Introduction
Cutaneous squamous cell carcinoma (cSCC) is one of
the primary types of skin cancer characterized by the malignant
proliferation of keratinizing cells of the epidermis (1). Worldwide, cSCC is the second most common
non-melanoma skin carcinoma (NMSC), following basal cell carcinoma
(BCC) (2). Despite early stage
diagnosis in most cases, cSCC tumors account for the majority of
NMSC-associated mortalities (3), and
~5% of cSCC tumors metastasize to regional lymph nodes or distant
sites (4). Although there has been
considerable progress in understanding the various etiologies of
cSCC, the molecular mechanisms of invasiveness have not been
clarified.
Nicotinamide N-methyltransferase (NNMT) is an
S-adenosyl-L-methionine-dependent cytoplasmic enzyme that
catalyzes the N-methylation of nicotinamide, pyridines and
their structural analogs (5,6). NNMT is predominantly expressed in the
liver, where its activity leads to the metabolism of drugs and
xenobiotic compounds (5,6). Elevated levels of NNMT expression have
been identified in various tumors, suggesting that NNMT serves an
important role in the progression of these cancer types (7–16).
Moreover, increased NNMT expression levels have been detected in
bodily fluids including the serum (14,17),
saliva (18) and urine (19). These findings suggest that NNMT may
act as a potential non-invasive cancer biomarker. Previous studies
have also indicated that NNMT dysregulation may affect the
proliferative, migratory, invasive and differentiation profiles of
various cancer types (16,20,21).
Notably, the upregulation of NNMT in cancer has been linked to poor
prognosis and treatment resistance (10,15,22,23).
However, the expression profile and biological functions of NNMT in
cSCC are yet to be elucidated.
Enhanced metastasis and rapid tumor growth are
associated with poor prognosis in cSCC (24,25).
Epithelial-mesenchymal transition (EMT) is a transcriptional
process that is activated in embryonic development and wound
healing, and contributes to cancer initiation and progression,
particularly during metastasis (26,27).
During EMT, epithelial cells lose their polarity and intercellular
adhesion properties, the cytoskeleton is reorganized, and
mesenchymal characteristics are gained, including migratory
activity. Invasion and metastasis are among the principal
characteristics of cancer (28), and
cancerous cells routinely acquire an invasive phenotype following
EMT. Various signaling pathways have been found to regulate EMT,
enhancing cell motility and invasiveness (29). Considering that it is necessary for
skin healing and can also contribute to tumor invasion and
metastasis, an understanding of the underlying mechanisms of EMT is
crucial.
The present study aimed to identify differentially
expressed proteins in an invasive, compared with those in a
non-invasive cSCC cell line, and to elucidate the mechanism of
invasiveness associated with specific genes, particularly NNMT.
NNMT was revealed to be upregulated in SCC12 cells, and
NNMT-targeted shRNA suppressed cell growth and invasion via the
downregulation of EMT-related genes.
Materials and methods
Cell culture
The human skin squamous carcinoma cell lines SCC12
and SCC13 were kindly donated by Dr James Rheinwald (Brigham and
Women's Hospital, Harvard Medical School, Boston, MA, USA). The
cells were cultured in growth medium containing a 3:1 ratio of
Dulbecco's modified Eagle's medium (DMEM) and Ham's F-12 Nutrient
Mixture (Gibco; Thermo Fisher Scientific, Inc.), supplemented with
10% fetal bovine serum (FBS), 5 µg/ml insulin, 0.5 µg/ml
hydrocortisone and 10 ng/ml epidermal growth factor. The cells were
maintained at 37°C (5% CO2) in a humidified
incubator.
Protein extraction and two-dimensional
gel electrophoresis (2-DE)
Cells were lysed on ice in lysis buffer (8 M urea,
4% CHAPS, 40 mM Tris-base and 100 mM dithiothreitol) supplemented
with a protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA). The
lysate was centrifuged at 13,000 × g for 10 min at 4°C to remove
insoluble cell debris, followed by trichloroacetic acid
precipitation. The precipitated proteins were resuspended in lysis
buffer containing 2% (w/v) ampholyte and quantified using a
Bradford protein assay kit (Bio-Rad Laboratories, Inc.). Equal
amounts of protein (30 µg) were subjected to first-dimension
isoelectric focusing (pH 4.0–7.0) and second-dimension gradient
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE; 7.5–17.5%).
Gel visualization and analysis
After 2-DE, the gels were fixed in methanol:acetic
acid:water (50:12:38) for 1.5 h, pretreated with 0.02%
Na2S2O3 for 1 min and washed twice
with deionized water. The gels were soaked in a solution containing
0.2% silver nitrate and 0.075% (v/v) formaldehyde, and incubated in
a developing solution [2% sodium bicarbonate, 0.0004% sodium
sulfoxide and 0.06% (v/v) formaldehyde]. The silver-stained gels
were scanned with a high-resolution scanner (GS-800 Calibrated
Imaging Densitometer; Bio-Rad Laboratories, Inc.) and all images
were analyzed using PDQuest™ software (version 8.0; Bio-Rad
Laboratories, Inc.).
Protein identification using
matrix-assisted laser desorption/ionization time-of-flight mass
spectrometry (MALDI-TOF-MS)
Protein spots of interest were excised from the 2-DE
gels and digested with trypsin (Promega Corp.). Subsequently, the
peptides were extracted using ammonium hydrogen carbonate buffer
with 5% formic acid. The extracted peptides were vacuum-dried and
reconstituted in a solution containing water, acetonitrile and
trifluoroacetic acid (93:5:2 by volume). The protein digest
spectrum was acquired in reflector mode and positive ion mode using
the Voyager-DE STR MALDI-TOF mass spectrometer (Applied Biosystems;
Thermo Fisher Scientific, Inc.). The following settings were used:
acceleration voltage 20 kV, guide wire voltage 0.01%, grid voltage
76% and delay time 150 nsec. Protein identification was performed
using the search program MS-FIT (http://prospector.ucsf.edu/prospector/mshome.htm).
Short hairpin RNA (shRNA)-mediated
NNMT gene silencing
The shRNA target sequence for NNMT
(5′-CATGATTGGTGAGCAGAAG-3′) was cloned into the pENTR™/U6 plasmid
(Invitrogen; Thermo Fisher Scientific, Inc.). The cloned pENTR-NNMT
plasmid was used to generate an adenovirus backbone by homologous
recombination with pAd/BLOCK-iT™-DEST. The target construct,
subjected to antibiotic-resistance screening and PCR confirmation,
was termed pAd-shNNMT, and the adenoviral plasmid containing the
GFP sequence was termed pAd-GFP. After endonuclease digestion with
PacI, linearized pAd-shNNMT and pAd-GFP were transfected
into 293A cells using Lipofectamine® 3000. Adenovirus
was collected from the culture supernatants and showed a cytopathic
effect of >80%. Adenovirus amplification was performed by
infecting 293A cells with the harvested adenoviral stocks. To
confirm NNMT gene-silencing, SCC12 cells were seeded into 6-well
plates (1×105 cells per well). At 70–80% confluence (24
h), the cells were infected with Ad-shNNMT or Ad-GFP adenovirus at
a multiplicity of infection (MOI) of 20, and harvested 72 h
post-infection for protein extraction and immunoblotting.
Cell viability
The effect of NNMT knockdown on the viability of
SCC12 cells was determined using the RealTime-Glo™ MT Cell
Viability Assay (Promega Corp.) in an opaque-walled assay plate,
according to the manufacturer's instructions but with minor
modifications. Briefly, SCC12 cells were seeded into 6-well plates
(1×105 cells per well) and incubated for 24 h at 37°C.
The cells were then infected with Ad-GFP or Ad-shNNMT adenovirus at
an MOI of 20. At 24 h post-infection, the growth medium was
replaced with the aforementioned normal culture medium. After an
additional 48 h incubation, the infected cells were detached with
trypsin/EDTA and replated in 96-well plates (5×102 cells
per well), and viability was assessed using the RealTime-Glo™ MT
Cell Viability Assay (Promega Corp.). To continuously monitor
viability, the pro-substrate and luciferase were added at the time
of cell replating. The luminescence intensity at the desired time
points was measured using the Glomax Multi Detection System
(Promega Corp.), and each sample was analyzed at least in
triplicate.
Colony formation assay
Ad-GFP and Ad-shNNMT adenovirus-infected SCC12 cells
were seeded into 6-well plates in triplicate (5×102
cells per well). The culture plates were incubated at 37°C for 7
days in the aforementioned growth medium, which was replaced every
3 days. The resulting colonies were stained with crystal violet
(1%, w/v; Sigma-Aldrich; Merck KGaA) for 2 h at room temperature
and counted under a microscope (Eclipse 80Ti, Nikon
Corporation).
Gap closure assay
Migration assays were performed using wound assay
chambers (Ibidi GmbH). Ad-GFP and Ad-shNNMT adenovirus-infected
SCC12 cells were trypsinized and resuspended in growth medium
containing 10% FBS. The cells were seeded (4×104 cells
per well) in the insert and allowed to adhere overnight. On the
following day, the inserts were removed and light microscopy images
were acquired (three for each sample). The cells were then cultured
in serum-free cuture medium for the indicated time, and images were
subsequently acquired using an inverted microscope equipped with an
image capture system (Nikon Corporation). The images were analyzed
using automated image analysis software (Nikon NIS Elements version
5.1 software; NIS-Elements Advanced Research).
Invasion assay
Cells infected with Ad-GFP or Ad-shNNMT adenovirus
(1×105 cells per well) were seeded into the upper
chamber of Matrigel-coated (BD Biosciences) Transwell inserts with
8-µm pore polycarbonate filters (Costar; Corning, Inc.). The lower
chamber was filled with DMEM/F12 containing 10% FBS, and the
inserts were incubated at 37°C for 24 h. Invaded cells were fixed
in 4% paraformaldehyde and stained with DAPI. The cells were
counted in five random fields and images were obtained using a
fluorescence microscope (magnification, ×100).
Western blot analysis
Cell extracts were prepared using RIPA lysis buffer
(Santa Cruz Biotechnology, Inc.) supplemented with protease and
phosphatase inhibitor cocktails (Thermo Fisher Scientific, Inc.).
The cells were sonicated for 2 min and centrifuged at 12,000 × g
for 10 min at 4°C to remove insoluble cell debris, and the protein
concentrations were determined using a bichinconinic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). A total of 30
µg protein per well were separated by 10% SDS-PAGE, and transferred
to a nitrocellulose membrane (EMD Millipore). After blocking and
washing following a standard protocol, the target proteins were
detected by incubation with primary antibodies against NNMT
(dilution 1:1,000; cat. no. AP13775C; Abgent, Inc.), E-cadherin
(dilution 1:1,000; cat. no. sc-21791; Santa Cruz Biotechnology,
Inc.), vimentin (dilution 1:1,000; cat. no. sc-6601; Santa Cruz
Biotechnology), zinc finger E-box-binding homeobox 1 (ZEB1;
dilution 1:1,000; cat. no. NBP1-05987; Novus biologicals, Ltd.),
Snail (dilution 1:1,000; cat. no. NBP1-80022; Novus Biologicals,
Ltd.), Slug (dilution 1:1,000; cat. no. ab128485; Abcam), MMP9
(dilution 1:1,000; cat. no. ab104686; Abcam), SPP1 (dilution 1:500;
cat. no. sc-73631; Santa Cruz Biotechnology, Inc.) and β-actin
(dilution 1:5,000; cat. no. A5441; Sigma-Aldrich; Merck KGaA).
Protein bands were visualized using an enhanced chemiluminescence
detection reagent (Pierce; Thermo Fisher Scientific, Inc.) and
images were captured using the ChemiDoc Touch Imaging System
(Bio-Rad Laboratories, Inc.).
Reverse-transcription-quantitative
(RT-q)PCR array
The RNA expression levels of 84 EMT-related genes
were simultaneously analyzed using the RT2 Profiler PCR
Array (cat. no. PAHS-090ZA; Qiagen, Inc.). Total RNA was purified
from SCC cells using the RNeasy kit (Qiagen, Inc.) according to the
manufacturer's protocol, and residual genomic DNA was removed using
the RNase-Free DNase Set (Qiagen, Inc.). Total RNA (5 µg) was
reversed transcribed into cDNA in a 10-µl reaction volume using the
RT2 First Strand Kit (Qiagen, Inc.). The cDNA was
diluted with RT2 SYBR® Green Mastermix
(Qiagen, Inc.) and distilled water to a total volume of 4,200 µl.
Then, 10 µl cDNA solution was used for each primer set in the
pathway-specific RT2 Profiler PCR Array, according to
the manufacturer's protocol. Amplification reactions were performed
using the Applied Biosystems ViiA7 Real-Time PCR System with
automated baseline and threshold cycle detection. The threshold
cycle number for each group of 84 genes was normalized to four
built-in housekeeping gene controls using the RT2
Profiler PCR Array Data Analysis web-based software (SABiosciences
Corp.).
Immunohistochemistry
Nine human skin cancer samples were collected at the
time of surgical resection at the Gyeongsang National University
Hospital (Jinju, Republic of Korea) during the period 2009–2015.
There were 5 males and 4 females, with a median age of 70.5 (range,
47–89 years). A 5-µm section was cut from the formalin-fixed,
paraffin-embedded tissue blocks of 9 patients with skin cancer.
Each section was mounted on charged slides, deparaffinized and
rehydrated in a graded alcohol series. To block endogenous
peroxidase activity, each slide was incubated with 0.3% hydrogen
peroxide for 30 min at room temperature. Antigen retrieval was
conducted by boiling in 10 mM sodium citrate for 30 min, and
immunostaining was performed using a rabbit polyclonal NNMT
antibody (dilution 1:200) with a labeled streptavidin-biotin kit
(Dako; Agilent Technologies, Inc.) according to the manufacturer's
instructions. The slides were lightly counterstained with Mayer's
hematoxylin and mounted.
RT-qPCR
Total RNA was extracted from SCC cells using Trizol
reagent (Thermo Fisher Scientific, Inc.), and first-strand cDNA was
generated using random hexamer primers provided in a first-strand
cDNA synthesis kit (Bio-Rad Laboratories, Inc.), according to the
manufacturer's instructions. The primers and probes (GAPDH; cat.
no. Hs02758991-g1; NNMT; cat. no. Hs00196287) were obtained
commercially (TaqMan® Gene Expression Assay; Thermo
Fisher Scientific, Inc.) and the cDNA was amplified using the
TaqMan® Gene Expression Master Mix (Thermo Fisher
Scientific, Inc.). Amplification was performed in 96-well plates
using the ViiA™ 7 Real-Time PCR System (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The thermocycling conditions were as
follows: 50°C for 2 min and 95°C for 10 min, followed by 40 cycles
at 94°C for 15 sec and 60°C for 1 min. GAPDH was used as the
internal control, and three repetitive experiments were conducted.
The comparative Ct (2−ΔΔCq) method (30) was used to calculate the relative
levels of EMT-related genes.
Statistical analysis
Each experiment was conducted at least three times
independently, and the data are presented as the mean ± standard
deviation. The difference between two groups was assessed using the
two-tailed Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Migration and invasion capacities of
SCC12 cells
Cell migration and invasion are integral to the
metastatic process. To characterize the phenotypic differences
between two cutaneous SCC cell lines, the cell migration and
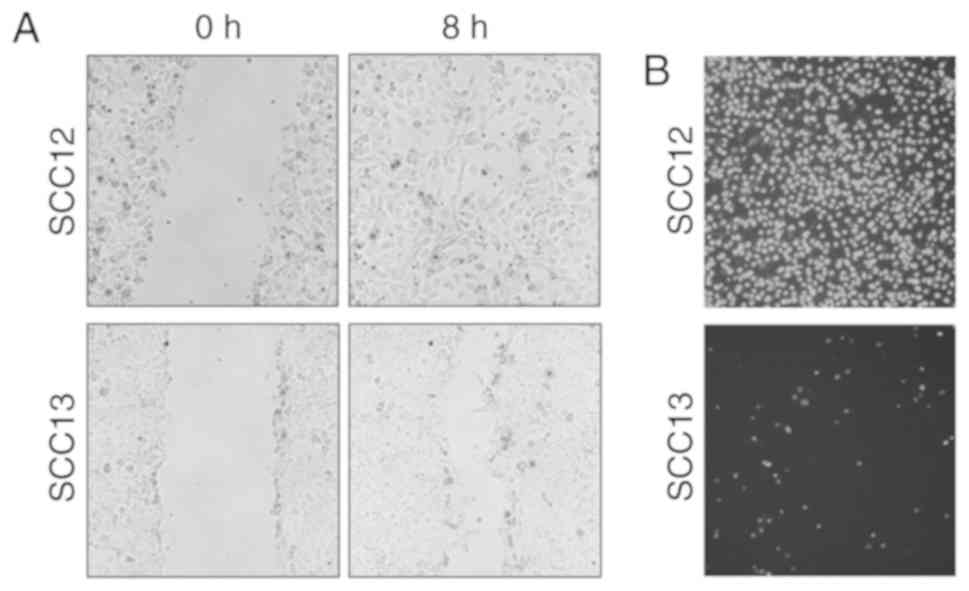
invasion capacities of SCC12 and SCC13 cells were determined using
wound healing and Matrigel invasion assays, respectively. SCC12
cells exhibited a progressive increase in the rate of cell
migration (Fig. 1A), and the
migration ability was considerably greater than that of SCC13
cells. In addition, the invasive ability of SCC12 cells was
increased compared with that of SCC13 cells (Fig. 1B). During cultivation, the phenotype
of cancerous cells is altered. However, the present study revealed
higher motility and invasive potential in SCC12 cells compared with
SCC13 cells. Therefore, these cell lines represent an ideal model
for characterizing genomic and proteomic features that result in a
metastatic phenotype.
Identification of molecular markers
for metastasis-related genes in SCC cells using proteomic
analysis
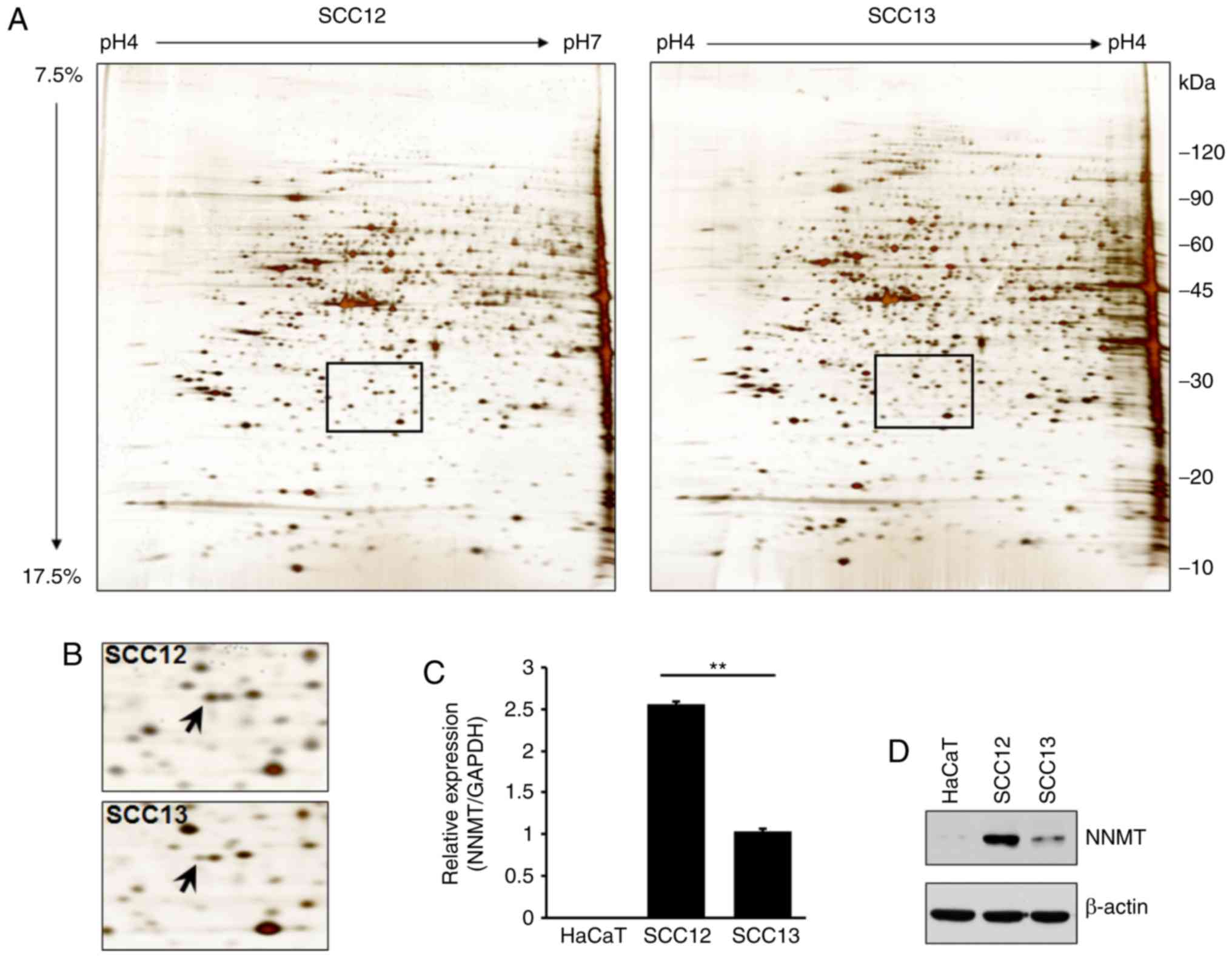
2-DE and LC-MS were used to analyze the differential
expression levels of total proteins in two cSCC cell lines. A
number of proteins were differentially expressed in SCC12 cells
compared with SCC13 cells (Fig. 2A
and Table I). Specifically, a protein
spot identified as NNMT by mass spectrometry was upregulated in
SCC12 cells (Fig. 2B). To verify the
results of 2-DE, RT-qPCR and western blot analyses were also
performed using the total RNA or cell lysates isolated from SCC12
and SCC13 cells, and the keratinocyte HaCaT cell line. Similarly,
the NNMT mRNA expression level in SCC12 cells was 2.5-fold higher
than that in the SCC13 cells (Fig.
2C). The corresponding level of NNMT protein expression was
also higher in SCC12 than in SCC13 cells (Fig. 2D).
 | Table I.Differentially expressed proteins in
SCC12 cellsa. |
Table I.
Differentially expressed proteins in
SCC12 cellsa.
| Spot# | Protein name | Accession no. | MW (kDa)/pI | No. of matched
peptides | Seq. coverage
(%) |
P-valueb |
|---|
| 2010 | Cytokeratin 19 | P08727 | 44.1/5.04 | 14/23 | 32 | 0.0002 |
| 4008 | Cytokeratin 19 | P08727 | 44.1/5.04 | 10/17 | 22 | 0.0036 |
| 4825 | Tropomyosin 1α
chain | P09493 | 32.7/4.69 | 5/13 | 15 | 0.0173 |
| 5008 | HSP27 | O14530 | 26.5/5.61 | 6/15 | 23 | 0.0004 |
| 6112 | Cytokeratin 8 | P05787 | 53.5/5.52 | 6/16 | 19 | 0.0125 |
| 3704 | Cytokeratin 8 | P05787 | 53.5/5.52 | 6/12 | 17 | 0.0081 |
| 5405 | Cytokeratin 8 | P05787 | 53.5/5.52 | 10/18 | 26 | 0.0133 |
| 2010 |
Adenosylhomosysteinase | P23526 | 47.6/5.92 | 8/17 | 18 | 0.0007 |
| 4216 | Nicotinamide
N-methyltransferase (NNMT) | P40261 | 29.6/5.56 | 9/22 | 42 | 0.0002 |
| 2010 | Glutathione
S-transferase omega 1 | P78417 | 27.6/6.24 | 5/17 | 21 | 0.0015 |
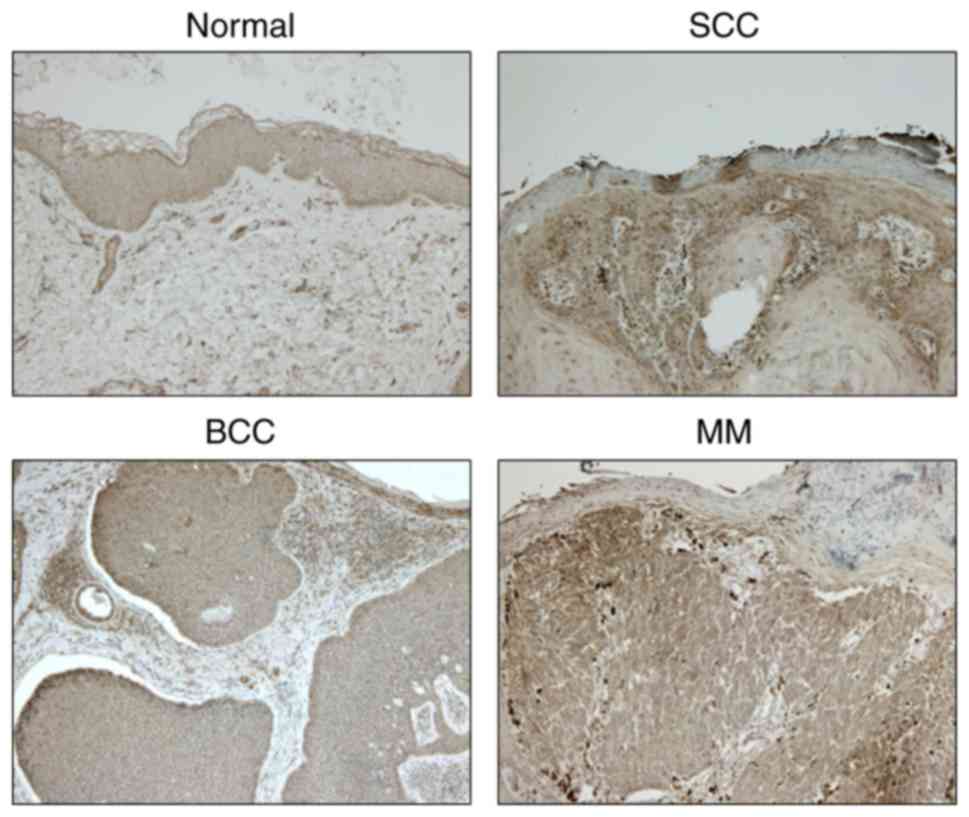
Immunohistochemical examination
The expression levels of NNMT in healthy, SCC, basal
cell carcinoma (BCC) and malignant melanoma (MM) tissues were
examined using immunohistochemistry. NNMT protein was strongly
expressed in SCC and MM tissues, and mildly expressed in BCC
tissues; however, it was not expressed in healthy tissues (Fig. 3).
Evaluation of EMT-related gene
expression in invasive and non-invasive SCC cells
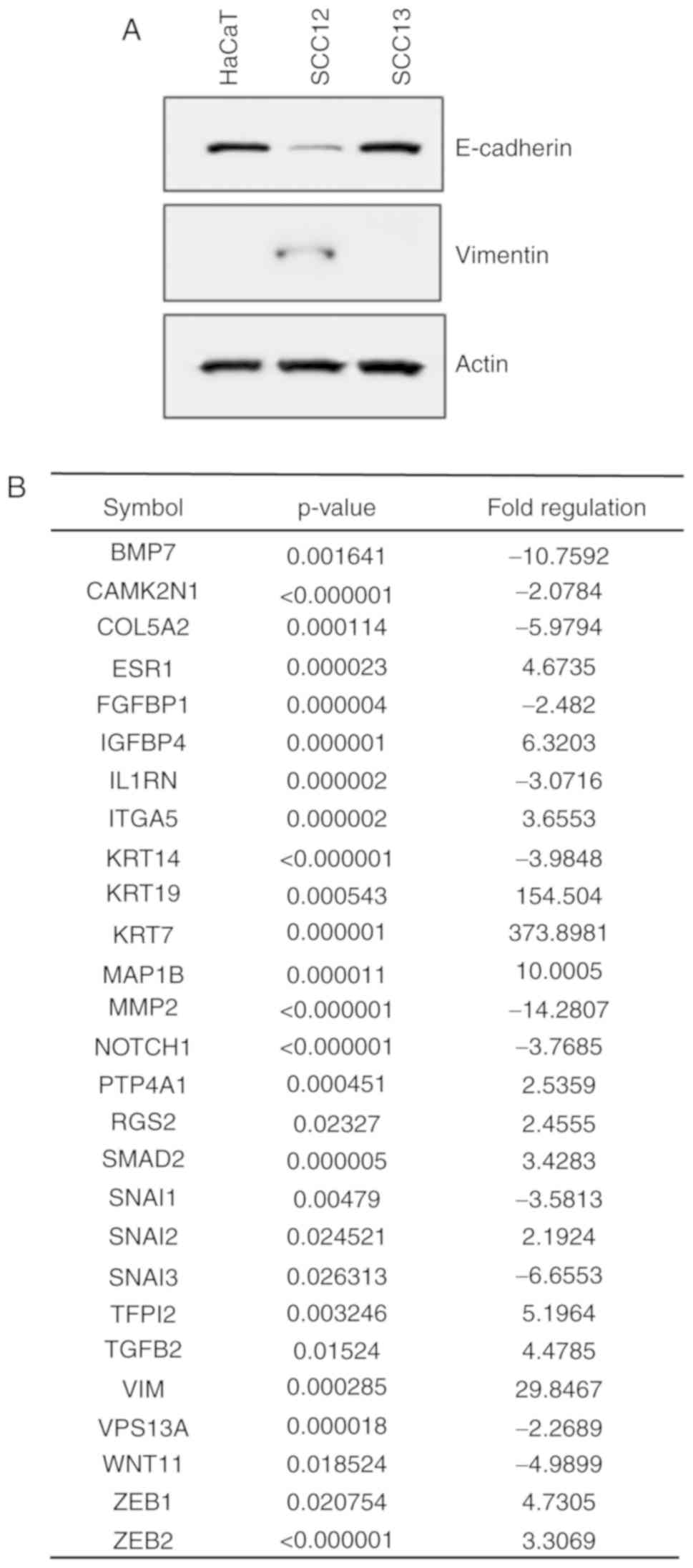
To determine whether NNMT was able to regulate human
SCC metastasis, the expression levels of EMT mediators were
assessed in the HaCaT, SCC12 and SCC13 cell lines using western
blotting. SCC12 cells showed a high expression level of the
mesenchymal marker vimentin, and a low expression level of the
epithelial marker E-cadherin, consistent with a migratory phenotype
(Fig. 4A). To identify genes
associated with the invasive phenotype of SCC12 cells, changes in
the expression of EMT-related genes were determined using the
RT2 Profiler PCR Array for 84 epithelial and
mesenchymal-associated genes. As shown in Fig. 4B, 27 EMT-related genes were
upregulated and suppressed in SCC12 cells with certain genes
exhibiting more than 2-fold changes in expression levels compared
with SCC13 cells. Several genes, including EMT-related vimentin,
cytokeratin 19 (KRT19) (upregulated), cytokeratin 7
(KRT7) (upregulated), bone morphogenetic protein 7
(BMP7) (downregulated) and matrix metalloproteinase 2
(MMP2) (downregulated) were differentially expressed
(Fig. 4B).
Effect of NNMT knockdown on cell
proliferation and density-dependent growth
To investigate the effects of NNMT overexpression on
SCC cells, NNMT knockdown was performed in SCC12 cells using
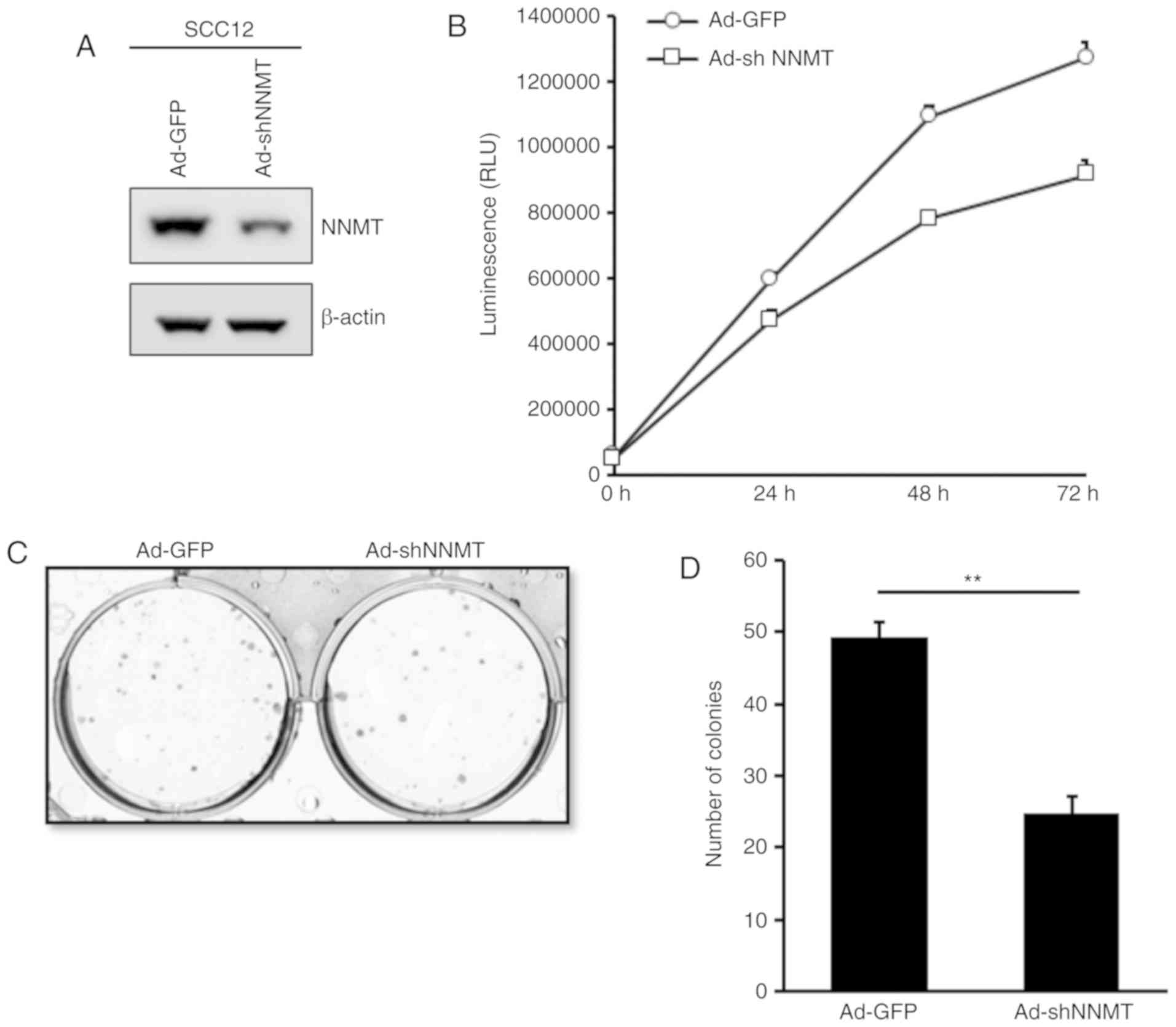
shRNA-expressing adenovirus. The protein expression level of NNMT
was determined and was markedly decreased in the
Ad-shNNMT-infected, compared with the Ad-GFP-infected SCC12 cells
(Fig. 5A). To assess the effects of
NNMT-silencing on the proliferation of SCC12 cells, a
bioluminescence assay was performed using the RealTime-Glo™ MT Cell
Viability Assay. The proliferation of Ad-shNNMT-infected SCC12
cells was considerably impaired compared with that of the
corresponding Ad-GFP-infected control cells (Fig. 5B). These results indicate that NNMT
may promote the proliferation of SCC cells.
Furthermore, the effect of NNMT knockdown on the
anchorage-dependent growth of SCC12 cells was investigated using a
colony formation assay. A similar inhibitory effect was observed;
the mean colony number was 49.3 in the control group and 24 in the
Ad-shNNMT group (Fig. 5C and D),
indicating that NNMT knockdown reduced the colony forming capacity
of SCC12 cells, and suggesting an inhibitory effect on cell
proliferation.
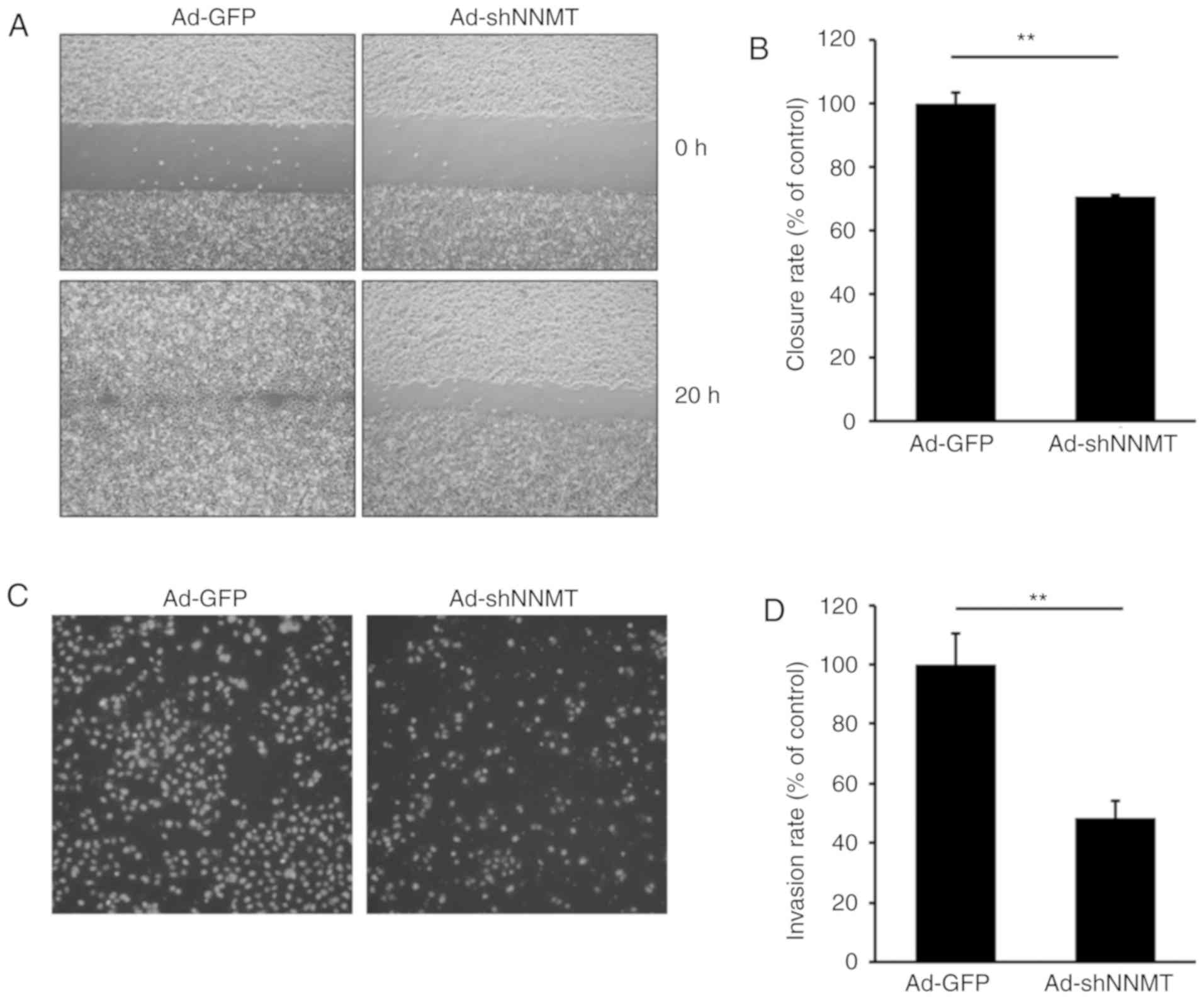
Effect of NNMT knockdown on cell
migration and invasion
An increase in cell motility is known to affect the
metastatic potential of cancer cells. Therefore, the potential
effect of NNMT knockdown on cell motility was investigated in SCC12
cells, a cell line with high metastatic potential. A wound healing
assay was subsequently performed on SCC12 cells infected with
Ad-GFP or Ad-shNNMT adenovirus. Gap closure was predominantly due
to cell migration rather than proliferation, at least at the early
time points. The wound was ~90% closed by the Ad-GFP-infected
cells, though only ~50% closed by Ad-shNNMT-infected cells
(Fig. 6A and B), suggesting that NNMT
knockdown inhibited the motility of SCC12 cells and resulted in
reduced gap closure.
The potential effects of NNMT knockdown on SCC12
cell invasiveness were also investigated. As shown in Fig. 6C and D, the depletion of NNMT using
Ad-shNNMT markedly reduced the invasive ability of SCC12 cells. The
number of invaded knockdown cells was ~47% that of the control
cells, suggesting that NNMT may promote SCC cell invasion.
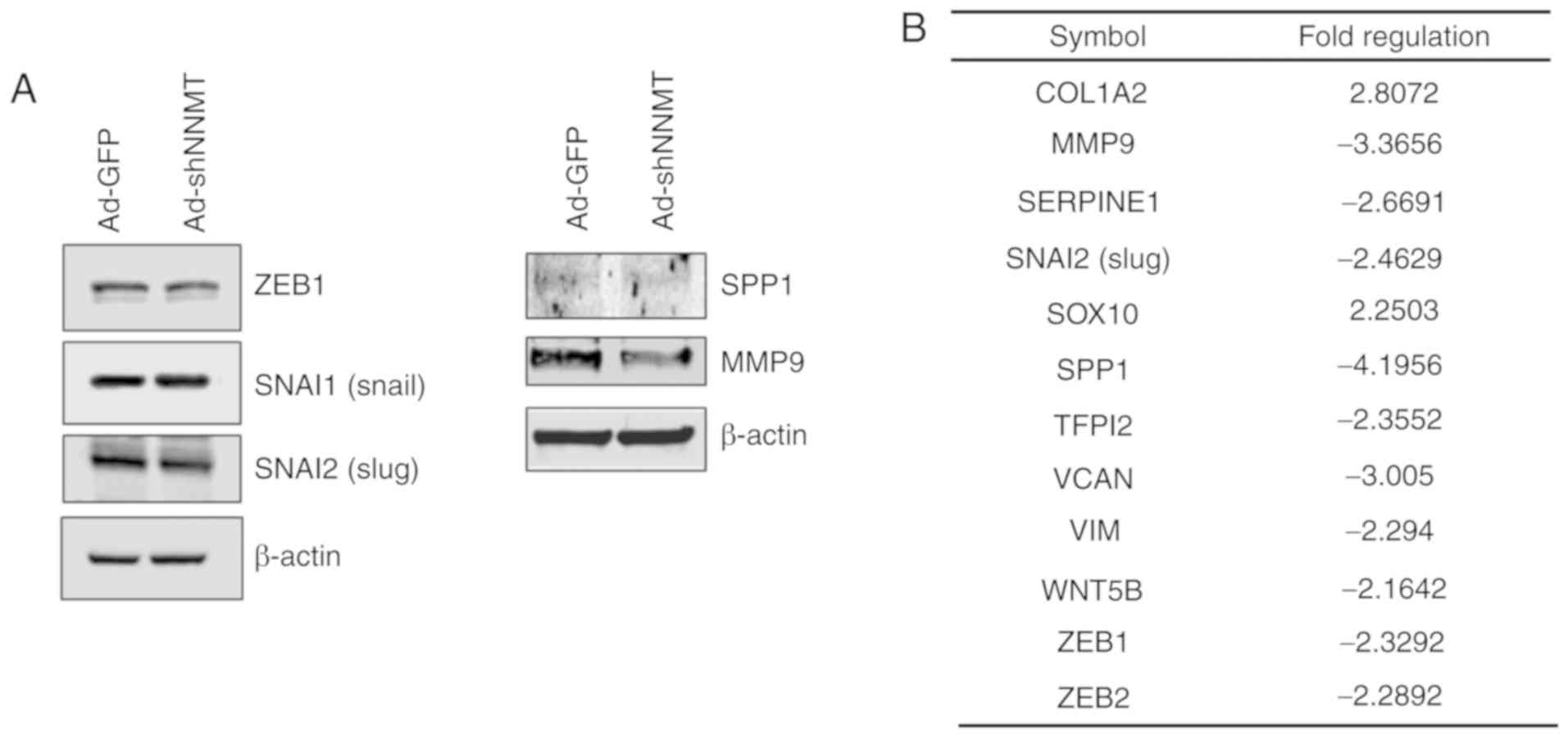
Effect of NNMT knockdown on
EMT-related gene expression
The role of NNMT in EMT regulation was further
investigated by detecting the protein expression levels of key EMT
effectors, including ZEB1, Slug and Snail. Notably, NNMT knockdown
did not have a considerable effect on ZEB1 and Snail, but the
expression level of Slug, MMP9 and osteopontin (SPP1) in
NNMT-knockdown cells was notably altered (Fig. 7A). To identify genes associated with
the invasiveness of SCC cells, and that are regulated by NNMT, the
expression of EMT-related genes was assessed using the
RT2 Profiler PCR Array. NNMT knockdown was found to
downregulate 10 of the 84 EMT-related genes assessed (Fig. 7B), negatively affecting the expression
levels of MMP9, SPP1 and versican core protein
(VCAN), which are involved in the regulation of
extracellular matrix (ECM) structure and function. In addition, the
mRNA expression level of Slug was inhibited by NNMT
knockdown. Collectively, these results indicate the metastatic
potential of SCC12 cells.
Discussion
Despite considerable progress in reducing its
incidence and mortality rates, and improving the survival of
affected patients, cancer remains a major threat to public health
worldwide. Metastasis to several organs, including the peritoneum
and lymph nodes, is the cause of >90% of all cancer-associated
mortalities (30). An EMT-induced
increase in the invasive ability of cancer cells is an important
process in carcinogenesis (26,29).
Metastasis, the process by which cancer cells leave the primary
tumor, disseminate to anatomically distant sites and form secondary
tumors, is a complex multistep process that results from an
intricate molecular cascade (29).
Numerous studies have provided insights into the molecular
components that facilitate this process, and understanding the
molecular mechanisms that regulate metastasis will allow the
development of novel therapeutic strategies. In the present study,
differentially expressed genes involved in tumor metastasis in cSCC
cells were identified using quantitative proteomics approaches.
After separating the proteins by 2-DE, LC-MS analysis was performed
to identify differentially expressed proteins between SCC12 and
SCC13 cells. A total of 39 proteins whose expression was increased
or decreased in SCC12 compared with SCC13 cells (>2-fold
increase or decrease; P<0.05) were identified. Among these
proteins, cytokeratin 19, tropomyosin 1 α chain, heat shock protein
β-1, cytokeratin 8, adenosylhomocysteinase, NNMT and glutathione
S-transferase omega 1 were significantly upregulated (P<0.05;
Fig. 2 and Table I). In particular, a protein spot
identified as NNMT by mass spectrometry, was upregulated at the
mRNA and protein levels in the invasive SCC12 cells, compared with
non-invasive SCC13 cells (Fig.
2).
NNMT catalyzes the N-methylation of
nicotinamide and other pyridines, regulating the metabolism of
drugs and xenobiotics (5,6). Various studies have revealed that NNMT
serves important roles in a wide range of biological processes
(7–16), suggesting that it may serve as a
diagnostic and prognostic marker, and a therapeutic target for
various tumors. Additionally, NNMT upregulation is involved in
tumor cell migration, and it may be used as a potential biomarker
for predicting tumor metastasis (16,20).
However, its role in cSCC has not previously been described. In the
present study, NNMT was found to be upregulated in an invasive,
compared with a non-invasive cSCC cell line. To further evaluate
the role of NNMT in invasive SCC12 cells, the effect of NNMT
depletion on various biological functions was assessed. This
demonstrated that NNMT knockdown suppressed the proliferation,
colony formation, migration and invasion capacities of SCC12 cells.
Therefore, NNMT may possess a tumor-promoting function in cSCC
cells.
EMT is activated at the transcriptional level during
embryonic development and wound healing, and contributes to the
initiation and progression of cancer, especially during tumor
metastasis (26,27). During EMT, polarized epithelial cells
lose their polarity and intercellular adhesion properties,
resulting in cytoskeleton reorganization and increased mesenchymal
properties. Invasion and metastasis are two of the defining
features of cancer (28). Cancer
cells characteristically adopt an invasive phenotype through the
process of EMT, and several signaling pathways reportedly regulate
EMT, which enhances cell motility and invasiveness (29). An understanding of the underlying
mechanisms of EMT are crucial, considering its vital involvement in
skin healing and its contribution to tumor invasion and metastasis.
In the present study, changes in the expression of EMT-related
genes were profiled using the RT2 Profiler PCR Array.
Several EMT-associated genes were found to be differentially
expressed in SCC12 cells, including the upregulation of vimentin,
KRT19 and KRT7, and the downegulation of BMP7
and MMP2 (Fig. 4B). SCC12 and
SCC13 cells differ in their expression levels of MMP2 and MMP9. Low
activity of MMP2 were revealed in SCC12 cells analyzed by
zymography (data not shown); therefore, the differences in the
expression and activity of MMP2 and MMP9 in the two cell lines
suggests that they may be involved in cancer progression, and that
different MMPs may be active in different cell types. A recent
study reported that NNMT promoted EMT in gastric cancer cells
(31); the present study revealed
that NNMT silencing increased the mRNA expression levels of
collagen α-2(I) chain (COL1A2) and the transcription factor
SOX10, as well as suppressing those of MMP9,
plasminogen activator inhibitor 1 (SERPINE1), SNAI2,
SPP1, tissue factor pathway inhibitor 2 (TFP12),
versican (VCAN), vimentin (VIM), WNT5B, ZEB1
and ZEB2 (Fig. 7). NNMT
knockdown negatively impacted the expression of genes that regulate
ECM structure and function, which included MMP9, SPP1 and
VCAN (formerly CSPG2). These genes are reportedly
involved in the regulation of SCC progression (32). The present study indicated that NNMT
could facilitate the EMT of cSCC cells, suggesting that NNMT may be
a promising prognostic indicator of invasive potential.
Although abnormal NNMT expression has been
identified in several types of cancer, the biological roles of NNMT
expression and activity in pathological conditions, including
cancer-cell migration and invasion, remain poorly understood. Tang
et al (20) demonstrated the
crucial role of NNMT in the promotion of cellular invasion in clear
cell renal cell carcinoma (ccRCC) cell lines; Akt inhibitor IV
markedly attenuated the NNMT-induced invasion of ccRCC cells,
indicating that activation of the PI3K/Akt signaling pathway is
required for NNMT-dependent invasion. This finding suggests a
potential mechanism in which NNMT acts upstream of the PI3K/Akt
pathway. Nevertheless, how EMT-related gene expression is regulated
in an NNMT-dependent manner remains unclear, in addition to how
NNMT-induced EMT is directly associated with tumor cell
metastasis.
In conclusion, the present study indicated that NNMT
was upregulated in invasive SCC12 cells, and that it may serve as a
potential biomarker of invasive tumor cells. NNMT knockdown
inhibited tumor cell proliferation and invasion, and NNMT
facilitated the EMT of cSCC cells by regulating EMT-related genes.
Therefore, NNMT may present a novel prognostic biomarker and
therapeutic target for patients with cSCC.
Acknowledgements
Not applicable.
Funding
This research was supported by Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (NRF-2018R1D1A1B07050577
and NRF- 2017R1A2B2005612).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
EPH and TJY conceived and designed the present
study. YSH and HYC performed the experiments and collected the
data. SYJ and YSP analyzed and interpreted the data. YSH and EPH
drafted the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
Gyeongsang National University Hospital. Samples were taken from
Gyeongsang National University Hospital with official written
ethical consent from the patients.
Patient consent for publication
All patients provided their written informed consent
for Publication and agreed to the publication of their associated
data and any accompanying images as appropriate.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lomas A, Leonardi-Bee J and Bath-Hextall
F: A systematic review of worldwide incidence of nonmelanoma skin
cancer. Br J Dermatol. 166:1069–1080. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Karia PS, Han J and Schmults CD: Cutaneous
squamous cell carcinoma: Estimated incidence of disease, nodal
metastasis, and deaths from disease in the United States 2012. J Am
Acad Dermatol. 68:957–956. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Alam M and Ratner D: Cutaneous
squamous-cell carcinoma. N Engl J Med. 344:975–983. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
O'Hara J, Ferlito A, Takes RP, Rinaldo A,
Strojan P, Shaha AR, Rodrigo JP and Paleri V: Cutaneous squamous
cell carcinoma of the head and neck metastasizing to the parotid
glandea review of current recommendations. Head Neck. 33:1789–1795.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rini J, Szumlanski C, Guerciolini R and
Weinshilboum RM: Human liver nicotinamide N-methyltransferase:
Ion-pairing radiochemical assay, biochemical properties and
individual variation. Clin Chim Acta. 186:359–374. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aksoy S, Szumlanski CL and Weinshilboum
RM: Human liver nicotinamide N-methyltransferase. cDNA cloning,
expression, and biochemical characterization. J Biol Chem.
269:14835–14840. 1994.PubMed/NCBI
|
|
7
|
Xu J, Moatamed F, Caldwell JS, Walker JR,
Kraiem Z, Taki K, Brent GA and Hershman JM: Enhanced expression of
nicotinamide N-methyltransferase in human papillary thyroid
carcinoma cells. J Clin Endocrinol Metab. 88:4990–4996. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lim BH, Cho BI, Kim YN, Kim JW, Park ST
and Lee CW: Overexpression of nicotinamide N-methyltransferase in
gastric cancer tissues and its potential post-translational
modification. Exp Mol Med. 38:455–465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim DS, Choi YP, Kang S, Gao MQ, Kim B,
Park HR, Choi YD, Lim JB, Na HJ, Kim HK, et al: Panel of candidate
biomarkers for renal cell carcinoma. J Proteome Res. 9:3710–3719.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yao M, Tabuchi H, Nagashima Y, Baba M,
Nakaigawa N, Ishiguro H, Hamada K, Inayama Y, Kishida T, Hattori K,
et al: Gene expression analysis of renal carcinoma: Adipose
differentiation-related protein as a potential diagnostic and
prognostic biomarker for clear-cell renal carcinoma. J Pathol.
205:377–387. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sartini D, Muzzonigro G, Milanese G,
Pierella F, Rossi V and Emanuelli M: Identification of nicotinamide
N-methyltransferase as a novel tumor marker for renal clear cell
carcinoma. J Urol. 176:2248–2254. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Iacobuzio-Donahue CA, Maitra A, Shen-Ong
GL, van Heek T, Ashfaq R, Meyer R, Walter K, Berg K, Hollingsworth
MA, Cameron JL, et al: Discovery of novel tumor markers of
pancreatic cancer using global gene expression technology. Am J
Pathol. 160:1239–1249. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tsuchiya A, Sakamoto M, Yasuda J, Chuma M,
Ohta T, Ohki M, Yasugi T, Taketani Y and Hirohashi S: Expression
profiling in ovarian clear cell carcinoma: Identification of
hepatocyte nuclear factor-1 beta as a molecular marker and a
possible molecular target for therapy of ovarian clear cell
carcinoma. Am J Pathol. 163:2503–2512. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Roessler M, Rollinger W, Palme S, Hagmann
ML, Berndt P, Engel AM, Schneidinger B, Pfeffer M, Andres H, Karl
J, et al: Identification of nicotinamide N-methyltransferase as a
novel serum tumor marker for colorectal cancer. Clin Cancer Res.
11:6550–6557. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim J, Hong SJ, Lim EK, Yu YS, Kim SW, Roh
JH, Do IG, Joh JW and Kim DS: Expression of nicotinamide
N-methyltransferase in hepatocellular carcinoma is associated with
poor prognosis. J Exp Clin Cancer Res. 28:202009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wu Y, Siadaty MS, Berens ME, Hampton GM
and Theodorescu D: Overlapping gene expression profiles of cell
migration and tumor invasion in human bladder cancer identify
metallothionein 1E and nicotinamide N-methyltransferase as novel
regulators of cell migration. Oncogene. 27:6679–6689. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tomida M, Mikami I, Takeuchi S, Nishimura
H and Akiyama H: Serum levels of nicotinamide N-methyltransferase
in patients with lung cancer. J Cancer Res Clin Oncol.
135:1223–1229. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sartini D, Pozzi V, Renzi E, Morganti S,
Rocchetti R, Rubini C, Santarelli A, Lo Muzio L and Emanuelli M:
Analysis of tissue and salivary nicotinamide N-methyltransferase in
oral squamous cell carcinoma: Basis for the development of a
noninvasive diagnostic test for early-stage disease. Biol Chem.
393:505–511. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sartini D, Muzzonigro G, Milanese G, Pozzi
V, Vici A, Morganti S, Rossi V, Mazzucchelli R, Montironi R and
Emanuelli M: Upregulation of tissue and urinary nicotinamide
N-methyltransferase in bladder cancer: Potential for the
development of a urine-based diagnostic test. Cell Biochem Biophys.
65:473–483. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang SW, Yang TC, Lin WC, Chang WH, Wang
CC, Lai MK and Lin JY: Nicotinamide N-methyltransferase induces
cellular invasion through activating matrix metalloproteinase-2
expression in clear cell renal cell carcinoma cells.
Carcinogenesis. 32:138–145. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Emanuelli M, Santarelli A, Sartini D,
Ciavarella D, Rossi V, Pozzi V, Rubini C and Lo Muzio L:
Nicotinamide N-Methyltransferase upregulation correlates with
tumour differentiation in oral squamous cell carcinoma. Histol
Histopathol. 25:15–20. 2010.PubMed/NCBI
|
|
22
|
Chen C, Wang X, Huang X, Yong H, Shen J,
Tang Q, Zhu J, Ni J and Feng Z: Nicotinamide N-methyltransferase: A
potential biomarker for worse prognosis in gastric carcinoma. Am J
Cancer Res. 6:649–663. 2016.PubMed/NCBI
|
|
23
|
Xie X, Liu H, Wang Y, Zhou Y, Yu H, Li G,
Ruan Z, Li F, Wang X and Zhang J: Nicotinamide N-methyltransferase
enhances resistance to 5-fluorouracil in colorectal cancer cells
through inhibition of the ASK1-p38 MAPK pathway. Oncotarget.
7:45837–45848. 2016.PubMed/NCBI
|
|
24
|
Hesse K, Satzger I, Schacht V, Köther B,
Hillen U, Klode J, Schaper K and Gutzmer R: Characterisation of
prognosis and invasion of cutaneous squamous cell carcinoma by
podoplanin and E-cadherin expression. Dermatology. 232:558–565.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cañueto J, Martín-Vallejo J,
Cardeñoso-Álvarez E, Fernández-López E, Pérez-Losada J and
Román-Curto C: Rapid growth rate is associated with poor prognosis
in cutaneous squamous cell carcinoma. Clin Exp Dermatol.
43:876–882. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial- mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liang L, Zeng M, Pan H, Liu H and He Y:
Nicotinamide N-methyltransferase promotes epithelial-mesenchymal
transition in gastric cancer cells by activating transforming
growth factor-β1 expression. Oncol Lett. 15:4592–4598.
2018.PubMed/NCBI
|
|
32
|
Kashyap MK, Marimuthu A, Kishore CJ, Peri
S, Keerthikumar S, Prasad TS, Mahmood R, Rao S, Ranganathan P,
Sanjeeviah RC, et al: Genomewide mRNA profiling of esophageal
squamous cell carcinoma for identification of cancer biomarkers.
Cancer Biol Ther. 8:36–46. 2009. View Article : Google Scholar : PubMed/NCBI
|