Introduction
Oral cancers account for about 2% of all cancer
cases diagnosed worldwide (1). More
than 350,000 individuals are diagnosed with oral cancer every year,
and oral cancers prove fatal for approximately 170,000 of these
people. Major risk factors for oral cancer include the use of
alcohol and tobacco (2). Although
decreased drinking and smoking have resulted in a decline in the
incidence of oral cancer, recent studies have reported an increase
in the number of young patients diagnosed with these diseases
(3,4).
CD44 is known to be expressed in many cell types,
including epithelial cells, fibroblasts, endothelial cells, and
leukocytes (5). CD44 plays
important roles in cell proliferation, adhesion, and migration
(6). The CD44 gene consists of 20
exons (7). The smallest isoform is
the standard form of CD44 (CD44s), which possesses 10 exons; other
possible isoforms are categorized as CD44 variants (CD44v), which
are generated by alternatively spliced transcripts (8). Post-translational modifications such
as N- and O-glycosylation and heparan sulfate
modification also augment the diversity of CD44 (9,10).
Both CD44s and CD44v are overexpressed in many cancers; however, a
pattern of expression remains to be elucidated.
One of the CD44 variants, CD44v6, was first
identified as contributing to cancer metastasis, and
CD44v6-specific monoclonal antibodies (mAbs) were found to inhibit
metastasis of rat pancreatic cancers (11,12).
Some CD44v6 isoforms act as co-receptors for receptor tyrosine
kinases (RTKs) such as MET and vascular endothelial growth factor
receptor (VEGFR)-2 (13–15). The transfection of CD44v4-7 cDNA
confers a metastatic phenotype in non-metastatic cells (16). Another CD44 variant, CD44v3, binds
to several heparan sulfate-binding growth factors such as
fibroblast growth factors (FGFs) and heparin-binding epidermal
growth factor (HB-EGF), and induces tumor progression (17,18).
Several CD44 variants were also reported as prognostic markers in
head and neck, lung, colorectal, breast, and hepatocellular cancers
(19–23).
Many mAbs have been developed to target CD44
(24–26). mAbs that neutralize contact between
hyaluronic acid and CD44 have been shown to inhibit
anchorage-independent growth of murine mammary carcinoma cells and
human colon carcinoma cells (24).
Anti-CD44 mAbs were also found to exhibit significant antitumor
activity in mouse xenograft models of human cancers (25,26).
Previously, we established clone C44Mab-5
(IgG1, kappa) using Cell-Based Immunization and
Screening (CBIS) (27).
C44Mab-5 recognized both CD44s and CD44v isoforms, and
demonstrated high sensitivity for flow cytometry and
immunohistochemical analysis in oral cancers. Because the
IgG1 subclass of C44Mab-5 lacks
antibody-dependent cellular cytotoxicity (ADCC) and
complement-dependent cytotoxicity (CDC), antitumor activity of
C44Mab-5 could not be determined.
In this study, we converted the IgG1
subclass C44Mab-5 into a mouse IgG2a subclass
mAb, 5-mG2a, and further produced a defucosylated
version, 5-mG2a-f, using FUT8-deficient ExpiCHO-S cells
(28). We then investigated whether
5-mG2a-f exhibited ADCC, CDC and antitumor activities
against oral cancers.
Materials and methods
Cell lines
Oral squamous carcinoma cell lines including HSC-2
(oral cavity) and SAS (tongue) were obtained from the Japanese
Collection of Research Bioresources Cell Bank (JCRB; Osaka, Japan).
Chinese hamster ovary (CHO)-K1 was obtained from the American Type
Culture Collection (ATCC). CD44v3-10 plus N-terminal PA16
tag-overexpressed CHO-K1 (CHO/PA16-CD44v3-10) was generated by
transfection of pCAG/PA16-CD44v3-10 to CHO-K1 cells using the Neon
Transfection System (Thermo Fisher Scientific, Inc.). The PA16 tag
consists of 16 amino acids (GLEGGVAMPGAEDDVV) (27). HSC-2 and SAS cells were cultured in
Dulbecco's modified Eagle's medium (DMEM; Nacalai Tesque, Inc.),
and CHO-K1 and CHO/PA16-CD44v3-10 were cultured in RPMI-1640 medium
(Nacalai Tesque, Inc.), supplemented with 10% heat-inactivated
fetal bovine serum (FBS; Thermo Fisher Scientific Inc.), 100
units/ml of penicillin, 100 µg/ml streptomycin, and 0.25 µg/ml
amphotericin B (Nacalai Tesque, Inc.) at 37°C in a humidified
atmosphere containing 5% CO2.
Antibodies
Mouse anti-CD44s mAb C44Mab-5
(IgG1, kappa) was developed as previously described
(27). Mouse IgG was purchased from
Sigma-Aldrich Corp. (Merck KGaA). To generate recombinant
C44Mab-5 (recC44Mab-5), cDNAs of
C44Mab-5 heavy and light chains were subcloned into
pCAG-Neo and pCAG-Ble vectors (FUJIFILM Wako Pure Chemical
Corporation), respectively. To generate 5-mG2a-f,
appropriate VH cDNA of mouse C44Mab-5 and
CH of mouse IgG2a were subcloned into
pCAG-Neo vector, and light chain of C44Mab-5 was
subcloned into pCAG-Ble vector. Vectors were transfected into
ExpiCHO-S or BINDS-09 (FUT8-deficient ExpiCHO-S cells) using the
ExpiCHO Expression System (28).
recC44Mab-5 and 5-mG2a-f were purified using
Protein G-Sepharose (GE Healthcare Bio-Sciences).
Animals
All animal experiments were performed in accordance
with relevant guidelines (e.g. ARRIVE guidelines) and regulations
(e.g. 3R regulations) to minimize animal suffering and distress in
the laboratory (29,30). Seventy female BALB/c nude mice (6
weeks old, 15–18 g) were purchased from Charles River (Kanagawa,
Japan). Animal studies for ADCC and antitumor activity were
approved by the Institutional Committee for Experiments of the
Institute of Microbial Chemistry (permit number: 2020-003). Mice
were maintained in a pathogen-free environment (23±2°C, 55±5%
humidity) on 11 h light/13 h dark cycle with food and water
supplied ad libitum during the experimental period. Mice
were monitored for health and weight every 1 or 5 days. Experiment
duration was three weeks. A bodyweight loss exceeding 25% and a
maximum tumor size exceeding 3,000 mm3 were identified
as humane endpoints. Mice were euthanized by cervical dislocation,
and the death was verified by respiratory arrest and cardiac
arrest.
Enzyme-linked immunosorbent assay
(ELISA)
C44Mab-5 and 5-mG2a-f were
immobilized on Nunc Maxisorp 96-well immunoplates (Thermo Fisher
Scientific Inc.) at 1 µg/ml for 30 min. After blocking using
SuperBlock buffer (Thermo Fisher Scientific Inc.) containing 0.5 mM
CaCl2, the plates were incubated with biotin-labeled
lectins, such as Aleuria aurantia lectin (AAL; Vector
Laboratories), Pholiota squarrosa lectin (PhoSL; J-OIL
MILLS, Inc.) (31), and
concanavalin A (ConA; Vector Laboratories), followed by 1:3,000
diluted peroxidase-conjugated streptavidin (Agilent Technologies).
The enzymatic reaction was produced using a 1-Step Ultra TMB-ELISA
(Thermo Fisher Scientific Inc.). The optical density was measured
at 655 nm using an iMark microplate reader (Bio-Rad Laboratories,
Inc.).
Flow cytometry
Cells were harvested by brief exposure to 0.25%
trypsin/1 mM ethylenediaminetetraacetic acid (EDTA; Nacalai Tesque,
Inc.). After washing with 0.1% bovine serum albumin (BSA) in
phosphate-buffered saline (PBS), cells were treated with primary
mAbs for 30 min at 4°C and subsequently with Alexa Fluor
488-conjugated anti-mouse IgG (1:1,000; Cell Signaling Technology,
Inc.). Fluorescence microscopy data were collected using an EC800
Cell Analyzer (Sony Corp.).
Immunohistochemical analyses
Histologic sections (4-µm thick) of an oral cancer
tissue microarray (catalogue number: OR481; US Biomax Inc.) were
directly autoclaved in citrate buffer (pH 6.0; Agilent Technologies
Inc.) for 20 min. Sections were then incubated with 1 µg/ml primary
mAbs for 1 h at room temperature and treated using an Envision+ Kit
(Agilent Technologies) for 30 min. Color was developed using
3,3′-diaminobenzidine tetrahydrochloride (DAB; Agilent Technologies
Inc.) for 2 min, and sections were then counterstained with
hematoxylin (FUJIFILM Wako Pure Chemical Corporation). Hematoxylin
and eosin (H&E) staining (FUJIFILM Wako Pure Chemical
Corporation) was performed using consecutive tissue sections. Leica
DMD108 (Leica Microsystems GmbH) was used to examine the sections
and obtain images.
Determination of the binding
affinity
Cells were suspended in 100 µl of serially diluted
mAbs (0.3 ng/ml-5 µg/ml), followed by the addition of Alexa Fluor
488-conjugated anti-mouse IgG (1:200; Cell Signaling Technology,
Inc.). Fluorescence microscopy data were collected using an EC800
Cell Analyzer (Sony Corp.). The dissociation constant
(KD) was calculated by fitting binding isotherms
to built-in one-site binding models in GraphPad PRISM 8 (GraphPad
Software, Inc.).
Western blot analysis
Cell lysates (10 µg) were boiled in sodium dodecyl
sulfate (SDS) sample buffer (Nacalai Tesque, Inc.). Proteins were
separated on 5–20% polyacrylamide gels (FUJIFILM Wako Pure Chemical
Corporation) and transferred onto polyvinylidene difluoride (PVDF)
membranes (Merck KGaA). After blocking with 4% skim milk (Nacalai
Tesque, Inc.) in PBS with 0.05% Tween 20, the membranes were
incubated with 10 µg/ml of an anti-CD44 mAb [clone
C44Mab-46 (mouse IgG1, kappa)]; available
from Antibody Bank of Tohoku University (ABTU; Miyagi, Japan);
http://www.med-tohoku-antibody.com/topics/001_paper_antibody_PDIS.htm#antiCD44)
or 1 µg/ml of anti-β-actin (clone AC-15; cat. no. A5441;
Sigma-Aldrich Corp.; Merck KGaA). This was followed by incubation
with peroxidase-conjugated anti-mouse immunoglobulins (Agilent
Technologies Inc.). Finally, protein bands were detected with
ImmunoStar LD (FUJIFILM Wako Pure Chemical Corporation) using a
Sayaca-Imager (DRC Co., Ltd.).
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNAs were prepared from cell lines using an
RNeasy Mini Prep Kit (Qiagen Inc.). The initial cDNA strand was
synthesized using SuperScript IV Reverse Transcriptase (Thermo
Fisher Scientific, Inc.) by priming nine random oligomers and an
oligo(dT) primer according to the manufacturer's instructions. We
performed 35 cycles of PCR for amplification using HotStarTaq DNA
Polymerase (Qiagen Inc.) with 0.2 µM of primer sets: Human CD44
sense (5′-GAAAGGAGCAGCACTTCAGG-3′), human CD44 antisense
(5′-ACTGCAATGCAAACTGCAAGC-3′), GAPDH sense
(5′-CAATGACCCCTTCATTGACC-3′), and GAPDH antisense
(5′-GTCTTCTGGGTGGCAGTGAT-3′).
ADCC
Six six-week-old female BALB/c nude mice were
purchased from Charles River (Kanagawa, Japan). After euthanization
by cervical dislocation, spleens were removed aseptically and
single-cell suspensions obtained by forcing spleen tissues through
a sterile cell strainer (352360, BD Falcon, Corning, Inc.) using a
syringe. Erythrocytes were lysed with a 10-sec exposure to ice-cold
distilled water. Splenocytes were washed with DMEM and resuspended
in DMEM with 10% FBS and used as effector cells. Target cells were
labeled with 10-µg/ml Calcein AM (Thermo Fisher Scientific, Inc.)
and resuspended in the same medium. The target cells
(2×104 cells/well) were plated in 96-well plates and
mixed with effector cells, anti-CD44s antibodies, or control IgG
(mouse IgG2a) (Sigma-Aldrich Corp.; Merck KGaA). After a
5-h incubation, the Calcein AM release of supernatant from each
well was measured. Fluorescence intensity was determined using a
microplate reader (Power Scan HT) (BioTek Instruments) with an
excitation wavelength of 485 nm and an emission wavelength of 538
nm. Cytolytic activity (as % of lysis) was calculated using the
equation: % lysis=(E-S)/(M-S) ×100, where E is the fluorescence of
the combined target and effector cells, S is the spontaneous
fluorescence of the target cells only, and M is the maximum
fluorescence measured after lysing all cells with a buffer
containing 0.5% Triton X-100, 10 mM Tris-HCl (pH 7.4), and 10 mM of
EDTA.
CDC
Cells in DMEM supplemented with 10% FBS were plated
in 96-well plates (2×104 cells/well), and incubated for
5 h at 37°C with either anti-CD44s antibodies or control IgG (mouse
IgG2a) (Sigma-Aldrich Corp.; Merck KGaA) and 10% rabbit
complement (Low-Tox-M Rabbit Complement; Cedarlane Laboratories).
To assess cell viability, an MTS
[3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium;
inner salt] assay was performed using a CellTiter
96®AQueous assay kit (Promega Corp.).
3D cell proliferation assay
3D cell proliferation was measured with the
CellTiter-Glo® 3D cell viability assay (Promega Corp.)
according to the manufacturer's instructions. Briefly, the cells
were plated (2,000 cells/100 µl/well) in triplicate in 96-well
ultra low attachment plates (Corning Inc.) with PBS or 100 µg/ml of
mouse IgG2a and an anti-CD44 mAb (5-mG2a-f)
in DMEM containing 10% FBS. The cell viability was measured after
48 h of incubation. The CellTiter-Glo® 3D reagent was
added into wells in a 1:1 dilution (100 µl volume in well:100 µl of
reagent) and then the plates were shaken for 5 min on an orbital
shaker and incubated at room temperature for an additional 25 min.
The luminescent signal was read using an EnSpire multi-plate reader
(Perkin Elmer). Images were taken using an Evolution MP camera
(Media Cybernetics). The proliferation rate was calculated relative
to the control (PBS was added instead of the antibodies).
Antitumor activity of
5-mG2a-f in the xenografts of oral cancers
Sixty-four six-week-old female BALB/c nude mice were
purchased from Charles River (Kanagawa, Japan) and used at 10 weeks
of age. HSC-2 and SAS cells (0.3 ml of 1.33×108 cells/ml
in DMEM) were mixed with 0.5 ml BD Matrigel Matrix Growth Factor
Reduced (BD Biosciences). One hundred microliters of this
suspension (5×106 cells) was injected subcutaneously
into the left flank. After day 1 (protocol-1) or day 7
(protocol-2), 100 µg of 5-mG2a-f and control mouse IgG
(Sigma-Aldrich Corp.; Merck KGaA) in 100 µl PBS were injected
intraperitoneally (i.p.) into treated and control mice,
respectively. Additional antibodies were then injected on days 7
and 14 (protocol-1) or on days 14 and 21 (protocol-2). Nineteen
days (protocol-1) or 27 days (protocol-2) after cell implantation,
all mice were euthanized by cervical dislocation and tumor
diameters and volumes were determined as previously described
(32).
Statistical analyses
All data are expressed as mean ± standard error of
the mean (SEM). Statistical analysis was carried out using ANOVA
following Tukey-Kramer's test for ADCC and CDC. Sidak's multiple
comparisons test was used for tumor volume and mouse weight, or
Welch's t test for tumor weight and 3D cell proliferation assay
using GraphPad Prism 7 (GraphPad Software, Inc.). P<0.05 was
adopted as a level of statistical significance.
Results
Production and characterization of
5-mG2a-f, a core-fucose-deficient mouse
IgG2a-type anti-CD44 antibody
As mouse IgG2a possesses high ADCC and
CDC activities (33), we first
produced a mouse IgG2a version of mouse IgG1
C44Mab-5 by subcloning appropriate VH cDNA of
C44Mab-5 and CH of mouse IgG2a
into pCAG-Neo vector, and light chain of C44Mab-5 into
pCAG-Ble vector. This IgG2a-type of C44Mab-5
is henceforth referred to as 5-mG2a. We additionally
produced a core-fucose-deficient type of 5-mG2a,
henceforth referred to as 5-mG2a-f, using the BINDS-09
cell line (FUT8-knockout Expi-CHO-S cell line) (28). Defucosylation of 5-mG2a-f
was confirmed using lectins such as Aleuria aurantia lectin
(AAL, fucose binder) (34) and
Pholiota squarrosa lectin (PhoSL, core fucose binder)
(31). Concanavalin A (ConA,
mannose binder) (35) was used as a
control. Both C44Mab-5 and 5-mG2a-f were
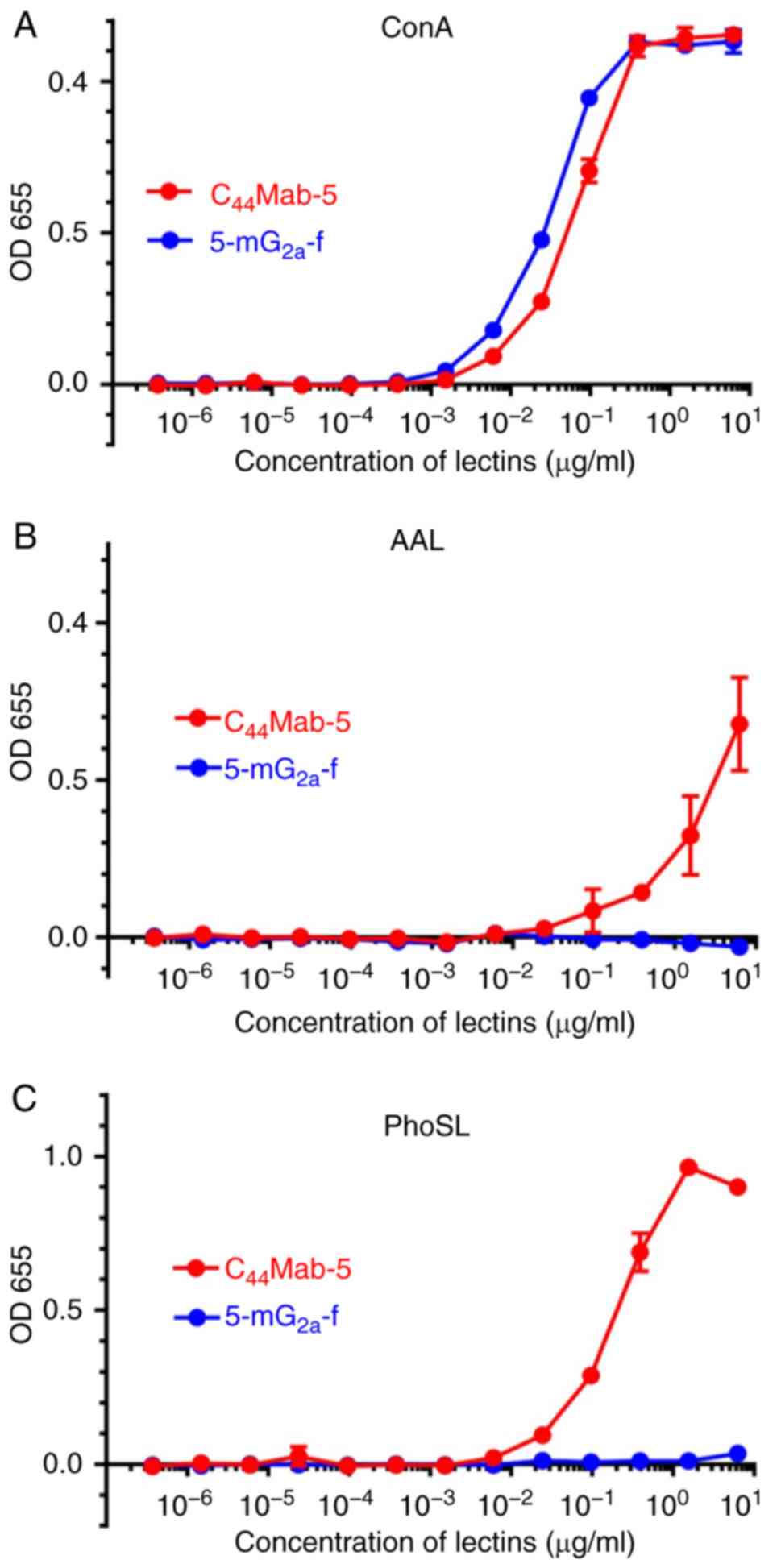
detected using ConA (Fig. 1A).
C44Mab-5, but not 5-mG2a-f, was detected
using AAL (Fig. 1B) or PhoSL
(Fig. 1C), indicating that
5-mG2a-f was defucosylated.
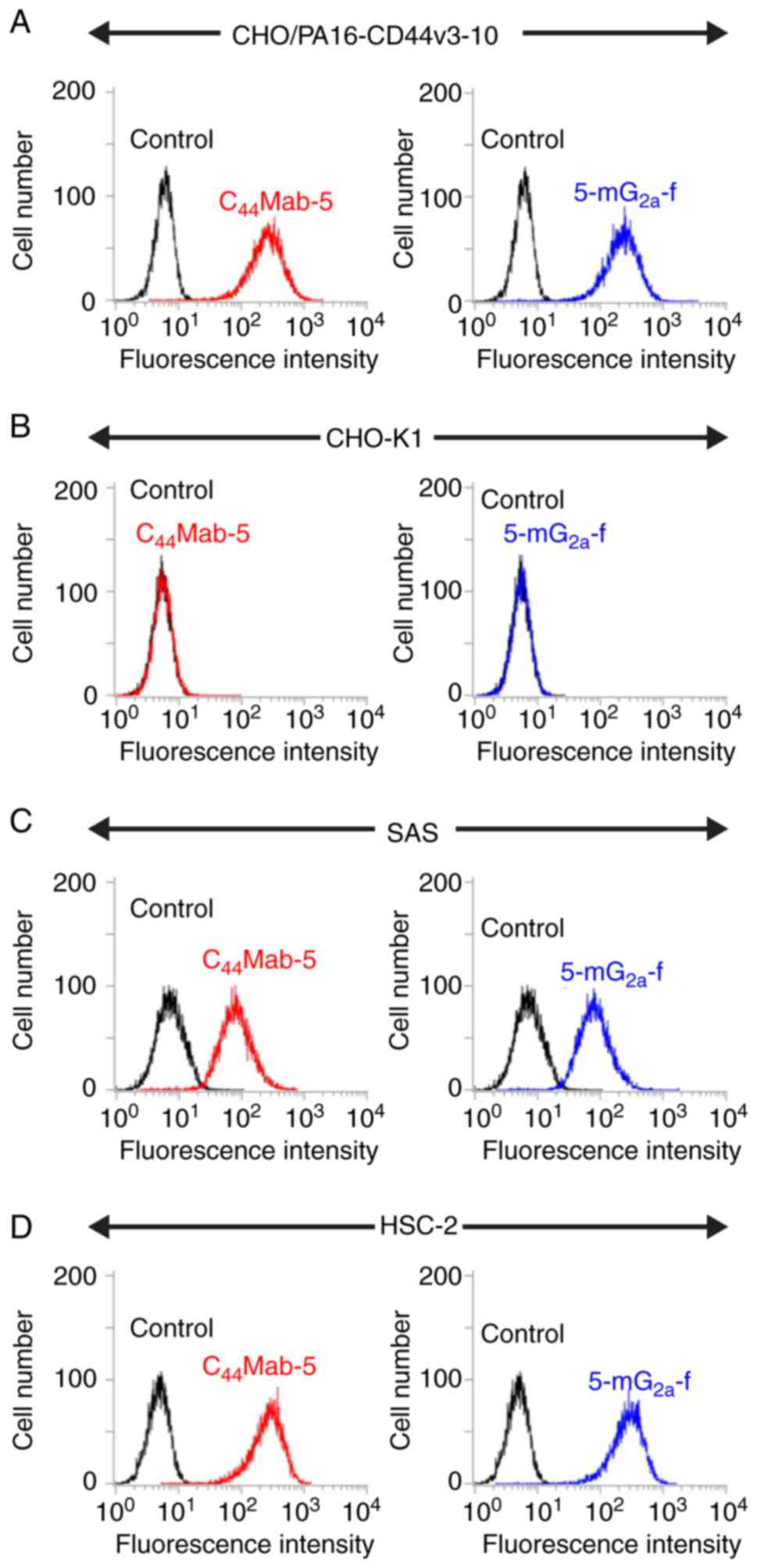
We examined the sensitivity of 5-mG2a-f
in CHO cells expressing CD44v3-10 plus N-terminal PA16 tag
(CHO/PA16-CD44v3-10) and in oral squamous cell carcinoma (OSCC)
cell lines (SAS and HSC-2) using flow cytometry. Both
C44Mab-5 and 5-mG2a-f reacted with
CHO/PA16-CD44v3-10 cells (Fig. 2A),
but not with CHO-K1 cells (Fig.
2B). Both C44Mab-5 and 5-mG2a-f reacted
with SAS cells (Fig. 2C) and HSC-2
cells (Fig. 2D), indicating that
both mAbs showed high sensitivity against SAS and HSC-2 cells.
As shown in Fig.
S1A, CD44 was not detected by an anti-CD44s mAb
(C44Mab-46) in both SAS and HSC-2 cells presumably
because the CD44 expression level in those cells might be low for
the detection in western blot analysis. Then, we performed RT-PCR
analysis for detection of CD44. As shown in Fig. S1B, the multiple bands of CD44v were
detected in SAS and HSC-2 cells using PCR, indicating that CD44v is
expressed in SAS and HSC-2 cells.
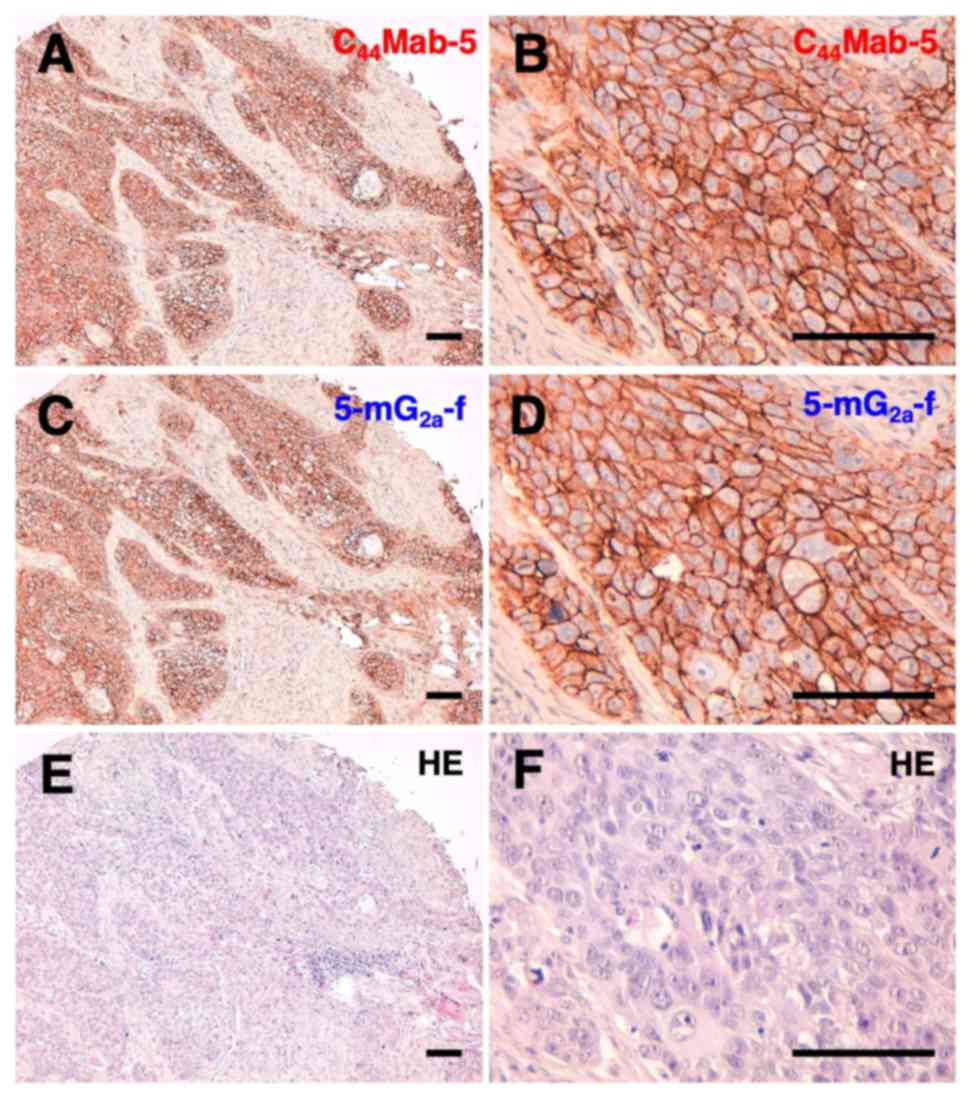
Next, we performed immunohistochemical analysis on
oral cancer cell lines. Representative images are shown in Fig. 3. Both C44Mab-5 (Fig. 3A and B) and 5-mG2a-f
(Fig. 3C and D) stained the plasma
membrane of oral cancer cells. The sensitivity of
5-mG2a-f was similar with that of C44Mab-5.
Both C44Mab-5 and 5-mG2a-f stained 33/38
cases (86.8%) of OSCCs of the tissue microarray. Hematoxylin &
eosin (H&E) staining of consecutive tissue sections of OSCC is
shown in Fig. 3E and F.
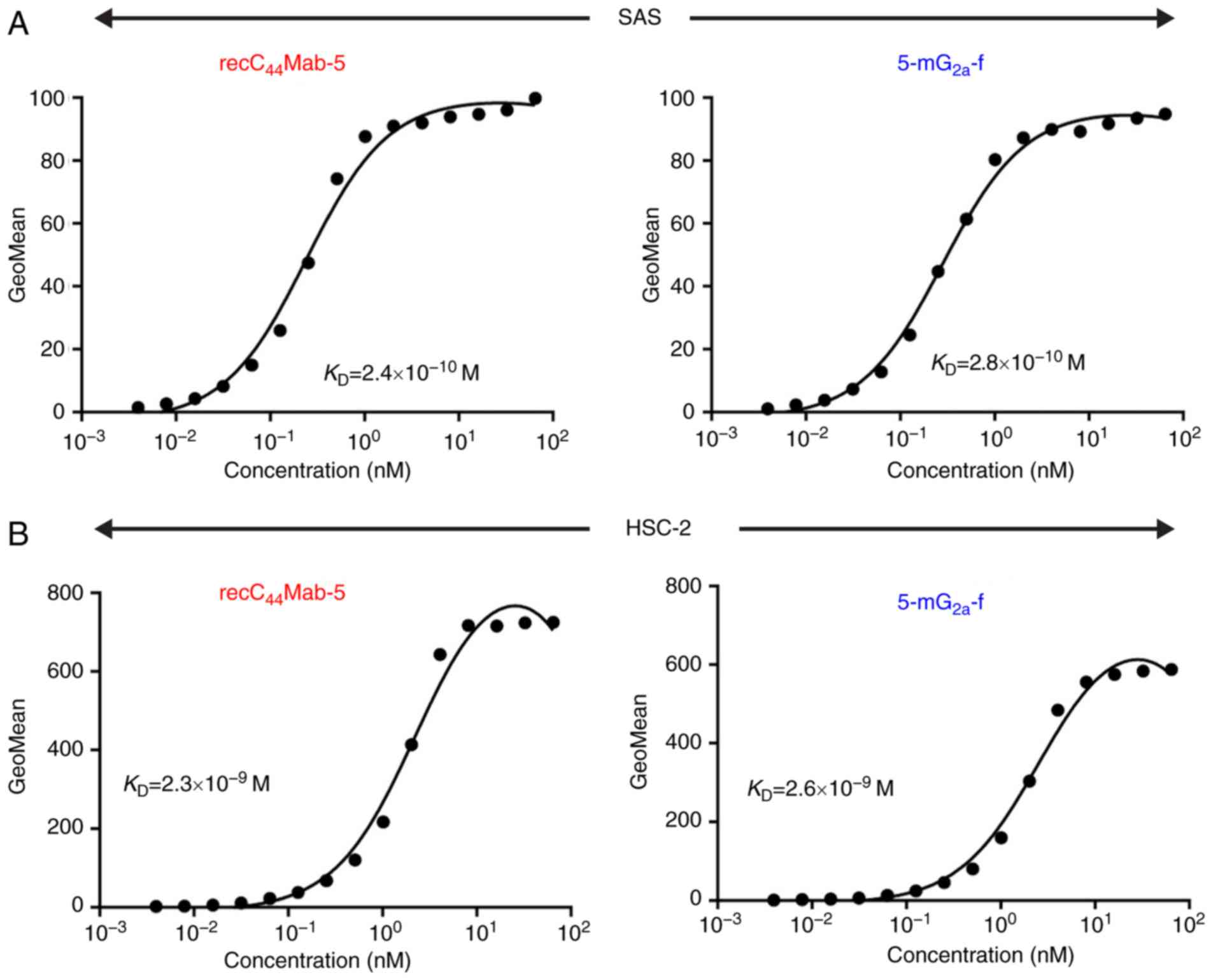
We performed a kinetic analysis of the interactions
of recC44Mab-5 and 5-mG2a-f with SAS and
HSC-2 oral cancer cell lines using flow cytometry. As shown in
Fig. 4A, the dissociation constant
(KD) for recC44Mab-5 in SAS cells was
2.4×10−10 M. In contrast, the KD for
5-mG2a-f in SAS cells was 2.8×10−10 M
(Fig. 4A). The binding affinity of
5-mG2a-f in SAS cells was very similar to that of
recC44Mab-5. Likewise, the KD for
recC44Mab-5 against HSC-2 was 2.3×10−9 M
(Fig. 4B). In contrast, the
KD for 5-mG2a-f in HSC-2 cells was
2.6×10−9 M (Fig. 4B).
The binding affinity of 5-mG2a-f in HSC-2 cells was very
similar to that of recC44Mab-5. The binding affinity of
5-mG2a-f in SAS was 9.3-fold higher than that against
HSC-2.
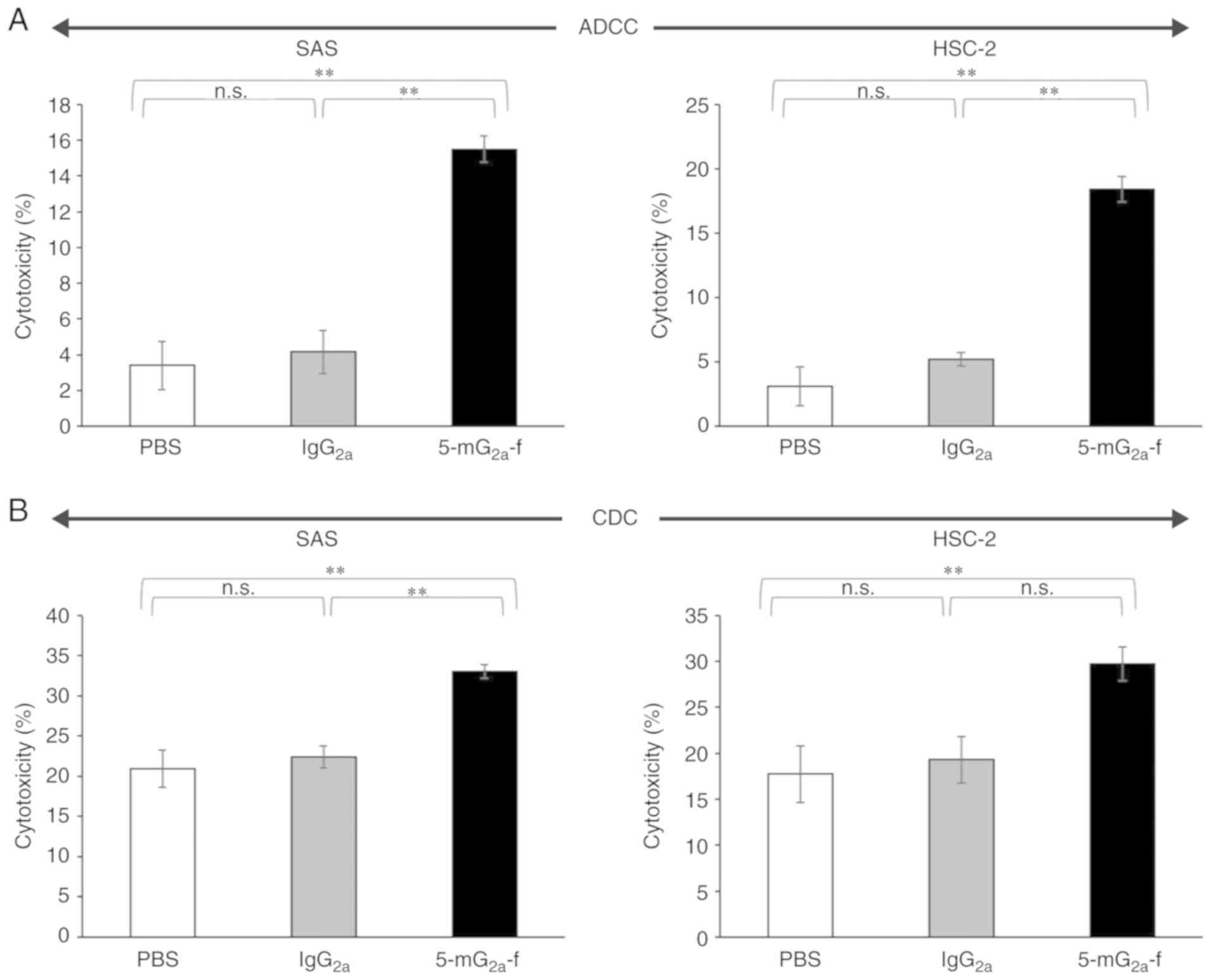
ADCC and CDC activities of
5-mG2a-f in oral cancer cell lines
Because the mouse IgG1 subclass of
C44Mab-5 does not possess ADCC or CDC activities, we
synthesized a mouse IgG2a subclass mAb, and further
defucosylated it to augment those activities. In this study, we
examined whether the developed 5-mG2a-f induced ADCC and
CDC in CD44-expressing oral cancer cell lines, such as SAS and
HSC-2 cells. As shown in Fig. 5A,
5-mG2a-f exhibited higher ADCC (16% cytotoxicity) in SAS
cells compared with that of control PBS (3.4% cytotoxicity;
P<0.01) and control mouse IgG2a treatment (4.2%
cytotoxicity; P<0.01). Similarly, 5-mG2a-f exhibited
higher ADCC (18% cytotoxicity) against HSC-2 cells compared with
that of control PBS (3.1% cytotoxicity; P<0.01) and control
mouse IgG2a treatment (5.2% cytotoxicity; P<0.01),
indicating that ADCC in SAS cells is similar with that in HSC-2
cells, despite the binding affinity of 5-mG2a-f in SAS
cells being 9.3-fold higher than in HSC-2 cells (Fig. 4). As shown in Fig. 5B, 5-mG2a-f exhibited
slightly higher CDC (33% cytotoxicity) in SAS cells compared with
control PBS (21% cytotoxicity; P<0.01) and control mouse
IgG2a treatment (22% cytotoxicity; P<0.01).
Similarly, 5-mG2a-f exhibited slightly higher CDC (30%
cytotoxicity) in HSC-2 cells compared with control PBS (18%
cytotoxicity; P<0.01) and control mouse IgG2a
treatment (19% cytotoxicity; not significant). Although ADCC/CDC
activities of 5-mG2a-f in oral cancer cells are not
outstanding, 5-mG2a-f may exert antitumor activity
against oral cancer cells in vivo.
The influence of 5-mG2a-f
in oral cancer cell lines in anchorage-independent condition
Next, we investigated whether 5-mG2a-f
inhibits cell growth of SAS and HSC-2 cells in
anchorage-independent condition. As shown in Fig. S2A, both SAS and HSC-2 cells grew in
anchorage-independent condition for 48 h. In contrast, an anti-CD44
mAb (5-mG2a-f) did not inhibit the growth of SAS or
HSC-2 compared to control mouse IgG2a (Fig. S2B), indicating that
5-mG2a-f did not affect the cell growth of oral cancer
cell lines in anchorage-independent condition.
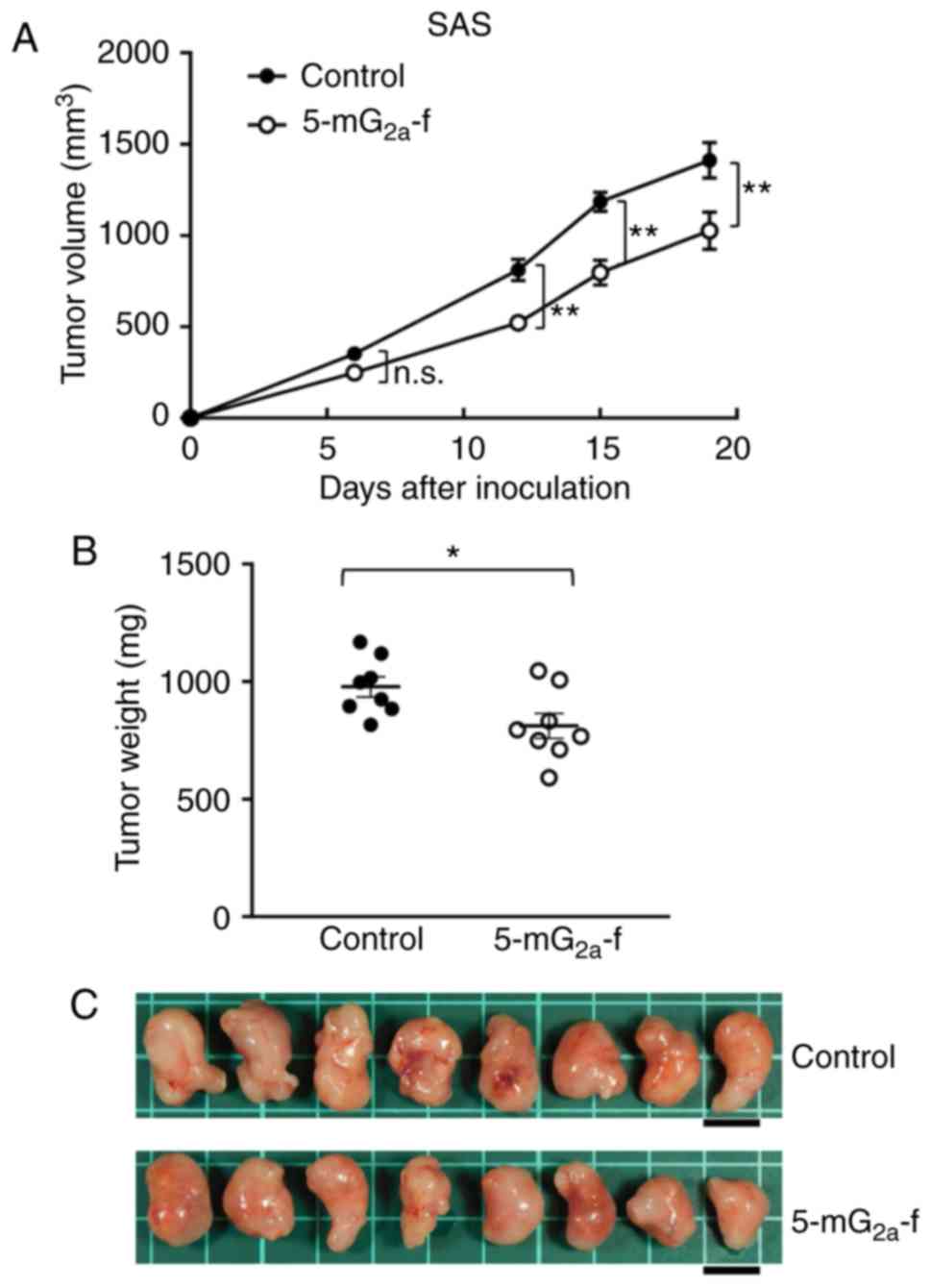
Antitumor activities of
5-mG2a-f in the mouse xenografts of SAS oral cancer
cells
SAS cells were subcutaneously implanted into the
flanks of nude mice. In protocol-1, 5-mG2a-f (100 µg)
and control mouse IgG (100 µg) were injected i.p. three times into
the mice, on days 1, 7, and 14 after SAS cell injections. Tumor
volume was measured on days 6, 12, 15, and 19. Tumor development
was significantly reduced in the 5-mG2a-f-treated mice
on days 12, 15, and 19 in comparison to the IgG-treated control
mice (Fig. 6A). Tumor volume on day
19 was reduced by 27% after 5-mG2a-f treatment. Tumors
from 5-mG2a-f-treated mice weighed significantly less
than tumors from IgG-treated control mice (16.9% reduction,
Fig. 6B). Resected tumors on day 19
are depicted in Fig. 6C. Control
and 5-mG2a-f-treated SAS xenograft mice are shown on day
19 in Fig. S3A and B,
respectively. Total body weights did not significantly differ
between the two groups (Fig.
S3C).
In protocol-2 of the SAS xenograft models, tumor
formation of 16 SAS-bearing mice was observed on day 7. Then, these
16 SAS-bearing mice were divided into a 5-mG2a-f-treated
group and a control group. On days 7, 14, and 21 after SAS cell
injections into the mice, 5-mG2a-f (100 µg) and control
mouse IgG (100 µg) were injected i.p. into the mice. Tumor
formation was observed in mice in both treated and control groups.
Tumor volume was measured on days 7, 12, 15, 19, 22, and 27.
5-mG2a-f-treated mice displayed significantly reduced
tumor development on days 22 and 27 in comparison to IgG-treated
control mice (Fig. 7A). Tumor
volume reduction by 5-mG2a-f was 43% on day 27. Tumors
from the 5-mG2a-f-treated mice weighed significantly
less than tumors from the IgG-treated control mice (27.1%
reduction, Fig. 7B). Resected
tumors on day 27 are depicted in Fig.
7C. Control and 5-mG2a-f-treated SAS xenograft mice
are shown on day 27 in Fig. S4A and
B, respectively. Total body weights did not significantly
differ between the two groups (Fig.
S4C). These results indicate that 5-mG2a-f reduced
the growth of SAS xenografts effectively, even when
5-mG2a-f was injected 7 days post-SAS cell injections in
mice.
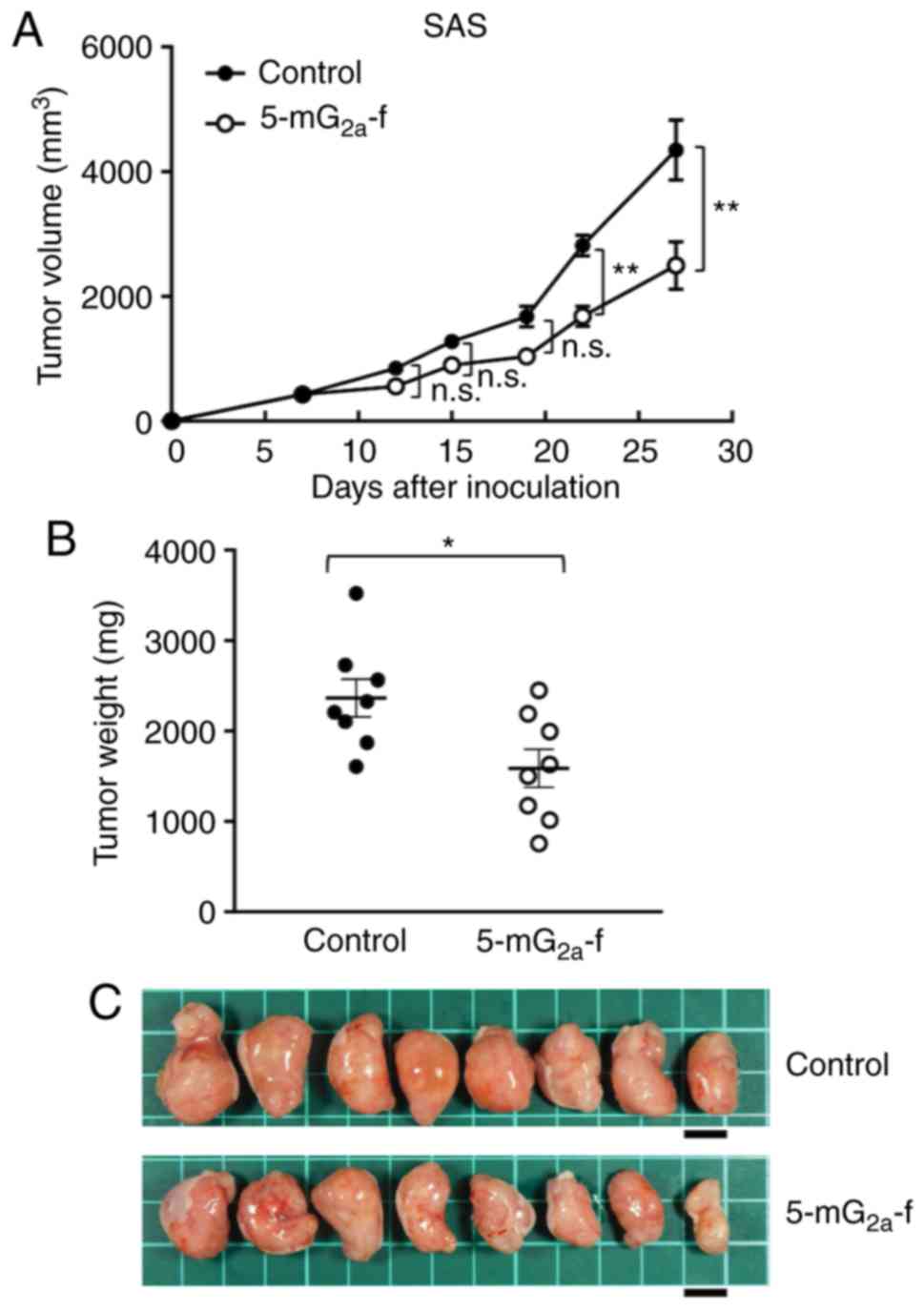 | Figure 7.Evaluation of antitumor activity of
5-mG2a-f (from day 7) in SAS xenografts. (A) SAS cells
(5×106 cells) were injected subcutaneously into the left
flank. After day 7, 100 µg of 5-mG2a-f and control mouse
IgG in 100 µl PBS were injected i.p. into treated and control mice,
respectively. Additional antibodies were then injected on days 14
and 21. Tumor volume was measured on days 7, 12, 15, 19, 22, and
27. Values are mean ± SEM. Asterisk indicates statistical
significance (**P<0.01; n.s., not significant; ANOVA and Sidak's
multiple comparisons test). (B) Tumors of SAS xenografts were
resected from 5-mG2a-f and control mouse IgG groups.
Tumor weight on day 27 was measured from excised xenografts. Values
are mean ± SEM. Asterisk indicates statistical significance
(*P<0.05, Welch's t test). (C) Resected tumors of SAS xenografts
from 5-mG2a-f and control mouse IgG groups on day 27.
Scale bar, 1 cm. |
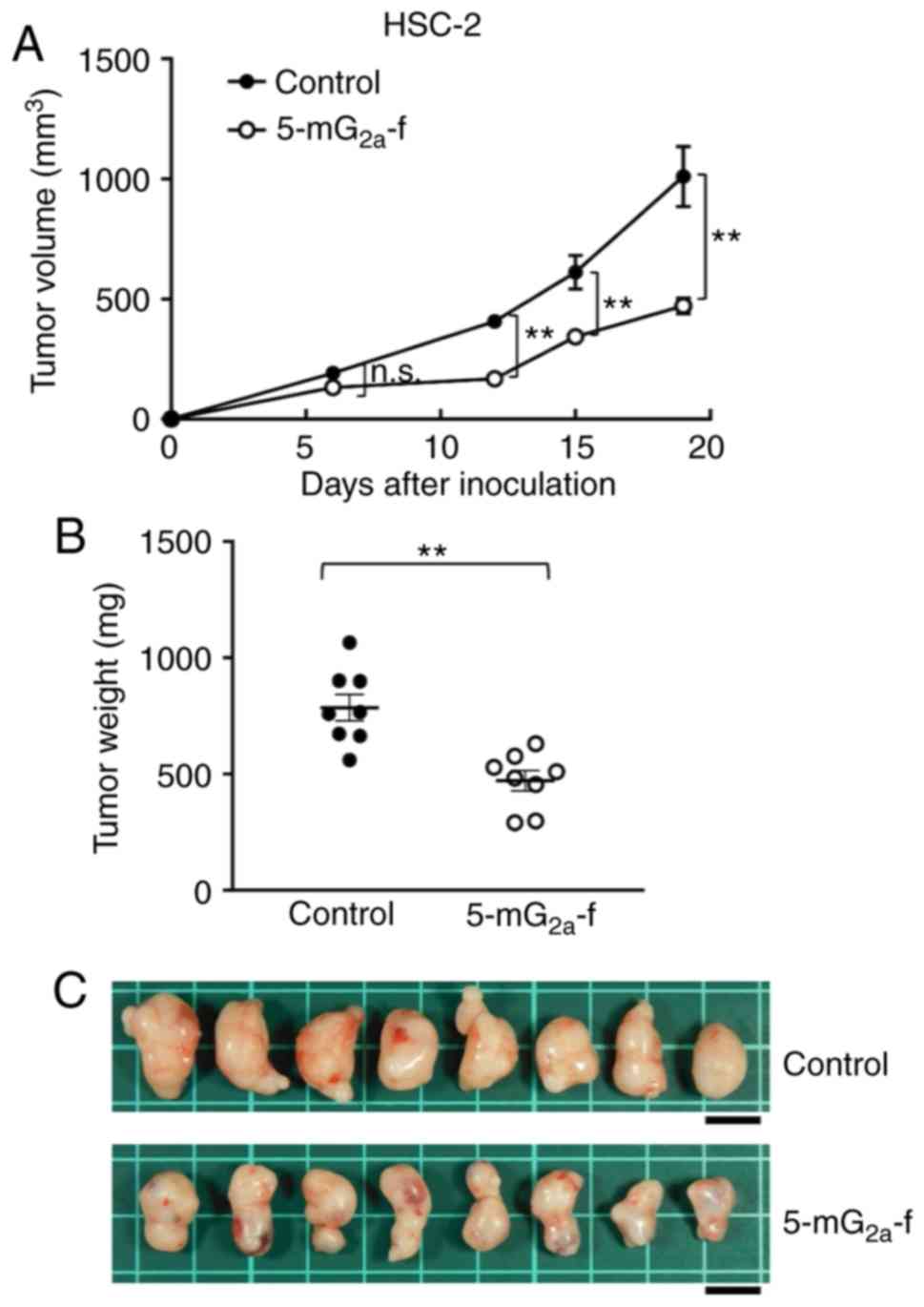
Antitumor activities of
5-mG2a-f in mouse xenografts of HSC-2 oral cancer
cells
In a second xenograft model of oral cancers, HSC-2
cells were subcutaneously implanted into the flanks of nude mice.
In protocol-1 of HSC-2 ×enograft models, 5-mG2a-f (100
µg) and control mouse IgG (100 µg) were injected i.p. three times
into the mice, on days 1, 7, and 14 after HSC-2 cell injections
into the mice. Tumor volume was measured on days 6, 12, 15, and 19.
5-mG2a-f-treated mice displayed significantly reduced
tumor development on days 12, 15, and 19 in comparison to
IgG-treated control mice (Fig. 8A).
Tumor volume reduction by 5-mG2a-f was 53% on day 19.
Tumors from 5-mG2a-f-treated mice weighed significantly
less than HSC-2 tumors from IgG-treated control mice (44.1%
reduction, Fig. 8B). Resected
tumors on day 19 are depicted in Fig.
8C. Control and 5-mG2a-f-treated HSC-2 ×enograft
mice are shown on day 19 in Fig. S5A
and B, respectively. Total body weights did not significantly
differ between the two groups (Fig.
S5C).
In protocol-2 of the HSC-2 ×enograft models, tumor
formation of 16 HSC-2-bearing mice was observed on day 7. Then,
these 16 HSC-2-bearing mice were divided into a
5-mG2a-f-treated group and a control group. On days 7,
14, and 21 after cell injections into the mice, 5-mG2a-f
(100 µg) and control mouse IgG (100 µg) were injected i.p. into the
mice. Tumor volume was measured on days 7, 12, 15, 19, 22, and 27.
5-mG2a-f-treated mice displayed significantly reduced
tumor development on days 22 and 27 in comparison to IgG-treated
control mice (Fig. 9A). Tumor
volume reduction in 5-mG2a-treated mice was 32% on day
27. Tumors from 5-mG2a-f-treated mice weighed
significantly less than tumors from IgG-treated control mice (27.1%
reduction, Fig. 9B). Resected
tumors on day 27 are depicted in Fig.
9C. Control and 5-mG2a-f-treated HSC-2 ×enograft
mice are shown on day 27 in Fig. S6A
and B, respectively. Total body weights did not significantly
differ between the two groups (Fig.
S6C). These results indicate that 5-mG2a-f reduced
the growth of HSC-2 ×enografts effectively, even when
5-mG2a-f was injected 7 days post-HSC-2 cell injections
in mice.
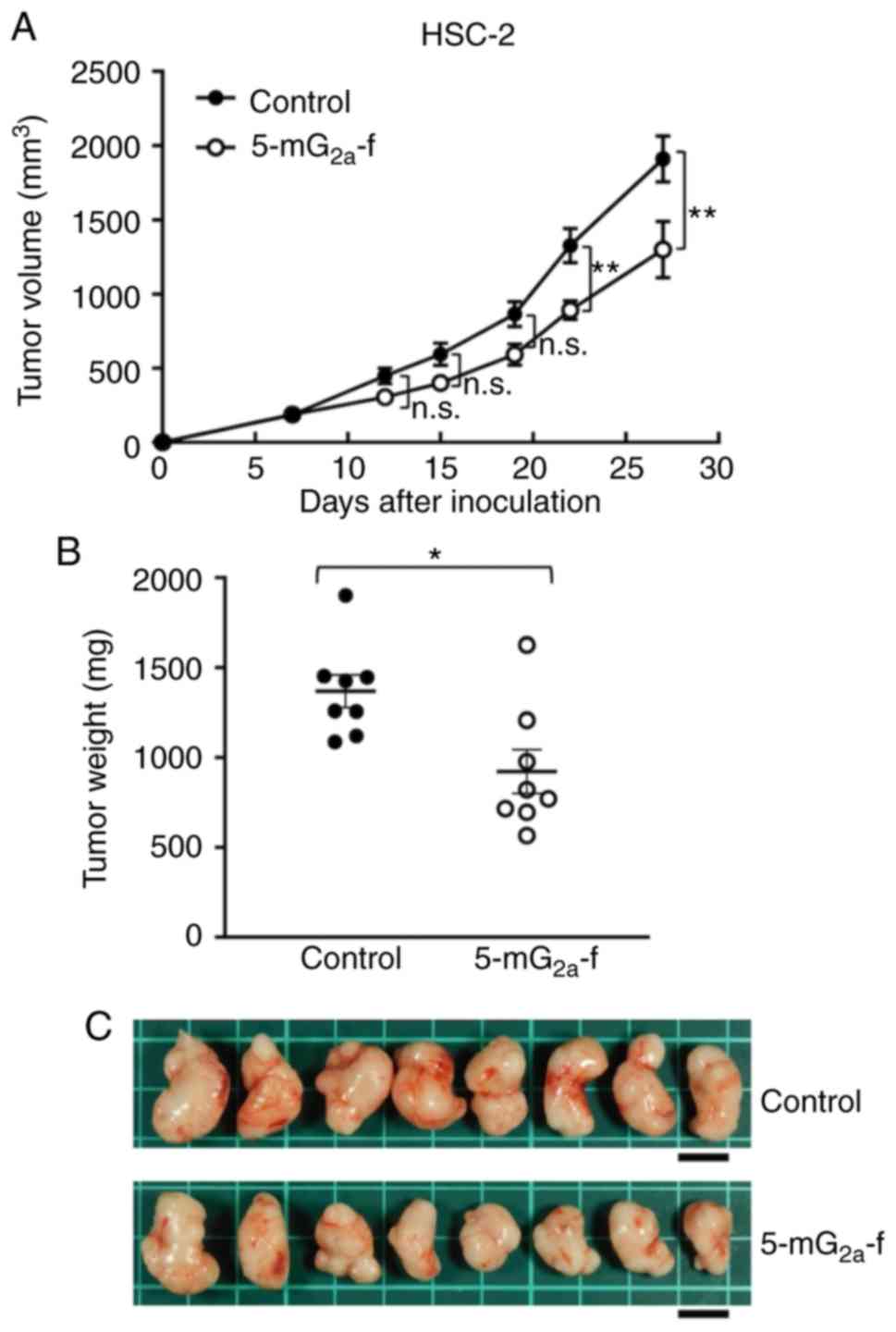 | Figure 9.Evaluation of antitumor activity of
5-mG2a-f (from day 7) in HSC-2 ×enografts. (A) HSC-2
cells (5×106 cells) were injected subcutaneously into
the left flank. After day 7, 100 µg of 5-mG2a-f and
control mouse IgG in 100 µl PBS were injected i.p. into treated and
control mice, respectively. Additional antibodies were then
injected on days 14 and 21. Tumor volume was measured on days 7,
12, 15, 19, 22, and 27. Values are mean ± SEM. Asterisk indicates
statistical significance (**P<0.01; n.s., not significant; ANOVA
and Sidak's multiple comparisons test). (B) Tumors of HSC-2
×enografts were resected from 5-mG2a-f and control mouse
IgG groups. Tumor weight on day 27 was measured from excised
xenografts. Values are mean ± SEM. Asterisk indicates statistical
significance (*P<0.05, Welch's t test). (C) Resected tumors of
HSC-2 ×enografts from 5-mG2a-f and control mouse IgG
groups on day 27. Scale bar, 1 cm. |
Discussion
In the present study, we investigated whether
anti-CD44 mAbs are advantageous for the treatment of oral cancers.
We previously developed a sensitive and specific anti-CD44 mAb,
C44Mab-5, but were unable to demonstrate antitumor
activity because the IgG1 subclass does not possess
ADCC/CDC activities (27). In this
study, we developed this antibody into an IgG2a subclass
antibody, and augmented ADCC activity using a defucosylated
variant. Oral cancers comprise several histological tumor types,
such as squamous cell carcinoma (SCC), adenocarcinoma,
mucoepidermoid carcinoma, and adenoid cystic carcinoma. Among
these, SCC accounts for over 90% of all oral cancers (36). Therefore, we used SAS and HSC-2 cell
lines, which are derived from SCC, and investigated ADCC/CDC and
antitumor activities.
The most effective treatment of OSCC depends upon
its clinical stage. Stage-I and -II (early stages) are treated via
surgery or radiotherapy alone. In contrast, stage-III and -IV
(advanced stages) require a combination of surgery, radiotherapy,
and chemotherapy (37). For
chemotherapy of OSCCs, cisplatin is mainly used, and is often
combined with docetaxel and 5-fluorouracil (38,39).
Other anticancer agents, such as paclitaxel, carboplatin, and
methotrexate can be also used for OSCCs (40), but effective molecular targeting
drugs, such as antibody therapies, are limited.
Recently, cetuximab, a mouse-human chimeric mAb
(IgG1) that targets epidermal growth factor receptor
(EGFR), was approved by the Food and Drug Administration (FDA) in
the USA for treatment of oral cancers (41). Cetuximab has been shown effective
against locoregionally advanced head and neck cancer and recurrent
or metastatic squamous cell carcinoma of the head and neck
(41–43). Although advances in diagnosis and
therapeutic techniques have improved the overall 5-year survival
rate to 70%, the 5-year survival rate in stage IV is only 40%;
therefore, further treatments must be developed (44). In our recent study, we also
developed a sensitive and specific mAb (EMab-17) against EGFR, and
demonstrated its ADCC/CDC and antitumor activity against SAS and
HSC-2 ×enografts (32). Although we
showed that EMab-17 could potentially be used for antibody-based
therapy for EGFR-expressing OSCC, the difference between cetuximab
and EMab-17 has not been clarified. Several studies characterizing
EMab-17, including epitope mapping and signal induction in OSCC
cells, are currently ongoing.
In another recent study, HER2 was shown to be
expressed in oral cancers, and an anti-HER2 mAb
(H2Mab-19) demonstrated antitumor activity (45). Therefore, anti-HER2 therapies using
trastuzumab could be effective for the treatment of oral cancers.
HER2 expression was reported in only 1.4% of immunohistochemical
analyses of oral cancer (46),
although it is expressed in 10.4% of breast cancers (47). Therefore, targeting only HER2 may be
insufficient for conquering oral cancers. As antitumor effects of
combined gefitinib and trastuzumab or cetuximab and trastuzumab
treatment on head and neck SCC (HNSCC) were demonstrated in
vitro (48,49), those few oral cancer patients
displaying HER2 overexpression/amplification may possibly benefit
from anti-HER2 therapy.
Furthermore, we previously investigated whether
podocalyxin (PODXL) may be a therapeutic target in OSCC using
anti-PODXL mAbs (50). We
engineered an anti-PODXL mAb of IgG1 subclass (PcMab-47)
into a mouse IgG2a-type mAb (47-mG2a) to
increase ADCC. We further developed 47-mG2a-f, a core
fucose-deficient variant of 47-mG2a to further augment
its ADCC. In vivo analysis revealed that
47-mG2a-f, but not 47-mG2a, exhibited
antitumor activity in SAS and HSC-2 ×enograft models at a dose of
100 µg/mouse/week administered three times. Although both
47-mG2a and 47-mG2a-f exhibited antitumor
activity in HSC-2 ×enograft models at a dose of 500 µg/mouse/week
administered twice, 47-mG2a-f also demonstrated higher
antitumor activity than 47-mG2a, indicating that a core
fucose-deficient anti-PODXL mAb could be useful for antibody-based
therapy against PODXL-expressing OSCCs. Therefore, we used the
core-fucose-deficient anti-CD44 mAb (5-mG2a-f) for
treating CD44-expressing oral cancers.
In this study, we demonstrated that
5-mG2a-f exerts ADCC/CDC activities in vitro, and
antitumor activities in vivo. Importantly,
5-mG2a-f effectively reduced the growth of SAS and HSC-2
×enografts, even when 5-mG2a-f was injected 7 days after
cell implantations into the mice. However, tumor volume reduction
of SAS and HSC-2 on day 27 by 5-mG2a-f was still only 43
and 32%, respectively, indicating that anti-CD44 therapy might not
be robust enough for conquering most oral cancers. One potential
reason for this weak antitumor activity is the lower ADCC activity
and CDC activity of 5-mG2a-f, despite high binding
activity in SAS cells (KD: 2.8×10−10
M) and HSC-2 cells (KD: 2.6×10−9
M).
In our previous report, C44Mab-5
detected CD44s (27). Although the
binding epitope of C44Mab-5 may potentially be located
between exon-1 and exon-5, we have not been able to determine the
exact binding epitope of C44Mab-5, likely because
C44Mab-5 recognizes the tertiary structure of CD44
rather simple peptides or glycans. Because the binding epitope is
critical for ADCC/CDC activities of mAbs, other anti-CD44 mAbs of
various epitopes will need to be developed in future studies.
Targeting multiple targets, such as EGFR, HER2,
PODXL, and CD44 may be needed for effective therapy to conquer oral
cancers. Another important goal is the targeting of cancer-specific
antigens using a cancer-specific mAb (CasMab) because EGFR, HER2,
PODXL, and CD44 are widely expressed in normal tissues. We
previously established CasMab against podoplanin (PDPN), which is
expressed in many cancers, including oral cancers (51–54).
In xenograft models with HSC-2 cells, a mouse-human chimeric mAb,
chLpMab-23, exerted antitumor activity using human natural killer
cells, indicating that chLpMab-23 may be useful for antibody
therapy against PDPN-expressing oral cancers (54). In the future study, cancer-specific
anti-CD44 mAbs may also be developed that can reduce the adverse
effects of traditional antibody therapy.
Supplementary Material
Supporting Data
Acknowledgements
We thank Ms. Saori Handa and Mr. Yu Komatsu
(Department of Antibody Drug Development, Tohoku University
Graduate School of Medicine) for technical assistance concerning
the in vitro experiments, and Ms. Akiko Harakawa [Institute
of Microbial Chemistry (BIKAKEN), Numazu, Microbial Chemistry
Research Foundation] for technical assistance regarding the animal
experiments.
Funding
This research was supported in part by Japan Agency
for Medical Research and Development (AMED) under grant nos.
JP20am0401013 (YK), JP20am0101078 (YK), and JP20ae0101028 (YK), and
by the Japan Society for the Promotion of Science (JSPS)
Grants-in-Aid for Scientific Research (KAKENHI) grant nos. 17K07299
(MKK), 19K07705 (YK), and 20K16322 (MS).
Availability of data and materials
The datasets used and/or analyzed during the study
are available from the corresponding author on reasonable
request.
Authors' contributions
HHo, JT, TO, TN, MS, TA, YS, and MY performed the
experiments. MKK analyzed the experimental data. MK, HHa, and YK
designed the current study and wrote the manuscript. All authors
read and approved the manuscript and agree to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
Animal studies for ADCC and the antitumor activity
were approved by the Institutional Committee for Experiments of the
Institute of Microbial Chemistry (Numazu-shi, Shizuoka, Japan)
(permit no. 2020-003).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
AAL
|
Aleuria aurantia lectin
|
|
ABTU
|
Antibody Bank of Tohoku
University
|
|
ADCC
|
antibody-dependent cellular
cytotoxicity
|
|
ATCC
|
American Type Culture Collection
|
|
BSA
|
bovine serum albumin
|
|
CasMab
|
cancer-specific mAb
|
|
CBIS
|
cell-based immunization and
screening
|
|
CDC
|
complement-dependent cytotoxicity
|
|
CD44s
|
CD44 standard
|
|
CD44v
|
CD44 variant
|
|
CHO
|
Chinese hamster ovary
|
|
Con A
|
concanavalin A
|
|
DMEM
|
Dulbecco's modified Eagle's
medium
|
|
EDTA
|
ethylenediaminetetraacetic acid
|
|
ELISA
|
enzyme-linked immunosorbent assay
|
|
FBS
|
fetal bovine serum
|
|
FDA
|
Food and Drug Administration
|
|
FGF
|
fibroblast growth factor
|
|
HNSCC
|
head and neck squamous cell
carcinoma
|
|
HB-EGF
|
heparin-binding epidermal growth
factor
|
|
JCRB
|
Japanese Collection of Research
Bioresources Cell Bank
|
|
mAb
|
monoclonal antibody
|
|
OSCC
|
oral squamous cell carcinoma
|
|
PBS
|
phosphate-buffered saline
|
|
PDPN
|
podoplanin
|
|
PhoSL
|
Pholiota squarrosa lectin
|
|
PODXL
|
podocalyxin
|
|
PVDF
|
polyvinylidene difluoride
|
|
RTKs
|
receptor tyrosine kinases
|
|
RT-PCR
|
reverse transcription-polymerase
chain reaction
|
|
SCC
|
squamous cell carcinoma
|
|
SDS
|
sodium dodecyl sulfate
|
|
SEM
|
standard error of the mean
|
|
VEGFR
|
vascular endothelial growth factor
receptor
|
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hashibe M, Brennan P, Chuang SC, Boccia S,
Castellsague X, Chen C, Curado MP, Dal Maso L, Daudt AW, Fabianova
E, et al: Interaction between tobacco and alcohol use and the risk
of head and neck cancer: Pooled analysis in the international head
and neck cancer epidemiology consortium. Cancer Epidemiol
Biomarkers Prev. 18:541–550. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tota JE, Anderson WF, Coffey C, Califano
J, Cozen W, Ferris RL, St John M, Cohen EE and Chaturvedi AK:
Rising incidence of oral tongue cancer among white men and women in
the United States, 1973–2012. Oral Oncol. 67:146–152. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hussein AA, Helder MN, de Visscher JG,
Leemans CR, Braakhuis BJ, de Vet HCW and Forouzanfar T: Global
incidence of oral and oropharynx cancer in patients younger than 45
years versus older patients: A systematic review. Eur J Cancer.
82:115–127. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sherman L, Sleeman J, Herrlich P and Ponta
H: Hyaluronate receptors: Key players in growth, differentiation,
migration and tumor progression. Curr Opin Cell Biol. 6:726–733.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Slevin M, Krupinski J, Gaffney J, Matou S,
West D, Delisser H, Savani RC and Kumar S: Hyaluronan-mediated
angiogenesis in vascular disease: Uncovering RHAMM and CD44
receptor signaling pathways. Matrix Biol. 26:58–68. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rodrigo JP, Dominguez F, Alvarez C,
Gonzalez MV, Herrero A and Suárez C: Clinicopathologic significance
of expression of CD44s and CD44v6 isoforms in squamous cell
carcinoma of the supraglottic larynx. Am J Clin Pathol. 118:67–72.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zöller M: CD44: Can a cancer-initiating
cell profit from an abundantly expressed molecule? Nat Rev Cancer.
11:254–267. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Greenfield B, Wang WC, Marquardt H,
Piepkorn M, Wolff EA, Aruffo A and Bennett KL: Characterization of
the heparan sulfate and chondroitin sulfate assembly sites in CD44.
J Biol Chem. 274:2511–2517. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Skelton TP, Zeng C, Nocks A and
Stamenkovic I: Glycosylation provides both stimulatory and
inhibitory effects on cell surface and soluble CD44 binding to
hyaluronan. J Cell Biol. 140:431–446. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hofmann M, Rudy W, Zöller M, Tölg C, Ponta
H, Herrlich P and Günthert U: CD44 splice variants confer
metastatic behavior in rats: Homologous sequences are expressed in
human tumor cell lines. Cancer Res. 51:5292–5297. 1991.PubMed/NCBI
|
|
12
|
Günthert U, Hofmann M, Rudy W, Reber S,
Zöller M, Haussmann I, Matzku S, Wenzel A, Ponta H and Herrlich P:
A new variant of glycoprotein CD44 confers metastatic potential to
rat carcinoma cells. Cell. 65:13–24. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tremmel M, Matzke A, Albrecht I, Laib AM,
Olaku V, Ballmer-Hofer K, Christofori G, Héroult M, Augustin HG,
Ponta H and Orian-Rousseau V: A CD44v6 peptide reveals a role of
CD44 in VEGFR-2 signaling and angiogenesis. Blood. 114:5236–5244.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Orian-Rousseau V, Morrison H, Matzke A,
Kastilan T, Pace G, Herrlich P and Ponta H: Hepatocyte growth
factor-induced Ras activation requires ERM proteins linked to both
CD44v6 and F-actin. Mol Biol Cell. 18:76–83. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Orian-Rousseau V, Chen L, Sleeman JP,
Herrlich P and Ponta H: CD44 is required for two consecutive steps
in HGF/c-Met signaling. Genes Dev. 16:3074–3086. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Rudy W, Hofmann M, Schwartz-Albiez R,
Zöller M, Heider KH, Ponta H and Herrlich P: The two major CD44
proteins expressed on a metastatic rat tumor cell line are derived
from different splice variants: Each one individually suffices to
confer metastatic behavior. Cancer Res. 53:1262–1268.
1993.PubMed/NCBI
|
|
17
|
Jackson DG, Bell JI, Dickinson R, Timans
J, Shields J and Whittle N: Proteoglycan forms of the lymphocyte
homing receptor CD44 are alternatively spliced variants containing
the v3 exon. J Cell Biol. 128:673–685. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bennett KL, Jackson DG, Simon JC, Tanczos
E, Peach R, Modrell B, Stamenkovic I, Plowman G and Aruffo A: CD44
isoforms containing exon V3 are responsible for the presentation of
heparin-binding growth factor. J Cell Biol. 128:687–698. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hu S, Wu X, Zhou B, Xu Z, Qin J, Lu H, Lv
L, Gao Y, Deng L, Yin J and Li G: IMP3 combined with CD44s, a novel
predictor for prognosis of patients with hepatocellular carcinoma.
J Cancer Res Clin Oncol. 140:883–893. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Auvinen P, Tammi R, Kosma VM, Sironen R,
Soini Y, Mannermaa A, Tumelius R, Uljas E and Tammi M: Increased
hyaluronan content and stromal cell CD44 associate with HER2
positivity and poor prognosis in human breast cancer. Int J Cancer.
132:531–539. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fang YJ, Zhang L, Wu XJ, Lu ZH, Li JB, Ou
QJ, Zhang MF, Ding PR, Pan ZZ and Wan DS: Impact of ERβ and CD44
expression on the prognosis of patients with stage II colon cancer.
Tumour Biol. 33:1907–1914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kokko LL, Hurme S, Maula SM, Alanen K,
Grénman R, Kinnunen I and Ventelä S: Significance of site-specific
prognosis of cancer stem cell marker CD44 in head and neck
squamous-cell carcinoma. Oral Oncol. 47:510–516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nguyen VN, Mirejovský T, Melinová L and
Mandys V: CD44 and its v6 spliced variant in lung carcinomas:
Relation to NCAM, CEA, EMA and UP1 and prognostic significance.
Neoplasma. 47:400–408. 2000.PubMed/NCBI
|
|
24
|
Ghatak S, Misra S and Toole BP: Hyaluronan
oligosaccharides inhibit anchorage-independent growth of tumor
cells by suppressing the phosphoinositide 3-kinase/Akt cell
survival pathway. J Biol Chem. 277:38013–38020. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Marangoni E, Lecomte N, Durand L, de
Pinieux G, Decaudin D, Chomienne C, Smadja-Joffe F and Poupon MF:
CD44 targeting reduces tumour growth and prevents post-chemotherapy
relapse of human breast cancers xenografts. Br J Cancer.
100:918–922. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jin L, Hope KJ, Zhai Q, Smadja-Joffe F and
Dick JE: Targeting of CD44 eradicates human acute myeloid leukemic
stem cells. Nat Med. 12:1167–1174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Yamada S, Itai S, Nakamura T, Yanaka M,
Kaneko MK and Kato Y: Detection of high CD44 expression in oral
cancers using the novel monoclonal antibody, C44Mab-5.
Biochem Biophys Rep. 14:64–68. 2018.PubMed/NCBI
|
|
28
|
Kato Y, Mizuno T, Yamada S, Nakamura T,
Itai S, Yanaka M, Sano M and Kaneko MK: Establishment of P38Bf, a
core-fucose-deficient mouse-canine chimeric antibody against dog
podoplanin. Monoclon Antib Immunodiagn Immunother. 37:218–223.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kilkenny C, Browne WJ, Cuthill IC, Emerson
M and Altman DG: Improving bioscience research reporting: The
ARRIVE guidelines for reporting animal research. PLoS Biol.
8:e10004122010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aske KC and Waugh CA: Expanding the 3R
principles: More rigour and transparency in research using animals.
EMBO Rep. 18:1490–1492. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi Y, Tateno H, Dohra H, Moriwaki
K, Miyoshi E, Hirabayashi J and Kawagishi H: A novel core
fucose-specific lectin from the mushroom Pholiota squarrosa.
J Biol Chem. 287:33973–33982. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Takei J, Kaneko MK, Ohishi T, Kawada M,
Harada H and Kato Y: A novel anti-EGFR monoclonal antibody
(EMab-17) exerts antitumor activity against oral squamous cell
carcinomas via antibody-dependent cellular cytotoxicity and
complement-dependent cytotoxicity. Oncol Lett. 19:2809–2816.
2020.PubMed/NCBI
|
|
33
|
Kato Y, Kunita A, Fukayama M, Abe S,
Nishioka Y, Uchida H, Tahara H, Yamada S, Yanaka M, Nakamura T, et
al: Antiglycopeptide mouse monoclonal antibody LpMab-21 exerts
antitumor activity against human podoplanin through
antibody-dependent cellular cytotoxicity and complement-dependent
cytotoxicity. Monoclon Antib Immunodiagn Immunother. 36:20–24.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wimmerova M, Mitchell E, Sanchez JF,
Gautier C and Imberty A: Crystal structure of fungal lectin:
Six-bladed beta-propeller fold and novel fucose recognition mode
for Aleuria aurantia lectin. J Biol Chem. 278:27059–27067.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sumner JB, Howell SF and Zeissig A:
Concanavalin a and Hemagglutination. Science. 82:65–66. 1935.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Rivera C: Essentials of oral cancer. Int J
Clin Exp Pathol. 8:11884–11894. 2015.PubMed/NCBI
|
|
37
|
Güneri P and Epstein JB: Late stage
diagnosis of oral cancer: Components and possible solutions. Oral
Oncol. 50:1131–1136. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vokes EE: Induction chemotherapy for head
and neck cancer: Recent data. Oncologist. 15 (Suppl 3):S3–S7. 2010.
View Article : Google Scholar
|
|
39
|
Marcazzan S, Varoni EM, Blanco E, Lodi G
and Ferrari M: Nanomedicine, an emerging therapeutic strategy for
oral cancer therapy. Oral Oncol. 76:1–7. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Furness S, Glenny AM, Worthington HV,
Pavitt S, Oliver R, Clarkson JE, Macluskey M, Chan KK and Conway
DI: Interventions for the treatment of oral cavity and
oropharyngeal cancer: Chemotherapy. Cochrane Database Syst Rev.
2011:CD0063862011.
|
|
41
|
Bonner JA, Harari PM, Giralt J, Azarnia N,
Shin DM, Cohen RB, Jones CU, Sur R, Raben D, Jassem J, et al:
Radiotherapy plus cetuximab for squamous-cell carcinoma of the head
and neck. N Engl J Med. 354:567–578. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Vermorken JB, Mesia R, Rivera F, Remenar
E, Kawecki A, Rottey S, Erfan J, Zabolotnyy D, Kienzer HR, Cupissol
D, et al: Platinum-based chemotherapy plus cetuximab in head and
neck cancer. N Engl J Med. 359:1116–1127. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Naruse T, Yanamoto S, Matsushita Y,
Sakamoto Y, Morishita K, Ohba S, Shiraishi T, Yamada SI, Asahina I
and Umeda M: Cetuximab for the treatment of locally advanced and
recurrent/metastatic oral cancer: An investigation of distant
metastasis. Mol Clin Oncol. 5:246–252. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Amit M, Yen TC, Liao CT, Chaturvedi P,
Agarwal JP, Kowalski LP, Ebrahimi A, Clark JR, Kreppel M, Zöller J,
et al: Improvement in survival of patients with oral cavity
squamous cell carcinoma: An international collaborative study.
Cancer. 119:4242–4248. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Takei J, Kaneko MK, Ohishi T, Kawada M,
Harada H and Kato Y: H2Mab-19, an anti-human epidermal
growth factor receptor 2 monoclonal antibody exerts antitumor
activity in mouse oral cancer xenografts. Exp Ther Med. 20:846–853.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hanken H, Gaudin R, Gröbe A, Fraederich M,
Eichhorn W, Smeets R, Simon R, Sauter G, Grupp K, Izbicki JR, et
al: Her2 expression and gene amplification is rarely detectable in
patients with oral squamous cell carcinomas. J Oral Pathol Med.
43:304–308. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yan M, Schwaederle M, Arguello D, Millis
SZ, Gatalica Z and Kurzrock R: HER2 expression status in diverse
cancers: Review of results from 37,992 patients. Cancer Metastasis
Rev. 34:157–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Kondo N, Ishiguro Y, Kimura M, Sano D,
Fujita K, Sakakibara A, Taguchi T, Toth G, Matsuda H and Tsukuda M:
Antitumor effect of gefitinib on head and neck squamous cell
carcinoma enhanced by trastuzumab. Oncol Rep. 20:373–378.
2008.PubMed/NCBI
|
|
49
|
Kondo N, Tsukuda M, Sakakibara A,
Takahashi H, Hyakusoku H, Komatsu M, Niho T, Nakazaki K and Toth G:
Combined molecular targeted drug therapy for EGFR and HER-2 in head
and neck squamous cell carcinoma cell lines. Int J Oncol.
40:1805–1812. 2012.PubMed/NCBI
|
|
50
|
Itai S, Ohishi T, Kaneko MK, Yamada S, Abe
S, Nakamura T, Yanaka M, Chang YW, Ohba SI, Nishioka Y, et al:
Anti-podocalyxin antibody exerts antitumor effects via
antibody-dependent cellular cytotoxicity in mouse xenograft models
of oral squamous cell carcinoma. Oncotarget. 9:22480–22497. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kato Y and Kaneko MK: A cancer-specific
monoclonal antibody recognizes the aberrantly glycosylated
podoplanin. Sci Rep. 4:59242014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yamada S, Ogasawara S, Kaneko MK and Kato
Y: LpMab-23: A cancer-specific monoclonal antibody against human
podoplanin. Monoclon Antib Immunodiagn Immunother. 36:72–76. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Kaneko MK, Yamada S, Nakamura T, Abe S,
Nishioka Y, Kunita A, Fukayama M, Fujii Y, Ogasawara S and Kato Y:
Antitumor activity of chLpMab-2, a human-mouse chimeric
cancer-specific antihuman podoplanin antibody, via
antibody-dependent cellular cytotoxicity. Cancer Med. 6:768–777.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Kaneko MK, Nakamura T, Kunita A, Fukayama
M, Abe S, Nishioka Y, Yamada S, Yanaka M, Saidoh N, Yoshida K, et
al: ChLpMab-23: Cancer-specific human-mouse chimeric
anti-podoplanin antibody exhibits antitumor activity via
antibody-dependent cellular cytotoxicity. Monoclon Antib
Immunodiagn Immunother. 36:104–112. 2017. View Article : Google Scholar : PubMed/NCBI
|