Introduction
According to statistics from the World Health
Organization, the mortality of lung cancer in 2018 was ~1.8 million
worldwide, accounting for 18.4% of total cancer deaths (1), resulting in lung cancer being one of
the deadliest malignant tumors worldwide. Non-small cell lung
cancer (NSCLC) accounts for ~80% of deaths from lung cancer
(2). In NSCLC cells, the mechanism
of cell death regulation is seriously unbalanced, leading to
uncontrolled cancer cell proliferation (3). Therefore, inducing cancer cell death
is the primary focus of clinical lung cancer treatment (4). Currently, the side effects and
toxicity of chemotherapy and radiotherapy have created a bottleneck
in clinical curative treatments (5). Therefore, there is an urgent need to
discover alternative reagents or novel strategies to treat lung
cancer.
Tripterygium wilfordii Hook F is a promising
antitumor Chinese herb with the ability to regulate disease by
promoting blood circulation, improving blood stasis and detoxifying
harmful metabolites (6). Triptolide
(TP), a lactone compound with a unique diterpene and triperoxide
structural form, is the major biologically active component
extracted from the root, stem, rhizome and leaf of Tripterygium
wilfordii Hook F (7). TP has
previously been applied to treat rheumatoid arthritis and nephritis
due to its immunosuppressive and anti-inflammatory actions
(8). Recently, studies have
revealed that TP effectively represses the growth of multiple tumor
cells by inhibiting cell proliferation, and inducing apoptosis both
in vivo and in vitro (9). The heterogeneity of tumors in
different organs makes it necessary to explore the unique
mechanisms of specific tumors for targeted therapy. Our previous
results showed that TP inhibits NSCLC cells by blocking the cell
cycle and inducing apoptosis (10),
and an isobaric tags for relative and absolute quantitation-based
proteomics analysis revealed that the ribosome biogenesis pathway
was significantly altered in TP-treated NSCLC cells (11).
Ribosome biogenesis has attracted increasing
attention in molecular research over the past decade due to its
beneficial role in controlling the translation of all proteins in
the cell, and thus governing cell growth and proliferation. The
nucleolus, a subnuclear unit of the nucleus, contains ribosomal
(r)DNA genes, which are transcribed by RNA polymerase I (Pol I),
resulting in a single transcript known as 47S pre-rRNA (12). 47S-pre-rRNA is initially processed
into 45S rRNA and further spliced into 18S rRNA (13). The upstream binding factor (UBF),
together with RNA Pol I, constitutes an important component of the
pre-initiation complex that participates in the initiation of
ribosome biogenesis (14). UBF
binds to the whole length of the rDNA transcript units and is
implicated in the control of the elongation process (15). When cells are subjected to stress,
the nucleolus acts as a sensor for cellular stress signals,
resulting in the recruitment of ribosomal proteins (RPs) to mouse
double minute 2 protein (MDM2), thereby disrupting the interaction
between p53 and MDM2 (16). As a
result, p53 is no longer degraded by MDM2-mediated ubiquitination,
which further induces cell cycle arrest or apoptosis (16). In addition, TP inhibits the activity
of RNA Pol I (17), suggesting that
TP may be involved in regulating ribosome biogenesis. However, the
underlying mechanism remains unclear. Thus, it was hypothesized
that perturbation of ribosome biogenesis may be related to
TP-induced apoptosis in NSCLC cells. The aim of the present study
was to determine the effect of TP on ribosome biogenesis in A549
cells, and reveal the molecular mechanisms mediating TP-induced
apoptosis and cell cycle arrest for improved utilization of TP in
lung cancer therapy.
Materials and methods
Cell culture
Human lung cancer A549 cells (cat. no. CCL-185™;
American Type Culture Collection) were cultured at 37°C in a
humidified atmosphere with 5% CO2 in Dulbecco's modified
Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.)
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.) and 1%
penicillin-streptomycin (Thermo Fisher Scientific, Inc.). The cell
line was authenticated using short tandem repeat profiling and
mycoplasmas were tested to confirm that the cells were free of
contamination.
Chemicals
TP (≥98%) and actinomycin D (Act D, 20 ng/ml) were
obtained from Sigma-Aldrich (Merck KGaA). These drugs were
aliquoted upon delivery, stored at 100 µg/ml in DMSO at −80°C, and
diluted to the indicated concentrations using serum-free culture
medium when used. Cells were treated with Act D for 36 h prior to
relevant experiments.
Cell cycle, apoptosis and
proliferation
Cells were treated with TP at 12.5, 50 and 100 ng/ml
for 36 h. Apoptosis, cell cycle and cell viability were detected
according to the manufacturer's instructions. For apoptosis,
5×105 cells were collected with EDTA-free trypsin,
washed with cold PBS, incubated with 5 µl Annexin V-FITC and 10 µl
propidium iodide (PI; 20 µg/ml) at room temperature for 15 min in
the dark, and analyzed by a FACS Canto II instrument (BD
Biosciences); the apoptotic rate was calculated as the sum of early
+ late apoptotic cells.
For cell cycle analysis, 5×105 cells were
fixed with absolute ethanol at −20°C overnight, washed with cold
PBS, incubated with 5 mg/ml RNase at 37°C for 30 min, stained with
5 mg/ml PI in the dark at room temperature for 30 min, and last
analyzed by a FACS Canto II. Flow cytometry data were analyzed with
FlowJo 7.6 software (FlowJo LLC).
For cell viability, 3×103 cells were
seeded in 96-well plates and cultured with 100 µl DMEM. Cell
Counting Kit-8 solution (Dojindo Molecular Technologies, Inc.) was
incubated with cells at 37°C for 3 h. Absorbance at 450 nm was
measured, and cell viability was evaluated according to the
manufacturer's instructions.
Reverse transcription-quantitative PCR
(RT-qPCR)
TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) was used to extract total cellular RNA. RT
was conducted using a PrimeScript RT reagent kit (Takara Bio, Inc.)
according to the manufacturer's instructions. Amplification of 45S
and 18S rRNA was performed using a C1000 Thermal Cycler detection
system (Bio-Rad Laboratories, Inc.) in triplicate using 1 ng of
cDNA, 125 nM forward and reverse primer and 25 µl 2X
SYBR® Premix Ex Taq™ (Takara Bio, Inc.) in a 50-µl
reaction. The reaction parameters were 95°C for 1 min, followed by
42 cycles of 95°C for 15 sec, 56°C for 25 sec and 72°C for 30 sec.
The following primers were used: 45S forward,
5′-GCGGAACCCTCGCTTCTC-3′ and reverse, 5′-CTCCGTTATGGTAGCGCTGC-3′;
18S forward, 5′-CTCTCCGGAATCGAACCCTGA-3′ and reverse,
5′-CGACGACCCATTCGAACGTCT-3′; and GAPDH forward,
5′-AGCCACATCGCTCAGAACAC-3′ and reverse,
5′-GAGGCATTGCTGATGATCTTG-3′. The relative mRNA expression levels
were quantified using the 2−∆∆Cq method (18); GAPDH was used as the internal
reference.
Immunoprecipitation (IP) assay
Cells were washed and scraped with cold PBS,
centrifuged at 16,000 × g at 4°C for 15 min, and suspended in IP
lysis buffer (Cell Signaling Technology, Inc.). Protein
concentrations were measured using a bicinchoninic acid (BCA)
protein assay kit (Thermo Fisher Scientific, Inc.). Protein (100
µg) was incubated with anti-MDM2 (1:100; cat. no. 86934; Cell
Signaling Technology, Inc.) or IgG (1:500; cat. no. sc-2025; Santa
Cruz Biotechnology, Inc.) antibodies mixed with 500 µl of TBS-0.5%
Tween 20 (TBST) along with 10 µl of Dynabeads protein G on a
rotating platform at 4°C overnight. Protein beads were then washed
three times with 0.6 ml of ice-cold TBST buffer. A micropipette was
used to remove the last traces of buffer. Antibody-protein
complexes were eluted with 1X SDS sample buffer, electrophoresed
and subjected to western blotting.
Western blot analysis
TP-treated cells were collected with Cell Lysis
Buffer (cat. no. 9803S; Cell Signaling Technology, Inc.)
supplemented with phenylmethanesulfonylfluoride fluoride (cat. no.
8553S; Cell Signaling Technology, Inc.) at the indicated times. The
BCA assay kit was used to determine the protein concentrations.
Proteins (30 µg) were denatured in boiling water for 10 min and
fractioned by using SDS-PAGE with 10% denaturing acrylamide gels.
The fractioned proteins were transferred to polyvinylidene fluoride
membranes. The blotted membranes were blocked with 5% nonfat milk
in TBST for 1 h at room temperature, followed by incubation with
the diluted primary antibodies against MDM2 (1:1,000; cat. no.
86934, Cell Signaling Technology, Inc), phosphorylated (p)-p53
(1:1,000; cat. no. 2521; Cell Signaling Technology, Inc), cleaved
(c)-caspase 9 (1:1,000; cat. no. 20750; Cell Signaling Technology,
Inc), c-caspase 3 (1:1,000; cat. no. 9664; Cell Signaling
Technology, Inc), GAPDH (1:1,000; cat. no. 5174; Cell Signaling
Technology, Inc.), RPL23 (1:3,000, cat. no. ab241088; Abcam), p53
(1:5,000; cat. no. 10442-1-AP; ProteinTech Group, Inc.), p53
upregulated modulator of apoptosis (1:1,000; PUMA; cat. no.
55120-1-AP; ProteinTech Group, Inc.) and BCL2 (1:1,000; cat. no.
12789-1-AP; ProteinTech Group, Inc.) at 4°C for overnight. The
membranes were then incubated with horseradish
peroxidase-conjugated secondary antibody (1:2,000; cat. nos. 7076
and 7074; Cell Signaling Technology, Inc.) for 1.5 h at room
temperature. Enhanced chemiluminescence reagents (EMD Millipore)
were used to visualize the immunoreactive proteins, and
densitometric analysis was performed by using ImageJ v1.8.0
software (National Institutes Of Health).
Immunofluorescence assay
Cells were seeded into confocal microscopy dishes,
treated with 50 ng/ml TP for 24 h and fixed in 4% paraformaldehyde
at room temperature for 20 min. Next, 0.2% Triton X-100 was used to
permeabilize the membranes. The prepared cells were washed with
cold PBS and blocked with 3% bovine serum albumin (BSA;
Sigma-Aldrich; Merck KGaA) at room temperature for 30 min. Primary
antibodies against nucleophosmin (B23; 1:200; cat. no. ab15440;
Abcam; or 1:100; cat. no. 10306-1-AP; ProteinTech Group, Inc.), RNA
Pol I (1:100; cat. no. sc-48385; Santa Cruz Biotechnology, Inc.),
UBF (1:100; cat. no. sc-13125; Santa Cruz Biotechnology, Inc.) and
nucleolin (NCL; 1:1,000; cat. no. 14574; Cell Signaling Technology,
Inc.) were added and incubated at 4°C overnight. Dylight™ 488
(green) or 594 (red)-conjugated secondary antibodies (1:500; cat.
nos. A23210 and A23420; Abbkine Scientific Co., Ltd.) were applied
for 1 h at room temperature. DAPI (Thermo Fisher Scientific, Inc.)
was used to counterstain the nuclei. Immunofluorescent images of
the prepared cells were viewed under a fluorescence laser-scanning
confocal microscope (magnification, ×63; Carl Zeiss AG) in 20
randomly selected fields.
Incorporation of 5-fluorouridine
(5-FU) into rRNA
Cells were seeded on coverslips and incubated with
DMEM for adherence. Then, 50 ng/ml TP was used to stimulate the
cells for 24 h. Next, 2 mM 5-FU (Sigma-Aldrich; Merck KGaA) was
incubated with the cells growing on coverslips at room temperature
for 15 min. The processes of washing, fixation and membrane
permeabilization were performed as described in the
Immunofluorescence assay section. Specific monoclonal antibodies
recognizing halogenated uridine (1:400; cat. no. B8434; Merck KGaA)
and NCL (cat. no. 14574; Cell Signaling Technology, Inc.) were used
to incubate with the prepared cells at 4°C overnight.
Dylight-conjugated secondary antibody incubation, nuclear
counterstaining and immunofluorescent imaging were conducted as
previously described.
Chromatin IP(ChIP) analysis
ChIP assays were performed according to the
manufacturer's instructions (EZ ChIP™ kit; cat. no. 17-371; EMD
Millipore). Briefly, the prepared cells were cross-linked with 1%
formaldehyde for 10 min at room temperature. The unreacted
formaldehyde was quenched using 0.125 M glycine. The cells were
washed with cooled PBS containing protease inhibitor cocktail II
included in the kit for 5 min, and then scraped and pelleted. The
cell pellets were lysed in 1% SDS lysis buffer. Sonication (pulse
for 5 sec, pause for 9 sec, cycle 23 times) on wet ice (0°C) was
conducted for DNA fragmentation. An aliquot of the sheared lysate
was preserved at 4°C until use as an input control. Sheared lysate
(100 µl) was diluted with dilution buffer and incubated with
anti-UBF (1:50; cat. no. sc-13125; Santa Cruz Biotechnology, Inc.)
and anti-RNA Pol I antibodies (1:50; cat. no. sc-48385; Santa Cruz
Biotechnology, Inc.) on a rotating rack at 4°C for overnight. The
immunocomplexes containing DNA, protein and antibodies were
captured using protein A-agarose. Immunocomplexes conjugated with
beads were eluted using elution buffer including 1% SDS and 100 mM
NaHCO3 and reverse cross-linked at 65°C overnight. DNA
fragments were collected, and UBF and RNA Pol I enrichment in the
rDNA promoter were amplified using RT-qPCR as previously described.
The primers for augmenting the rRNA promoter DNA were as follows:
Forward, 5′-CGATGGTGGCGTTTTTGG-3′ and reverse,
5′-CCGACTCGGAGCGAAAGATA-3′. The negative control comprised the same
assay conducted using a human normal IgG antibody (cat. no.
sc-2025; Santa Cruz Biotechnology, Inc.
Animal studies
A549 cells (5×106) were injected
subcutaneously into 10 4-week-old male BALB/cAnNCr-nu/nu nude mice
(18–22 g; Sippr-BK Laboratory Animal Co., Ltd.). All mice were fed
with standard mouse irradiated food and tap water ad
libitum, and maintained under conditions of 25°C and 50%
humidity with a 12-h light/dark cycle. After 3 weeks, the mice were
randomly divided into two groups of five, receiving subcutaneous TP
(1.5 mg/kg) or saline (mock) injections every day for 5 weeks. The
body weight and tumor volume were measured every 2 days. The tumor
volume was calculated as (length × width × width)/2. Then, all mice
were euthanized using carbon dioxide in according with the
Guidelines for Euthanasia of Rodents Using Carbon Dioxide issued by
the National Institutes of Health. After euthanasia, death was
further verified by cardiac arrest and cadaveric rigidity. The
animal experiments were approved by the Animal Experimentation
Ethics Committee of the Tongde Hospital of Zhejiang Province.
Terminal
deoxynucleotidyl-transferase-mediated dUTP nick end labeling
(TUNEL) assay
TUNEL assays were performed to investigate the
apoptotic cells in tumor tissues using an in situ cell death
detection kit obtained from Roche Diagnostics. Tumor tissue was
fixed with a mixture of 70% alcohol, formaldehyde and glacial
acetic acid (16:1:1) at 4°C for 24 h, and paraffin-embedded tumor
sections (5-µm) were dewaxed and rehydrated. Proteinase K was
applied at 37°C for 25 min. Membranes were dissolved at room
temperature for 20 min. Then, TdT and dUTP were mixed at 1:9 and
the mixture incubated with tissues at 37°C for 2 h. DAPI solution
was used to stain nuclei in the dark at room temperature for 10
min. A fluorescence microscope was used to observe and photograph
the apoptotic cells. Positive cells were marked by green
fluorescence. TUNEL-positive cells and total cells in sections were
measured using ImageJ v1.8.0 software.
Immunohistochemistry
The tissue samples (5 µm) were deparaffinized and
rehydrated. A timer-controlled water bath was used to conduct
heat-induced antigen retrieval at 98°C for 20 min. TBS plus 0.025%
Triton X-100 was used to wash the slides with gentle agitation. The
slides were then blocked with 10% BSA in TBS at room temperature
for 2 h. Primary antibodies against p53 (1:150; cat. no. 2527; Cell
Signaling Technology, Inc.), RPL23 (1:50; cat. no. 16086-1-AP;
ProteinTech Group, Inc.) and NCL (1:400; cat. no. 14574; Cell
Signaling Technology, Inc.) were incubated with the samples
overnight at 4°C, and 50 µl horseradish peroxidase-conjugated
secondary antibody (cat. no. 8114S; Cell Signaling Technology,
Inc.) was used to recognize the primary antibody at room
temperature for 1 h. The tissues were then stained with
3,3′-diaminobenzidine and subsequently hematoxylin at room
temperature for 4 min, and the samples were observed under a light
microscope (magnification, ×40) in 20 fields of vision and
quantified by ImageJ v1.8.0 software.
Statistical analyses
Experiments were performed in triplicate at least
three times, independently. Data are presented as the mean ±
standard deviation. One-way, two-way or mixed ANOVA followed by
Tukey's, Dunnett's or Sidak's multiple comparisons test, or
Student's t-test in GraphPad Prism 6 (GraphPad Software, Inc.) were
used to analyze the statistical significance between groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
TP induces apoptosis and cell cycle
arrest in A549 cells
The effects of TP were first determined on A549
cells. An Annexin V/PI staining-based assay with
fluorescence-activated cell sorting was performed to analyze the
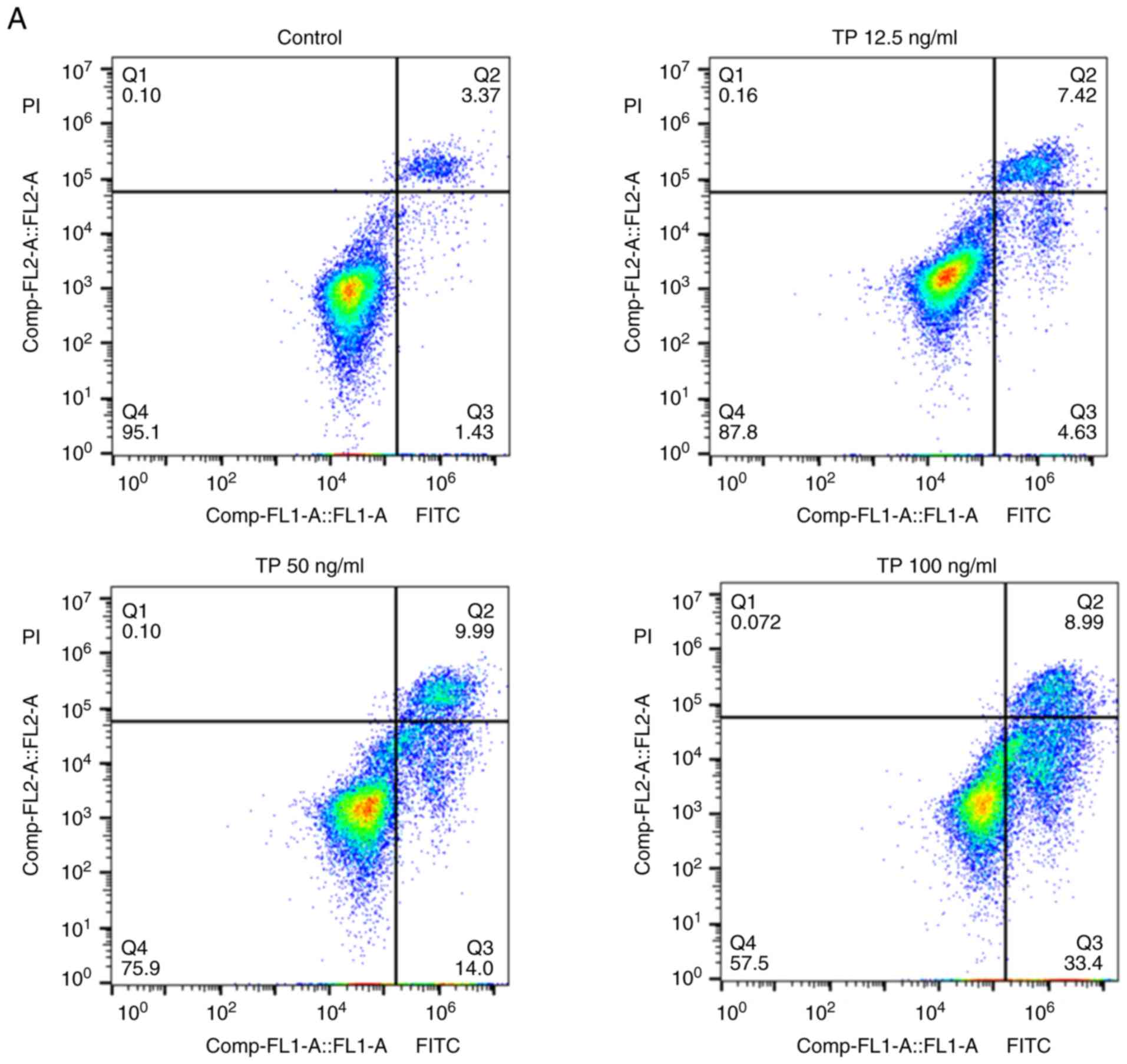
population of apoptotic cells. The results showed that TP increased
the apoptotic index in a dose-dependent manner (Fig. 1A). The percentage of apoptotic cells
gradually increased with increasing TP concentration, from
4.1±1.34% in control cells to 12.3±1.72, 26.2±2.16 and 45.8±3.28%
in TP-treated cells (Fig. 1C).
Then, PI staining was performed to analyze the cell cycle (Fig. 1B). TP caused the G0/G1 fraction to
decrease from 48.6±3.56% in control cells to 14.9±1.22% in 100
ng/ml TP-treated cells, and the G2/M fraction increased from
14.2±2.42% in the control to 19.2±2.68% (12.5 ng/ml TP), 25.2±3.11%
(50 ng/ml TP) and 38.9±3.62% (100 ng/ml TP; Fig. 1D). These results indicated that TP
arrested A549 cells at the G2/M phase. Then, the effect of TP on
cell viability was evaluated. As presented in Fig. 1E, TP induced a marked dose-dependent
reduction in cell viability; increasing the duration of treatment
from 24 h to 48 h significantly enhanced this inhibition.
TP induces nucleolar RNA Pol I and UBF
translocation and nucleolar disintegration
The expression and distribution of Pol I in the
nucleus was investigated. Cells were treated with a negative
control, TP, or a positive control, Act D, for 36 h. Act D is a
well-characterized nucleolar disruptor (19). The prepared cells were dually
stained with antibodies against the RNA Pol I antigen and B23; B23
is associated with ribosome biogenesis and nucleolar
ribonucleoproteins (20). As
presented in Fig. 2A, endogenous
RNA Pol I was present primarily in the nucleolus in the control
cells. Treatment with TP and Act D resulted in abnormal nuclear
morphology and nucleolar disintegration, and faint traces of RNA
Pol I were observed in TP-treated cells. For UBF, robust protein
expression was observed in almost all nucleoli in the negative
control cells; TP and Act D induced nucleolar disintegration and
further diffused the distribution of UBF throughout the nucleus
rather than being focused in the nucleolus (Fig. 2B). The RNA Pol I and UBF relative
fluorescent intensities were significantly reduced in the
TP-treated cells.
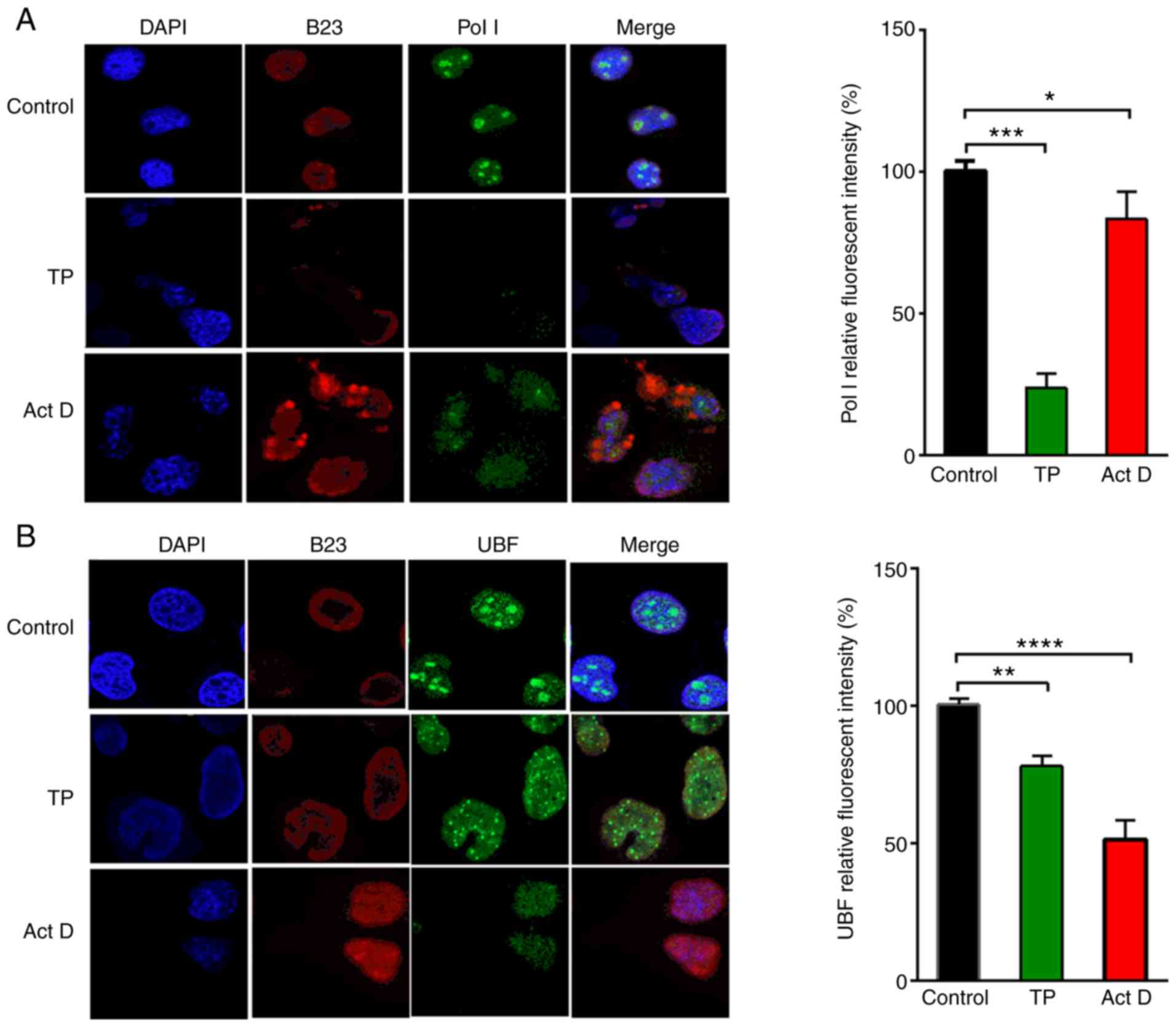 | Figure 2.TP induces nucleolus disintegration
and suppresses ribosomal DNA transcription factors. A549 cells
(2×104) were seeded on coverslips were treated with TP
(50 ng/ml), Act D (20 ng/ml) or DMSO for 24 h, fixed and stained
with (A) anti-Pol I or (B) anti-UBF antibodies; B23 was used to
stain the nucleoli. Data are presented as the mean ± SD of three
independent experiments. Pictures were captured under a confocal
microscope (magnification, ×63). Data were analyzed using one-way
ANOVA combined with Dunnett's multiple comparisons test.
*P<0.05, **P<0.01, ***P<0.001, ****P<0.0001. TP,
triptolide; Act D, actinomycin D; B23, nucleophosmin; Pol I, RNA
polymerase I; UBF, upstream binding factor. |
TP interrupts rRNA synthesis and rRNA
processing
Burger et al (21) demonstrated that the disintegration
of nucleolar structures is related to rRNA synthesis. Therefore,
immunofluorescence assays were performed to detect whether TP plays
a role in the process of RNA synthesis. NCL, a multifunctional
protein that mainly regulates ribosome biogenesis and
transcription, was used as a control to evaluate the integrity of
the nucleoli. The results in Fig.
3A revealed that TP treatment significantly reduced the
incorporation of 5-FU into nascent rRNA, suggesting that TP
treatment inhibited rRNA synthesis. The ChIP assay demonstrated
that TP treatment caused significant dissociation of Pol I and UBF
from the promoter region of the rDNA in a dose-dependent manner
(Fig. 3B and C); these results were
consistent with the decreased localization of RNA Pol I and UBF
observed in Fig. 2A and B. 45S rRNA
is the transcriptional product driven by Pol I and UBF, and
maturation of the primary transcript leads to the generation of 18S
rRNA (16). RT-qPCR analysis was
performed to determine the effect of TP on primary rRNA transcript
45S and modified 18S. The results in Fig. 3D showed that TP significantly
reduced the expression of 45S pre-rRNA in a dose-dependent manner,
and the expression of 18S expression was also significantly
impaired. This result indicated that TP inhibited rRNA synthesis by
inhibiting Pol I and UBF, thus further interfering with rRNA
processing.
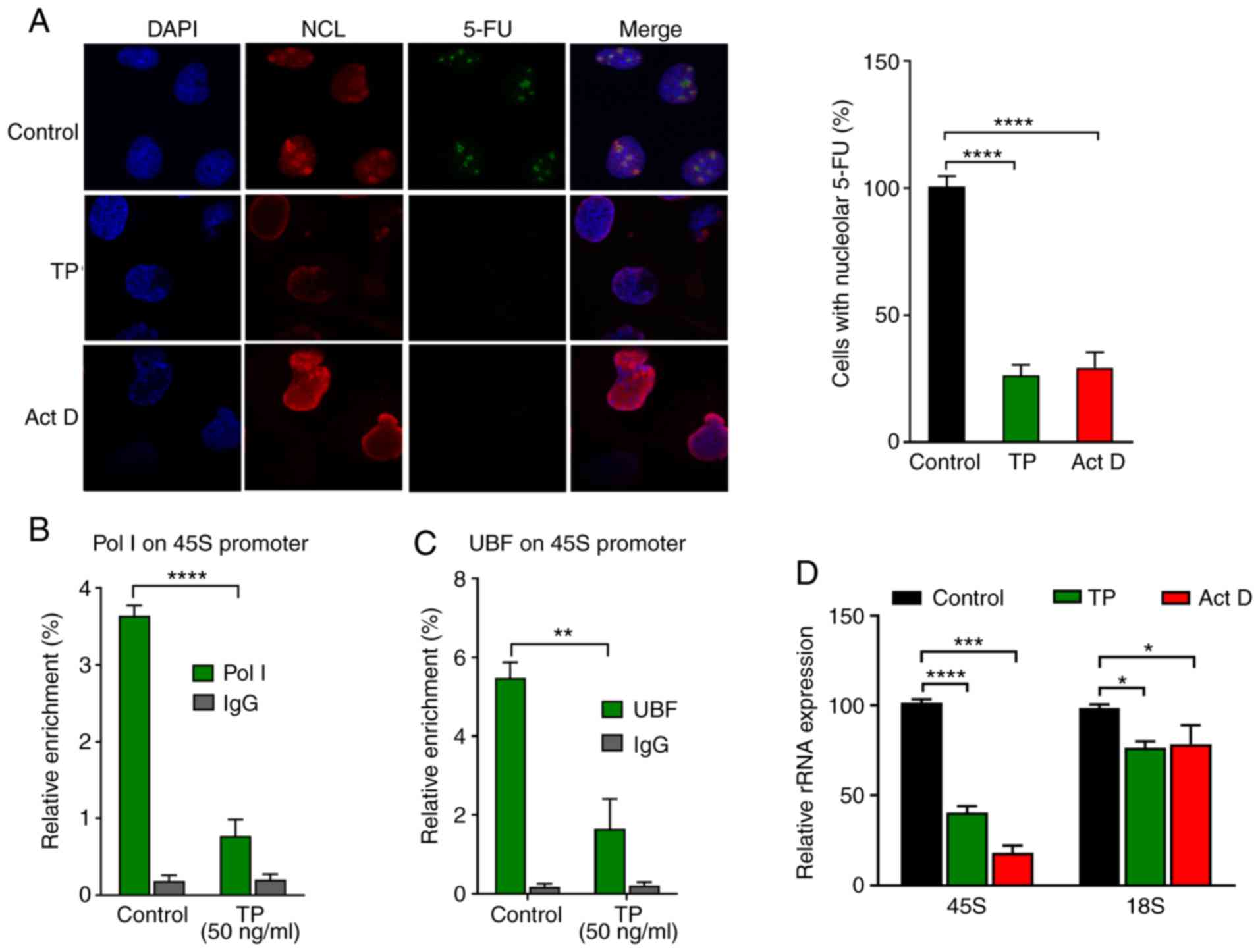 | Figure 3.TP reduces rRNA synthesis and
suppresses rRNA processing. (A) A549 (2×104) cells were
seeded on coverslips and first treated with TP (50 ng/ml) or Act D
(20 ng/ml) for 24 h; newly synthesized RNA was labeled with 5-FU
for 15 min before the cells were harvested, and Alexa
Fluor-conjugated monoclonal antibodies were used to mark
5-FU-labeled RNA. NCL was used to stain the nucleoli, and DAPI was
used to counterstain the nuclei. Images were captured under a
confocal microscope (magnification, ×63). A549 Cells
(1×107) treated with TP (50 ng/ml, 24 h) were subjected
to chromatin immunoprecipitation assays for (B) Pol I and (C) UBF
occupancy of the promoter region of 45S rRNA. (D) Reverse
transcription-quantitative PCR analysis was used to evaluate 45S
and 18S expression. Data are presented as the mean ± SD of at least
three independent experiments, and were analyzed using one-way
ANOVA combined with Dunnett's multiple comparisons test or two-way
ANOVA combined with Sidak's multiple comparisons test. *P<0.05,
**P<0.01, ***P<0.001, ****P<0.0001. TP, triptolide; Act D,
actinomycin D; r, ribosomal; NCL, nucleolin; Pol I, RNA polymerase
I; UBF, upstream binding factor; 5-FU, 5-fluorouridine. |
TP induces apoptosis and cell cycle
arrest by increasing the binding of RPL23 to MDM2
When ribosome biogenesis is interrupted, RPs may not
be used for ribosome biogenesis and can bind to MDM2, which
inhibits MDM2 from mediating p53 degradation (22,23).
To determine whether there are intrinsic links between TP-induced
ribosome biogenesis disorder and cell survival inhibition, A549
cells were treated with TP or a positive control, and co-IP assays
were carried out with an anti-MDM2 antibody to pull down the
associated proteins. The results showed that TP treatment increased
the binding of RPL23 to MDM2 and reduced the binding of p53 to MDM2
(Fig. 4A). These results suggested
that there may be competitive binding of RPL23 and p53 to MDM2. The
increased binding of RPL23 to MDM2 led to reduced binding of p53,
resulting in decreased MDM2-mediated degradation of p53.
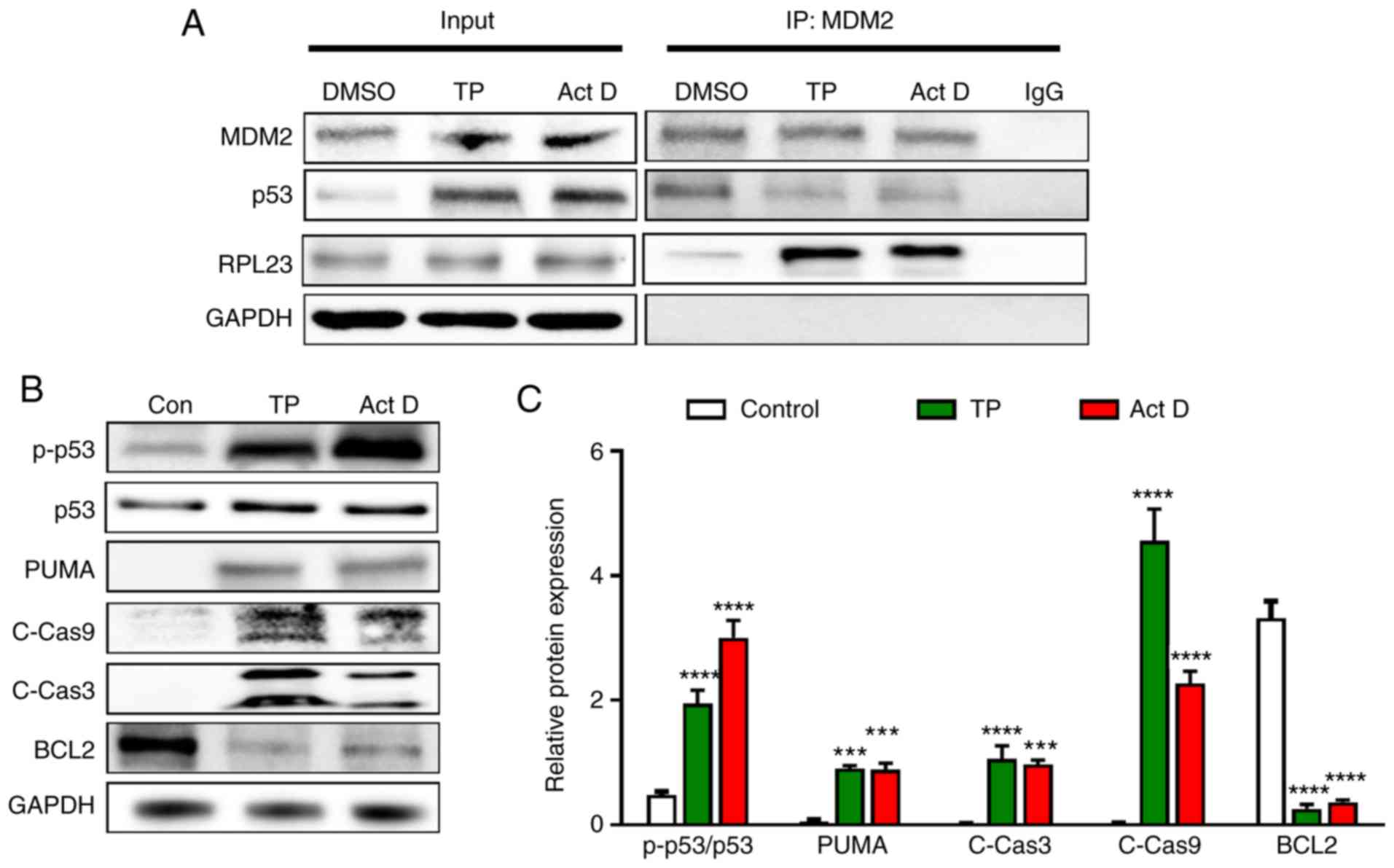 | Figure 4.TP increases the binding of RPL23
with MDM2 and activates p53 and p53-regulated proteins. (A) Cell
lysates of TP-treated cells (5×106) were
immunoprecipitated with anti-MDM2 antibodies, followed by
immunoblotting with anti-MDM2, p53 and RPL23 antibodies. For each
lysate, 20% of the quantity used for IP was loaded as an input
control. (B) Cell lysates from 5×106 A549 cells
receiving various treatments for 48 h were subjected to western
blot analysis with the indicated antibodies; GAPDH was used as an
internal control. (C) Densitometric analysis of protein expression.
Data are presented as the mean ± SD of at least three independent
experiments. Data were analyzed using one-way ANOVA combined with
Tukey's multiple comparisons test. ***P<0.001, ****P<0.0001
vs. control. TP, triptolide; MDM2, mouse double minute 2; RPL23,
ribosomal protein L23; PUMA, p53-upregulated modulator of
apoptosis, C-Cas, cleaved caspase; p, phosphorylated; IP,
immunoprecipitation. |
p53 stabilization can evoke signaling events
involved in cell survival, such as apoptotic and cell cycle
pathways (24). As shown in
Fig. 4B and C, TP treatment induced
increases in p-p53, and activation of caspase 9 and caspase 3.
Reduced levels of BCL2 were also induced by TP and the positive
control. These results suggested that TP induced apoptosis and cell
cycle arrest via p53 activation.
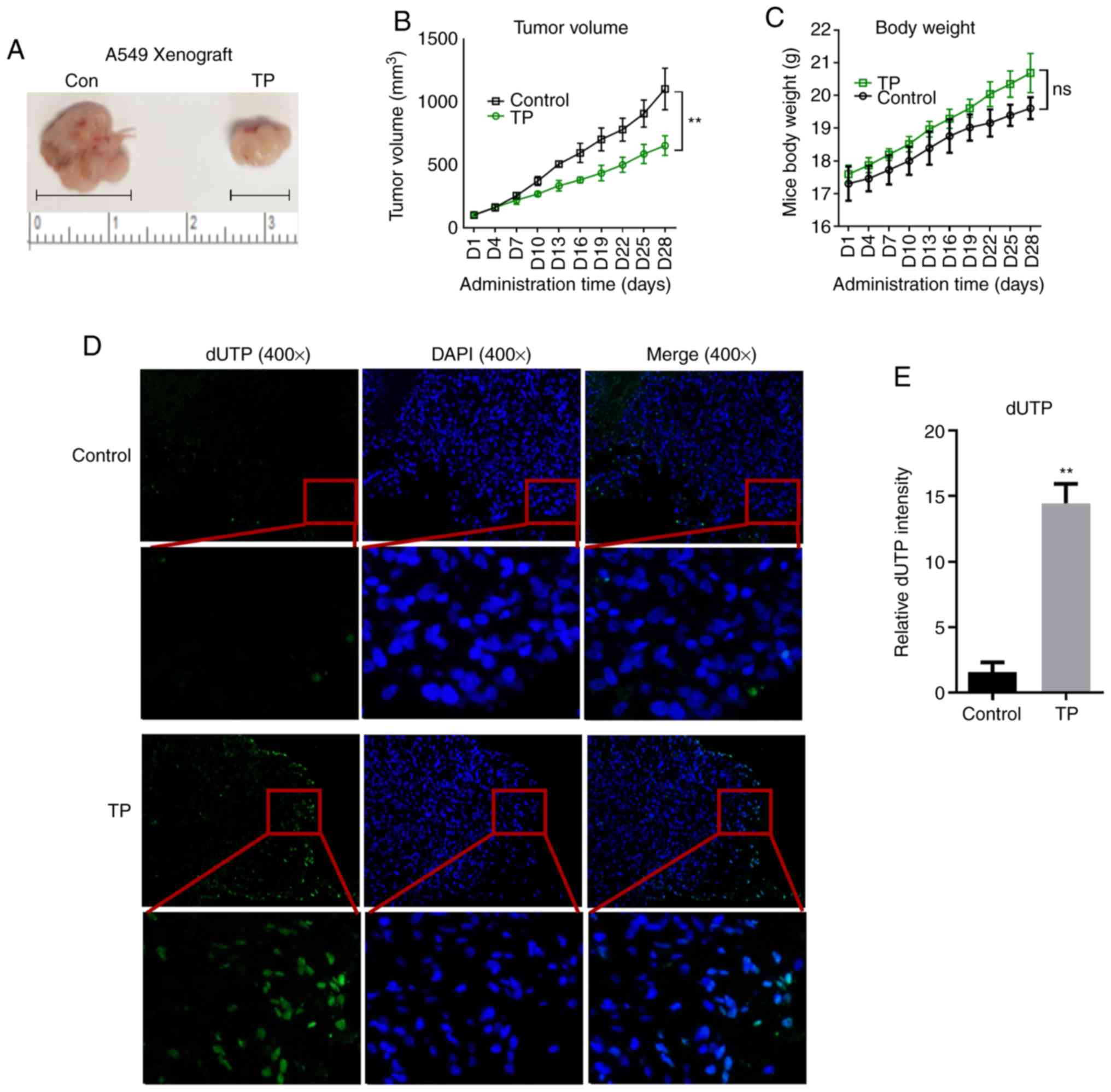
TP suppresses tumor growth by inducing
ribosomal stress in vivo
To further validate the effects of TP on A549 cell
survival, A549 ×enografts were implanted in nude mice. BALB/c mice
were injected subcutaneously on their right limb. At 3 weeks later,
the mice were divided randomly into two groups of five mice. Each
group was treated with saline or TP once a day for 4 weeks. The
results showed that TP treatment notably inhibited tumor growth
from day 13 (Fig. 5A and B).
Moreover, the body weights of the TP-treated mice were
significantly higher than that of the control group (Fig. 5C). TP treatment induced enhanced
expression of dUTP, which binds mainly to the terminal 3-OH of
fragmented DNA, indicating that apoptosis in TP-treated xenografts
was markedly activated (Fig. 5D and
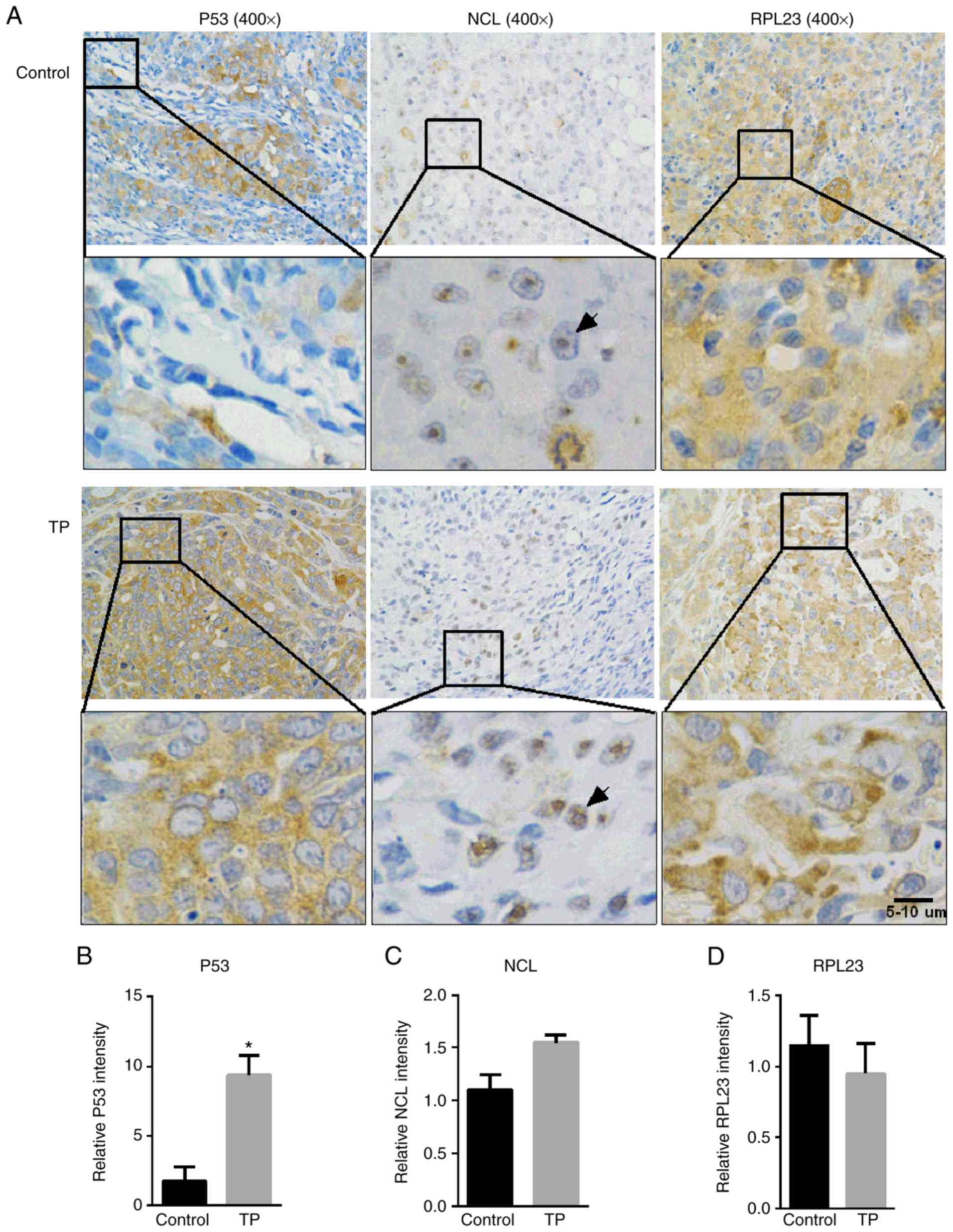
E). Consistent with the in vitro experiments,
immunohistochemistry revealed robust p53 expression in response to
TP administration (Fig. 6A and B).
In the control xenografts, NCL was found predominantly in the
nucleolus; however, TP treatment caused it to disperse to the
nucleus (Fig. 6A), suggesting that
the nucleolus was disrupted. NCL and RPL23 exhibited nearly the
same expression intensity between the two groups (Fig. 6C and D); however, TP treatment
resulted in RPL23 being more likely to be located at the edge of
the cell membrane (Fig. 6A).
Discussion
The nucleolus is a pressure sensor that contains
various proteins involved in ribosome synthesis (25). B23, NCL, RNA Pol I and UBF are the
main sensory proteins located in the nucleoli (16). In the present study, it was found
that B23, NCL, RNA Pol I and UBF were located primarily in the
nucleolus in control-treated NSCLC cells, whereas in the TP-treated
cells, the nucleolar structure disappeared, and B23, NCL, RNA Pol I
and UBF were translocated and dispersed throughout the nucleus.
Downregulation of NCL is involved in cell cycle arrest, growth
inhibition and apoptosis (26).
Inhibition of B23 in atypical teratoid or rhabdoid tumors arrests
the cell cycle at G1 phase (27),
suggesting that reduced expression and abnormal distribution of NCL
and B23 induced by TP are implicated in cell cycle arrest,
apoptosis and growth inhibition. These results are consistent with
previous findings that ribosomal stress caused by acrolein induced
nucleolar disintegration and nucleolus-related molecule
translocation (16).
The abnormal distribution and expression of sensory
proteins inevitably leads to abnormal ribosomal rRNA synthesis
(28,29). rRNA synthesis is the first step of
ribosome biogenesis in the nucleolus. RNA Pol I and UBF are
essential members of the pre-initiation complex that promotes the
initiation of rDNA promoter transcription (30). Disrupting the binding ability of UBF
and RNA Pol I to the rDNA promoter decreases rDNA transcription and
rRNA synthesis (31). Previous
studies have reported that TP is a known inhibitor of RNA
polymerase I; therefore, it was hypothesized that the inhibition of
rRNA synthesis by TP in the present study was a result of
suppression of RNA Pol I. It was shown that TP reduced UBF1
expression in the nucleoli and reduced UBF1 binding to rDNA, which
exacerbated the decrease in rRNA synthesis. In addition, the
TP-mediated reduction in the expression of precursor 45S rRNA also
significantly reduced the level of 18S rRNA, which is a processed
product from precursor 45S rRNA. 18S rRNA participates in the
biogenesis of the 60S ribosome subunit (32), suggesting that TP is involved in
disruption of the 60S ribosome subunit. This is consistent with our
previous proteomics analysis showing that TP treatment
significantly downregulated proteins involved in ribosome
processing (11).
Disruption of ribosome biogenesis evokes signaling
pathways that lead to cell cycle arrest, cellular senescence or
apoptosis (16). The recruitment
and binding of RPs to MDM2 increases significantly after ribosome
biogenesis is disrupted; therefore, p53 is freed from MDM2 and no
longer degraded by MDM2-mediated ubiquitination, leading to p53
activation (33). In the present
study, it was found that ribosomal stress induced by TP cascaded
via disintegrating the ribosomal structure, inhibiting rRNA
biogenesis and freeing RPL23. Consequently, the released RPL23
robustly and competitively combines with MDM2 to cause the
dissociation and activation of p53, consistent with previous
observations that RPL23 activates p53 in response to ribosomal
perturbation (23). Activated p53
further causes cleavage of caspase 9, which is implicated in the
activation of caspase cascades by cleaving and activating caspase 3
(16). In the present study,
increased levels of both c-caspase 3 and c-caspase 9 were observed
following TP treatment. The increase in p53 phosphorylation was
associated with an increase in PUMA, which is expressed at very low
levels under normal conditions (34,35).
The increase in PUMA levels further promoted cellular apoptosis via
feedback interactions with p53 (36). In addition, the reduction in BCL2
levels suggested that the apoptosis induced by TP is
mitochondria-dependent, consistent with a previous report that TP
treatment activated the mitochondrial apoptosis pathway (10).
Disturbances in ribosome synthesis can lead to cell
cycle arrest through the retinoblastoma protein pathway and
p53-dependent pathway (37,38). In the present study, it was found
that TP induced disturbances in ribosome synthesis, suggesting that
cell cycle arrest in lung cancer cells induced by TP could be
attributed, at least in part, to disturbances in ribosome
synthesis. Protein synthesis by ribosomes is a major metabolic
event that regulates proliferation and cellular growth. TP-induced
inhibition of rRNA synthesis may result in abnormal lung cancer
cell proliferation, consistent with reports that ribosome
heterogeneity impairs global protein synthesis and normal growth
(39). The MDM2-p53 axis is a key
signaling pathway that regulates the tumor cell cycle,
proliferation and apoptosis (40).
Downregulation of MDM2 and p53 ubiquitination promotes the
proliferation and cisplatin chemoresistance of cancer cells
(41). Zhou et al (42) reported that activating the
RPL40-MDM2-p53 pathway induced cell cycle arrest and apoptosis. In
the present study, it was found that TP inhibited cell cycle,
induced apoptosis and activated the p53-MDM2 pathway, indicating
that the inhibitory effect of TP on lung cancer cells may be
related to the activation of the p53-MDM2 pathway.
Numerous studies have reported that ribosome
synthesis disorder induces the suppression of cancers (22,43).
TP has also been shown to markedly induce lung cancer cell
apoptosis via multiple mechanisms (8,10);
however, the mechanism by which TP inhibits lung cancer by inducing
disordered ribosomes has not been reported. The present findings
are the first, to our knowledge, to show that TP induced apoptosis,
cell cycle arrest, and suppression of cell proliferation and tumor
growth via nucleolar disintegration and rRNA synthesis inhibition,
potentially via the ribosome-RPL23-MDM2-p53 signaling pathway
(Fig. 7). Additional research into
the mechanisms of TP's effects on ribosomal biosynthesis is
required; however, these results could lead to the development of
anticancer therapeutic strategies involving the use of TP.
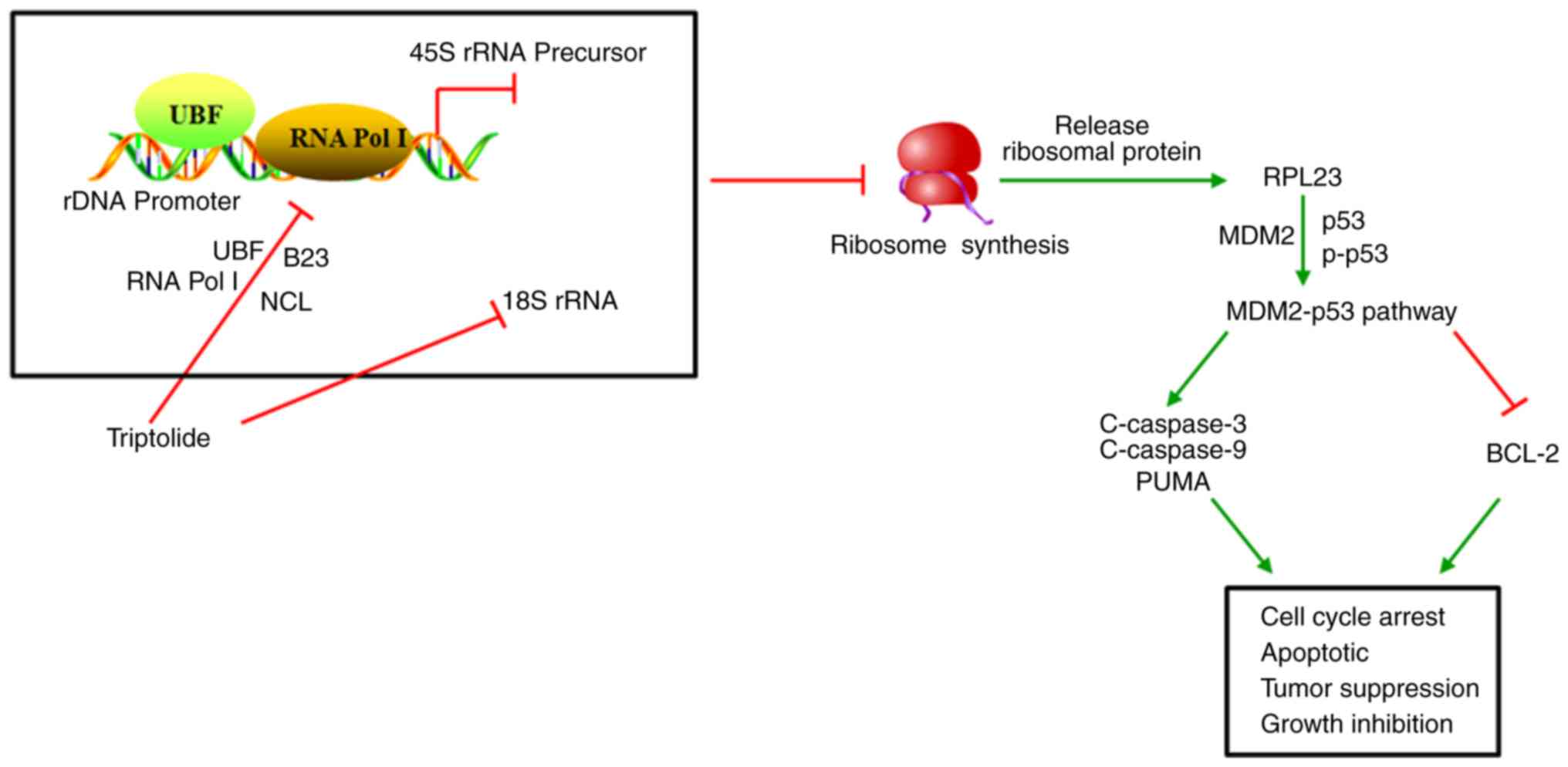 | Figure 7.Schematic diagram of the mechanism of
TP-induced lung cancer cell inhibition via ribosomal stress
initiation. TP induces nucleolus degradation and inhibits 45S rRNA
synthesis by reducing the binding of Pol I and UBF to the rDNA
promoter. The 18S rRNA degraded by TP disrupts ribosomal synthesis,
resulting in the liberation of RPL23. The combination of RPL23 with
MDM2 activates p53, which drives apoptosis, growth inhibition and
cell cycle arrest. TP, triptolide; Pol I, RNA polymerase I; UBF,
upstream binding factor; RPL23, ribosomal protein L23; MDM2, mouse
double minute 2; NCL, nucleolin; B23, nucleophosmin; PUMA,
p53-upregulated modulator of apoptosis; C, cleaved; p,
phosphorylated; r, ribosomal. |
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the National
Natural Science Foundation of China (grant no. 81774026) and the
Natural Science Foundation of Zhejiang Province (grant nos.
LQ19H100002 and Q17H290005).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WW and JW designed the research. JW, ZQZ, FQL, JNC,
XG, and BBC performed the experiments. JW and ZZQ analyzed the
data. JW and ZQZ wrote the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The study was approved by the Animal Experimentation
Ethics Committee of the Tongde Hospital of Zhejiang Province, and
was performed in accordance with the approved guidelines.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Osmani L, Askin F, Gabrielson E and Li QK:
Current WHO guidelines and the critical role of immunohistochemical
markers in the subclassification of non-small cell lung carcinoma
(NSCLC): Moving from targeted therapy to immunotherapy. Semin
Cancer Biol. 52:103–109. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jain NA and Otterson GA: Immunotherapy in
inoperable stage III non-small cell lung cancer: A review. Drugs
Context. 8:2125782019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang D, Jin Q, Jiang C, Gao M, Ni Y and
Zhang J: Imaging cell death: Focus on early evaluation of tumor
response to therapy. Bioconjug Chem. Mar 23–2020.(Epub ahead of
print). View Article : Google Scholar
|
|
4
|
Li-Weber M: Targeting apoptosis pathways
in cancer by Chinese medicine. Cancer Lett. 332:304–312. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Almeida EM, Ferreira HJ, Alves DR and
Da Silva WMB: Therapeutic potential of medicinal plants indicated
by the Brazilian public health system in treating the collateral
effects induced by chemotherapy, radiotherapy, and
chemoradiotherapy: A systematic review. Complement Ther Med.
49:1022932020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Luo D, Zuo Z, Zhao H, Tan Y and Xiao C:
Immunoregulatory effects of Tripterygium wilfordii Hook F and its
extracts in clinical practice. Front Med. 13:556–563. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Noel P, Von Hoff DD, Saluja AK, Velagapudi
M, Borazanci E and Han H: Triptolide and its derivatives as cancer
therapies. Trends Pharmacol Sci. 40:327–341. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Chen SR, Dai Y, Zhao J, Lin L and Wang Y
and Wang Y: A mechanistic overview of triptolide and celastrol,
natural products from tripterygium wilfordii Hook F. Front
Pharmacol. 9:1042018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yan P and Sun X: Triptolide: A new star
for treating human malignancies. J Cancer Res Ther. 14:S271–S275.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Meng G, Wang W, Chai K, Yang S, Li F and
Jiang K: Combination treatment with triptolide and
hydroxycamptothecin synergistically enhances apoptosis in A549 lung
adenocarcinoma cells through PP2A-regulated ERK, p38 MAPKs and Akt
signaling pathways. Int J Oncol. 46:1007–1017. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Li F, Zhao D, Yang S, Wang J, Liu Q, Jin X
and Wang W: ITRAQ-based proteomics analysis of triptolide on human
A549 lung adenocarcinoma cells. Cell Physiol Biochem. 45:917–934.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goodfellow SJ and Zomerdijk JC: Basic
mechanisms in RNA polymerase I transcription of the ribosomal RNA
genes. Subcell Biochem. 61:211–236. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang M and Pestov DG: Quantitative
northern blot analysis of mammalian rRNA processing. Methods Mol
Biol. 1455:147–157. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Russell J and Zomerdijk JC: The RNA
polymerase I transcription machinery. Biochem Soc Symp. 73:203–216.
2006. View Article : Google Scholar
|
|
15
|
O'Sullivan AC, Sullivan GJ and McStay B:
UBF binding in vivo is not restricted to regulatory sequences
within the vertebrate ribosomal DNA repeat. Mol Cell Biol.
22:657–668. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang HT, Chen TY, Weng CW, Yang CH and
Tang MS: Acrolein preferentially damages nucleolus eliciting
ribosomal stress and apoptosis in human cancer cells. Oncotarget.
7:80450–80464. 2016.PubMed/NCBI
|
|
17
|
Vispé S, DeVries L, Créancier L, Besse J,
Bréand S, Hobson DJ, Svejstrup JQ, Annereau JP, Cussac D, Dumontet
C, et al: Triptolide is an inhibitor of RNA polymerase I and
II-dependent transcription leading predominantly to down-regulation
of short-lived mRNA. Mol Cancer Ther. 8:2780–2790. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Boulon S, Westman BJ, Hutten S, Boisvert
FM and Lamond AI: The nucleolus under stress. Mol Cell. 40:216–227.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Mitrea DM, Cika JA, Guy CS, Ban D,
Banerjee PR, Stanley CB, Nourse A, Deniz AA and Kriwacki RW:
Nucleophosmin integrates within the nucleolus via multi-modal
interactions with proteins displaying R-rich linear motifs and
rRNA. Elife. 5:e135712016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Burger K, Mühl B, Harasim T, Rohrmoser M,
Malamoussi A, Orban M, Kellner M, Gruber-Eber A, Kremmer E, Hölzel
M and Eick D: Chemotherapeutic drugs inhibit ribosome biogenesis at
various levels. J Biol Chem. 285:12416–12425. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Turi Z, Lacey M, Mistrik M and Moudry P:
Impaired ribosome biogenesis: Mechanisms and relevance to cancer
and aging. Aging (Albany NY). 11:2512–2540. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Dai MS, Zeng SX, Jin Y, Sun XX, David L
and Lu H: Ribosomal protein L23 activates p53 by inhibiting MDM2
function in response to ribosomal perturbation but not to
translation inhibition. Mol Cell Biol. 24:7654–7668. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen J: The cell-cycle arrest and
apoptotic functions of p53 in tumor initiation and progression.
Cold Spring Harb Perspect Med. 6:a0261042016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Deregowska A, Adamczyk J, Kwiatkowska A,
Gurgul A, Skoneczny M, Skoneczna A, Szmatola T, Jasielczuk I, Magda
M, Rawska E, et al: Shifts in rDNA levels act as a genome buffer
promoting chromosome homeostasis. Cell Cycle. 14:3475–3487. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang W, Luo J, Xiang F, Liu X, Jiang M,
Liao L and Hu J: Nucleolin down-regulation is involved in
ADP-induced cell cycle arrest in S phase and cell apoptosis in
vascular endothelial cells. PLoS One. 9:e1101012014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Phi JH, Sun CH, Lee SH, Lee S, Park I,
Choi SA, Park SH, Lee JY, Wang KC, Kim SK, et al: NPM1 as a
potential therapeutic target for atypical teratoid/rhabdoid tumors.
BMC Cancer. 19:8482019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Okur MN, Lee JH, Osmani W, Kimura R,
Demarest TG, Croteau DL and Bohr VA: Cockayne syndrome group A and
B proteins function in rRNA transcription through nucleolin
regulation. Nucleic Acids Res. 48:2473–2485. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Murano K, Okuwaki M, Hisaoka M and Nagata
K: Transcription regulation of the rRNA gene by a multifunctional
nucleolar protein, B23/nucleophosmin, through its histone chaperone
activity. Mol Cell Biol. 28:3114–3126. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Engel C, Gubbey T, Neyer S, Sainsbury S,
Oberthuer C, Baejen C, Bernecky C and Cramer P: Structural basis of
RNA polymerase I transcription initiation. Cell. 169:120–131.e22.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin YM, Chu PH and Ouyang P: Ectopically
expressed pNO40 suppresses ribosomal RNA synthesis by inhibiting
UBF-dependent transcription activation. Biochem Biophys Res Commun.
516:381–387. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sugiyama T, Li S, Kato M, Ikeuchi K,
Ichimura A, Matsuo Y and Inada T: Sequential ubiquitination of
ribosomal protein uS3 triggers the degradation of non-functional
18S rRNA. Cell Rep. 26:3400–3415.e7. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang Y and Lu H: Signaling to p53:
Ribosomal proteins find their way. Cancer Cell. 16:369–377. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li XQ, Yu Q, Chen FS, Tan WF, Zhang ZL and
Ma H: Inhibiting aberrant p53-PUMA feedback loop activation
attenuates ischaemia reperfusion-induced neuroapoptosis and
neuroinflammation in rats by downregulating caspase 3 and the NF-κB
cytokine pathway. J Neuroinflammation. 15:2502018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Garufi A, Pistritto G, Baldari S, Toietta
G, Cirone M and D'Orazi G: p53-Dependent PUMA to DRAM antagonistic
interplay as a key molecular switch in cell-fate decision in
normal/high glucose conditions. J Exp Clin Cancer Res. 36:1262017.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen D, Ni HM, Wang L, Ma X, Yu J, Ding WX
and Zhang L: p53 up-regulated modulator of apoptosis induction
mediates acetaminophen-induced necrosis and liver injury in mice.
Hepatology. 69:2164–2179. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lessard F, Igelmann S, Trahan C, Huot G,
Saint-Germain E, Mignacca L, Del Toro N, Lopes-Paciencia S, Le
Calvé B, Montero M, et al: Senescence-associated ribosome
biogenesis defects contributes to cell cycle arrest through the Rb
pathway. Nat Cell Biol. 20:789–799. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Del Toro N, Fernandez-Ruiz A, Mignacca L,
Kalegari P, Rowell MC, Igelmann S, Saint-Germain E, Benfdil M,
Lopes-Paciencia S, Brakier-Gingras L, et al: Ribosomal protein
RPL22eL22 regulates the cell cycle by acting as an inhibitor of the
CDK4-cyclin D complex. Cell Cycle. 18:759–770. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Heissenberger C, Liendl L, Nagelreiter F,
Gonskikh Y, Yang G, Stelzer EM, Krammer TL, Micutkova L, Vogt S,
Kreil DP, et al: Loss of the ribosomal RNA methyltransferase NSUN5
impairs global protein synthesis and normal growth. Nucleic Acids
Res. 47:11807–11825. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Liu X, Tan Y, Zhang C, Zhang Y, Zhang L,
Ren P, Deng H, Luo J, Ke Y and Du X: NAT10 regulates p53 activation
through acetylating p53 at K120 and ubiquitinating Mdm2. EMBO Rep.
17:349–366. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xing Y, Liu Y, Liu T, Meng Q, Lu H, Liu W,
Hu J, Li C, Cao M, Yan S, et al: TNFAIP8 promotes the proliferation
and cisplatin chemoresistance of non-small cell lung cancer through
MDM2p53 pathway. Cell Commun Signal. 16:432018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Zhou Q, Hou Z, Zuo S, Zhou X, Feng Y, Sun
Y and Yuan X: LUCAT1 promotes colorectal cancer tumorigenesis by
targeting the ribosomal protein L40-MDM2-p53 pathway through
binding with UBA52. Cancer Sci. 110:1194–1207. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lempiäinen H and Shore D: Growth control
and ribosome biogenesis. Curr Opin Cell Biol. 21:855–863. 2009.
View Article : Google Scholar : PubMed/NCBI
|