertinib and IR. This antitumor effect was further confirmed in
xenograft models. Following the completion of treatment, inhibition
of tumor growth was observed for an additional 25 days. This may be
indicative of the durable cellular suppression of osimertinib when
combined with radiation. However, the optimal combination pattern
of osimertinib and IR, such as the fractioned dose, the fractioned
number, the timing of osimertinib administration and so on, has not
been clear until now. It has been reported that the
autophosphorylation of EGFR could be activated following radiation
and serves an important role in leading to radiation resistance
(28,29). Thus, osimertinib was administrated
prior to radiation in the present study. Notably, cell
proliferation was inhibited by treatment with osimertinib combined
with radiation with the SER >1, which demonstrated the
inhibition of proliferation was at least partly associated with the
radio-sensitivity induced by osimertinib. However, the underlying
mechanisms still require further investigations.
Radio-sensitization is a consequence of the repair
of DNA DSBs mediated through a blockade in EGFR-signaling events
(30,31). It has been reported that irradiation
can directly activate EGFR signaling to in turn activate DNA DSB
repair (28–29,32). A
number of processes in the DNA damage response are central to
radio-sensitivity, including checkpoint activation and repair
(33,34). G2/M-phase-arrest allows cells to
repair damaged DNA and can cause IR-resistance. γ-H2AX is thought
to be a reporter of tumor radio-sensitivity, induced by IR at DNA
DSB sites (35,36). In the present study it was revealed
that osimertinib decreased IR-induced G2/M-phase arrest in a
dose-dependent manner, and that the level of γ-H2AX in the
combination treatment group was increased when compared with that
of IR alone. These results may suggest that, when combined with
osimertinib, IR shortens G2/M-phase arrest and inhibition of DNA
damage repair.
It has been reported that receptor tyrosine kinases
and AKT signaling may be activated by IR in NSCLC cells (37–39).
AKT is involved in the regulation of cell cycle progression. Third
generation agents can induce a switch to multiple signaling
mechanisms such as the ERK and AKT signaling pathways, thereby
bypassing EGFR-dependency (40–42).
In the present study, the slight increase in p-EGFR (1068), p-EGFR
(1173) and p-AKT following irradiation was observed, but not for
pERK. Notably, osimertinib treatment significantly inhibited
p-EGFR, p-AKT and p-ERK protein expression alone or in combination
with IR in vitro; it also significantly decreased the
phosphorylation of EGFR in vivo. Furthermore, the
combination treatment was revealed to increase the rate of
apoptosis. This may be another reason for the radio-sensitization
role of osimertinib in NSCLC cells exhibiting EGFR mutations.
Further investigations are required to determine whether
alterations of a specific gene or gene set involved in EGFR
expression and apoptosis-associated signaling pathways are
modulated by osimertinib.
In conclusion, osimertinib was revealed to enhance
the radio-sensitivity of T790M/L858R NSCLC in vitro and
in vivo, suggesting a potential clinical impact for the use
of this combination therapeutic strategy. Improved understanding of
the molecular mechanisms underpinning the radio-sensitizing effects
of osimertinib may enable the development of novel approaches to
optimize the treatment of lung cancer.
Acknowledgements
The authors would like to thank Mrs. Minghui Hu
(Asia Innovative Medicines and Early Development, AstraZeneca,
Shanghai, China) for cell culture assistance, and Mr. Kunji Liu
(Asia Innovative Medicines and Early Development, AstraZeneca) for
his work on the in vivo modeling. They would also like to
thank Mrs. Haihua Hu (Asia Innovative Medicines and Early
Development, AstraZeneca) for her work on the in vitro
immunofluorescence assay.
Funding
The present study was supported by grants from the
National Health and Family Planning Commission of China (grant no.
201402011), the National Natural Science Foundation of China (grant
no. 81472812), the Postdoctoral Innovation of Shandong Province
(grant no. 201501010), the Postdoctoral Science Foundation of China
(grant no. 2016M590640) and the Innovation Project of the Shandong
Academy of Medical Science.
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
LW, LX, ZY, LZheng and JY designed the study. NW,
JW, LZhu, CL, XM and SL performed the experiments and analyzed the
data. NW drafted the manuscript. LW, XM and LX revised the
manuscript. All authors reviewed and approved the final
manuscript.
Ethics approval and consent to
participate
All of the animal studies were approved by the
Institutional Animal Care and Use Committee of AstraZeneca
(Shanghai, China).
Patient consent for publication
Not applicable.
Competing interests
The authors Mrs. Jia Wang, Mrs. Lifang Zhu, Mrs.
Changting Liu, Mrs. Shaorong Li, Mrs. Li Zheng and Mrs. Zhenfan
Yang are affiliated with Asia Innovative Medicines and Early
Development, AstraZeneca (Shanghai, China). The osimertinib
(AZD9291) used in the present study was provided by
AstraZeneca.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reinersman JM, Johnson ML, Riely GJ,
Chitale DA, Nicastri AD, Soff GA, Schwartz AG, Sima CS, Ayalew G,
Lau C, et al: Frequency of EGFR and KRAS mutations in lung
adenocarcinomas in African Americans. J Thorac Oncol. 6:28–31.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gahr S, Stoehr R, Geissinger E, Ficker JH,
Brueckl WM, Gschwendtner A, Gattenloehner S, Fuchs FS, Schulz C,
Rieker RJ, et al: EGFR mutational status in a large series of
Caucasian European NSCLC patients: Data from daily practice. Br J
Cancer. 109:1821–1828. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shi Y, Au JS, Thongprasert S, Srinivasan
S, Tsai CM, Khoa MT, Heeroma K, Itoh Y, Cornelio G and Yang PC: A
prospective, molecular epidemiology study of EGFR mutations in
Asian patients with advanced non-small-cell lung cancer of
adenocarcinoma histology (PIONEER). J Thorac Oncol. 9:154–162.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
NC CN NCCN Clinical Practice Guidelines in
Oncology NSCLC (version 7 2015). 2015, http://www.nccn.org/professionals/physician_gls/pdf/nscl.pdfJuly
16–2015
|
|
6
|
Tan DS, Yom SS, Tsao MS, Pass HI, Kelly K,
Peled N, Yung RC, Wistuba II, Yatabe Y, Unger M, et al: The
International Association for the Study of Lung Cancer consensus
statement on optimizing management of EGFR mutation-positive
non-small cell lung cancer: Status in 2016. J Thorac Oncol.
11:946–963. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu HA, Arcila ME, Rekhtman N, Sima CS,
Zakowski MF, Pao W, Kris MG, Miller VA, Ladanyi M and Riely GJ:
Analysis of tumor specimens at the time of acquired resistance to
EGFR-TKI therapy in 155 patients with EGFR-mutant lung cancers.
Clin Cancer Res. 19:2240–2247. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ku BM, Bae YH, Koh J, Sun JM, Lee SH, Ahn
JS, Park K and Ahn MJ: AZD9291 overcomes T790 M-mediated resistance
through degradation of EGFR(L858R/T790M) in non-small cell lung
cancer cells. Invest New Drugs. 34:407–415. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Oxnard GR, Arcila ME, Sima CS, Riely GJ,
Chmielecki J, Kris MG, Pao W, Ladanyi M and Miller VA: Acquired
resistance to EGFR tyrosine kinase inhibitors in EGFR-mutant lung
cancer: Distinct natural history of patients with tumors harboring
the T790M mutation. Clin Cancer Res. 17:1616–1622. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cross DA, Ashton SE, Ghiorghiu S, Eberlein
C, Nebhan CA, Spitzler PJ, Orme JP, Finlay MR, Ward RA, Mellor MJ,
et al: AZD9291, an irreversible EGFR TKI, overcomes T790M-mediated
resistance to EGFR inhibitors in lung cancer. Cancer Discov.
4:1046–1061. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Finlay MR, Anderton M, Ashton S, Ballard
P, Bethel PA, Box MR, Bradbury RH, Brown SJ, Butterworth S,
Campbell A, et al: Discovery of a potent and selective EGFR
inhibitor (AZD9291) of both sensitizing and T790M resistance
mutations that spares the wild type form of the receptor. J Med
Chem. 57:8249–8267. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
FDA FDA approves new pill to treat certain
patients with non-small cell lung cancer, 2015. Available at
Osimertinib in models of EGFR-mutant NSCLC brain metastases.
http://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm472525.htmNovember
13–2015
|
|
13
|
Ramalingam SS, Reungwetwattana T and
Chewaskulyong B: Osimertinib versus standard-of-care EGFR-TKI as
first-line treatment in patients with EGFRm advanced NSCLC: FLARA.
Presented at the ESMO Congress, Madrid, (abstract LBA2). 2017,
https://oncologypro.esmo.org/Meeting-Resources/ESMO-2017-Congress/Osimertinib-vs-standard-of-care-SoC-EGFR-TKI-as-first-line-therapy-in-patients-pts-with-EGFRm-advanced-NSCLC-FLAURA
|
|
14
|
Delaney G, Barton M, Jacob S and Jalaludin
B: A model for decision making for the use of radiotherapy in lung
cancer. Lancet Oncol. 4:120–128. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bokobza SM, Jiang Y, Weber AM, Devery AM
and Ryan AJ: Short-course treatment with gefitinib enhances
curative potential of radiation therapy in a mouse model of human
non-small cell lung cancer. Int J Radiat Oncol Biol Phys.
88:947–954. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang S, Zheng X, Huang H, Wu K, Wang B,
Chen X and Ma S: Afatinib increases sensitivity to radiation in
non-small cell lung cancer cells with acquired EGFR T790M mutation.
Oncotarget. 6:5832–5845. 2015.PubMed/NCBI
|
|
17
|
Kriegs M, Gurtner K, Can Y, Brammer I,
Rieckmann T, Oertel R, Wysocki M, Dorniok F, Gal A, Grob TJ, et al:
Radiosensitization of NSCLC cells by EGFR inhibition is the result
of an enhanced p53-dependent G1 arrest. Radiother Oncol.
115:120–127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ready N, Jänne PA, Bogart J, Dipetrillo T,
Garst J, Graziano S, Gu L, Wang X, Green MR and Vokes EE: Cancer,
Leukemia Group B, Chicago, IL: Chemoradiotherapy and gefitinib in
stage III non-small cell lung cancer with epidermal growth factor
receptor and KRAS mutation analysis: Cancer and leukemia group B
(CALEB) 30106, a CALGB-stratified phase II trial. J Thorac Oncol.
5:1382–1390. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kelly K, Chansky K, Gaspar LE, Albain KS,
Jett J, Ung YC, Lau DH, Crowley JJ and Gandara DR: Phase III trial
of maintenance gefitinib or placebo after concurrent
chemoradiotherapy and docetaxel consolidation in inoperable stage
III non-small-cell lung cancer: SWOG S0023. J Clin Oncol.
26:2450–2456. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Choong NW, Mauer AM, Haraf DJ, Lester E,
Hoffman PC, Kozloff M, Lin S, Dancey JE, Szeto L, Grushko T, et al:
Phase I trial of erlotinib-based multimodality therapy for
inoperable stage III non-small cell lung cancer. J Thorac Oncol.
3:1003–1011. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Komaki R, Blumenschein GR, Wistuba II, Lee
JJ, Allen P, Wei X, Welsh J, O'Reilly M, Herbst RS, Tang X, et al:
Phase II trial of erlotinib and radiotherapy following
chemoradiotherapy for patients with stage III non-small cell lung
cancer. J Clin Oncol. 29((Suppl 15)): 70202011. View Article : Google Scholar
|
|
22
|
Kalvass JC, Maurer TS and Pollack GM: Use
of plasma and brain unbound fractions to assess the extent of brain
distribution of 34 drugs: Comparison of unbound concentration
ratios to in vivo p-glycoprotein efflux ratios. Drug Metab Dispos.
35:660–666. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Casal Rubio J, Fírvida-Pérez JL,
Lázaro-Quintela M, Barón-Duarte FJ, Alonso-Jáudenes G, Santomé L,
Afonso-Afonso FJ, Amenedo M, Huidobro G, Campos-Balea B, et al: A
phase II trial of erlotinib as maintenance treatment after
concurrent chemoradiotherapy in stage III non-small-cell lung
cancer (NSCLC): A Galician Lung Cancer Group (GGCP) study. Cancer
Chemother Pharmacol. 73:451–457. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sequist LV, Martins RG, Spigel D, Grunberg
SM, Spira A, Jänne PA, Joshi VA, McCollum D, Evans TL, Muzikansky
A, et al: First-line gefitinib in patients with advanced
non-small-cell lung cancer harboring somatic EGFR mutations. J Clin
Oncol. 26:2442–2449. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xu Y, Zheng Y, Sun X, Yu X, Gu J, Wu W,
Zhang G, Hu J, Sun W and Mao W: Concurrent radiotherapy with
gefitinib in elderly patients with esophageal squamous cell
carcinoma: Preliminary results of a phase II study. Oncotarget.
6:38429–38439. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Park SY, Kim YM and Pyo H: Gefitinib
radiosensitizes non-small cell lung cancer cells through inhibition
of ataxia telangiectasia mutated. Mol Cancer. 9:2222010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Das AK, Sato M, Story MD, Peyton M, Graves
R, Redpath S, Girard L, Gazdar AF, Shay JW, Minna JD, et al:
Non-small-cell lung cancers with kinase domain mutations in the
epidermal growth factor receptor are sensitive to ionizing
radiation. Cancer Res. 66:9601–9608. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Das AK, Chen BP, Story MD, Sato M, Minna
JD, Chen DJ and Nirodi CS: Somatic mutations in the tyrosine kinase
domain of epidermal growth factor receptor (EGFR) abrogate
EGFR-mediated radioprotection in non-small cell lung carcinoma.
Cancer Res. 67:5267–5274. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Dittmann K, Mayer C, Fehrenbacher B,
Schaller M, Kehlbach R and Rodemann HP: Nuclear EGFR shuttling
induced by ionizing radiation is regulated by phosphorylation at
residue Thr654. FEBS Lett. 584:3878–3884. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dittmann K, Mayer C, Fehrenbacher B,
Schaller M, Raju U, Milas L, Chen DJ, Kehlbach R and Rodemann HP:
Radiation-induced epidermal growth factor receptor nuclear import
is linked to activation of DNA-dependent protein kinase. J Biol
Chem. 280:31182–31189. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Toulany M, Kasten-Pisula U, Brammer I,
Wang S, Chen J, Dittmann K, Baumann M, Dikomey E and Rodemann HP:
Blockage of epidermal growth factor receptor-phosphatidylinositol
3-kinase-AKT signaling increases radiosensitivity of K-RAS mutated
human tumor cells in vitro by affecting DNA repair. Clin Cancer
Res. 12:4119–4126. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rodemann HP, Dittmann K and Toulany M:
Radiation-induced EGFR-signaling and control of DNA-damage repair.
Int J Radiat Biol. 83:781–791. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kriegs M, Kasten-Pisula U, Rieckmann T,
Holst K, Saker J, Dahm-Daphi J and Dikomey E: The epidermal growth
factor receptor modulates DNA double-strand break repair by
regulating non-homologous end-joining. DNA Repair (Amst).
9:889–897. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Myllynen L, Rieckmann T, Dahm-Daphi J,
Kasten-Pisula U, Petersen C, Dikomey E and Kriegs M: In tumor cells
regulation of DNA double strand break repair through EGF receptor
involves both NHEJ and HR and is independent of p53 and K-Ras
status. Radiother Oncol. 101:147–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Löbrich M, Shibata A, Beucher A, Fisher A,
Ensminger M, Goodarzi AA, Barton O and Jeggo PA: gammaH2AX foci
analysis for monitoring DNA double-strand break repair: Strengths,
limitations and optimization. Cell Cycle. 9:662–669. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lord CJ and Ashworth A: The DNA damage
response and cancer therapy. Nature. 481:287–294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li B, Yuan M, Kim IA, Chang CM, Bernhard
EJ and Shu HK: Mutant epidermal growth factor receptor displays
increased signaling through the phosphatidylinositol-3 kinase/AKT
pathway and promotes radioresistance in cells of astrocytic origin.
Oncogene. 23:4594–4602. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun Y, Moretti L, Giacalone NJ, Schleicher
S, Speirs CK, Carbone DP and Lu B: Inhibition of JAK2 signaling by
TG101209 enhances radiotherapy in lung cancer models. J Thorac
Oncol. 6:699–706. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dent P, Yacoub A, Contessa J, Caron R,
Amorino G, Valerie K, Hagan MP, Grant S and Schmidt-Ullrich R:
Stress and radiation-induced activation of multiple intracellular
signaling pathways. Radiat Res. 159:283–300. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Walter AO, Sjin RT, Haringsma HJ, Ohashi
K, Sun J, Lee K, Dubrovskiy A, Labenski M, Zhu Z, Wang Z, et al:
Discovery of a mutant-selective covalent inhibitor of EGFR that
overcomes T790M-mediated resistance in NSCLC. Cancer Discov.
3:1404–1415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ercan D, Xu C, Yanagita M, Monast CS,
Pratilas CA, Montero J, Butaney M, Shimamura T, Sholl L, Ivanova
EV, et al: Reactivation of ERK signaling causes resistance to EGFR
kinase inhibitors. Cancer Discov. 2:934–947. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Cortot AB, Repellin CE, Shimamura T,
Capelletti M, Zejnullahu K, Ercan D, Christensen JG, Wong KK, Gray
NS and Jänne PA: Resistance to irreversible EGF receptor tyrosine
kinase inhibitors through a multistep mechanism involving the IGF1R
pathway. Cancer Res. 73:834–843. 2013. View Article : Google Scholar : PubMed/NCBI
|