Introduction
Budd-Chiari syndrome (BCS), a disease named after
the British physician George Budd and the Australian pathologist
Hans Chiari, is a relatively rare hepatic vascular disease, which
is mainly caused by obstruction of the hepatic vein outflow tract
(1–3). The incidence rate of membranous
obstruction of the inferior vena cava (MOVC) is the highest among
all types of BCS in China; in MOVC, the diaphragm forms at the
opening of the inferior vena cava and/or hepatic vein (4). This condition results in considerable
physiological damage and psychological trauma and increases the
economic burden of the family of affected patients.
Abnormal expressions of von Willebrand factor (vWF),
endothelin-1, vascular endothelial growth factor (VEGF) and other
cytokines have been detected in the plasma of patients with MOVC
compared with those of normal individuals. vWF and VEGF are
recognized as pro-angiogenic factors and are associated with
angiogenesis (5,6). Furthermore, a Chinese study reported
that granulation tissue and neovascularization were present in the
lesion of the inferior vena cava and that the luminal surface as
covered by a layer of intact endothelial cells (7). Therefore, the formation of the
inferior vena cava diaphragm possibly begins with proliferation and
angiogenesis after vascular endothelial injury.
microRNAs (miRNAs/miRs) are a class of non-coding,
endogenous and small RNA molecules with a length of ~22-28
nucleotides. miRNAs, as novel gene regulators, are able to inhibit
protein production by inducing mRNA degradation or combining with
the 3′-untranslated regions (UTRs) of target mRNAs to suppress the
process of translation (8,9). In this manner, miRNAs have a vital
function in a number of pathological and physiological processes,
including cell proliferation, death, immune function, apoptosis and
angiogenesis (10,11). A previous study by our group has
indicated that the expression of miR-3133 is significantly lower in
the plasma of patients with MOVC than in healthy controls (12). Therefore, downregulation of miR-3133
may be involved in the pathogenesis of MOVC through regulating the
proliferation and angiogenesis of endothelial cells. However, how
miR-3133 regulates the proliferation and angiogenesis of
endothelial cells has remained elusive and was therefore
investigated in the present study.
Materials and methods
Cell lines and reagents
Human umbilical vein endothelial cells (HUVECs; cat.
no. GNhu39) were purchased from the cell bank of the Chinese
Academy of Sciences. The cells were cultured in RPMI-1640 medium
(KeyGen Biotech Co., Ltd.) containing 10% fetal bovine serum
(Tianhang Biotechnology Co., Ltd.). The cells were incubated at
37°C in a humidified atmosphere with 5% CO2.
Plasmids and transient
transfections
miR-3133 mimics and their respective negative
control (NC), as well as miR-3133 inhibitor and its NC, were
purchased from RiboBio Biotechnology Co., Ltd. The
pc-DNA3.1–3×Flag-JUNB and pc-DNA3.1-3×Flag-vector were obtained
from Youbao Biotechnology Co., Ltd. The hilymax reagent was used to
transiently transfect plasmids for the miR-3133 mimics and
inhibitor into the cells according to the manufacturers protocol.
These transfections were performed when the HUVECs were cultured in
a 12-well plate at 1.2×105 cells/well with 1 ml fresh
medium for 24 h and reached 60–70% confluency.
Cell proliferation assay
HUVECs were cultured in a 12-well plate at
1.2×105 cells/well with 1 ml fresh medium for 24 h and
transfected with plasmids after the density reached 60–70%. For the
proliferation assay, these transfected cells were seeded into
96-well plate at 5×103 cells/well in 100 µl fresh medium
and culture was continued for 0, 24, 48 and 72 h prior to the assay
with the Cell Counting Kit-8 (CCK-8) (Dojindo).
Tube formation assay
For the tube formation assay, pre-chilled 96-well
plates were coated with 50 µl Matrigel™ (BD Biosciences) and
incubated at 37°C for 30 min. Subsequently, 1×104 cells
resuspended in 100 µl serum-free medium were added to each well and
then incubated at 37°C for another 6 h prior to capturing of images
under a microscope and counting the complete tubular
structures.
Reverse transcription-quantitative
(RT-q)PCR assay
The total RNA was isolated from cultured cells by
using TRIzol (Invitrogen; Thermo Fisher Scientific, Inc.). To
quantify gene expression, the complementary DNA of miR-3133 was
synthesized using the miRNA First Strand cDNA Synthesis Kit (by
stem-loop; Vazyme), while the RT reaction for JUNB and VEGF was
performed using the HiScript II First Strand cDNA Synthesis Kit
(Vazyme). The internal controls for miR-3133 and JUNB/VEGF were U6
and GAPDH, respectively. The 7500HT qPCR system thermal cycler
(Applied Biosystems; Thermo Fisher Scientific, Inc.) was used for
the amplification of the obtained complementary DNA in triplicate
with AceQ qPCR SYBR-Green Master Mix (Vazyme). The program settings
were as follows: Pre-denaturation at 95°C for 5 min, followed by 40
cycles of 10 sec at 95°C, annealing at 60°C for 30 sec and
extension at 95°C for 15 sec. The relative gene expression was
determined using the 2−ΔΔCq method (13). The primers used were as follows:
5′-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACATTGGG-3′
(stem-loop), 5′-CGCGCGTAAAGAACTCTTAAAA-3′ (forward) and
5′-AGTGCAGGGTCCGAGGTATT-3′ (reverse) for miR-3133;
5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-AACGCTTCACGAATTTGCGT-3′
(reverse) for U6; 5′-TGGAACAGCCCTTCTACCAC-3′ (forward) and
5′-TGGAACAGCCCTTCTACCAC-3′ (reverse) for JUNB;
5′-TGCCCGCTGCTGTCTAATG-3′ (forward) and 5′-GCGAGTCTGTGTTTTTGCAG-3′
(reverse) for VEGF; 5′-GCCGGTGCTGAGTATGTC-3′ (forward) and
5′-CTTCTGGGTGGCAGTGAT-3′ (reverse) for GAPDH.
Western blot analysis
Following transfection, the HUVECs were collected
and lysed using radioimmunoprecipitation assay lysis buffer
containing 100 mM phenylmethylsulfonyl fluoride and ultrasound. The
supernatant was collected after centrifuging at 12435 × g for 5 min
at 4°C and the protein concentration was measured using a
bicinchoninic acid protein assay kit (Vazyme) according to the
manufacturers protocol. Equal amounts of protein were separated by
10% SDS-PAGE and then transferred to a polyvinylidene difluoride
membrane. The membranes were blocked with Tris-buffered saline
containing 0.1% Tween-20 (TBST) and 5% skimmed milk at room
temperature for 2 h. The primary antibodies, including anti-JUNB
(cat. no. ab128878; 1:1,000 dilution; Abcam) and anti-VEGF (cat.
no. ER30607; 1:1,000 dilution; Huabio) were incubated at 4°C
overnight. The internal control of proteins was α-tubulin (cat. no.
10094-1-AP; 1:1,000 dilution; Beyotime Institute of Biotechnology,
Inc.). The membranes were washed three times with TBST and
incubated with horseradish peroxidase (HRP)-labeled goat
anti-rabbit IgG secondary antibodies (cat. no. A0208; 1:1,000
dilution; Beyotime Institute of Biotechnology, Inc.) at room
temperature for 1 h. The bands were visualized using Immobilon
Western Chemiluminescent HRP Substrate (EMD Millipore). The results
were analyzed by Image Lab 5.2.1 (Bio-Rad Laboratories, Inc.) and
each blot was performed at least three times.
ELISA
HUVECS (2.5×105 cells/well) were seeded
into a six-well plate and the medium was replaced with 1 ml
serum-free fresh medium at 24 h after transfection. After another
24 h, the supernatant was collected and centrifuged at 1,000 × g
for 20 min at 4°C to perform ELISA. The human VEGF ELISA kit
(Elabscience) was used to detect the concentration of VEGF
according to the manufacturers protocol.
Bioinformatics prediction and
luciferase reporter assay
MicroRNA.org (www.microrna.org/microrna/; August 2010 release; last
update, 1 November 2010; Species, human) was used to predict the
putative targets of miR-3133 (14).
The wild-type (WT) 3′-UTR of JUNB containing predicted miR-3133
binding sequences and the mutant (Mut) 3′-UTR of JUNB were
chemically produced by ABM Biotechnology Co., Ltd. The WT and Mut
3UTR of JUNB were subcloned into the psiCHECK-2 vector. HUVECs at
5×104 cells/well in 24-well plates were co-transfected
with 0.5 µg reporter plasmid and miR-3133 mimics or miR-3133 mimics
NC using hilymax reagent for 36 h, and cell lysates (Promega Corp.)
were then prepared according to the manfacturers instructions.
Luciferase activity was measured with a dual-Luciferase Reporter
Assay System (Promega Corp.) and the activity was normalized to the
Renilla luciferase gene.
Statistical analysis
Values are expressed as the mean ± standard
deviation and analyzed with SPSS 20.0 (IBM Corp.). The two-tailed
Students t-test (two groups) or one-way analysis of variance were
used to evaluate the inter-group differences. When comparing
between any two means, the data satisfying the normality and the
homogeneity of the variance were compared with the least
significant differences t-test, and otherwise, Dunnetts T3 test was
used. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-3133 regulates the proliferation
and angiogenesis of HUVECs
miR-3133 mimics, inhibitor and their respective NCs
were transfected into cells using the hilymax reagent. RT-qPCR
demonstrated that the mimics increased the levels of miR-3133,
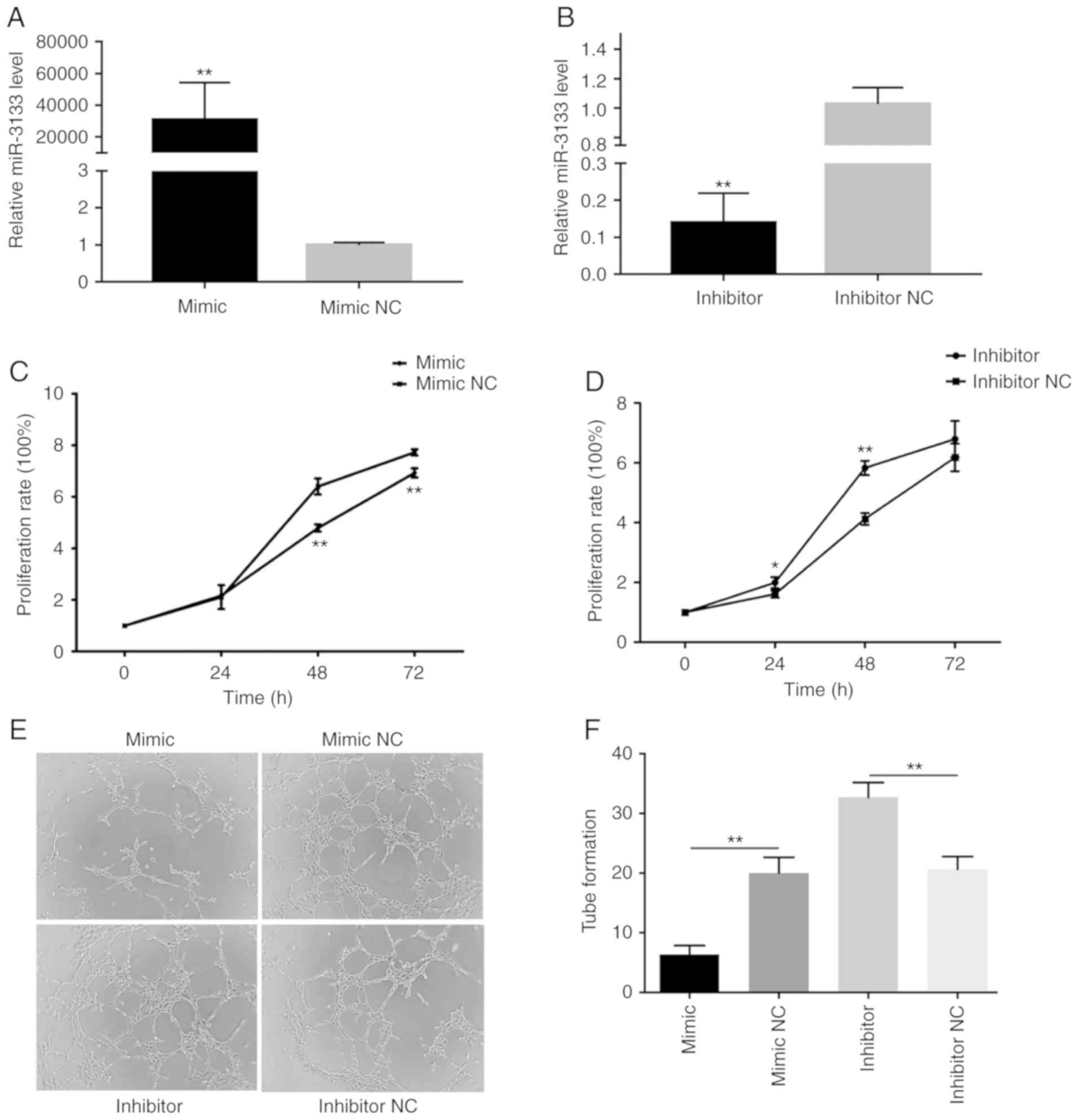
whereas the inhibitor reduced the levels of miR-3133 (Fig. 1A and B). The results of the CCK-8
assay indicated that, compared with their respective NCs, the
miR-3133 mimics attenuated the proliferation of HUVECs, whereas the
miR-3133 inhibitor promoted it (Fig. 1C
and D). In addition, the tube formation assay demonstrated that
the miR-3133 mimics reduced the formation of tube-like structures,
whereas miR-3133 inhibitor caused their stimulation (Fig. 1E and F). Taken together, these
results indicated that miR-3133 suppresses proliferation and
angiogenesis.
JUNB is a target gene of miR-3133
MicroRNA.org was used to predict
target genes of miR-3133 to study the molecular mechanism of
miR-3133. The mirSVR score and PhastCons score are the indicators
in the miRanda evaluation. Genes with a lower mirSVR score and a
higher PhastCons score were more likely to be a target gene of
miR-3133. The prediction result suggested that JUNB (mirSVR
score=−1.2373, PhastCons score=0.5511) was a potential target gene
of miR-3133 (Fig. 2A).
Subsequently, a dual-luciferase assay was performed, revealing that
the luciferase activity in cells cotransformed with miR-3133 mimics
and wild-type JUNB was significantly reduced compared with that in
cells cotransformed with miR-3133 mimics and mutated JUNB. However,
the luciferase activity in cells cotransformed with miR-3133 mimics
NC and wild-type JUNB was not significantly different compared with
that in cells cotransformed with miR-3133 mimics NC and mutant-type
JUNB (Fig. 2B).
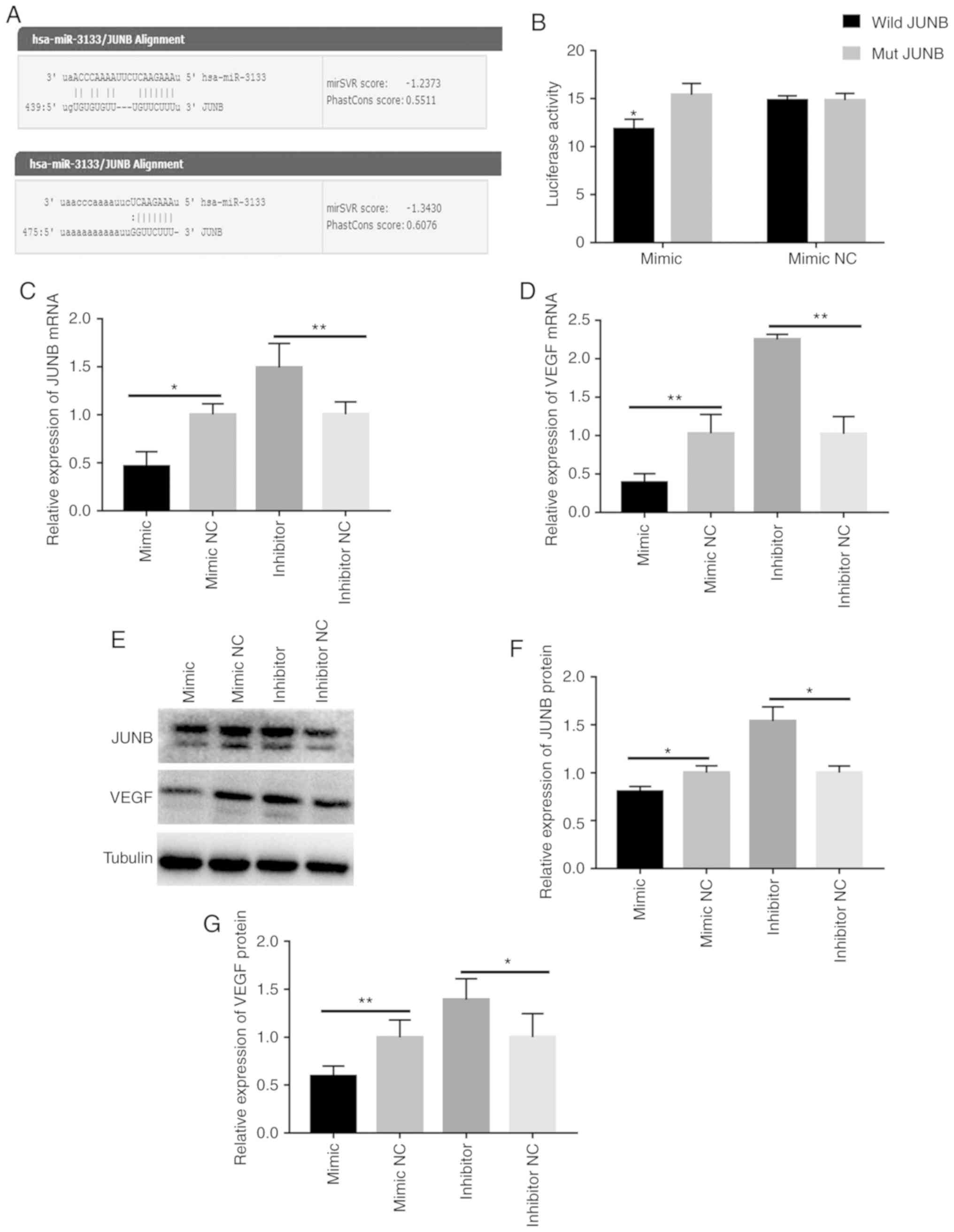 | Figure 2.JUNB is a direct target of miR-3133,
whereas miR-3133 inhibits the expression of JUNB and VEGF. (A)
Putative hsa-miR-3133 binding and mutated sites in the 3′UTR of
JUNB. (B) JUNB 3′UTR reporter activity in the presence of miR-3133
mimics or mimics NC was measured by a luciferase assay and
normalized to the activity of Renilla luciferase. (C)
RT-qPCR analysis of JUNB expression in HUVECs after transfection.
(D) RT-qPCR analysis of VEGF expression in HUVECs after
transfection. (E and F) Western blot images and analysis of JUNB
and VEGF, respectively. (G) ELISA of VEGF concentration in the
supernatant. Values are expressed as the mean ± standard deviation.
*P<0.05 and **P<0.01 vs. the respective NC. miR, microRNA;
NC, negative control; HUVECs, human umbilical vein endothelial
cells; RT-qPCR, reverse transcription-quantitative PCR; VEGF,
vascular endothelial growth factor; hsa, Homo sapiens; wild,
wild-type; mut, mutant; UTR, untranslated region. |
miR-3133 inhibits the expression of
JUNB and VEGF in HUVECs
After transfection, the expression of JUNB and VEGF
was detected at the mRNA and protein level by RT-qPCR and western
blot analysis, respectively. The results indicated that, compared
with the respective NC, the expression levels of JUNB and VEGF were
significantly decreased in the mimics group and were higher in the
inhibitor group (Fig. 2C-F). In
addition, ELISA was used to detect the secretion levels of VEGF
after transfection, and the results suggested that, compared with
the respective control group, the secretion of VEGF was reduced in
the miR-3133 mimics group and upregulated in the miR-3133 inhibitor
group (Fig. 2G). These results
demonstrate that miR-3133 is able to negatively regulate the
expression of JUNB and VEGF.
Overexpression of JUNB promotes
proliferation and angiogenesis in HUVECs
The forced expression of JUNB in endothelial cells
was previously reported to stimulate the formation of a tip-like
cell morphology and angiogenesis and induces VEGF expression
(15–17). In the present study, it was assessed
whether JUNB has a role in the proliferation and angiogenesis of
HUVECs. The JUNB overexpression plasmid (JUNBOE) was constructed
and the empty vector (Cont) was used as a control. The expression
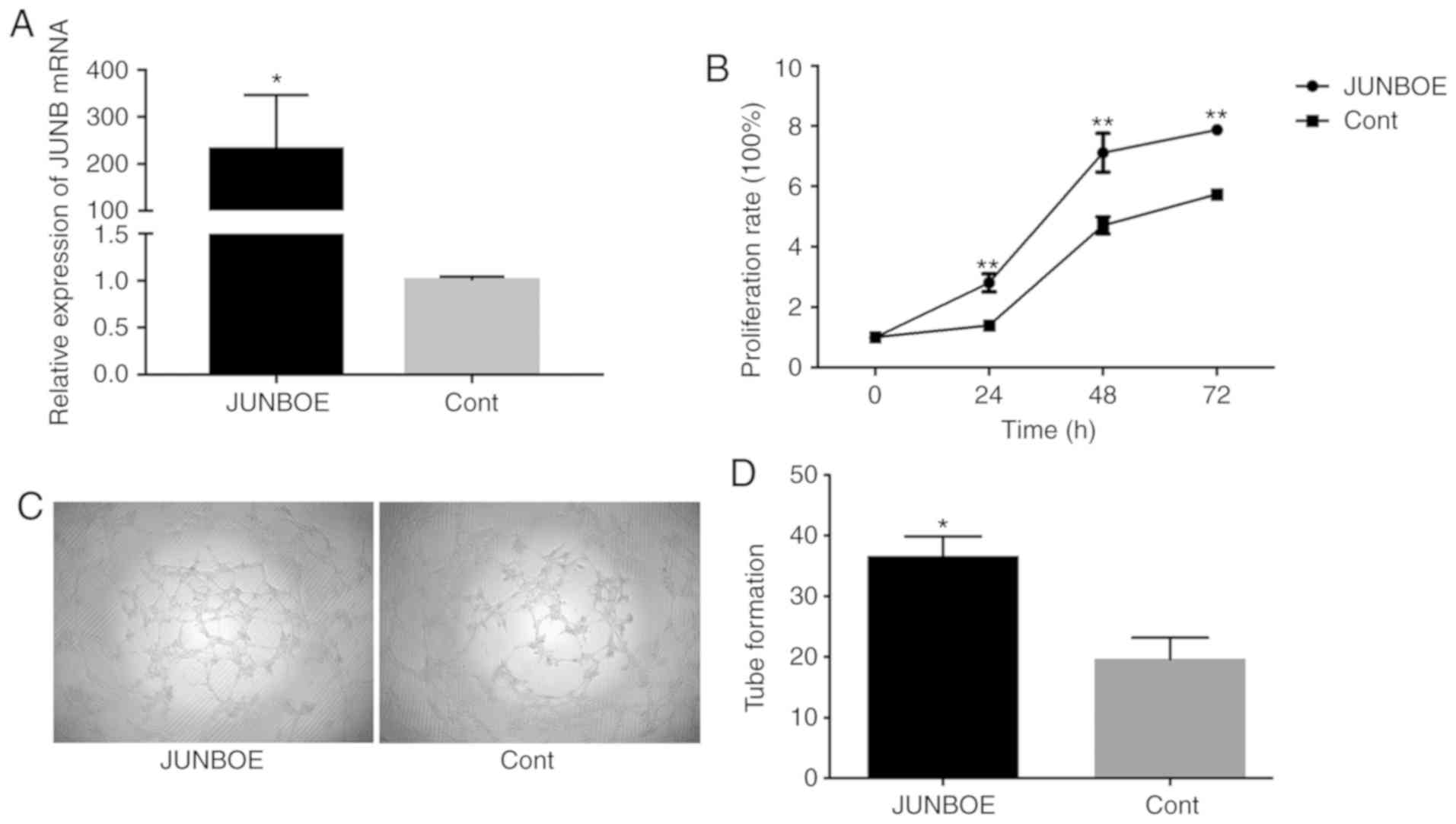
of JUNB was detected using RT-qPCR to determine whether the
overexpression with JUNBOE was effective. The results indicated
that, compared with the Cont group, JUNB was highly expressed in
the JUNBOE group (Fig. 3A). The
CCK-8 assay was then used to detect cell proliferation after
transfection and tube formation was evaluated by a tube formation
assay. The results suggested that the proliferation of HUVECs in
the JUNBOE group was higher than that in the Cont group at 24 h
after transfection (Fig. 3B). In
addition, the tube formation ability in the JUNBOE group increased
compared with that in the Cont group (Fig. 3C and D).
JUNB positively regulates the
expression and secretion of VEGF in HUVECs
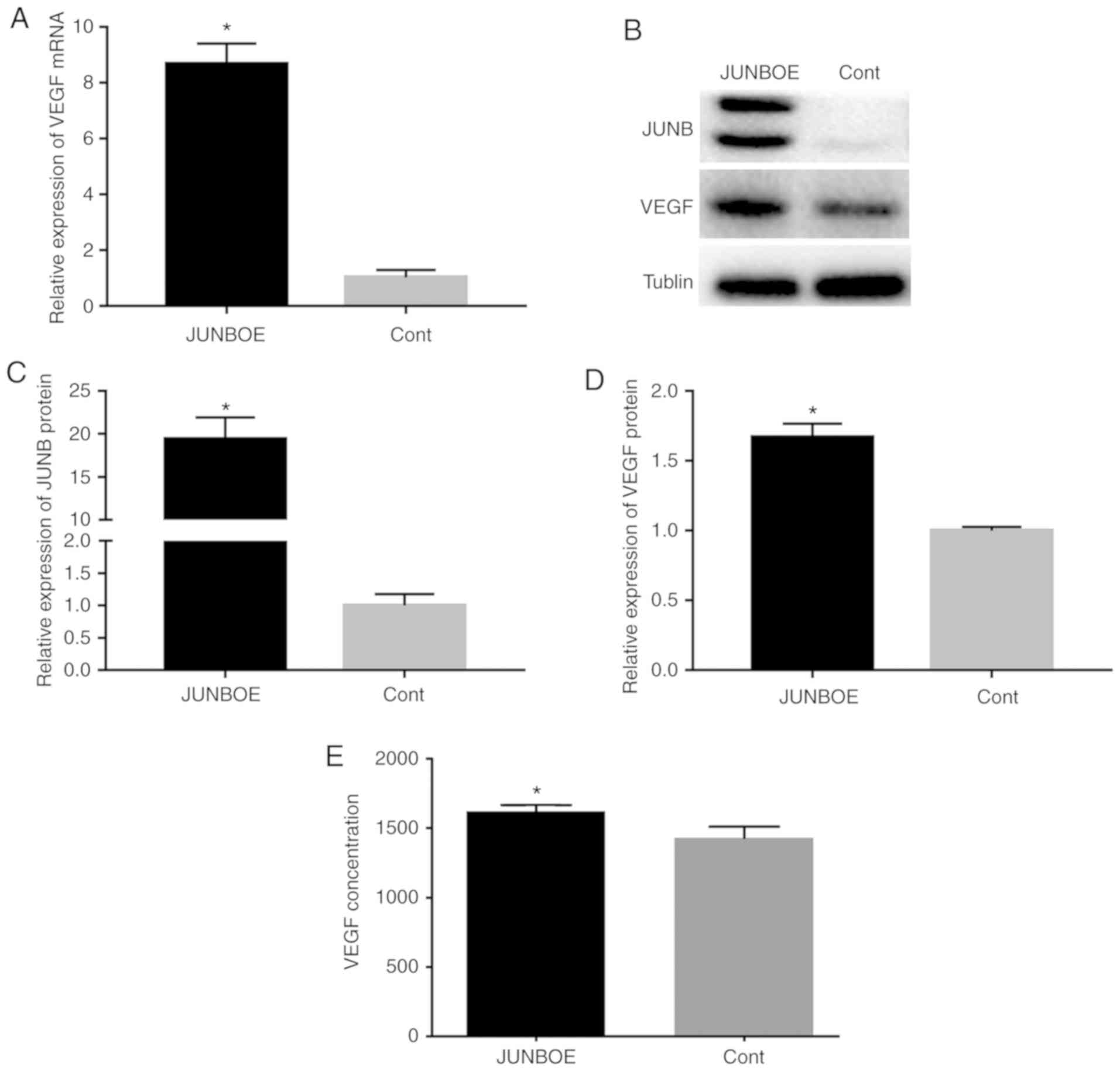
The expression of VEGF at the mRNA and protein
levels was assessed after transfection with JUNBOE or Cont to
verify the role of JUNB in the regulation of VEGF. The results
indicated that the mRNA (Fig. 4A)
and protein expression levels (Fig.
4B-D) of VEGF increased when JUNB was overexpressed.
Furthermore, JUNB overexpression induced the secretion of VEGF
(Fig. 4E).
miR-3133 reduces the proliferation and
angiogenesis ability of HUVECs promoted by JUNB overexpression
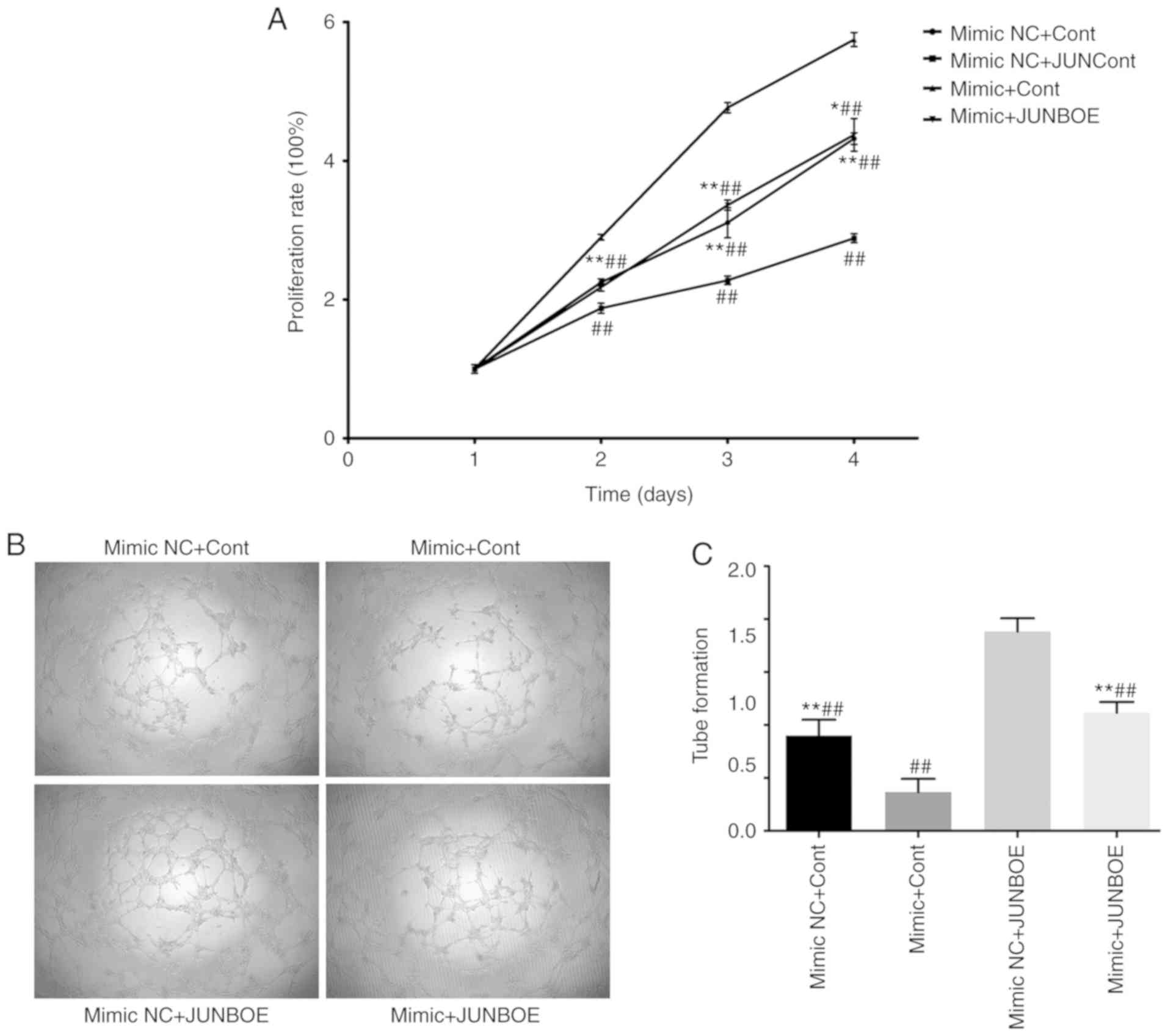
miR-3133 mimics or miR-3133 mimics NC was
transfected into cells together with JUNBOE or Cont. A total of
four groups were included in the experiment, namely mimics NC+Cont,
mimics+Cont, mimics NC+JUNBOE and mimics+JUNBOE. The results of the
CCK-8 assay indicated that the mimics NC+JUNBOE group had the
highest cell proliferation rate, whereas the mimics+Cont group had
the lowest rate (Fig. 5A). In
addition, in the tube formation assay, the number of tubular
structures in the four groups exhibited the same trend (Fig. 5B and C). This result indicated that
miR-3133 partially inhibited the positive regulation of
proliferation and angiogenesis by JUNB.
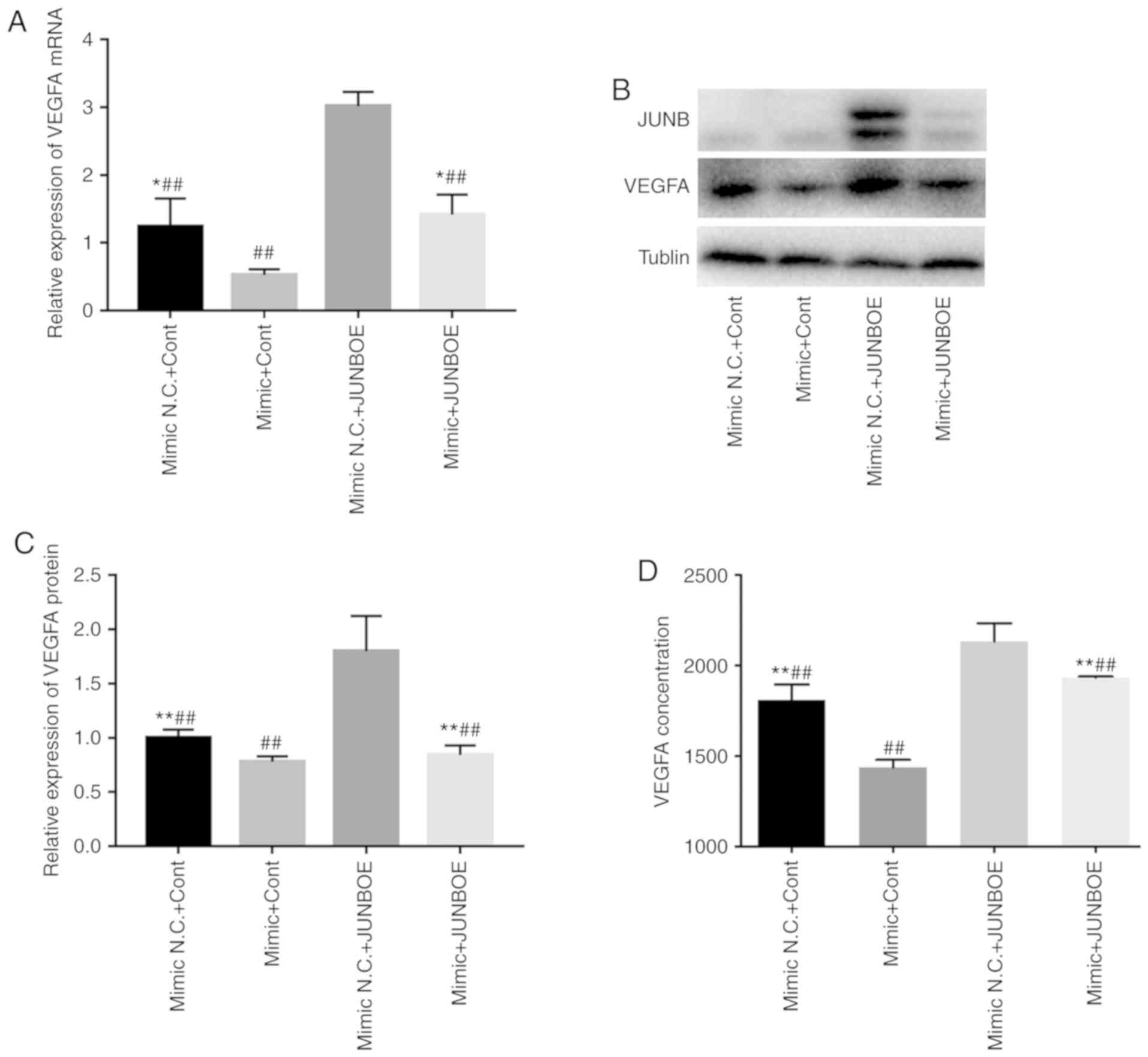
miR-3133 abrogates the induction of
VEGF expression caused by overexpression of JUNB
VEGFA expression was detected at the mRNA and
protein level by RT-qPCR and western blot analysis, respectively.
The results indicated that compared with the mimics NC+control
group, the expression of VEGFA was markedly upregulated in the
mimics NC+JUNBOE group, while it was decreased in the mimics+Cont
group. Co-transfection with mimics abrogated the effect of JUNBOE
to upregulate VEGFA. Furthermore, JUNB was downregulated in the
mimics+JUNBOE group compared with the mimics NC+JUNBOE group, which
indicates that miR-3133 exerts its inhibitory effect on VEGFA
expression via inhibiting the expression of JUNB (Fig. 6A-C). In addition, the results of the
ELISA revealed that, compared with the mimics+Cont group, the
concentration of VEGFA secreted into the culture medium by HUVECs
after cotransformation of miR-3133 and/or JUNBOE was markedly
upregulated in the mimics NC+JUNBOE group (Fig. 6D).
Discussion
BCS is a blood reflux disorder caused by complete or
incomplete obstruction of the hepatic vein outflow tract or
inferior vena cava hepatic segment. Its clinical manifestations are
mainly portal hypertension or inferior vena cava hypertension
(18,19). From a global perspective, BCS is a
rare disease, but a large number of cases are reported in
relatively poor countries, including China, India, South Africa and
Nepal, whereas relatively few cases are reported in Western
countries (20). BCS is a complex
process involving numerous factors and the pathogenesis and
epidemiology exhibit marked differences between China and Western
countries (21). In China, MOVC is
the most common type of BCS. However, the specific pathogenesis
remains to be fully elucidated. Therefore, the present study aimed
to preliminarily assess the possible molecular mechanisms of the
occurrence of MOVC.
Riemens et al (22) reported that the inferior vena cava
septum is composed of small pieces of fibrous tissue, which contain
capillaries, and the surface is covered with endothelial cells. The
two surfaces of the membrane are composed of vascular endothelial
tissues and membrane formation occurs due to endothelial damage
(4,23). Therefore, abnormal proliferation and
angiogenesis of HUVECs are the key factors involved in membranous
formation.
Angiogenesis refers to the formation of new blood
vessels by the development of existing capillaries or
post-capillary veins; it includes the following processes:
Degradation of the vascular basement membrane; activation,
proliferation and migration of vascular endothelial cells; and
reconstruction of new blood vessels and vascular network (24).
Angiogenesis is a complex process that relies on the
coordination of angiogenic and inhibitory factors. Under normal
circumstances, these factors are in equilibrium. Once this balance
is broken, the vascular system is activated, causing excessive
angiogenesis or inhibiting the vascular system to degenerate blood
vessels. VEGF is a heparin-binding angiogenic growth factor
displaying high specificity for vascular endothelial cells and
regulates proliferation, migration and angiogenesis (25–27).
As a key regulator to sustain endothelial function, VEGF correlates
with the thrombus organization and modulates the function of
vascular endothelial cells (25,26,28).
In a preliminary study by our group, the plasma samples from nine
patients with MOVC and five healthy control were analyzed (12). Analysis of these samples indicated
that the serum concentration of VEGF in patients with MOVC was
increased compared with that in subjects without BCS or any other
types of BCS.
In the human genome, almost one-third of the genes
are thought to be regulated by miRNAs. miRNAs function as important
endogenous regulators of gene expression and are thereby implicated
in modulating various biological processes through complementary
miRNA-mRNA binding to form silencing complexes (25,29–35).
Recent evidence has indicated that the dysregulation of miRNA is
involved in numerous pathological processes (36–38),
and miRNA inhibitors have been demonstrated to have tremendous
therapeutic potential in a multitude of studies (39,40).
Therefore, continuous exploration of the roles of miRNAs will
provide possible therapeutic targets for MOVC.
A previous study by our group reported that a series
of miRNAs was abnormally expressed in patients with MOVC (12). Among these miRNAs, miR-3133 was
significantly downregulated. Thus, in the present study, miR-3133
mimics, mimics NC, inhibitor and inhibitor NC were individually
transfected into HUVECs to detect the association between miR-3133
and processes of septum formation. A subsequent CCK-8 assay
demonstrated that miR-3133 mimics attenuated the proliferation
HUVECS, whereas miR-3133 inhibitor promoted the proliferation as
compared with the respective NC. In addition, the tube formation
assay demonstrated that miR-3133 mimics reduced, whereas miR-3133
inhibitor stimulated the formation of tube-like structures by
HUVECs. Taken together, miR-3133 has an inhibitory role in the
proliferation and angiogenesis of HUVECs.
The results of MicroRNA.org
and a dual-Luciferase reporter assay proved that JUNB was a target
gene of miR-3133. Furthermore, miR-3133 was able to negatively
regulate JUNB and VEGF at the mRNA and protein level. In addition,
ELISA demonstrated that the secretion of VEGF by HUVECs was reduced
following transfection with miR-3133 overexpression vector.
Yoshitomi et al (15) and
Ryzhov et al (41) proved
that JUNB not only positively regulates VEGF at the mRNA and
protein level but also promotes VEGF secretion. The proliferative
and angiogenic capacity of cells was enhanced when JUNB was
overexpressed. The inhibitory effects of miR-3133 mimics on the
proliferation and angiogenic capacity were abrogated by
co-transfection with JUNB, which indicated that miR-3133 inhibited
the expression of JUNB so that it could not exert its role in
promoting proliferation and angiogenesis and reduced the expression
and secretion of VEGF.
In conclusion, the present study indicated that
miR-3133 regulated the angiogenesis potential of HUVECs through the
JUNB/VEGF pathway, and may have an important role in membrane
formation as a key pathogenic process of MOVC. However, this is
only a possible mechanism. In order to further validate the
angiogenic role of miR-3133 in MOVC in future studies, it will be
attempted to examine pathological HUVECs, plasma samples and their
corresponding clinical features to study the expression levels of
miR-3133 in pathological HUVECs and the association between
miR-3133 and the disease severity.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from the
National Natural Science Foundation of China (grant no.
81872647).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors contributions
GS, LX and YC conceived and designed the
experiments. MX, LC, XZ, YuZ, YiZ and QW performed the experiments.
MX, LC and XZ analyzed the experimental data and wrote the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MOVC
|
membranous obstruction of the inferior
vena cava
|
|
BCS
|
Budd Chiari syndrome
|
|
VEGF
|
vascular endothelial growth factor
|
|
JUNB
|
JunB proto-oncogene
|
|
HUVECs
|
human umbilical vein endothelial
cells
|
|
CCK-8
|
Cell Counting Kit-8
|
References
|
1
|
Menon KV, Shah V and Kamath PS: The
Budd-Chiari syndrome. N Engl J Med. 350:578–585. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martens P and Nevens F: Budd-Chiari
syndrome. United European Gastroenterol J. 3:489–500. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zanetto A, Pellone M and Senzolo M:
Milestones in the discovery of Budd-Chiari syndrome. Liver Int.
39:1180–1185. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dang X, Li L and Xu P: Research status of
Budd-Chiari syndrome in China. Int J Clin Exp Med. 7:4646–4652.
2014.PubMed/NCBI
|
|
5
|
Yuan W, Qian M, Li ZX, Zhao CL, Zhao J and
Xiao JR: Endothelin-1 activates the notch signaling pathway and
promotes tumorigenesis in giant cell tumor of the spine. Spine
(Phila Pa 1976). 44:E1000–E1009. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tsao CJ, Pandolfi L, Wang X, Minardi S,
Lupo C, Evangelopoulos M, Hendrickson T, Shi A, Storci G, Taraballi
F and Tasciotti E: Electrospun patch functionalized with
nanoparticles allows for spatiotemporal release of VEGF and PDGF-BB
promoting in vivo neovascularization. ACS Appl Mater Interfaces.
10:44344–44353. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Qiao SS, Dang XW, Xu DQ, Wu Y, Li J, Zhang
HX, Chen KS, Xu PQ and Zhang SJ: Morphological features of
pathological membrane of inferior vena cava associated with
Budd-Chiari syndrome. Chin J Exp Sur. 29:1598–1600. 2012.
|
|
8
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kontomanolis EN and Koukourakis MI:
MicroRNA: The potential regulator of endometrial carcinogenesis.
Microrna. 4:18–25. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Urbich C, Kuehbacher A and Dimmeler S:
Role of microRNAs in vascular diseases, inflammation, and
angiogenesis. Cardiovasc Res. 79:581–588. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer. 96
(Suppl):R40–R44. 2007.PubMed/NCBI
|
|
12
|
Sun GX, Su Y, Li Y, Zhang YF, Xu LC, Zu
MH, Huang SP, Zhang JP and Lu ZJ: Circulating microRNA profile in
patients with membranous obstruction of the inferior vena cava. Exp
Ther Med. 11:811–817. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in Drosophila. Genome Biol.
5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yoshitomi Y, Ikeda T, Saito H, Yoshitake
Y, Ishigaki Y, Hatta T, Kato N and Yonekura H: JunB regulates
angiogenesis and neurovascular parallel alignment in mouse
embryonic skin. J Cell Sci. 130:916–926. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zou Y, Li Q, Xu Y, Yu X, Zuo Q, Huang S,
Chu Y, Jiang Z and Sun L: Promotion of trophoblast invasion by
lncRNA MVIH through inducing Jun-B. J Cell Mol Med. 22:1214–1223.
2018.PubMed/NCBI
|
|
17
|
Sadri D, Farhadi S and Nourmohamadi P:
Angiogenesis in odontogenic keratocyst and dentigerous cyst:
Evaluation of JunB and VEGF expression. Dent Res J (Isfahan).
16:327–332. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Dang XW, Xu PQ, Ma XX, Xu DQ, Zhu YJ and
Zhang YS: Surgical treatment of Budd-Chiari syndrome: Analysis of
221 cases. Hepatobiliary Pancreat Dis Int. 10:435–438. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li SL, Zu MH and Lu ZJ: A review on the
research status and trends of Budd-Chiari syndrome. Zhonghua Liu
Xing Bing Xue Za Zhi. 31:1192–1195. 2010.(In Chinese). PubMed/NCBI
|
|
20
|
Wang ZG, Zhang FJ, Yi MQ and Qiang LX:
Evolution of management for Budd-Chiari syndrome: A team's view
from 2564 patients. ANZ J Surg. 75:55–63. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Qi XS, Guo XZ and Fan DM: Difference in
Budd-Chiari syndrome between the West and China. Hepatology.
62:6562015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Riemens SC, Haagsma EB, Kok T, Gouw AS and
van der Jagt EJ: Familial occurrence of membranous obstruction of
the inferior vena cava: Arguments in favor of a congenital
etiology. J Hepatol. 22:404–409. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Teng F, Zu MH and Hua QJ: Correlations of
iodide ions with vascular endothelial growth factor and its
receptors during the proliferation of vascular endothelial cells.
Genet Mol Res. 13:6439–6447. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chamorro-Jorganes A, Lee MY, Araldi E,
Landskroner-Eiger S, Fernández-Fuertes M, Sahraei M, Quiles Del Rey
M, van Solingen C, Yu J, Fernández-Hernando C, et al: VEGF-induced
expression of miR-17-92 cluster in endothelial cells is mediated by
ERK/ELK1 activation and regulates angiogenesis. Circ Res.
118:38–47. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shibuya M: Vascular endothelial growth
factor and its receptor system: Physiological functions in
angiogenesis and pathological roles in various diseases. J Biochem.
153:13–19. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kim S, Jun JH, Kim J, Kim DW, Jang YH, Lee
WJ, Chung HY and Lee SJ: HIF-1α and VEGF expression correlates with
thrombus remodeling in cases of intravascular papillary endothelial
hyperplasia. Int J Clin Exp Pathol. 6:2912–2918. 2013.PubMed/NCBI
|
|
29
|
Zhou XL, Wu JH, Wang XJ and Guo FJ:
Integrated microRNA-mRNA analysis revealing the potential roles of
microRNAs in tongue squamous cell cancer. Mol Med Rep. 12:885–894.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hao Y, Yang J, Yin S, Zhang H, Fan Y, Sun
C, Gu J and Xi JJ: The synergistic regulation of VEGF-mediated
angiogenesis through miR-190 and target genes. RNA. 20:1328–1336.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chen L, Li ZY, Xu SY, Zhang XJ, Zhang Y,
Luo K and Li WP: Upregulation of miR-107 inhibits glioma
angiogenesis and VEGF expression. Cell Mol Neurobiol. 36:113–120.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li X, Zhang J, Gao L, McClellan S, Finan
MA, Butler TW, Owen LB, Piazza GA and Xi Y: MiR-181 mediates cell
differentiation by interrupting the Lin28 and let-7 feedback
circuit. Cell Death Differ. 19:378–386. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liu XD, Cai F, Liu L, Zhang Y and Yang AL:
MicroRNA-210 is involved in the regulation of postmenopausal
osteoporosis through promotion of VEGF expression and osteoblast
differentiation. Biol Chem. 396:339–347. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jiang FS, Tian SS, Lu JJ, Ding XH, Qian
CD, Ding B, Ding ZS and Jin B: Cardamonin regulates miR-21
expression and suppresses angiogenesis induced by vascular
endothelial growth factor. Biomed Res Int. 2015:5015812015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mei H, Lin ZY and Tong QS: The roles of
microRNAs in neuroblastoma. World J Pediatr. 10:10–16. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Su YF, Zang YF, Wang YH and Ding YL:
MiR-19-3p induces tumor cell apoptosis via targeting FAS in rectal
cancer cells. Technol Cancer Res Treat. 19:15330338209179782020.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lu H, Zhang L, Lu S, Yang D, Ye J, Li M
and Hu W: miR-25 expression is upregulated in pancreatic ductal
adenocarcinoma and promotes cell proliferation by targeting ABI2.
Exp Ther Med. 19:3384–3390. 2020.PubMed/NCBI
|
|
38
|
Tiwari A, Mukherjee B and Dixit M:
MicroRNA key to angiogenesis regulation: MiRNA biology and therapy.
Curr Cancer Drug Targets. 18:266–277. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yuan M, Huang LL, Chen JH, Wu J and Xu Q:
The emerging treatment landscape of targeted therapy in
non-small-cell lung cancer. Signal Transduct Target Ther. 4:612019.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhao X, Hu GF, Shi YF and Xu W: Research
progress in microRNA-based therapy for gastric cancer. Onco Targets
Ther. 12:11393–11411. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Ryzhov S, Biktasova A, Goldstein AE, Zhang
Q, Biaggioni I, Dikov MM and Feoktistov I: Role of JunB in
adenosine A2B receptor-mediated vascular endothelial growth factor
production. Mol Pharmacol. 85:62–73. 2014. View Article : Google Scholar : PubMed/NCBI
|