Introduction
In the past few decades, hepatocellular carcinoma
(HCC) has become one of the most common human malignancies,
accounting for >70% of all liver cancer cases worldwide
(1), and was the third leading
cause of cancer-related death worldwide in 2017 (2). Although considerable advances have
been made in HCC diagnosis and treatment, the prognosis of patients
is still unsatisfactory (3). The
5-year survival rate of HCC remains at a low level, which may be
due to the high recurrence rate after surgical resection (4). Studies have indicated that the
primary cause of HCC is infection with hepatitis B and C virus.
However, diabetes and excessive drinking can also contribute to HCC
(5). Therefore, a more thorough
exploration of HCC pathogenesis is imperative to developing new
diagnostic methods and identifying therapeutic targets.
Long noncoding RNAs (lncRNAs), >200 nucleotides
in length, have no protein-coding function (6). As a type of noncoding RNA, the
functions of lncRNAs have been widely investigated during the past
decade. Increasing numbers of studies have reported that
dysregulation of lncRNA expression is closely associated with the
occurrence and development of cancers (7–11).
Research has also indicated the critical roles of lncRNAs in
hepatocarcinogenesis, with regard to cellular proliferation,
apoptosis and metastasis (12).
For example, expression of LINRIS was upregulated in CRC tissues
and is associated with poor overall survival, and LINRIS stabilizes
insulin-like growth factor 2 mRNA-binding protein 2 to promote
colorectal cancer progression (13). Furthermore, lncRNA
anti-differentiation ncRNA was found to regulate the invasion and
metastasis abilities of breast cancer cells by affecting the
stability of histone-lysine N-methyltransferase EZH2 (14). Expression levels of lncRNA
colorectal neoplasia differentially expressed (CRNDE) have been
found to increase in various types of cancer. For example, higher
expression of lncRNA CRNDE promoted cellular proliferation and
chemoresistance by targeting microRNA
(miR)-181a-5p-Wnt/beta-catenin signaling in colorectal cancer
(15). CRNDE also bound and
reduced the protein stability of serine/arginine-rich splicing
factor 6 to regulate the chemoresistance of gastric cancer
(16). In the present study, the
molecular mechanism of CRNDE in HCC was further explored.
MicroRNAs (miRNAs/miRs) are also involved in the
progression of HCC. For example, miR-29c-3p expression was
downregulated in colorectal cancer tissues, and inhibited cancer
progression by negatively regulating circ-001971 and VEGFA
(17). Low expression levels of
miR-29c-3p were associated with shorter overall survival times of
patients with HCC, while overexpression of miR-29c-3p markedly
inhibited cellular proliferation and tumor growth (18). Nuclear autoantigenic sperm protein
(NASP) is reported to be upregulated in gastric cancer, ovarian
cancer and hepatocellular carcinoma (19–21).
However, the function of the miR-29c-3p-NASP axis in HCC remains
unclear.
In the present study, the expression level of CRNDE
in HCC was assessed using normal non-tumor tissue data from the
Gene Expression Profiling Interactive Analysis (GEPIA) database and
reverse transcription-quantitative (RT-q) PCR. miR-29c-3p was
predicted as a potential target of CRNDE, and its CRNDE regulatory
role was investigated. In addition, the effects of the
CRNDE/miR-29c-3p/NASP axis on cellular proliferation, migration and
invasiveness, and the underlying mechanisms of
CRNDE/miR-29c-3p/NASP in HCC, were determined.
Materials and methods
Clinical specimens
A total of 40 pairs of HCC and adjacent-normal
tissues were collected from The Second Hospital of Tianjin Medical
University (Tianjin, China) between January 2017 and December 2019.
HCC diagnosis was primarily confirmed by tumor marker detection and
liver imaging examinations. Patients were aged <50 years old
(n=17) and ≥50 years old (n=23), 22 of which were male, and 18
female. Patient inclusion criteria included: i) The patient was
diagnosed with primary liver cancer; and ii) the patient understood
the purpose of the study and provided written informed consent.
Exclusion criteria included: i) Patients diagnosed with other
tumors or liver insufficiency; ii) patients experiencing mental
illness; and iii) patients with other major diseases other than the
tumor. The distance between the tumor tissue and healthy tissues
was ≥3 cm, and tissues were immediately frozen in the liquid
nitrogen following resection. The patient clinicopathological
features are presented in Table I.
Surgical indications include the tumor lesion not exceeding half of
the liver, no distant metastases, and that patients exhibited
normal liver function. None of the enrolled patients had received
any pre-surgical treatment. The study was approved by the Ethical
Committee of the Second Hospital of Tianjin Medical University
(approval no. KY2020K093), and written informed consent was
obtained from each patient.
 | Table I.Association between CRNDE expression
and the clinicopathological characteristics of patients with HCC
(n=40). |
Table I.
Association between CRNDE expression
and the clinicopathological characteristics of patients with HCC
(n=40).
|
|
| CRNDE
expression |
|
|---|
|
|
|
|
|
|---|
|
Characteristics | n=40 | Low (n=20) | High (n=20) | P-value |
|---|
| Age, years |
|
|
|
|
|
<50 | 17 | 8 | 9 |
|
|
≥50 | 23 | 12 | 11 | 0.749 |
| Sex |
|
|
|
|
|
Male | 22 | 14 | 8 |
|
|
Female | 18 | 6 | 12 | 0.057 |
| Tumor size, cm |
|
|
|
|
| ≤5 | 14 | 10 | 4 |
|
|
>5 | 26 | 10 | 16 | 0.047 |
| TNM stage |
|
|
|
|
|
I+II | 18 | 13 | 5 |
|
|
III+IV | 22 | 7 | 15 | 0.011 |
Cell culture
Normal liver THLE-2 cells and liver cancer cells
(SNU-398 and Huh7) were purchased from the American Type Culture
Collection. All cell lines were authenticated by STR profiling. The
cells were cultured in RPMI 1640 medium supplemented with 10% fetal
bovine serum (FBS; both Gibco; Thermo Fisher Scientific, Inc.), and
maintained at 37°C in an incubator containing 5%
CO2.
Transfection
Lentivector-mediated short hairpin (sh) RNA CRNDE
(shCRNDE-1, 5′-GGTTAAGCTGTATTTGATTGC−3′; shCRNDE-2,
5′-GATGTGTTTCAATCTAGATGC−3′; and shCRNDE-3,
5′-GCTCGAGTGGTTTAAATATGT−3′) and the negative control plasmids
(shNC; 5′-CAACAAGATGAAGAGCACCAA−3′) were synthesized by Guangzhou
RiboBio Co., Ltd. The CRNDE overexpression plasmid and the empty
negative control plasmid (pcDNA3.1) were purchased from Shanghai
GenePharma Co., Ltd. Briefly, CRNDE-encoding cDNA was amplified and
cloned into the pcDNA3.1 plasmid; sequence sources for CRNDE and
NASP are Accession NR_034105.4 and NM_001195193, respectively. The
negative control (NC) mimics, miR-29c-3p mimics, miR-29c-3p
inhibitor and NC inhibitor were purchased from Guangzhou RiboBio
Co., Ltd., according to the mature miR-29c-3p sequence
(5′-UAGCACCAUUUGAAAUCGGUUA-3′). At 70% confluence, HCC cells were
transfected using Lipofectamine® 3000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
instructions. Briefly, 1 µg nucleic acid was used to transfect
cells; the nucleic acid and Lipofectamine were separately diluted
using Opti-MEM® (Invitrogen; Thermo Fisher Scientific,
Inc.), and then mixed and incubated at room temperature for 5 min.
Then, 48 h after transfection at 37°C, the cells were harvested for
subsequent experimentation.
RNA extraction and RT-qPCR
Total RNA was extracted from tissues and cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.) per the manufacturer's instructions, and quantified with a
NanoDrop 2000 instrument (Thermo Fisher Scientific, Inc.). Then, 1
µg RNA was reverse transcribed to cDNAs using the Reverse
Transcription System kit (Invitrogen; Thermo Fisher Scientific,
Inc.) in a 20 µl reaction. qPCR was then performed using the
SYBR® Premix EX Taq™ kit (Takara Bio, Inc.) with a
Real-Time PCR System (Thermo Fisher Scientific, Inc.): Total
reaction (20 µl) consisted of 10 µl master mix (TaqMan), 1 µl
forward and reverse primers, 2 µl cDNA and 7 µl DEPC water.
Reaction conditions: i) 95°C for 5 min; and ii) 40 cycles of 95°C
for 10 sec, 60°C for 30 sec, and 72°C for 30 sec. Gene expression
was quantified using the 2−ΔΔCq method (22). The expression of CRNDE and NASP was
normalized to that of GAPDH (the internal control), and miR-29c-3p
expression was normalized to U6. qPCR was performed with the
following primers: CRNDE forward, 5′-AAATTCATCCCAAGGCTGGT-3′ and
reverse, 5′-AAACCACTCGAGCACTTTGA-3′; NASP forward,
5′-AGATTGGGAACCTAGAGCTTGC-3′ and reverse,
5′-ACTTCTCCGAGTTTAAGATGTGC-3′; miR-29c-3p forward,
5′-ACACTCCAGCTGGTAGCACCATTGAAAT-3′ and reverse,
5′-TGGTGTCGTGGAGTCG-3′; miR-29a-3p forward,
5′-GGGTAGCACCATCTGAAAT-3′ and reverse, 5′-CAGTGCGTGTCGTGGAGT−3′;
miR-146a-5p forward, 5′-GGCGATGAGAACTGAATTCCA−3′ and reverse,
5′-GCAGGGACCGTGGT-3′; miR-455-3p forward,
5′-GCCGGATCCAGTCGTGTTGCTTTATCTACAGAGC−3′ and reverse,
5′-GCCGAATTCTCAGAGCGGTCAAGCAGTGA-3′; miR-28-5p forward,
5′-TGGTGTCGTGGGTCGA-3′ and reverse, 5′-CTCGCTTCGGCAGCACA-3′;
miR-338-3p forward, 5′-GGGTCCAGCATCAGTGATT-3′ and reverse,
5′-GTGCAGGGTCCGAGGT-3′; GAPDH forward, 5′-AATGGACAACTGGTCGTGGAC−3′
and reverse, 5′-CCCTCCAGGGGATCTGTTTG-3′; and U6 forward,
5′-AGTAAGCCCTTGCTGTCAGTG−3′ and reverse,
5′-CCTGGGTCTGATAATGCTGGG.
Cell viability assay
The Cell Counting kit 8 (CCK-8; Dojindo Molecular
Technologies, Inc.) was used to evaluate cellular proliferation
after transfection. A total of 1×103 transfected cells
per well were seeded into 96-well plates. Then, 10 µl CCK-8
solution was added to each well at the indicated transfection time
points (24, 48 and 72 h) and incubated for a further 2 h according
to the manufacturer's instructions. The absorbance was detected at
450 nm using a micro-plate reader. Cell viability was calculated
according to the manufacturer's instructions.
Wound-healing assay
Transfected cells were seeded into a 6-well plate.
When reaching confluence, a wound was created in the monolayer
using a 10-µl pipette tip and the cells were cultured in medium
without serum. Images of wound width were captured at 0 and 24 h
using an inverted microscope (Thermo Fisher Scientific, Inc.). The
width of each wound was measured and the fold change was
calculated.
Dual-luciferase reporter assay
To confirm the interaction between miR-29c-3p and
CRNDE, as well as miR-29c-3p and NASP, wild-type (WT) and
mutant-type (MUT) sequences of CRNDE and NASP (containing the
putative binding site of miR-29c-3p) were cloned into the pGL3
reporter vector (Promega Corporation). 293T cells were
co-transfected with the WT or MUT reporter plasmids, and miR-29c-3p
mimics or NC mimics using Lipofectamine® 3000
(Invitrogen; Thermo Fisher Scientific, Inc.). After 48 h, the cells
were collected and lysed. The fluorescence intensity was measured
using the Dual-Luciferase Reporter Assay System (Promega
Corporation) and normalized by comparison with Renilla
luciferase activity, according to the manufacturer's
instructions.
Transwell assay
To assess cellular migration and invasion ability,
1×105 cells were seeded into the upper chamber of
Transwell inserts with serum-free medium to assess migration, and
medium containing 10% FBS was added to the lower chambers. For the
invasion assay, the upper chamber was pre-coated with Matrigel (BD
Biosciences, 1:8 dilution) at 37°C for 7 h. Cells were maintained
in an incubator at 37°C prior to fixing. After 24 h, the cells were
fixed with 4% paraformaldehyde for 10 min and stained with 0.4%
crystal violet for 10 min at room temperature. Then, the stained
cells were washed with PBS and observed using an inverted
microscope (Thermo Fisher Scientific, Inc.). The number of cells
was calculated in five randomly selected visual fields using ImageJ
1.51 software (National Institutes of Health).
Western blotting
Total protein samples were extracted form cells
using RIPA lysis buffer (Beyotime Institute of Biotechnology) with
PMSF. Protein concentration was quantified using a BCA kit
(Beyotime Institute of Biotechnology). Equal amounts of protein (30
µg) were separated by 10% SDS-PAGE and transferred to PVDF
membranes. The Membranes were then blocked with 5% non-fat milk for
1 h at room temperature, and incubated with the primary antibodies
overnight at 4°C. Then, the membranes were incubated with the
corresponding HRP-conjugated secondary antibodies for 1 h at room
temperature. The protein bands were visualized using an ECL kit
(Beyotime Institute of Biotechnology). The gray values of the
protein bands were quantified using ImageJ 1.51 software (National
Institutes of Health) and relative expression levels were
calculated. The following antibodies were employed: Anti-NASP
(1:1,000; cat. no. ab181169; Abcam), anti-N cadherin (1:3,000; cat.
no. ab76011; Abcam), anti-E cadherin (1:1,000; cat. no. ab231303;
Abcam), anti-vimentin antibody (1:1,000; cat. no. ab16700; Abcam),
anti-GAPDH (1:5,000; cat. no. ab8245; Abcam), goat anti-rabbit
(1:3,000; cat. no. 7074; CST Biological Reagents Co., Ltd.) and
horse anti-mouse (1:3,000; cat. no. 7076; CST Biological Reagents
Co., Ltd.).
Immunohistochemistry
Tissues were fixed with 4% paraformaldehyde at room
temperature for ≥48 h. Paraffin sections of 5 µm were cut from the
tumor tissues of patients with HCC, and were dewaxed with xylene
for 3 h at room temperature, and rehydration in a descending
alcohol series (absolute ethanol, 10 min; methanol with 3% hydrogen
peroxide, 10 min; 90% alcohol, 10 min; 70% alcohol, 5 min). The
samples were then washed with ddH2O. Antigen retrieval
was conducted at 95°C for 15 min, then the samples were cooled to
room temperature andwashed with ddH2O once again. The
sections were blocked with 10% goat serum at room temperature for 1
h, and then incubated with anti-NASP (1:100; cat. no. sc-514669;
Santa Cruz Biotechnology, Inc.) at 4°C overnight. After incubation
with an anti-mouse HRP-conjugated secondary antibody (1:1,000; cat.
no. 7076; CST Biological Reagents Co., Ltd.), the sections were
stained with DAB at room temperature for 10 min, and hematoxylin at
room temperature for 5 min, and NASP expression was observed using
a light microscope.
Bioinformatics analysis
CRNDE expression data were obtained from The Cancer
Genome Atlas (TCGA) in GEPIA (http://gepia.cancer-pku.cn/) and the Lnc2Cancer 3.0
(http://bio-bigdata.hrbmu.edu.cn/lnc2cancer/index.html)
database. Differentially expressed miRNAs were screened in dbDEMC
2.0 (https://www.picb.ac.cn/dbDEMC/index.html). Target
genes were analyzed using UALCAN (http://ualcan.path.uab.edu/). Potential target miRNAs
of CRNDE were predicted by starBase (http://starbase.sysu.edu.cn). Target genes of miRNA
were predicted by starBase, miRWalk (http://mirwalk.umm.uni-heidelberg.de/), TargetScan
(http://www.targetscan.org/vert_71/),
miRDB (http://mirdb.org/) and ONCOMIR (http://www.oncomir.org/oncomir/index.html).
Statistical analysis
Results are presented as the mean ± SD, and were
analyzed using SPSS 19.0 (IBM Corp) and GraphPad Prism 8.0 Software
(GraphPad Software, Inc.). One-way ANOVA and Tukey's test were used
for multiple comparison analyses. All data from normal vs. tumor
tissues were analyzed using paired t-tests. All other comparisons
between two groups were analyzed by unpaired t-tests. Associations
between CRNDE expression and clinicopathological characteristics
were assessed using the χ2 test. All experiments were
carried out in triplicate, and P<0.05 was considered to indicate
a statistically significant difference.
Results
CRNDE expression is elevated in HCC
tissues
First, CRNDE expression was detected in HCC tissues
using online database analysis and RT-qPCR. Results from TCGA
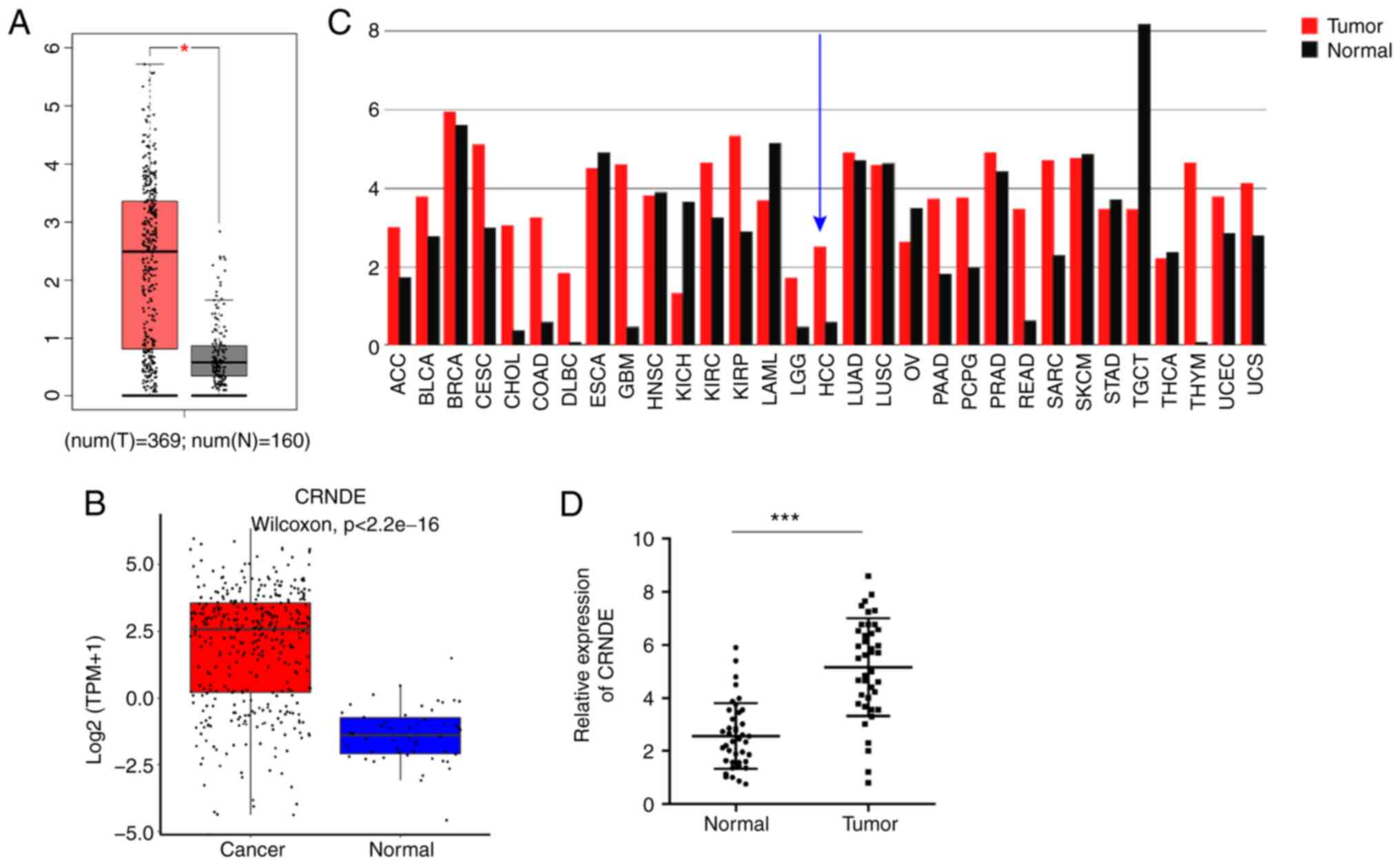
revealed that the expression of CRNDE was significantly increased
in HCC tissues compared with adjacent-normal tissues (Fig. 1A), and a similar result was
observed in the Lnc2Cancer 3.0 database (Fig. 1B). CRNDE expression was also
increased in a number of other cancer types, aside from HCC
(Fig. 1C). Furthermore, RT-qPCR
was performed to assess the expression of CRNDE in HCC and
non-cancer tissues. The results indicated that CRNDE expression was
significantly elevated in HCC tissues (Fig. 1D). Thus, these results suggested
that CRNDE is positively associated with HCC progression.
CRNDE-knockdown suppresses HCC cell
proliferation, migration and invasiveness
Next, the expression levels of CRNDE were assessed
in normal liver and HCC cell lines. Concertedly, CRNDE expression
was upregulated in cancer cells, but not in normal liver cells
(Fig. 2A). To determine its
function in HCC cells, CRNDE expression was silenced in SNU-398 and
Huh7 cells via shRNA-CRNDE transfection, and confirmed by RT-qPCR.
Due to superior knockdown efficiency, shCRNDE-2 and shCRNDE-3 were
selected for further experimentation (Fig. 2B). Subsequently, a CCK-8 assay was
used to evaluate the proliferative capacity of cells transfected
with shCRNDE, which was found to be significantly inhibited by
CRNDE-knockdown (Fig. 2C).
Moreover, Transwell and wound-healing assays revealed that the
migratory and invasive capacities were suppressed when CRNDE was
knocked down (Fig. 2D-F). These
results demonstrated that CRNDE-knockdown inhibited the
proliferation and metastatic potential of HCC cells in
vitro.
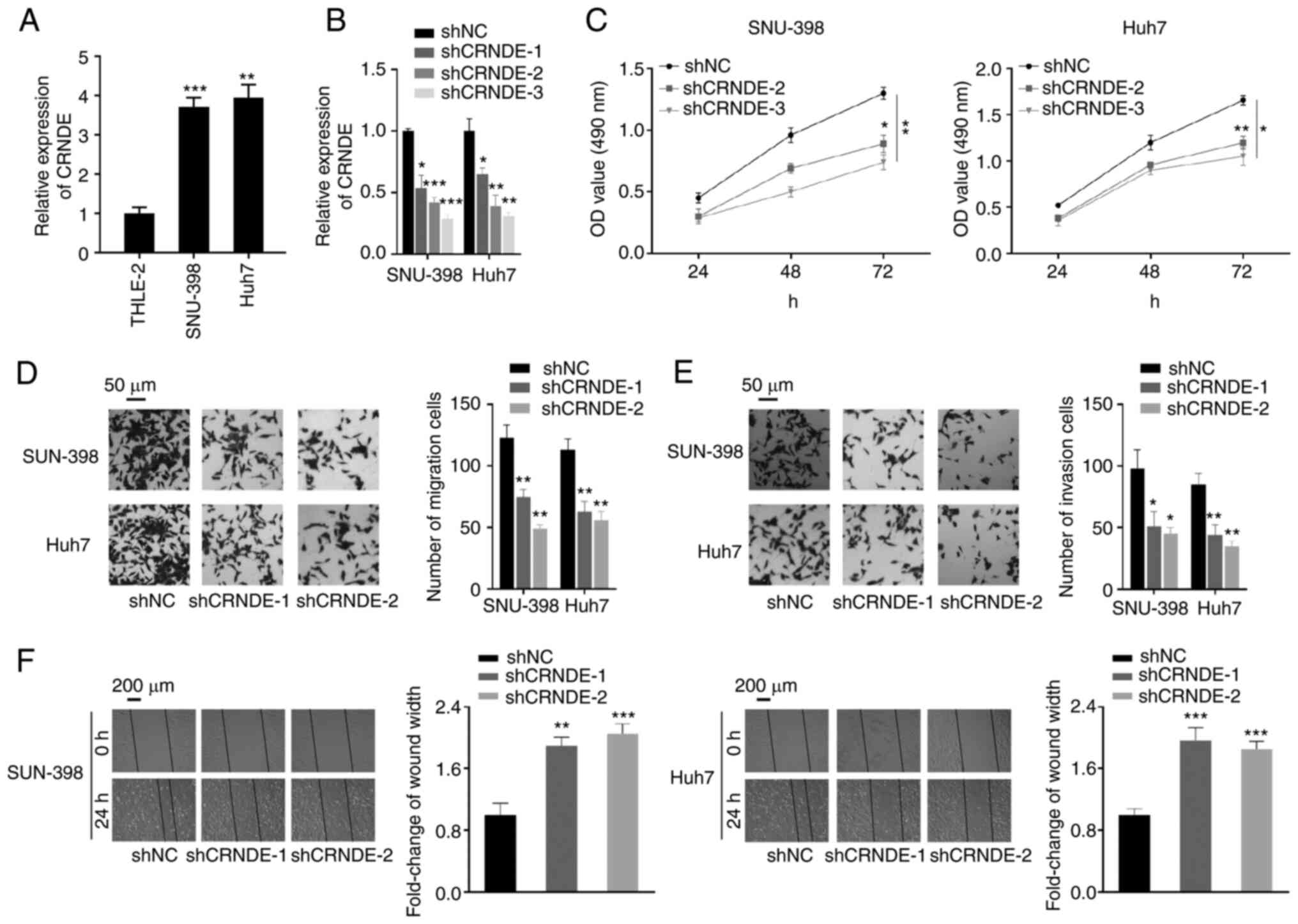 | Figure 2.CRNDE-knockdown inhibits HCC cell
proliferation, migration and invasiveness. (A) Relative expression
levels of CRNDE in normal liver cells and liver cancer cells. (B)
SNU-398 and Huh7 cells were transfected with shNC, shCRNDE-1,
shCRNDE-2 or shCRNDE-3, and CRNDE expression was determined by
reverse transcription-quantitative PCR. (C) HCC cell proliferation
was determined by Cell Counting kit 8 analysis after shCRNDE
transfection. (D and E) CRNDE-knockdown inhibited the migration and
invasiveness of HCC cells. (F) Migration ability was detected by
wound-healing assays in SNU-398 and Huh7 cells transfected with
shNC, shCRNDE-1 or shCRNDE-2. *P<0.05, **P<0.01 and
***P<0.001 vs. THLE-2 or shNC. CRNDE, lncRNA colorectal
neoplasia differentially expressed; HCC, hepatocellular carcinoma;
sh, short hairpin (RNA); NC, negative control. |
CRNDE targets and negatively regulates
miR-29c-3p expression
Increasing reports have indicated that lncRNAs can
act as competing endogenous RNAs (ceRNAs) to miRNAs (23,24).
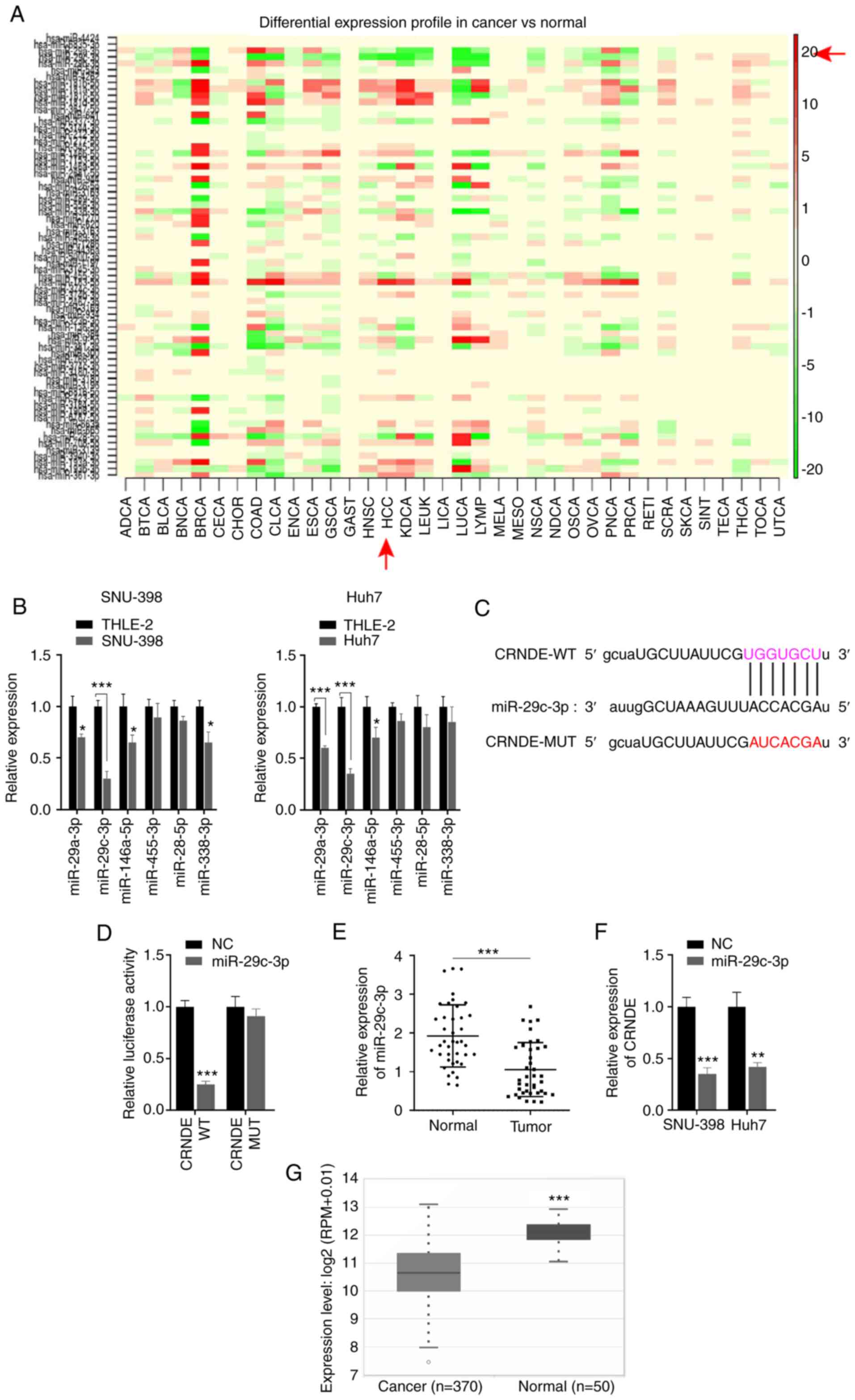
In the present study, 60 potential targets of CRNDE were predicted
using the starBase database. Furthermore, differentially expressed
miRNAs were screened in association with multiple cancers via
bioinformatics analysis (Fig. 3A).
In HCC, six miRNAs with low expression were selected, and their
expression assessed in SNU-398 and Huh7 cells. The RT-qPCR results
showed that among the six miRNAs, miR-29c-3p expression was the
lowest expressed in HCC cells (Fig.
3B). Therefore, miR-29c-3p was selected as a potential target
miRNA of CRNDE. The WT and MUT binding sequence between CRNDE and
miR-29c-3p is presented in Fig.
3C. To verify the interaction between CRNDE and miR-29c-3p, a
dual-luciferase reporter assay was performed, revealing that the
overexpression of miR-29c-3p markedly reduced the luciferase
activity of cells transfected with CRNDE-WT, but not CRNDE-MUT
(Fig. 3D). Additionally,
miR-29c-3p expression was markedly decreased in HCC tissues
(Fig. 3E). To determine the
effects of miR-29c-3p on CRNDE expression, miR-29c-3p expression
was assessed in HCC cells transfected with miR-29c-3p mimics or
inhibitor. The results showed that miR-29c-3p mimics significantly
increased miR-29c-3p expression, while the miR-29c-3p inhibitor
downregulated expression (Fig. S1A
and B). Furthermore, CRNDE expression was inhibited by
miR-29c-3p overexpression in HCC cells (Fig. 3F). Reduced expression of miR-29c-3p
was also observed in patients with HCC via bioinformatics analysis
(Fig. 3G). Collectively, these
results confirm that miR-29c-3p is a direct target of CRNDE, and is
decreased in HCC.
NASP is a downstream target gene of
miR-29c-3p
The downstream target genes of miR-29c-3p were
predicted using five databases, and a total of 17 genes co-existed
in five databases (Fig. 4A). The
heat map of gene expression indicated that the expression levels of
NASP, TMOD3, KCTD2 and CPR37 were higher in HCC compared with the
other genes investigated (Fig.
4B). In the present study, a binding site between miR-29c-3p
and NASP was predicted using TargetScan (Fig. 4C). Furthermore, dual-luciferase
reporter assays confirmed that the luciferase activity of
NASP-WT-transfected cells was significantly decreased following the
overexpression of miR-29c-3p, while miR-29c-3p mimics did not
affect the luciferase activity of the NASP-MUT group (Fig. 4D). RT-qPCR analysis indicated that
NASP mRNA was significantly increased in HCC compared with normal
tissues (Fig. 4E). Similarly,
higher NASP expression levels were also observed in HCC cancer
tissues via the GEPIA and UALCAN databases, respectively (Fig. 4F and G). Moreover, NASP expression
was positively associated with HCC tumor grade and individual
cancer stage (Fig. 4H and I), and
HCC patients with higher NASP expression experienced lower
disease-free survival rates (Fig.
4J).
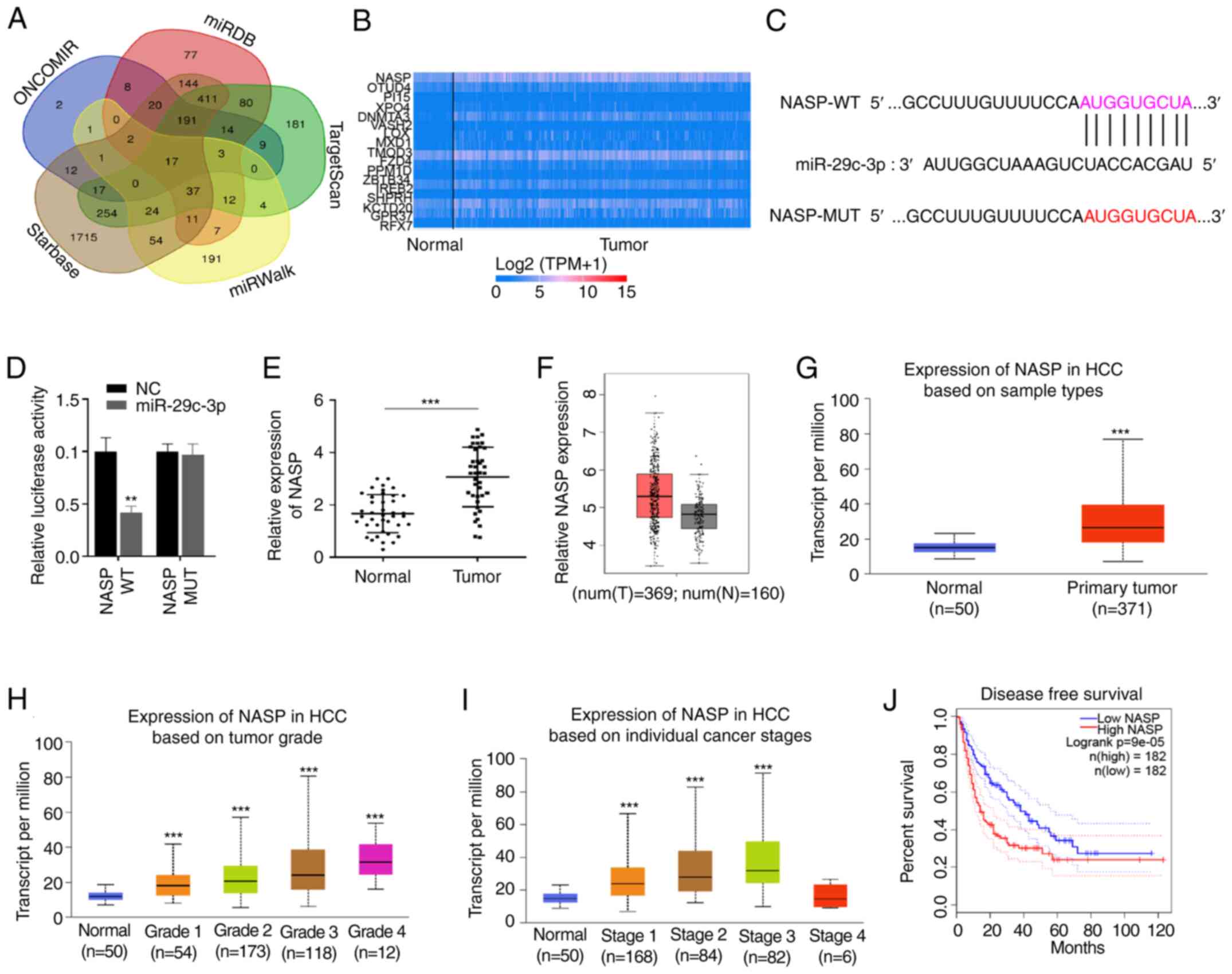 | Figure 4.miR-29c-3p targets NASP and NASP
expression is increased in patients with HCC. (A) Targets of
miR-29c-3p were predicted using the miRWalk, starBase, Oncomir,
TargetScan and miRDB databases. (B) Expression heatmap of 16 target
genes of miR-29c-3p was analyzed in HCC using the UALCAN database.
(C) Binding sites between miR-29c-3p and NASP. (D) Relationship
between miR-29c-3p and NASP was confirmed by dual luciferase
reporter assay. (E) NASP expression was detected by reverse
transcription-quantitative PCR in HCC and normal tissues. NASP
expression level analysis in (F) GEPIA and (G) UALCAN databases,
respectively. (H) HCC tumor grade was positively associated with
NASP expression. (I) Expression of NASP at different HCC patient
stages. (J) NASP expression was related to the disease-free
survival of patients with HCC in GEPIA database. **P<0.01 and
***P<0.001 vs. WT or Normal. miR, microRNA; NASP, nuclear
autoantigenic sperm protein; HCC, hepatocellular carcinoma; WT,
wild-type; MUT, mutant-type; GEPIA, Gene Expression Profiling
Interactive Analysis. |
CRNDE and miR-29c-3p regulate NASP
expression in HCC cells
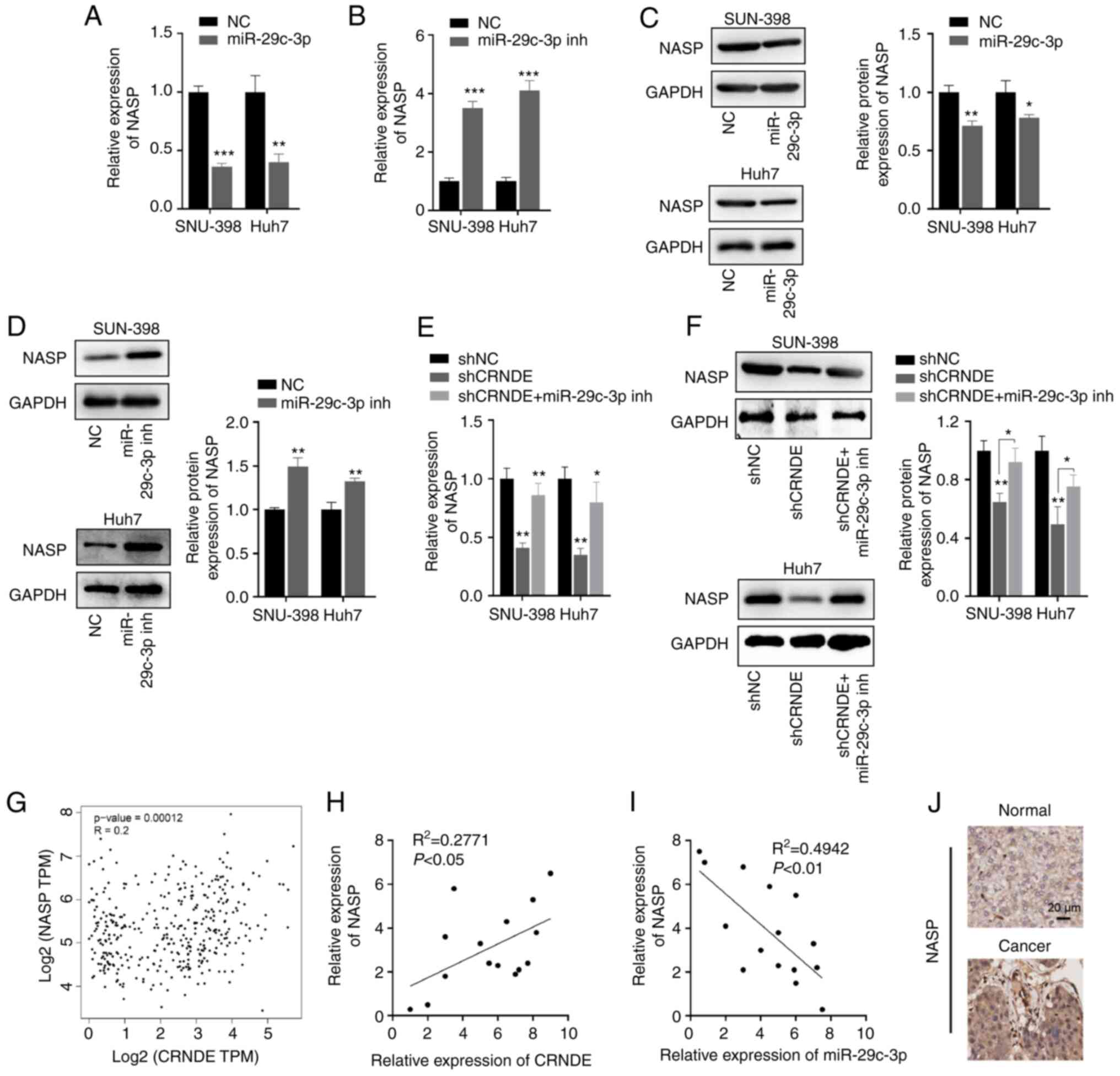
To determine the effects of CRNDE and miR-29c-3p on
NASP expression, HCC cells were transfected with miR-29c-3p mimics,
miR-29c-3p inhibitor, shCRNDE, CRNDE or their respective negative
controls. The results showed that NASP expression was inhibited by
miR-29c-3p mimics (Fig. 5A), but
increased by the miR-29c-3p inhibitor (Fig. 5B). Consistent with these results,
the protein expression levels of NASP were regulated by miR-29c-3p
mimics and inhibitor (Fig. 5C and
D). Furthermore, the effect of CRNDE on NASP expression was
assessed in HCC cells transfected with shCRNDE or CRNDE. The
results indicated that NASP expression at the mRNA and protein
levels were notably suppressed by shCRNDE, which was rescued by the
miR-29c-3p inhibitor (Fig. 5E and
F). Additionally, correlation analyses revealed that NASP
expression was positively associated with that of CRNDE, (Fig. 5G and H), but negatively associated
with miR-29c-3p expression (Fig.
5I). Immunohistochemistry also revealed that the expression of
NASP was also increased in the tumor tissues of patients with HCC,
compared with normal tissues (Fig.
5J). Thus, these results indicated that CRNDE regulated NASP
expression by inhibiting miR-29c-3p in HCC cells.
The CRNDE-miR-29c-3p-NASP axis
regulates HCC cell proliferation, migration and invasiveness
To further determine the roles of the
CRNDE-miR-29c-3p-NASP axis in HCC, the overexpression efficiency of
CRNDE and NASP was detected in HCC cells (Fig. S1C and D). Then, the indicated
transfections were performed, and rescue assays were conducted to
study the effects of miR-29c-3p on CRNDE and NASP. Firstly, CCK-8
assays indicated that cellular proliferation ability was promoted
in SNU-398 and Huh7 cells following CRNDE and NASP overexpression,
while the opposite effect was observed following miR-29c-3p mimics
transfection. However, rescue assays indicated that the promotion
of cell proliferation caused by CRNDE and NASP was significantly
weakened by miR-29c-3p mimics (Fig.
6A). To evaluate whether the CRNDE-miR-29c-3p-NASP axis affects
epithelial mesenchymal transition (EMT), western blot analysis was
carried out. The results indicated that the expression levels of
N-cadherin and vimentin were increased in cells transfected with
the CRNDE or NASP overexpression plasmids, while E-cadherin was
downregulated. However, the effects of CRNDE or NASP in EMT was
reversed by miR-29c-3p mimics. These results demonstrated that the
overexpression of CRNDE and NASP was conducive to EMT, but was
negatively regulated by miR-29c-3p mimics (Fig. 6B). Moreover, migration and
wound-healing assays indicated that CRNDE and NASP promoted
cellular migration capacity, which was reversed by miR-29c-3p
mimics (Fig. 6C and E). Consistent
with the migration results, the overexpression of miR-29c-3p also
successfully rescued the effects of CRNDE and NASP on cellular
invasiveness (Fig. 6D).
Collectively, these data demonstrated that CRNDE promoted HCC cell
proliferation, migration and invasiveness by regulating the
miR-29c-3p/NASP axis.
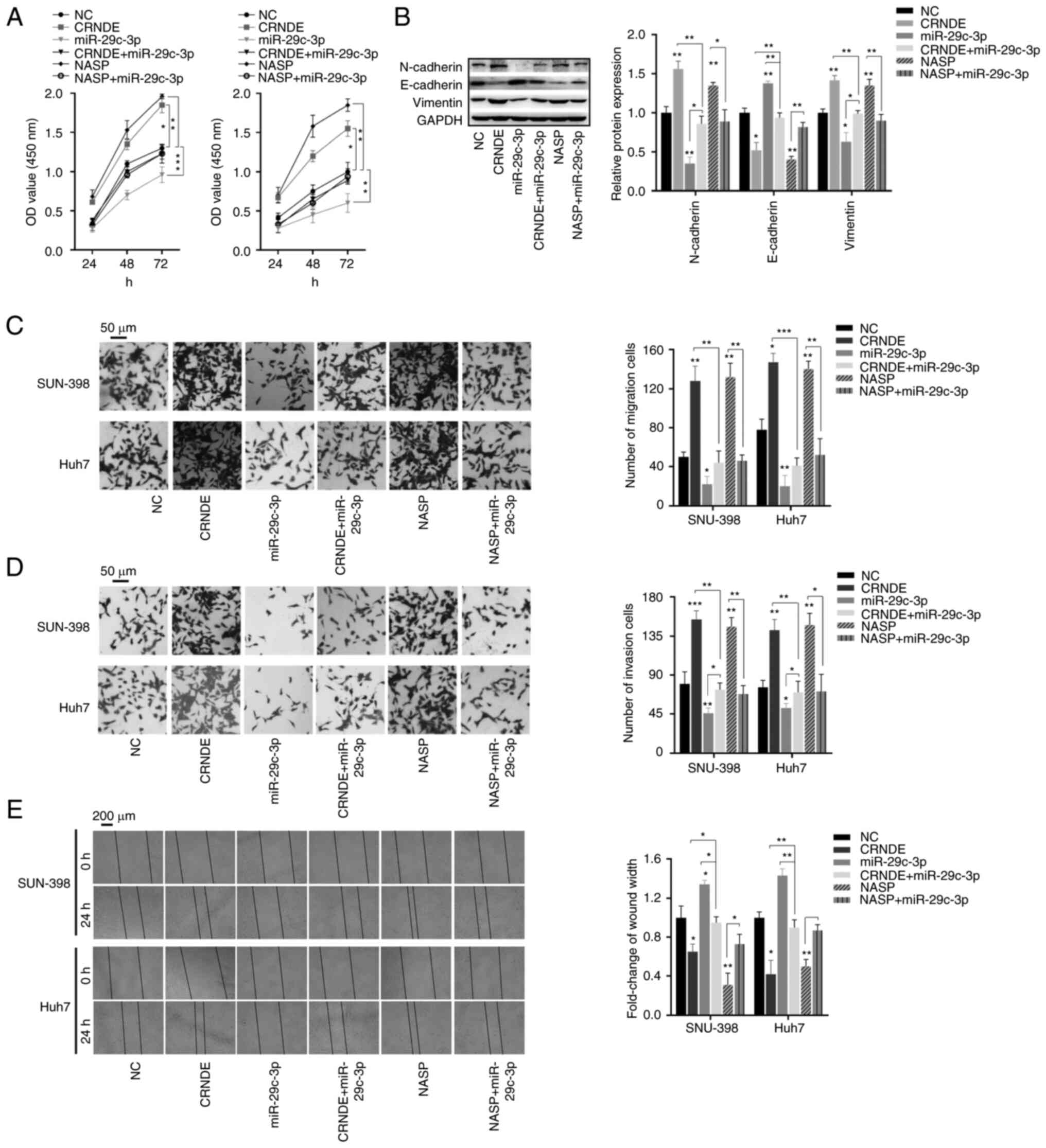 | Figure 6.CRNDE-miR-29c-3p-NASP axis regulates
HCC cell proliferation, migration and invasiveness. HCC cells were
transfected with CRNDE, miR-29c-3p mimics, NASP, CRNDE+miR-29c-3p
mimics, NASP+miR-29c-3p mimics or the corresponding negative
controls. (A) Proliferation ability of HCC cells following
transfection. (B) Expression levels of epithelial-mesenchymal
transition-related proteins were detected by western blotting in
HCC cells following transfection. Transwell assays were performed
to evaluate the (C) migration and (D) invasiveness of HCC cells
following transfection. (E) Wound-healing assays were used to
analyze HCC cell migration. *P<0.05, **P<0.01 and
***P<0.001. miR, microRNA; CRNDE, lncRNA colorectal neoplasia
differentially expressed; NASP, nuclear autoantigenic sperm
protein; HCC, hepatocellular carcinoma; sh, short hairpin
(RNA). |
Discussion
LncRNAs serve critical roles in various diseases,
including cancer. As such, dysregulation of lncRNAs is closely
involved in the tumorigenesis and clinical prognosis of cancer
(25). The lncRNA-mediated
progression of HCC has been widely studied. For example, knockdown
of lncRNA HOTAIR suppressed glycolysis by targeting the
miR-130a-3p/HIF1A axis in HCC during hypoxia (26). As a metastasis-associated molecule,
lncRNA AY927503 was found to be highly expressed and to correlate
with tumor metastasis and poor prognosis in patients with HCC by
stimulating ITGAV transcription (27). Furthermore, lncRNA SNHG16
accelerated HCC progression by regulating the expression of
autophagy-related gene P62 via miR-17-5p (28). In addition, lncRNA lncTCF7
activated wnt signaling to promote the self-renewal of liver cancer
stem cells (29). In the present
study, CRNDE was found to be significantly upregulated in HCC
compared with adjacent-normal tissues, suggesting that CRNDE is an
HCC-related lncRNA. These findings were consistent with previous
studies demonstrating that CRNDE functions as an oncogene in
multiple cancers, such as pancreatic cancer (30), hepatocellular carcinoma (31) and glioma (32). Thus, the function and clinical
significance of CRNDE warrants further investigation.
In loss-of-function assays, CRNDE-knockdown was
found to markedly suppress the proliferation, migration and
invasiveness of HCC cells, illustrating the oncogenic role of CRNDE
in HCC. However, the study did not reveal the role of CRNDE in HCC
in vivo, which requires additional investigation.
Furthermore, the molecular mechanism underlying the effects of
CRNDE in HCC remains unclear.
It is well known that lncRNAs perform their
functions via various mechanisms, such as RNA-protein interaction,
RNA-RNA pathways and the ceRNA pathway. In the present study,
miR-29c-3p was predicted as a target of CRNDE in HCC. As such, a
luciferase reporter assay confirmed a direct interaction between
CRNDE and miR-29c-3p. In previous studies, miR-29c-3p was
competitively bound by lncRNA RP11-480I12.5 to inhibit cancer
progression (33). As a tumor
suppressor, miR-29c-3p was negatively regulated by lncRNA MYOSLID
in gastric cancer (34). Combining
the revelation that miR-29c-3p expression was decreased in HCC
cells in the present study, miR-29c-3p was confirmed as a tumor
suppressor in HCC. Thus, we hypothesize that CRNDE plays an
oncogenic role in HCC by inhibiting miR-29c-3p expression.
Previous studies have indicated that the
overexpression of NASP promotes the proliferation, migration and
invasiveness of cancer cells (35). The carcinogenesis of NASP was also
discovered in gastric cancer (19), ovarian cancer (20) and HCC (21). In the present study, NASP was
identified as a direct target of miR-29c-3p in HCC cells. NASP was
also highly expressed in HCC tissues, which indicated poor
disease-free survival rates of patients with HCC. These findings
demonstrated that NASP plays the role of tumor promoter in HCC.
Furthermore, miR-29c-3p negatively regulated NASP expression in HCC
cells, but CRNDE exerted a positive effect on NASP expression.
Moreover, the positive and negative correlation of CRNDE or
miR-29c-3p on NASP, respectively, were determined in HCC tissues.
The overexpression of miR-29c-3p inhibited CRNDE and NASP
expression, and reduced the enhancement of CRNDE- or
NASP-associated proliferation, EMT, migration and invasiveness of
HCC cells. Thus, the results of the present study highlighted the
effect of the CRNDE/miR-29c-3p/NASP axis in HCC. The potential role
of the CRNDE/miR-29c-3p/NASP axis in HCC in vivo will be
investigated in future studies.
In summary, the results of the present study
indicated that CRNDE promoted the progression of HCC by inhibiting
cellular proliferation, migration and invasiveness. This was
achieved via direct targeting of miR-29c-3p to upregulate NASP
expression. Thus, the CRNDE/miR-29c-3p/NASP axis may be a potential
therapeutic target for HCC.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
GXi, YZ and BL designed and conceived the study. YZ,
BL, GXu and CH conducted the experiments. All authors analyzed the
data. GXi, YZ and BL drafted the manuscript. All authors have read
and approved the final manuscript. GXi and YZ confirmed the
authenticity of all the raw data.
Ethics approval and consent to
participate
The present study was approved by the Ethical
Committee of the Second Hospital of Tianjin Medical University.
Written informed consent was obtained from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yu LX and Schwabe RF: The gut microbiome
and liver cancer: Mechanisms and clinical translation. Nat Rev
Gastroenterol Hepatol. 14:527–539. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lau WY and Lai EC: Hepatocellular
carcinoma: Current management and recent advances. Hepatobiliary
Pancreat Dis Int. 7:237–257. 2008.PubMed/NCBI
|
|
4
|
Portolani N, Coniglio A, Ghidoni S,
Giovanelli M, Benetti A, Tiberio GA and Giulini SM: Early and late
recurrence after liver resection for hepatocellular carcinoma:
Prognostic and therapeutic implications. Ann Surg. 243:229–235.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kulik L and El-Serag HB: Epidemiology and
management of hepatocellular carcinoma. Gastroenterology.
156:477–491.e1. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mercer TR, Dinger ME and Mattick JS: Long
non-coding RNAs: Insights into functions. Nat Rev Genet.
10:155–159. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zeng Z, Dong J, Li Y, Dong Z, Liu Z, Huang
J, Wang Y, Zhen Y and Lu Y: The expression level and clinical
significance of lncRNA X91348 in hepatocellular carcinoma. Artif
Cells Nanomed Biotechnol. 47:3067–3071. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Yang MH, Zhao L, Wang L, Ou-Yang W, Hu SS,
Li WL, Ai ML, Wang YQ, Han Y, Li TT, et al: Nuclear lncRNA HOXD-AS1
suppresses colorectal carcinoma growth and metastasis via
inhibiting HOXD3-induced integrin β3 transcriptional activating and
MAPK/AKT signalling. Mol Cancer. 18:312019. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mu M, Niu W, Zhang X, Hu S and Niu C:
LncRNA BCYRN1 inhibits glioma tumorigenesis by competitively
binding with miR-619-5p to regulate CUEDC2 expression and the
PTEN/AKT/p21 pathway. Oncogene. 39:6879–6892. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Li H, Jia J, Yang L, Chu J, Sheng J, Wang
C, Meng W, Jia Z, Yin H, Wan J and He F: LncRNA MIR205HG drives
esophageal squamous cell carcinoma progression by regulating
miR-214/SOX4 axis. Onco Targets Ther. 13:13097–13109. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu Z, Zhou W, Lin C, Wang X, Zhang X,
Zhang Y, Yang R, Chen W and Cao W: Dysregulation of FOXD2-AS1
promotes cell proliferation and migration and predicts poor
prognosis in oral squamous cell carcinoma: A study based on TCGA
data. Aging (Albany NY). 13:2379–2396. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kan Z, Zheng H, Liu X, Li S, Barber TD,
Gong Z, Gao H, Hao K, Willard MD, Xu J, et al: Whole-genome
sequencing identifies recurrent mutations in hepatocellular
carcinoma. Genome Res. 23:1422–1433. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang Y, Lu JH, Wu QN, Jin Y, Wang DS, Chen
YX, Liu J, Luo XJ, Meng Q, Pu HY, et al: LncRNA LINRIS stabilizes
IGF2BP2 and promotes the aerobic glycolysis in colorectal cancer.
Mol Cancer. 18:1742019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li Z, Hou P, Fan D, Dong M, Ma M, Li H,
Yao R, Li Y, Wang G, Geng P, et al: The degradation of EZH2
mediated by lncRNA ANCR attenuated the invasion and metastasis of
breast cancer. Cell Death Differ. 24:59–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang F, Wang H, Yu J, Yao X, Yang S, Li
W, Xu L and Zhao L: LncRNA CRNDE attenuates chemoresistance in
gastric cancer via SRSF6-regulated alternative splicing of PICALM.
Mol Cancer. 20:62021. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen C, Huang Z, Mo X, Song Y, Li X, Li X
and Zhang M: The circular RNA 001971/miR-29c-3p axis modulates
colorectal cancer growth, metastasis, and angiogenesis through
VEGFA. J Exp Clin Cancer Res. 39:912020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu H, Zhang W, Wu Z, Liu Y, Shi Y, Gong J,
Shen W and Liu C: miR-29c-3p regulates DNMT3B and LATS1 methylation
to inhibit tumor progression in hepatocellular carcinoma. Cell
Death Dis. 10:482019. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yu B, Chen X, Li J, Gu Q, Zhu Z, Li C, Su
L and Liu B: microRNA-29c inhibits cell proliferation by targeting
NASP in human gastric cancer. BMC Cancer. 17:1092017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ali-Fehmi R, Chatterjee M, Ionan A, Levin
NK, Arabi H, Bandyopadhyay S, Shah JP, Bryant CS, Hewitt SM, O'Rand
MG, et al: Analysis of the expression of human tumor antigens in
ovarian cancer tissues. Cancer Biomark. 6:33–48. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kang X, Feng Y, Gan Z, Zeng S, Guo X, Chen
X, Zhang Y, Wang C, Liu K, Chen X, et al: NASP antagonize chromatin
accessibility through maintaining histone H3K9me1 in hepatocellular
carcinoma. Biochim Biophys Acta Mol Basis Dis. 1864:3438–3448.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang L, Cho KB, Li Y, Tao G, Xie Z and Guo
B: Long noncoding RNA (lncRNA)-mediated competing endogenous RNA
networks provide novel potential biomarkers and therapeutic targets
for colorectal cancer. Int J Mol Sci. 20:57582019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chan JJ and Tay Y: Noncoding RNA:RNA
regulatory networks in cancer. Int J Mol Sci. 19:13102018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
He Y, Meng XM, Huang C, Wu BM, Zhang L, Lv
XW and Li J: Long noncoding RNAs: Novel insights into hepatocelluar
carcinoma. Cancer Lett. 344:20–27. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hu M, Fu Q, Jing C, Zhang X, Qin T and Pan
Y: LncRNA HOTAIR knockdown inhibits glycolysis by regulating
miR-130a-3p/HIF1A in hepatocellular carcinoma under hypoxia. Biomed
Pharmacother. 125:1097032020. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kang CL, Qi B, Cai QQ, Fu LS, Yang Y, Tang
C, Zhu P, Chen QW, Pan J, Chen MH and Wu XZ: LncRNA AY promotes
hepatocellular carcinoma metastasis by stimulating ITGAV
transcription. Theranostics. 9:4421–4436. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhong JH, Xiang X, Wang YY, Liu X, Qi LN,
Luo CP, Wei WE, You XM, Ma L, Xiang BD and Li LQ: The lncRNA SNHG16
affects prognosis in hepatocellular carcinoma by regulating p62
expression. J Cell Physiol. 235:1090–1102. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang Y, He L, Du Y, Zhu P, Huang G, Luo J,
Yan X, Ye B, Li C, Xia P, et al: The long noncoding RNA lncTCF7
promotes self-renewal of human liver cancer stem cells through
activation of Wnt signaling. Cell Stem Cell. 16:413–425. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang G, Pan J, Zhang L, Wei Y and Wang C:
Long non-coding RNA CRNDE sponges miR-384 to promote proliferation
and metastasis of pancreatic cancer cells through upregulating
IRS1. Cell Prolif. 50:e123892017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ji D, Jiang C, Zhang L, Liang N, Jiang T,
Yang B and Liang H: LncRNA CRNDE promotes hepatocellular carcinoma
cell proliferation, invasion, and migration through regulating
miR-203/BCAT1 axis. J Cell Physiol. 234:6548–6560. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zheng J, Liu X, Wang P, Xue Y, Ma J, Qu C
and Liu Y: CRNDE promotes malignant progression of Glioma by
attenuating miR-384/PIWIL4/STAT3 axis. Mol Ther. 24:1199–1215.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lou W, Ding B, Zhong G, Yao J, Fan W and
Fu P: RP11-480I12.5-004 promotes growth and tumorigenesis of breast
cancer by relieving miR-29c-3p-mediated AKT3 and CDK6 degradation.
Mol Ther Nucleic Acids. 21:916–931. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Han Y, Wu N, Jiang M, Chu Y, Wang Z, Liu
H, Cao J, Liu H, Xu B and Xie X: Long non-coding RNA MYOSLID
functions as a competing endogenous RNA to regulate MCL-1
expression by sponging miR-29c-3p in gastric cancer. Cell Prolif.
52:e126782019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kong F, Li L, Wang C, Zhang Q and He S:
MiR-381-3p suppresses biological characteristics of cancer in
head-neck squamous cell carcinoma cells by targeting nuclear
autoantigenic sperm protein (NASP). Biosci Biotechnol Biochem.
84:703–713. 2020. View Article : Google Scholar : PubMed/NCBI
|