Introduction
Lung cancer is one of the most prevalent and
aggressive malignancies, and is a major cause of cancer-related
deaths in the world, having the highest incidence and mortality
rate among all cancers (1). Lung
cancer is routinely classified as non-small cell lung cancer
(NSCLC) and small-cell lung cancer (SCLC), and the former accounts
for approximately 80–85% of all lung cancers. Only 17.7% of the
patients who are diagnosed with lung cancer live more than 5 years.
Most patients with NSCLC are diagnosed in the advanced stages, and
the overall 5-year survival rate is low due to its recurrence and
metastasis (2–6). Therefore, there is an urgent need to
explore the mechanisms of NSCLC and to find effective biomarkers
for prognostic evaluation and therapy.
DEK is a potential biomarker and oncogene,
which is reported in many cancers (7–9). The
DEK gene encodes the DEK oncoprotein with one SAP domain
which binds to cruciform and superhelical DNA to then influence
various important nuclear processes, such as transcriptional
regulation, signal transduction, differentiation, apoptosis, RNA
processing and DNA replication (10–16).
Increased expression of DEK has been revealed to be associated with
various neoplasms, including breast, cervical, and neuroendocrine
prostate cancer, hematologic malignancies, gastric adenocarcinoma,
and lung cancer (8,17–23).
DEK was revealed to promote epithelial-mesenchymal transition (EMT)
and angiogenesis by activating the PI3K/AKT/mTOR pathway in triple
negative breast cancers (8).
Depletion of DEK suppressed the proliferation, migration, invasion,
and angiogenesis of breast cancer cells (8). The silencing of DEK induced apoptosis
and senescence via the upregulation of NF-κB (23) and downregulated tumorigenesis and
metastasis by the DEK/Gsk3β/β-catenin axis in cervical cancer cells
(22). In acute myelogenous
leukemia, DEK was involved in chromosomal translocation by binding
to the HIV-2 enhancer (11). In
neuroendocrine prostate cancer and gastric adenocarcinoma,
increased DEK expression was revealed to be an independent clinical
risk factor and associated with shorter disease-free survival of
cancer patients (9,21). These findings indicated that DEK may
be a potential biomarker and predicator for tumor progression and
therapy. However, the role and underlying mechanism of DEK in lung
cancer has been poorly explored.
In the present study, the expression data for DEK in
lung cancers was collected and the correlations between DEK
expression and clinicopathological parameters of lung cancers was
analyzed. The role of DEK in the proliferation and invasion of lung
cancer cells was also investigated and the effects of DEK on the
Wnt signaling pathway and EMT process were explored.
Materials and methods
The cancer genome atlas (TCGA) data
collection and analysis
The expression data of DEK in lung cancers and
normal lung tissues, and the correlations between DEK expression
and prognosis of lung cancers were obtained and analyzed using the
online databases Gene Expression Profiling Interactive Analysis
(GEPIA; http://gepia.cancer-pku.cn) and
UALCAN (http://ualcan.path.uab.edu) (24), which is based on The Cancer Genome
Atlas (TCGA).
Cell lines and transfection
All the cell lines used in the present study were
purchased from the Cell Bank of the Chinese Academy of Sciences
(Shanghai, China). The normal human bronchial epithelial (HBE) cell
line was cultured in minimal essential medium (Gibco; Invitrogen;
Thermo Fisher Scientific, Inc.), and the human lung cancer cell
lines A549, H1299, SK-MES-1, LK2, H460 and H661 were cultured in
RPMI-1640 medium (Gibco; Invitrogen; Thermo Fisher Scientific,
Inc.). Both culture mediums contained 10% fetal bovine serum (FBS)
(FB15015; Clark Bioscience). All these cells were maintained at
37°C in an atmosphere containing 5.0% carbon dioxide (25–27).
A549 and H1299 cells were plated in 6-well plates
for 24 h and cultured to 70–80% confluence before gene
transfection. The plasmids contained the DEK or DEK-shRNA
sequences that were synthesized by Shanghai GeneChem Co., Ltd. The
plasmids were transfected into cells using Lipofectamine 3000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. An empty vector or a plasmid that
contained a scrambled shRNA sequence served as a negative control
for the plasmid of DEK or DEK-shRNA,
respectively.
Western blot analysis
The total protein extracted from each sample was
separated by cell lysis buffer (Pierce; Thermo Fisher Scientific,
Inc.) and quantified using the Bradford method. Totally, 60 µg of
protein was separated by SDS-PAGE (10%) and transferred to a
polyvinylidene fluoride membrane (EMD Millipore). The bands were
blocked in 5% non-fat milk for 1 h at room temperature. The
membranes were incubated with primary antibodies (Table I) overnight at 4°C and then washed
with tris-buffered saline containing 0.1% Tween-20. The membranes
were incubated with an anti-rabbit or anti-mouse IgG antibody
(1:2,000; ProteinTech Group, Inc.) conjugated with horseradish
peroxidase (BIOSS) at 37°C for 2 h. Finally, the membranes were
visualized using an enhanced chemilumiscence reagent (Pierce;
Thermo Fisher Scientific, Inc.) and detected with a bioimaging
system (DNR Bio-Imaging System). The blots were stripped according
to the manufacturer's protocol using a stripping buffer (Beyotime
Institute of Biotechnology) and the bands that had similar/the same
molecular weight were re-probed. The relative protein levels were
analyzed with ImageJ software (version 1.47; National Institutes of
Health) using GAPDH as a loading control.
 | Table I.List of antibodies used for western
blotting. |
Table I.
List of antibodies used for western
blotting.
| Antibody name | Source | Catalog number | Host | Dilution |
|---|
| DEK | ProteinTech Group,
Inc. | 16448-1-AP | Rabbit | 1:2,000 |
| GSK3β | Cell Signaling
Technology, Inc. | 5676 | Rabbit | 1:1,000 |
| Axin | Santa Cruz
Biotechnology, Inc. | SC-293109 | Mouse | 1:100 |
| β-catenin | BD Biosciences | MAB13291 | Rabbit | 1:1,000 |
| Active
β-catenin | BD Biosciences | 5279 | Rabbit | 1:1,000 |
| c-Myc | BD Biosciences | MAB3696 | Rabbit | 1:1,000 |
| MMP7 | Santa Cruz
Biotechnology, Inc. | SC-515703 | Mouse | 1:100 |
| Cyclin D1 | Santa Cruz
Biotechnology, Inc. | SC-8396 | Rabbit | 1:100 |
| DVL1 | Santa Cruz
Biotechnology, Inc. | SC-8026 | Mouse | 1:100 |
| LRP6 | Santa Cruz
Biotechnology, Inc. | SC-25317 | Mouse | 1:100 |
| LEF1 | Santa Cruz
Biotechnology, Inc. | SC-374522 | Mouse | 1:100 |
| TCF4 | Santa Cruz
Biotechnology, Inc. | SC-166699 | Mouse | 1:100 |
| Snail | Cell Signaling
Technology, Inc. | 3879 | Rabbit | 1:500 |
| Vimentin | Cell Signaling
Technology, Inc. | 5741 | Rabbit | 1:1,000 |
| E-cadherin | Cell Signaling
Technology, Inc. | 3195 | Rabbit | 1:1,000 |
| N-cadherin | Cell Signaling
Technology, Inc. | 13116 | Rabbit | 1:1,000 |
| EGFR | ProteinTech Group,
Inc. | 18986-1-AP | Rabbit | 1:1,000 |
| KRAS | ProteinTech Group,
Inc. | 12063-1-AP | Rabbit | 1:1,000 |
| ALK | ProteinTech Group,
Inc. | 24184-1-AP | Mouse | 1:1,000 |
| GAPDH | Santa Cruz
Biotechnology, Inc. | SC-47724 | Rabbit | 1:1,000 |
Colony forming assay
Cells were plated in 6-well plates by seeding 1,000
cells/well 24 h after transfection and cultured for 14 days. The
medium was changed every 4 days. Then, the plates were washed with
phosphate-buffered saline (PBS). The cells were fixed with 4%
paraformaldehyde for 20 min and stained with hematoxylin for 10 min
at 37°C. The number of colonies formed with more than 50 cells were
counted using a BioImaging system.
Cell proliferation assay
Twenty-four hours after transfection, the cells were
plated into 96-well plates at a density of 3,000 cells/well in
medium containing 10% FBS. The quantitation of cell viability was
detected using Cell Counting Kit-8 (CCK-8; Dojindo Molecular
Technologies, Inc.). Each well was supplemented with CCK-8 reagent
at 1:10 (v/v) per 100 µl and incubated for 2 h at 37°C. The data
were quantitated spectrophotometrically with a test wavelength of
450 nm.
Cell migration and invasion
analyses
Cell migration and invasion assays were performed in
24-well Transwell chambers containing inserts with a pore size of 8
µm (Costar Corporation). For Matrigel invasion assays, the upper
side of an 8-µm pore was coated with Matrigel basement membrane
matrix (1:8 dilution; BD Biosciences) for at least 2 h at 37°C.
After gelation of the Matrigel, the cells were seeded
(1×105 cells/well) to the upper chambers of a 100-µl
medium supplemented with 2% FBS, and the lower chambers were filled
with 600 µl 20% FBS as the chemoattractant. For the migration
assays, the upper chambers were filled with 100 µl of medium
supplemented with 2% FBS not-coated with Matrigel, and the lower
chambers were filled with 600 µl 20% FBS as a chemoattractant.
After 20 h, the filters were fixed with 4% paraformaldehyde and
stained with hematoxylin for 10 min at 37°C. The non-invading cells
on the upper surface were cleared with a cotton swab. Ten randomly
selected high-power fields were observed under light microscopy
(magnification, ×200), and the number of migrated or invaded cells
was counted. All the experiments aforementioned were performed in
triplicate independently.
Statistical analysis
Statistical analyses were performed using
statistical software GraphPad Prism 6.0 (GraphPad Software, Inc.).
The results were analyzed using Student's t-test, Spearman's rank
correlation, and Kaplan-Meier analysis. A P<0.05 was considered
to indicate a statistically significant difference.
Results
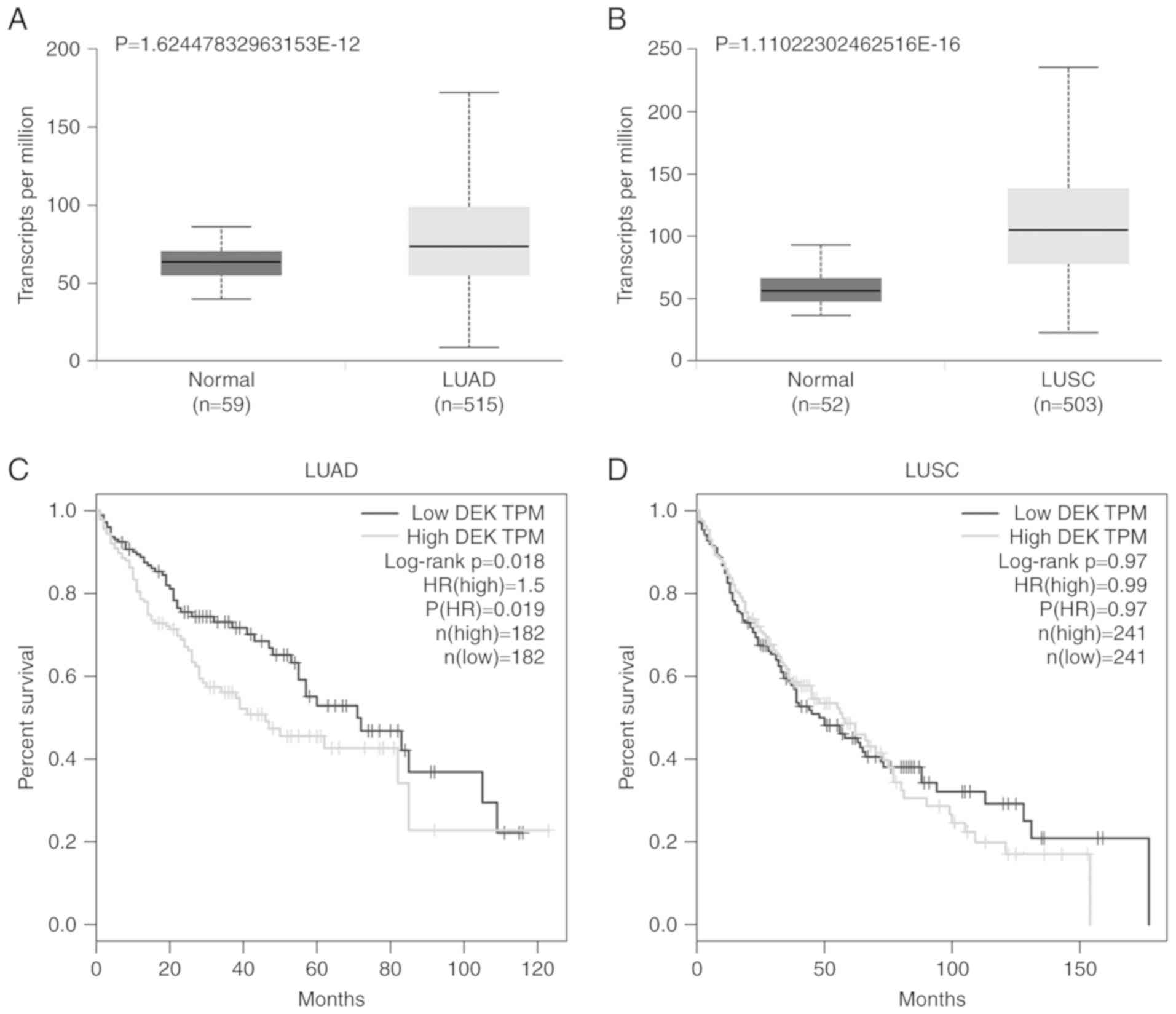
DEK expression is higher in lung
cancers than in normal lung tissues and correlated with poor
prognosis of lung adenocarcinomas
The expression data of DEK in lung cancers and
normal lung tissues were analyzed using the online database GEPIA,
UALCAN based on TCGA. The data from UALCAN revealed that the
expression level of DEK in lung adenocarcinomas (LUAD) (P<0.01)
and lung squamous cell carcinomas (LUSC) (P<0.01) was
significantly higher than that in normal lung tissues (Fig. 1A and B). According to the
Kaplan-Meier graph obtained from the GEPIA database, LUAD patients
with increased expression of DEK had significantly shorter overall
survival (P=0.018, Fig. 1C).
However, the expression of DEK was not correlated with the survival
of LUSC patients (P=0.97, Fig.
1D).
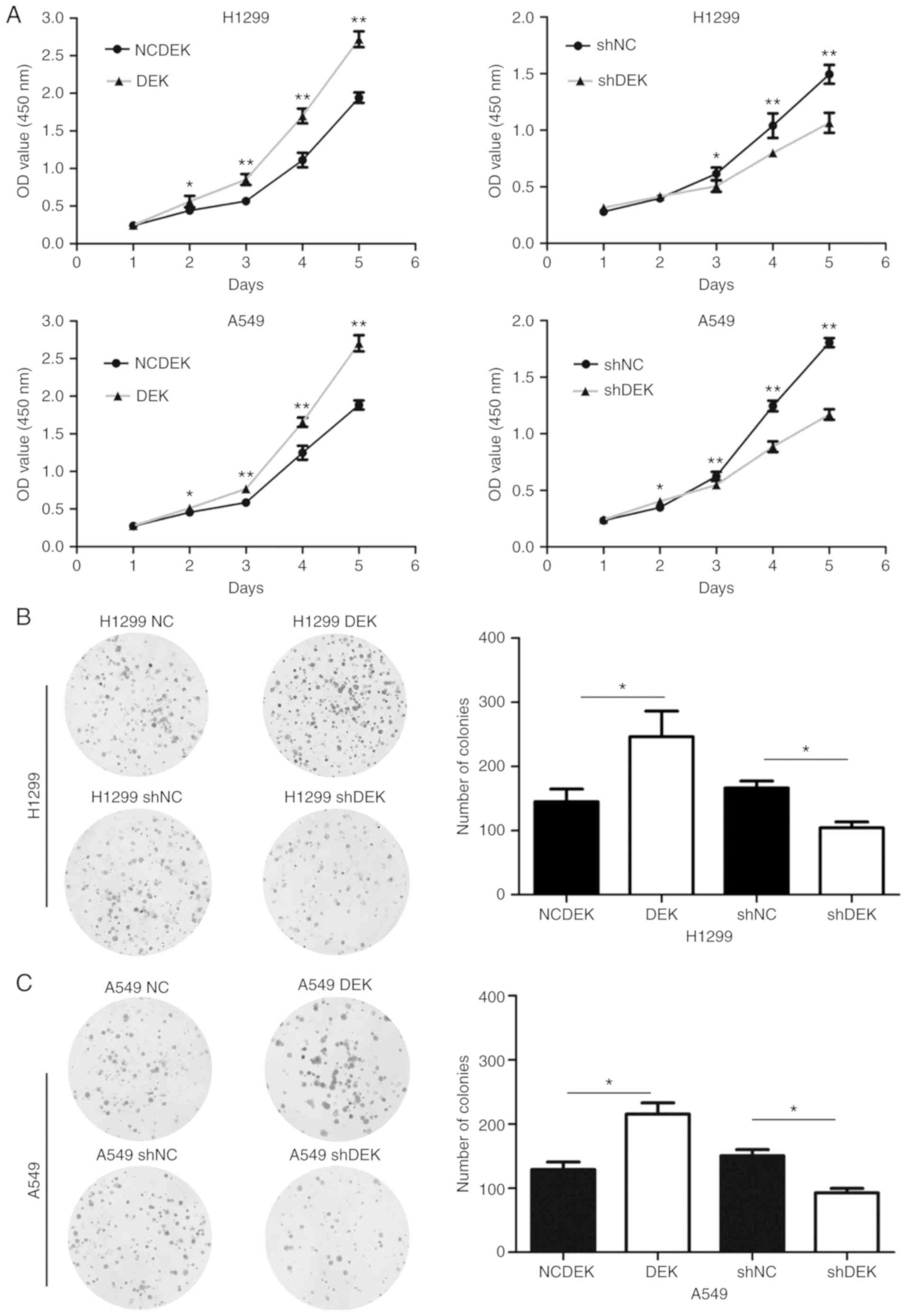
DEK promotes the proliferation, colony
formation, migration, and invasion of lung cancer cells
The expression levels of DEK in normal HBE and NSCLC
cell lines A549, H1299, SK, LK2, H460, and H661 were examined and
are revealed in the Fig. S1. DEK
was expressed at a relatively low level in A549, H460 and LK2
cells, a moderate level in HBE and H1299 cells, and a high level in
H661 and SK-MES-1 cells. We selected A549 and H1299 for further
experiments.
Overexpression of DEK by DEK transfection
enhanced the proliferation rate (H1299-DEK, P<0.05; A549-DEK,
P<0.05) and the ability of colony formation (H1299-DEK,
P<0.05; A549-DEK, P<0.05) of H1299-DEK and A549-DEK cells
compared to the control cells (Fig.
2). In contrast, knockdown of DEK expression by shRNA inhibited
the cellular proliferation rate (H1299-shDEK, P<0.05;
A549-shDEK, P<0.05) and the ability of colony formation of cells
(H1299-shDEK, P<0.05; A549-shDEK, P<0.05) of H1299-shDEK and
A549-shDEK, compared to the control cells (Fig. 2).
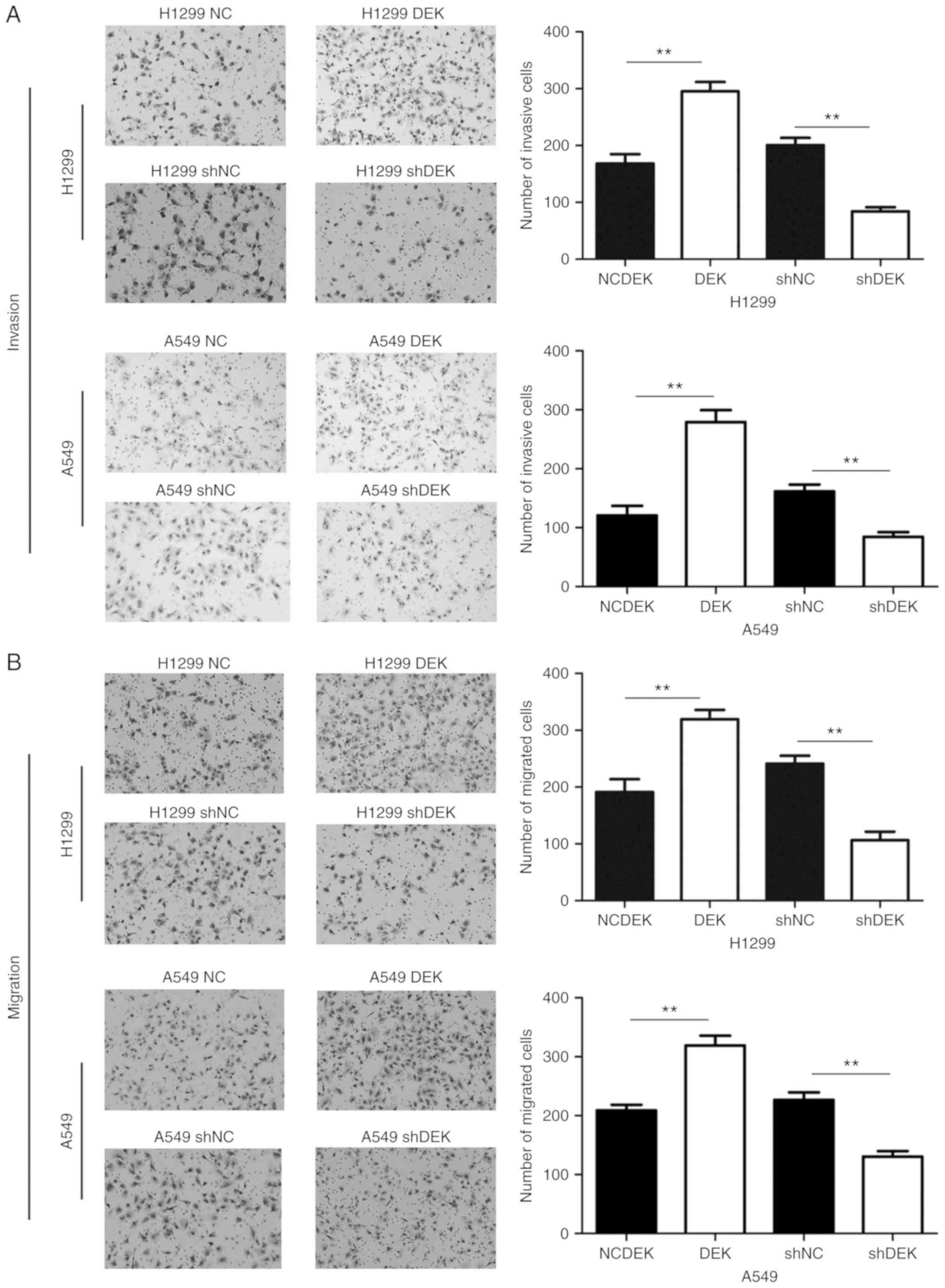
Furthermore, enhanced DEK expression promoted the
migrative and invasive abilities of A549-DEK and H1299-DEK cells
compared with the control cells (P<0.05, respectively).
Conversely, knockdown of DEK expression inhibited the migrative and
invasive abilities of H1299-shDEK and A549-shDEK cells (P<0.01,
respectively) (Fig. 3).
DEK upregulates the activities of
β-catenin and the Wnt signaling pathway
Enhanced DEK expression promoted an increase in the
expression levels of active-β-catenin and LEF1 and inhibited the
levels of Gsk3β and Axin in H1299-DEK and A549-DEK cells (P<0.05
in both cases). The expression levels of some target genes of the
Wnt pathway, such as cyclin D1, c-Myc, and MMP7, were also
significantly increased in H1299-DEK and A549-DEK cells (P<0.05
in both cases). However, the levels of total β-catenin, DVL1, LRP6
and TCF4 were not markedly altered (P>0.05) (Fig. 4A-C). Conversely, knockdown of DEK
expression downregulated the levels of active-β-catenin, LEF1,
cyclin D1, c-Myc, and MMP7, and increased the expression of
Gsk3β and Axin in H1299-ShDEK and A549-ShDEK cells (P<0.05). The
expression levels of total β-catenin, DVL1, LRP6 and TCF4 were not
markedly altered (P>0.05) (Fig.
5A-C).
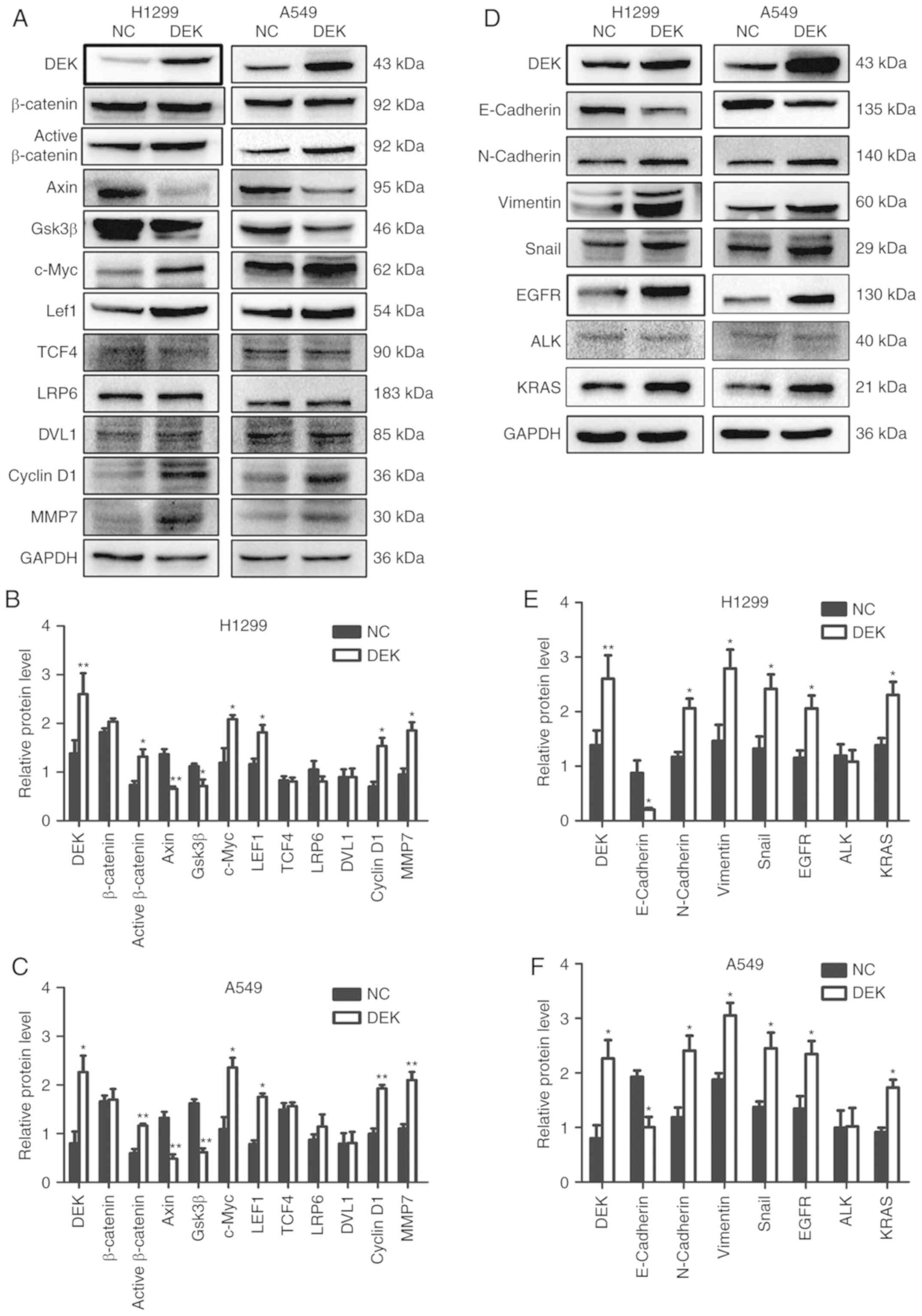 | Figure 4.Expression of Wnt and EMT-related
proteins, EGFR, KRAS, and ALK after forced upregulation of DEK in
lung cancer cells. Representative results of (A and D) western blot
analysis, and histograms of relative protein levels in (B and E)
H1299 cells and (C and F) A549 cells. GAPDH served as an internal
control. *P<0.05, **P<0.01. EGFR, epidermal growth factor
receptor; KRAS, Kirsten rat sarcoma viral oncogene homolog; ALK,
anaplastic lymphoma kinase; NC, negative control cells; DEK, cells
transfected with DEK. |
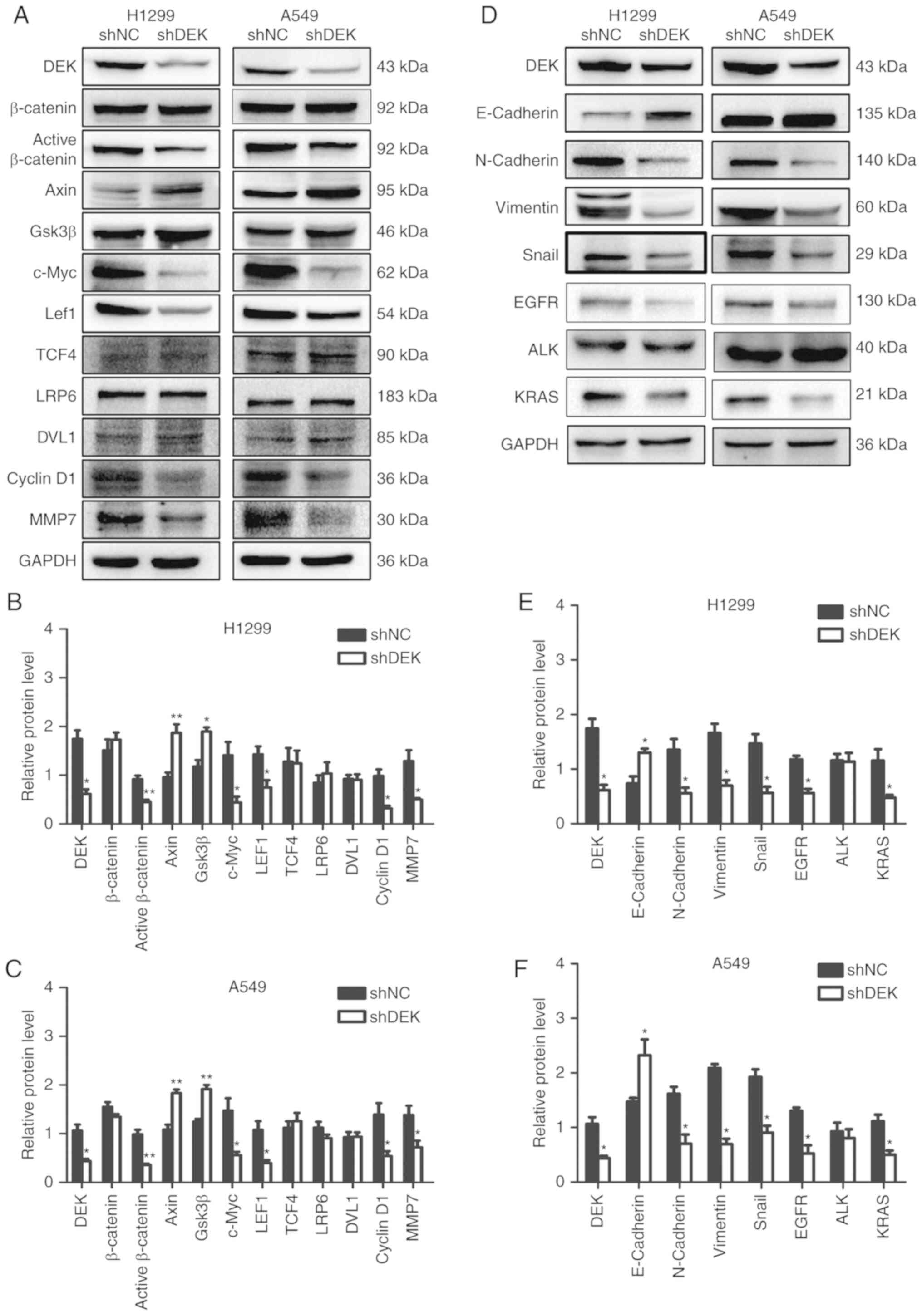 | Figure 5.Expression of Wnt and EMT-related
proteins, EGFR, KRAS, and ALK after forced downregulation of DEK in
lung cancer cells. Representative results of (A and D) western blot
analysis, and histograms of relative protein levels in (B and E)
H1299 cells and (C and F) A549 cells. GAPDH served as an internal
control. *P<0.05, **P<0.01. EGFR, epidermal growth factor
receptor; KRAS, Kirsten rat sarcoma viral oncogene homolog; ALK,
anaplastic lymphoma kinase; shNC, cells transfected with scrambled
shRNA; shDEK, cells transfected with DEK-shRNA. |
DEK promotes EMT
The expression levels of vimentin, Snail, and
N-cadherin were increased, while E-cadherin was decreased in
H1299-DEK and A549-DEK cells (P<0.05) (Fig. 4D-F). Conversely, in H1299-shDEK and
A549-shDEK cells, the expression levels of vimentin, Snail, and
N-cadherin were decreased, while E-cadherin was significantly
increased (P<0.05) (Fig.
5D-F).
DEK expression is correlated with the
levels of EGFR and KRAS in lung adenocarcinomas
According to the data from the GEPIA database, the
expression of DEK was correlated with the expression of EGFR and
KRAS (P<0.001, respectively), but was not correlated with the
expression of ALK (P=0.82) or c-ros oncogene 1 receptor kinase
(ROS1) (P=0.40) in LUADs (Fig. 6).
In LUSCs, DEK expression was not correlated with the expression of
EGFR (P=0.46), KRAS (P=0.061), ALK (P=0.82), or ROS1 (P=0.053)
(Fig. 7). It was revealed by
western blot analysis that enhanced DEK expression promoted the
expression levels of EGFR and KRAS in H1299-DEK and A549-DEK cells
(P<0.05 in both cases), however, the level of ALK was not
markedly altered (P>0.05) (Fig.
4D-F). Conversely, knockdown of DEK expression downregulated
the levels of EGFR and KRAS in H1299-shDEK and A549-shDEK cells
(P<0.05, respectively), however the level of ALK was not
markedly altered (P>0.05) (Fig.
5D-F).
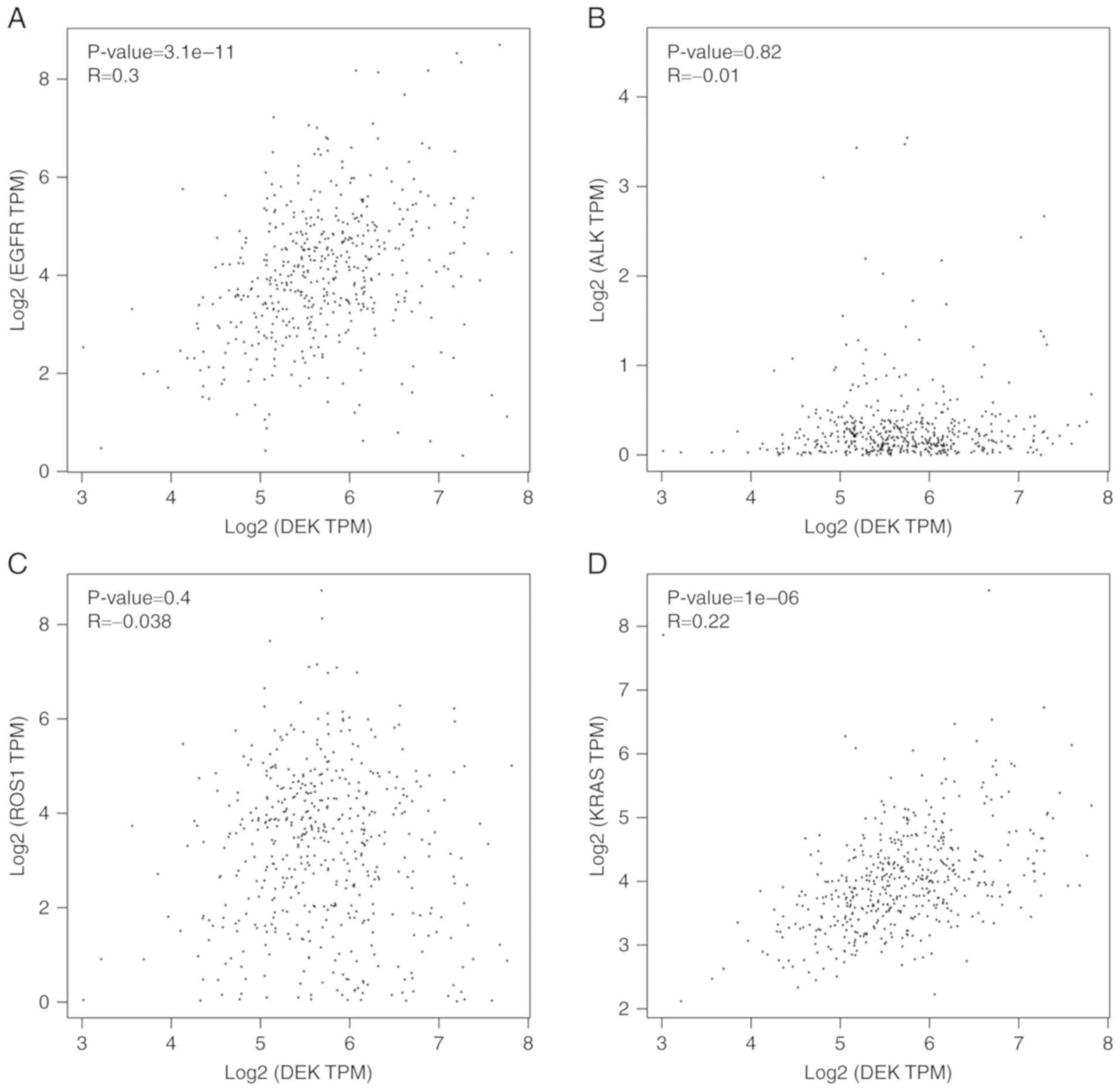 | Figure 6.Correlations between DEK and EGFR,
ALK, ROS1, and KRAS in LUAD. The scatter diagrams that display
correlations between DEK and (A) EGFR, (B) ALK, (C) ROS1, and (D)
KRAS in LUADs were obtained from GEPIA. EGFR, epidermal growth
factor receptor; ALK, anaplastic lymphoma kinase; ROS1, c-ros
oncogene 1 receptor kinase; KRAS, Kirsten rat sarcoma viral
oncogene homolog; LUAD, lung adenocarcinoma; GEPIA, Gene Expression
Profiling Interactive Analysis. |
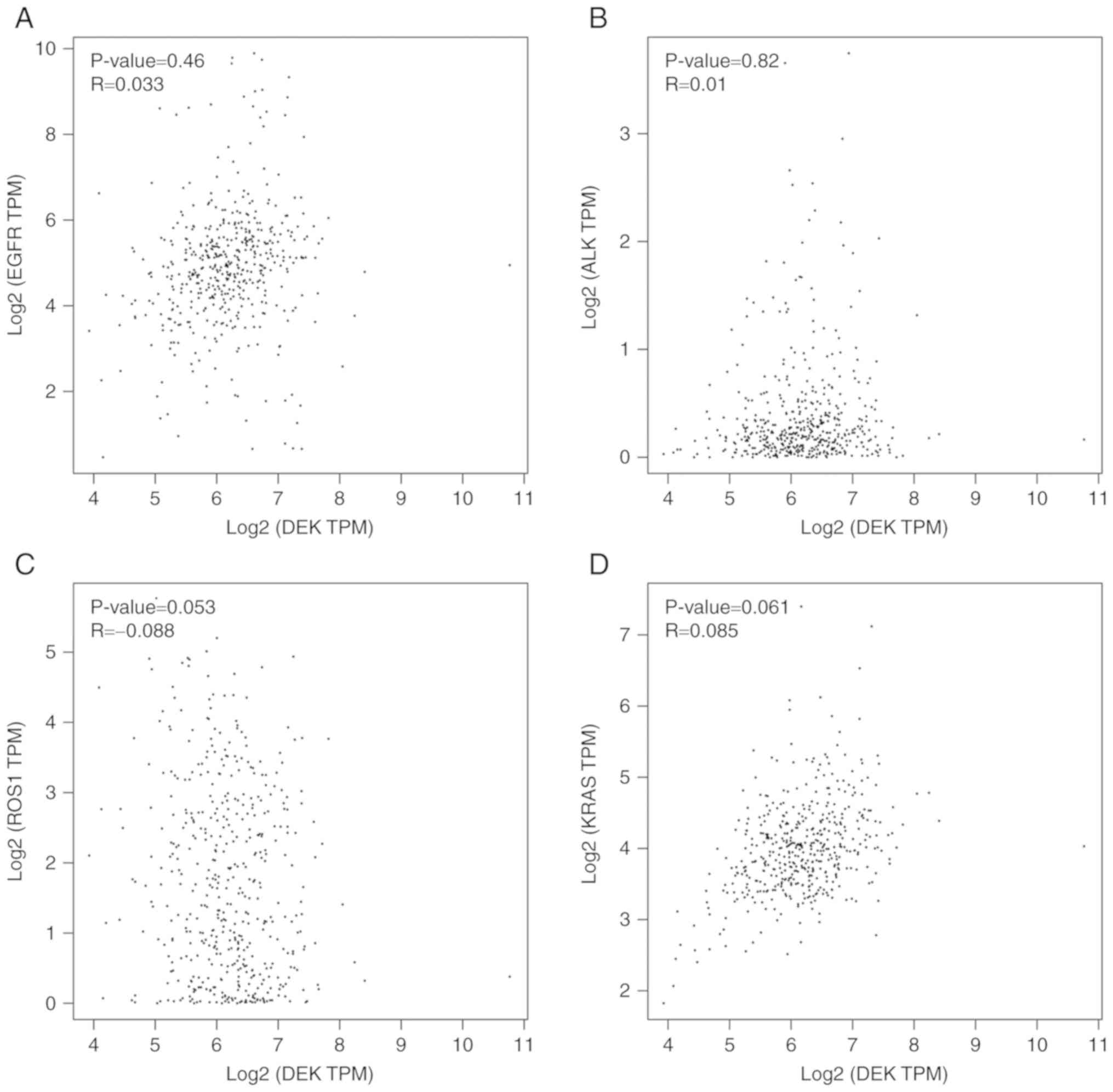 | Figure 7.Correlations between DEK and EGFR,
ALK, ROS1, and KRAS in LUSC. The scatter diagrams that display
correlations between DEK and (A) EGFR, (B) ALK, (C) ROS1, and (D)
KRAS in LUSCs were obtained from GEPIA. EGFR, epidermal growth
factor receptor; ALK, anaplastic lymphoma kinase; ROS1, c-ros
oncogene 1 receptor kinase; KRAS, Kirsten rat sarcoma viral
oncogene homolog; LUSC, lung squamous cell carcinoma; GEPIA, Gene
Expression Profiling Interactive Analysis. |
Discussion
DEK is located on chromosome 6, consists of
11 exons and is controlled by a TATA-less promoter that contains a
CpG island around the transcriptional start and several binding
sites for transcription factors (15). DEK was first isolated as part
of a fusion that arises in a subtype of acute myeloid leukemias
involving the (6,9) chromosomal translocation (28). DEK regulates hematopoiesis by
increasing the repopulating and self-renewal capacity of
hematopoietic stem cells while decreasing the number and cycling of
hematopoietic progenitor cells (29). DEK has been reported as an
oncogene in breast, cervical, and neuroendocrine prostate cancer,
acute myeloid leukemia, gastric adenocarcinoma, and lung cancer
(8,18,20–23,30).
Previous studies have documented the expression of
DEK in NSCLC (18,20,25),
which indicated that DEK expression was correlated with the
progression of NSCLCs. However, the effect and mechanism of DEK
underlying the progression and prognosis of NSCLCs still requires
further investigation. In the present study, by analyzing the data
from TCGA, it was confirmed that DEK was prominently overexpressed
in NSCLCs, when compared to DEK expression in normal lung tissues,
and that an increased DEK level predicted poor prognosis in LUAD
patients. Furthermore, DEK expression was correlated with EGFR,
KRAS in LUAD. EGFR is a common therapeutic target in NSCLCs
(31–33). Therefore, it is possible that DEK is
involved in the mechanism of gene target therapy. The level of DEK
may be a predictive indicator of the therapeutic effect of
gene-targeted drugs, which require further investigation.
The mechanisms by which DEK promotes the
proliferation and invasion of lung cancers are poorly investigated.
A study by Wang et al reported that DEK depletion
downregulated RhoA expression and indicated that DEK inhibited cell
migration by inactivation of the RhoA/ROCK/MLC signaling pathway
(25). To explore the regulating
mechanisms of DEK in NSCLCs, the effects of DEK on the Wnt
signaling pathway and EMT were examined. The results revealed that
DEK expression upregulated the levels of active β-catenin and Wnt
target genes, such as cyclin D1, c-Myc and MMP7.
Furthermore, DEK expression activated the EMT process by decreasing
the expression of E-cadherin, and enhancing the expression of
N-cadherin, vimentin, and Snail. The Wnt pathway is important in
embryonic development and is involved in the development of many
cancers (34,35). The present results indicated that
DEK promoted the proliferation and invasion of NSCLC and activated
the Wnt signaling pathway and EMT process. In the present study, it
was also observed that DEK did not regulate the total expression
level of β-catenin, but regulated only the expression level of
active β-catenin. In addition, DEK overexpression downregulated the
levels of Gsk3β and Axin, which are key proteins of the
Gsk3β/Axin/APC complex inducing the phosphorylation and degradation
of β-catenin. Thus, DEK may enhance the activity of β-catenin by
regulating the Gsk3β/Axin/APC complex, which requires further
confirmation.
In conclusion, DEK was revealed to be overexpressed
in lung cancers, which indicated poor prognosis in lung
adenocarcinomas. Overexpression of DEK activated the Wnt signaling
pathway and EMT process, and promoted the proliferation and
invasion of lung cancers. DEK is a potential prognostic biomarker,
and a potential target of gene therapy for NSCLC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81372497 to HTX) and
Program for Liaoning Excellent Talents in University (grant no.
LR2015067 to HTX).
Availability of data and material
The data supporting the conclusions of this article
are included within the article and its supplementary information
files.
Authors' contributions
HTX designed the study. MQY and HTX participated in
drafting the manuscript. MQY, LLB, LL, YWZ, ZW, ZHL, CCL and WJH
performed the experiments. All authors have read and approved the
final version of this manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TCGA
|
The Cancer Genome Atlas
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
NSCLC
|
non-small cell lung cancer
|
|
SCLC
|
small-cell lung cancer
|
|
HIV
|
human immunodeficiency virus
|
|
EMT
|
epithelial-mesenchymal transition
|
|
EGFR
|
epidermal growth factor receptor
|
|
ALK
|
anaplastic lymphoma kinase
|
|
KRAS
|
Kirsten rat sarcoma viral oncogene
homolog
|
|
ROS
|
tyrosine-protein kinase
|
|
MMP
|
matrix metalloproteinase
|
|
DVL
|
dishevelled homolog
|
|
TCF
|
transcription factor
|
|
LRP
|
low-density lipoprotein
receptor-related protein
|
|
Gsk
|
glycogen synthase kinase
|
|
APC
|
adenomatous polyposis coli
|
References
|
1
|
DeSantis CE, Miller KD, Goding Sauer A,
Jemal A and Siegel RL: Cancer statistics for African Americans,
2019. CA Cancer J Clin. 69:211–233. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Li J, Guo W, Ran J, Tang R, Lin H, Chen X,
Ning B, Li J, Zhou Y, Chen LC, et al: Five-year lung cancer
mortality risk analysis and topography in Xuan Wei: A
spatiotemporal correlation analysis. BMC Public Health. 19:1732019.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barta JA, Powell CA and Wisnivesky JP:
Global epidemiology of lung cancer. Ann Glob Health. 85:82019.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nedović-Vuković M, Laušević D, Ljaljević
A, Golubović M and Trajković G: Lung cancer mortality in
Montenegro, 1990 to 2015. Croat Med J. 60:26–32. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Haas K, Brillante C, Sharp L, Elzokaky AK,
Pasquinelli M, Feldman L, Kovitz KL and Joo M: Lung cancer
screening: assessment of health literacy and readability of online
educational resources. BMC Public Health. 18:13562018. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oze I, Ito H, Nishino Y, Hattori M,
Nakayama T, Miyashiro I, Matsuo K and Ito Y: Trends in small-cell
lung cancer survival in 1993–2006 based on population-based cancer
registry data in Japan. J Epidemiol. 29:347–353. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Riveiro-Falkenbach E, Ruano Y,
Garcia-Martin RM, Lora D, Cifdaloz M, Acquadro F, Ballestín C,
Ortiz-Romero PL, Soengas MS and Rodríguez-Peralto JL: DEK oncogene
is overexpressed during melanoma progression. Pigment Cell Melanoma
Res. 30:194–202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang Y, Gao M, Lin Z, Chen L, Jin Y, Zhu
G, Wang Y and Jin T: DEK promoted EMT and angiogenesis through
regulating PI3K/AKT/mTOR pathway in triple-negative breast cancer.
Oncotarget. 8:98708–98722. 2017.PubMed/NCBI
|
|
9
|
Lin D, Dong X, Wang K, Wyatt AW, Crea F,
Xue H, Wang Y, Wu R, Bell RH, Haegert A, et al: Identification of
DEK as a potential therapeutic target for neuroendocrine prostate
cancer. Oncotarget. 6:1806–1820. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Waldmann T, Scholten I, Kappes F, Hu HG
and Knippers R: The DEK protein-an abundant and ubiquitous
constituent of mammalian chromatin. Gene. 343:1–9. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Fu GK, Grosveld G and Markovitz DM: DEK,
an autoantigen involved in a chromosomal translocation in acute
myelogenous leukemia, binds to the HIV-2 enhancer. Proc Natl Acad
Sci USA. 94:1811–1815. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kappes F, Scholten I, Richter N, Gruss C
and Waldmann T: Functional domains of the ubiquitous chromatin
protein DEK. Mol Cell Biol. 24:6000–6010. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Matrka MC, Watanabe M, Muraleedharan R,
Lambert PF, Lane AN, Romick-Rosendale LE and Wells SI:
Overexpression of the human DEK oncogene reprograms cellular
metabolism and promotes glycolysis. PLoS One. 12:e01779522017.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Böhm F, Kappes F, Scholten I, Richter N,
Matsuo H, Knippers R and Waldmann T: The SAF-box domain of
chromatin protein DEK. Nucleic Acids Res. 33:1101–1110. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kappes F, Burger K, Baack M, Fackelmayer
FO and Gruss C: Subcellular localization of the human
proto-oncogene protein DEK. J Biol Chem. 276:26317–26323. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Waldmann T, Eckerich C, Baack M and Gruss
C: The ubiquitous chromatin protein DEK alters the structure of DNA
by introducing positive supercoils. J Biol Chem. 277:24988–24994.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sandén C and Gullberg U: The DEK
oncoprotein and its emerging roles in gene regulation. Leukemia.
29:1632–1636. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Xu Y, Liang Z, Li C, Yang Z and Chen L:
LCMR1 interacts with DEK to suppress apoptosis in lung cancer
cells. Mol Med Rep. 16:4159–4164. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boer J, Mahmoud H, Raimondi S, Grosveld G
and Krance R: Loss of the DEK-CAN fusion transcript in a child with
t(6;9) acute myeloid leukemia following chemotherapy and allogeneic
bone marrow transplantation. Leukemia. 11:299–300. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou QC, Deng XF, Yang J, Jiang H, Qiao
MX, Liu HH, Qian Z, Hou LL and Hu HG: Oncogene DEK is highly
expressed in lung cancerous tissues and positively regulates cell
proliferation as well as invasion. Oncol Lett. 15:8573–8581.
2018.PubMed/NCBI
|
|
21
|
Ou Y, Xia R, Kong F, Zhang X, Yu S, Jiang
L, Zheng L and Lin L: Overexpression of DEK is an indicator of poor
prognosis in patients with gastric adenocarcinoma. Oncol Lett.
11:1823–1828. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu X, Zou L, Yao Q, Zhang Y, Gan L and
Tang L: Silencing DEK downregulates cervical cancer tumorigenesis
and metastasis via the DEK/p-Ser9-GSK-3β/p-Tyr216-GSK-3β/β-catenin
axis. Oncol Rep. 38:1035–1042. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liu K, Feng T, Liu J, Zhong M and Zhang S:
Silencing of the DEK gene induces apoptosis and senescence in CaSki
cervical carcinoma cells via the up-regulation of NF-κB p65. Biosci
Rep. 32:323–332. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chandrashekar DS, Bashel B, Balasubramanya
SAH, Creighton CJ, Ponce-Rodriguez I, Chakravarthi BVSK and
Varambally S: UALCAN: A portal for facilitating tumor subgroup gene
expression and survival analyses. Neoplasia. 19:649–658. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang J, Sun L, Yang M, Luo W, Gao Y, Liu
Z, Qiu X and Wang E: DEK depletion negatively regulates
Rho/ROCK/MLC pathway in non-small cell lung cancer. J Histochem
Cytochem. 61:510–521. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang Y, Lei L, Zheng YW, Zhang L, Li ZH,
Shen HY, Jiang GY, Zhang XP, Wang EH and Xu HT: Odd-skipped related
1 inhibits lung cancer proliferation and invasion by reducing Wnt
signaling through the suppression of SOX9 and β-catenin. Cancer
Sci. 109:1799–1810. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lei L, Wang Y, Zheng YW, Fei LR, Shen HY,
Li ZH, Huang WJ, Yu JH and Xu HT: Overexpression of Nemo-like
kinase promotes the proliferation and invasion of lung cancer cells
and indicates poor prognosis. Curr Cancer Drug Targets. 19:674–680.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Soekarman D, von Lindern M, van der Plas
DC, Selleri L, Bartram CR, Martiat P, Culligan D, Padua RA,
Hasper-Voogt KP, Hagemeijer A, et al: Dek-can rearrangement in
translocation (6;9)(p23;q34). Leukemia. 6:489–494. 1992.PubMed/NCBI
|
|
29
|
Capitano ML and Broxmeyer HE: A role for
intracellular and extracellular DEK in regulating hematopoiesis.
Curr Opin Hematol. 24:300–306. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Sandén C, Nilsson HJ and Gullberg U: The
DEK oncoprotein is upregulated by multiple leukemia-associated
fusion genes. Blood Cells Mol Dis. 54:284–285. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Diaz-Serrano A, Gella P, Jimenez E,
Zugazagoitia J and Paz-Ares Rodriguez L: Targeting EGFR in lung
cancer: Current standards and developments. Drugs. 78:893–911.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Jurišić V, Obradovic J, Pavlović S and
Djordjevic N: Epidermal growth factor receptor gene in
non-small-cell lung cancer: The importance of promoter polymorphism
investigation. Anal Cell Pathol (Amst). 2018:61921872018.PubMed/NCBI
|
|
33
|
Bernicker EH, Allen TC and Cagle PT:
Update on emerging biomarkers in lung cancer. J Thorac Dis. 11
(Suppl 1):S81–S88. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
van Neerven SM and Vermeulen L: The
interplay between intrinsic and extrinsic Wnt signaling in
controlling intestinal transformation. Differentiation. 108:17–23.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Schaefer KN and Peifer M: Wnt/Beta-catenin
signaling regulation and a role for biomolecular condensates. Dev
Cell. 48:429–444. 2019. View Article : Google Scholar : PubMed/NCBI
|