Introduction
Esophageal cancer (EsC) ranks seventh among all
malignancy-related deaths worldwide (1). The prognosis of esophageal malignancies
is particularly poor because these tumors typically do not cause
any symptoms in the early stages. Therefore, most EsC patients are
diagnosed with advanced stages of the diseases which are
accompanied by distant metastasis, making them ineligible for
radical surgery (2,3). This complication leads to a low 5-year
survival rate, and although this rate has increased over time, it
remains at only 18% (2). Esophageal
squamous cell carcinoma (ESCC) is the dominant histological type of
EsC in southeastern Asia, especially in China (4). The risk factors for chronic cell damage
include smoking, alcohol abuse, thermal damage and nutrient
deficiencies. Furthermore, infiltration and metastasis are the two
main causes of death from ESCC (2,5). Recent
studies have revealed that tumor cells affect the surrounding
microenvironment, pierce through the surrounding barrier, migrate
into peritumoral tissues from the primary site, and contribute to
regional invasion and widespread metastasis (6). However, the concrete mechanisms by
which ESCC metastasizes remains unknown. Thus, identification of
the potential specific biomarkers involved in tumor metastasis will
help improve the chances of an early diagnosis, formulate
appropriate clinical treatment plans and prolong the survival of
ESCC patients.
Ras-association domain family 10 (RASSF10)
located on chromosome 11p15.2, is both a candidate tumor-suppressor
gene and the latest identified member of the RASSF family (7,8).
RASSF10, as well as RASSF7-9 of this family, which share a
conserved Ras association domain in the N-terminal region,
demonstrate several tumor-suppressive properties (8,9).
Accumulating evidence suggests that downregulation or inactivation
of RASSF10 by hypermethylation is extensively involved in the
promotion of cell proliferation and metastasis, which has been
observed in many human malignancies, such as astrocytic glioma,
malignant melanoma of the skin, hepatocellular carcinoma, thyroid
cancer and childhood acute lymphoblastic leukemia (10–14).
However, to the best of our knowledge, further studies of the
association between RASSF10 and ESCC, including the clinical
significance and cellular biological mechanisms underlying the
participation of RASSF10 in these malignant phenotypes, are
urgently warranted.
In the present study, we aimed to investigate the
clinical value of RASSF10 expression in ESCC patients. Furthermore,
on the strength of previous research on RASSF10, the potential
functions of RASSF10 in ESCC were investigated.
Materials and methods
Tissue specimens
This project was authorized by permission of the
Ethics Committee of the Affiliated Hospital of Nantong University
(Nantong, China), and signed informed-consent forms were collected
from all patients. Relevant clinical data on each patient were
collected from his or her medical records. The ESCC tissues and
paired normal adjacent mucosa tissues were derived from 187 ESCC
patients who had undergone surgery at the Affiliated Hospital of
Nantong University between January 2008 and December 2014. The mean
age of patients at the time of surgery was 53.81 years (range,
21–88 years). The percentage of male or female patients accounted
for 67.91 and 32.09% of the ESCC patients, respectively.
Histological classifications and stages per the TNM Classification
of Malignant Tumors (TNM stages) (https://www.uicc.org/resources/tnm-classification-malignant-tumours-8th-edition)
were determined independently by two senior pathologists according
to the classification criteria of the American Joint Committee on
Cancer. Resected samples were snap-frozen in liquid nitrogen until
RNA and protein extraction could be performed. After the enrolled
ESCC patients underwent surgery, they were treated in strict
accordance with therapeutic schedule on the basis of ESCC treatment
guidelines. The ESCC patients who were diagnosed with
pathologically confirmed lymph node metastases were able to select
treatment with chemotherapy of 2–4 cycles. Chemoradiotherapy using
albumin bound paclitaxel plus platinum antineoplastic agent was
used in the treatment of the patients with ESCC.
Cell lines and transfection
We purchased the normal esophageal epithelium cell
line Het-1a and ESCC cell lines (TE-10, ECA-109 and KYSE-150) from
the Shanghai Institute for Biological Sciences, Chinese Academy of
Sciences. Het-1a, TE-10, ECA-109 and KYSE-150 cells were maintained
in RPMI-1640 or DMEM, supplemented with 10% fetal bovine serum
(FBS), ampicillin (50 U/ml) and streptomycin (50 µg/ml), in a
humidified chamber at 37°C with 5% CO2. Notably, TE-10
and ECA-109 cells were transfected with plasmids encoding RASSF10,
or shRNA against RASSF10, along with a vector control. The cDNA
encoding full-length human RASSF10 was cloned into the PCDH vector.
The shRNA targeting sequence for RASSF10 was
5′-GGAAAUUCCUUGAAUGUUAAU-3′. The expression constructs were
verified by DNA sequencing. RASSF10 expression in the transfected
cells was validated by western blot analysis.
RNA isolation and quantitative
real-time polymerase chain reaction (qPCR)
Total RNA isolation and qPCR analyses of
RASSF10 expression were performed as previously reported
(15). The relative mRNA expression
levels of RASSF10 were calculated in human tissues using the
2−ΔΔCq method (16). The
primer sequences of RASSF10 were designed as follows:
forward primer (5′-CAGAGCAGGGTGAGGGTAGA-3′) and reverse primer
(5′-GCTCGGACCACGTCAATTTC-3′). The primer sequences of GAPDH
were designed as follows: forward primer
(5′-CAGGAGGCATTGCTGATGAT-3′) and reverse primer
(5′-GAAGGCTGGGGCTCATTT-3′). All experiments were repeated three
times.
Immunohistochemistry (IHC)
analysis
For the streptavidin-peroxidase (S-P) IHC studies,
the primary antibodies were as follows: anti-RASSF10 (1:200; cat.
no. Ab113105; Abcam), anti-E-cadherin (1:500; cat. no. ab40772;
Abcam) and anti-vimentin (1:500; cat. no. ab8978; Abcam). Staining
intensity and the fraction of positive cells were scored according
to 10 randomly selected visual fields. The immunoreactive scores
(IRS) for the proportion of positive cells and the staining grade
were calculated for each specimen.
Scoring criteria
The percentages of positive cells were calculated as
follows: 0 (≤5%), 1 (>5–25%), 2 (>25–50%) and 3 (>50%).
Staining intensity was quantified according to four classification:
0 (negative), 1 (weak positive), 2 (moderate positive), and 3
(strong positive). According to the scoring criteria above, the
expression levels of RASSF10 or E-cadherin or vimentin in the ESCC
tissues were classified into two groups: samples with IRS >3
defined as high expression and the samples with IRS ≤3 defined as
low expression.
Protein extraction and western blot
analysis
Protein isolation and western blot analysis were
performed as previously reported (17). The following commercial antibodies
against RASSF10 (1:500; cat. no. Ab113105), E-cadherin (1:20,000;
cat. no. Ab40772), vimentin (1:2,000; cat. no. Ab8978), β-catenin
(1:3,000; cat. no. Ab16051), GAPDH (1:3,000; cat. no. Ab9485) and
Lamin A (1:5,000; cat. no. Ab226198) followed by secondary antibody
antibodies (1:5,000; cat. no. Ab6728; 1:10,000; cat. no. Ab6728)
purchased from Abcam were used. All experiments were repeated three
times.
Cell proliferation assay
Cell proliferation was monitored using the Cell
Counting Kit-8 (CCK-8) based on the manufacturer's protocol
(Dojindo). The transfected ESCC cell lines (TE-10 and ECA-109) were
incubated at a density of 3,000 cells per well in a 96-well plate.
Cell viability was determined at daily intervals (every 24 h)
following the absorbance measured at 450 nm. All experiments were
repeated three times.
Colony formation assay
For the colony formation assay, the transfected ESCC
lines (TE-10 and ECA-109) in the logarithmic growth phase were
plated into 6-well plates at a density of 300 cells per well and
maintained for two weeks in medium containing G418 (1,000 µg/ml).
The colonies were washed with PBS three times, and then fixed with
paraformaldehyde and stained with crystal violet (0.1%) at 25°C for
10 min. The colony formation was measured by photographing the
stained colonies under a microscope (magnification, ×5; Olympus
Corp.). Colony formation was assessed using MShot Image Analysis
System (Guangzhou Mingmei Photoelectric Technology Co., Ltd.). All
experiments were repeated three times.
Immunofluorescence
Immunofluorescence staining analysis was performed
as previously reported using a confocal laser-scanning microscope
(Carl Zeiss) (18). To assess the
protein subcellular localization, the data were analyzed with Adobe
Photoshop 7.0 software.
Luciferase reporter assay
TOPflash/FOPflash reporters (Millipore) were used in
this assay. The cells were transiently transfected with 1 mg of
either TOPFlash or FOPFlash, and 0.1 mg pRLTK
(Renilla-TK-luciferase vector, Promega Corp.) following the
manufacturer's protocol. Firefly and Renilla luciferase
activities were conducted employing the Dual-Luciferase Reporter
Assay System (Promega Corp.) after a 48-h transfection. The
activity of firefly luciferase was normalized to that of
Renilla luciferase as a transfection control. All
experiments were repeated three times.
Transwell invasion assays
Cell motility was determined by cell invasion assays
using 8-µm Transwell filters (BD Biosciences) with Matrigel (BD
Biosciences). The treated ECA109 and TE-10 cells at a density of
5×104 were seeded into the upper chamber with medium
that did not contain serum; medium containing serum was placed into
the lower chambers as a chemoattractant. After incubation for 48 h,
the treated cells on the upper surface of the membrane were stained
with crystal violet (0.1%) at 25°C for 10 min followed by manual
counting under a microscope (magnification, ×20; Olympus Corp.) in
three randomly selected areas. The Transwell assays were assessed
using MShot Image Analysis System (Guangzhou Mingmei Photoelectric
Technology Co., Ltd.). All experiments were repeated three
times.
Wound healing assay
TE-10 and ECA-109 cells were plated onto 6-well
plates to encourage the formation of a monolayer of cells. A 200-µl
pipette tip was employed to vertically scratch the cells in the
middle of the well. The floating debris was then washed away. Fresh
medium containing 0.1% FBS was supplemented and images of the cells
were taken with a light microscope (magnification, ×100; Olympus
Corp.) at 0 and 48 h to determine cell migration. The wound healing
assays were assessed using MShot Image Analysis System (Guangzhou
Mingmei Photoelectric Technology Co., Ltd.). All experiments were
repeated three times.
Inhibitor treatment
The Wnt inhibitor IWR-1 was purchased from ENZO.
Approximately 24 h before transfection, the treated ECA109 cells
were plated in 6-well plates at 35–55% confluence. Cells were then
transfected with the Wnt inhibitor IWR-1 at a working concentration
of 10 µM, respectively, using Lipofectamine 2000 (Invitrogen;
Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. After 48–72 h, the cells were collected for subsequent
experiments.
Statistical analysis
Statistical analysis was carried out using SPSS
software 22.0 (SPSS, Inc.) and GraphPad Prism 5.0 software
(GraphPad Software, Inc.). The Chi-squared test or Student's t-test
was employed to compare the differences between two groups.
Spearman rank correlations test was used to detect the relationship
between expression of two genes. Other data were evaluated
employing the Mann-Whitney U test as applicable to determine
significant differences. Univariate and multivariate Cox regression
model analyses were used to determine the prognostic relevant
factors. Kaplan-Meier method was used to calculate the survival
function and draw the survival curve. A P-value <0.05 was
considered statistically significant.
Results
RASSF10 is downregulated in human ESCC
tissues
To explore the clinical significance of RASSF10 in
ESCC progression, qPCR was employed to measure the expression of
the RASSF10 mRNA level extracted from the collected 20 pairs
of ESCC patient tissues and matched adjacent normal tissues. Our
results revealed that the RASSF10 mRNA expression level in
the ESCC cancerous tissue samples was significantly downregulated
compared with that in the corresponding adjacent normal tissue
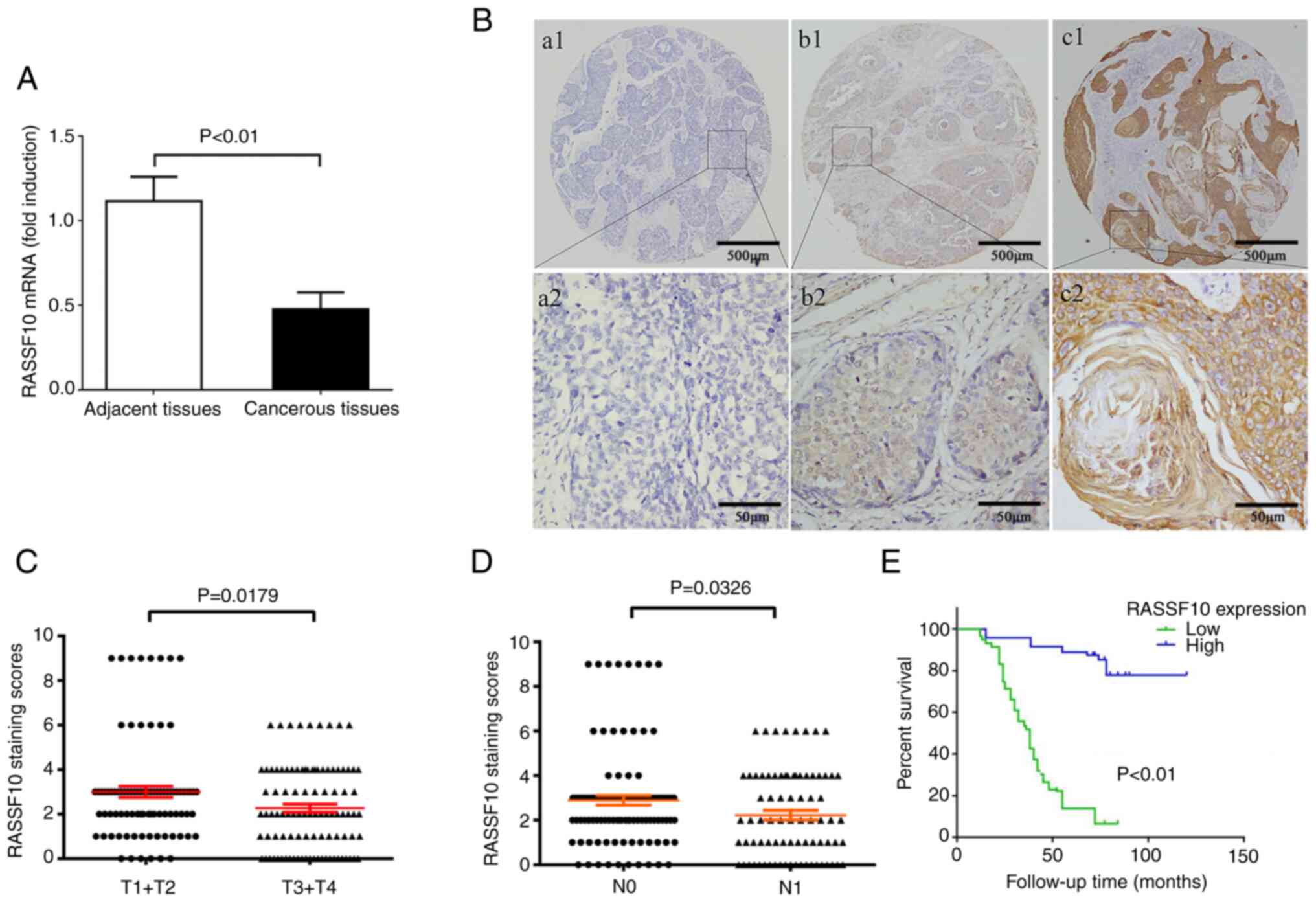
samples (Fig. 1A, P<0.01).
TMA-IHC was further conducted to determine the RASSF10 protein
status on a cohort of 187 samples of ESCC tissues and 154
peritumoral tissues. The distribution of the different levels of
RASSF10 expression in ESCC and peritumoral tissues is shown in
Table I. The expression levels of
RASSF10 were markedly higher in the peritumoral tissue samples than
that in the tumor tissues (P<0.001). Our results indicated that
the RASSF10 protein was mainly localized in the cytoplasm of ESCC
cells (Fig. 1B). These results
demonstrated that RASSF10 is downregulated in ESCC.
 | Table I.Immunohistochemical analysis of
RASSF10 expression in ESCC. |
Table I.
Immunohistochemical analysis of
RASSF10 expression in ESCC.
|
|
| RASSF10
expression |
|
|---|
|
|
|
|
|
|---|
| Group | No. of cases | Low (1–3) | High (4–9) | P-value |
|---|
| Adjacent normal
tissues | 154 | 51 | 103 | <0.001 |
| ESCC tissues | 187 | 142 | 45 |
|
Clinical significance of RASSF10 in
ESCC
Furthermore, we assessed the association between
activated cytoplasmic RASSF10 expression and the clinical
pathological parameters in ESCC patients. The data demonstrated
that RASSF10 was markedly associated with differentiation (P=0.001)
(Fig. 1B), primary tumor (P=0.014),
lymph node metastasis (P=0.02) and TNM stage (P 0.006) (Table II). As shown in Fig. 1C, the immunoreaction scores of the
RASSF10 status in T3-T4 stages was markedly lower in comparison
with that of T1-T2 stages (P=0.0179). It was also found that ESCC
patients who developed metastasis had markedly lower immunoreaction
scores for RASSF10 compared with than those of non-metastatic
patients (P=0.0326) (Fig. 1D).
| Table II.Association of RASSF10 expression
with clinicopathological features in the ESCC patients. |
Table II.
Association of RASSF10 expression
with clinicopathological features in the ESCC patients.
|
|
| RASSF10 expression
in tumor cells |
|---|
|
|
|
|
|---|
| Clinicopathologic
characteristics | Cases | Low or
negative | High | P-value |
|---|
| Total | 187 | 142 | 45 |
|
| Sex |
|
|
| 0.348 |
|
Male | 127 | 99 | 28 |
|
|
Female | 60 | 43 | 17 |
|
| Age (years) |
|
|
| 0.486 |
|
≤60 | 138 | 103 | 35 |
|
|
>60 | 49 | 39 | 10 |
|
| Tumor size |
|
|
| 0.394 |
| <3
cm | 114 | 89 | 25 |
|
| ≥3
cm | 73 | 53 | 20 |
|
|
Differentiation |
|
|
| 0.001a |
| Low
grade | 69 | 43 | 26 |
|
| Middle
grade | 59 | 45 | 14 |
|
| High
grade | 59 | 54 | 5 |
|
| Primary tumor |
|
|
| 0.014a |
|
T1+T2 | 88 | 74 | 14 |
|
|
T3+T4 | 99 | 68 | 31 |
|
| Lymph node
metastasis |
|
|
| 0.020a |
| N0 | 107 | 88 | 19 |
|
| N1 | 80 | 54 | 26 |
|
| TNM stage |
|
|
| 0.006a |
| I | 33 | 18 | 15 |
|
| II | 65 | 51 | 14 |
|
|
III | 89 | 73 | 16 |
|
Next, Kaplan-Meier analysis combined with IHC
analysis demonstrated that low expression of RASSF10 was correlated
with poor overall survival in ESCC patients (P<0.01) (Fig. 1E). As shown in Table III, multivariate Cox regression
analysis confirmed that low cytoplasmic RASSF10 expression was a
marked independent prognostic factor (P=0.002). Collectively, our
results indicated that there is a convincing association between
low RASSF10 expression and poor outcome in ESCC patients.
| Table III.Univariate and multivariate analysis
of the association of prognosis with the clinicopathologic
parameters and RASSF10 expression in the ESCC patients. |
Table III.
Univariate and multivariate analysis
of the association of prognosis with the clinicopathologic
parameters and RASSF10 expression in the ESCC patients.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Characteristic | HR | P-value | 95% CI | HR | P-value | 95% CI |
|---|
| RASSF10
expression | 3.661 | 0.007a | 1.437-9.326 | 3.444 | 0.002a | 1.560-7.604 |
| High
vs. low or negative |
|
|
|
|
|
|
| Sex | 0.661 | 0.096 | 0.406-1.077 |
|
|
|
| Male
vs. Female |
|
|
|
|
|
|
| Age (years) | 1.290 | 0.229 | 0.852-1.953 |
|
|
|
| ≤60 vs.
>60 |
|
|
|
|
|
|
| Tumor size
(cm) | 1.205 | 0.551 | 0.653-2.226 |
|
|
|
| <3
vs. ≥3 |
|
|
|
|
|
|
|
Differentiation | 0.799 | 0.115 | 0.605-1.056 |
|
|
|
| Low vs.
middle and high grade |
|
|
|
|
|
|
| Primary tumor | 1.785 | 0.035a | 1.041-3.061 | 1.871 | 0.018a | 1.112-3.147 |
| T1+T2
vs. T3+T4 |
|
|
|
|
|
|
| Lymph node
metastasis | 1.489 | 0.014a | 1.082-2.047 | 1.447 | 0.021a | 1.059-1.979 |
|
N0 vs.
N1 |
|
|
|
|
|
|
| TNM stage |
|
|
|
|
|
|
| I vs.
II vs. III | 1.880 | 0.022a | 1.139-3.101 | 1.686 | 0.033a | 1.042-2.727 |
Effect of RASSF10 on ESCC cell
proliferation
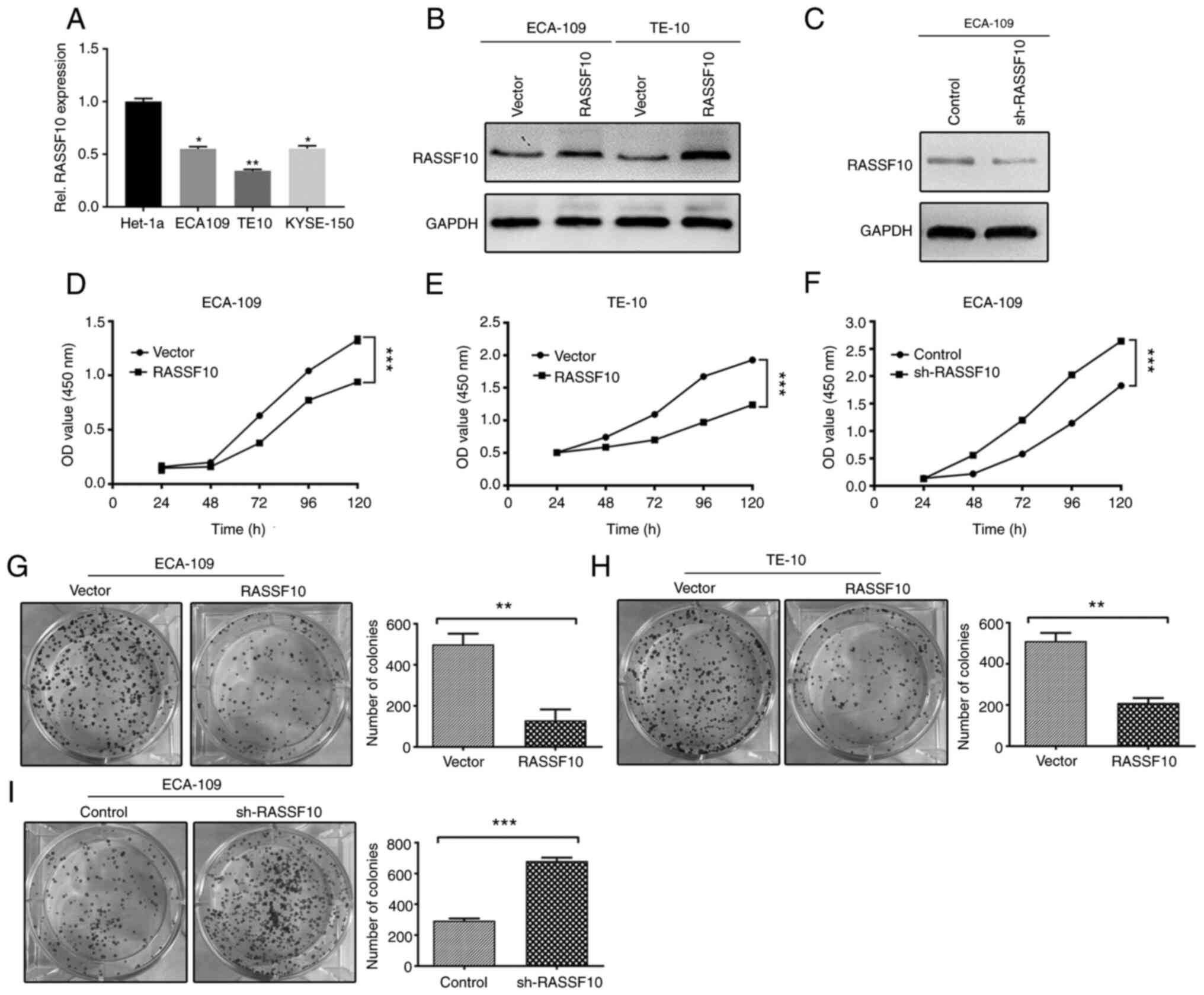
To explore the role of RASSF10 in the progression of
ESCC, endogenous expression of RASSF10 was assessed by qPCR
in normal esophageal epithelium cell line Het-1a and human ESCC
cell lines TE10, ECA109 and KYSE-150. RASSF10 mRNA levels
were lower in ESCC cell lines compared with that in the Het-1a
cells (Fig. 2A). We then generated
stable infected ECA-109 and TE-10 cells with RASSF10
overexpression, and the ECA-109 cell line was selected to silence
RASSF10 expression. Western blot analysis was used to verify the
transfection efficiency of the treated cell lines (Fig. 2B and C). Increased proliferation is
one of most malignant phenotypes of cancer cells and plays an
important part in cancer progression (19). Thus, CCK-8 and colony formation
assays were used to evaluated the proliferative capabilities of the
mentioned cell models. As shown in Fig.
2D and E, ectopic expression of RASSF10 dramatically suppressed
the cell growth of the ECA-109 and TE-10 cells compared with the
empty vector group, and the number of colonies which had formed in
the RASSF10-transfected ECA-109 and TE-10 cells were significantly
decreased in keeping with this finding (Fig. 2G and H), In contrast, RASSF10
knockdown resulted in markedly reverse effects (Fig. 2F and I). These results suggest that
RASSF10 is crucial for inhibiting ESCC cell growth.
Effect of RASSF10 on ESCC cell
migration and invasion
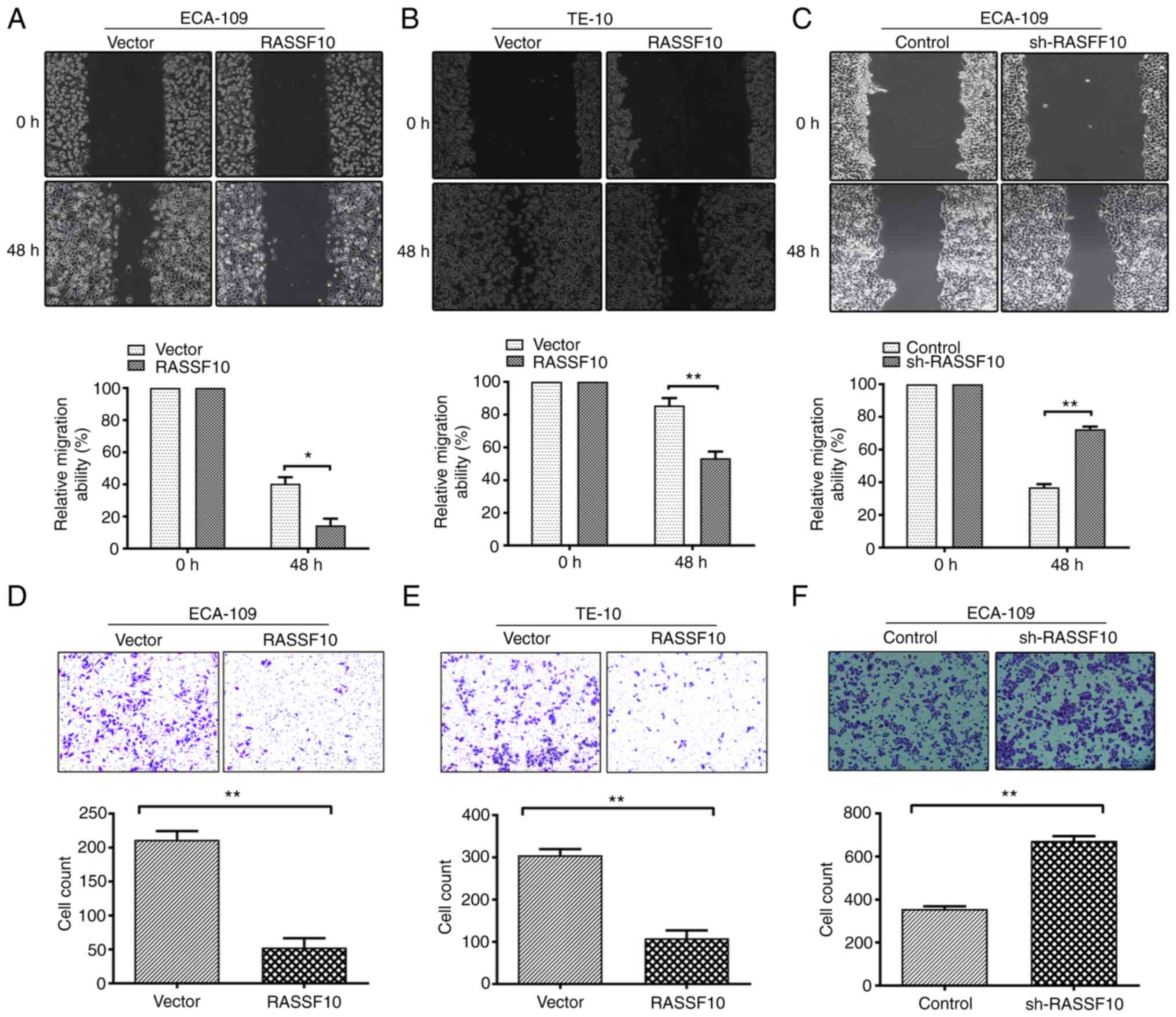
Since increased capability for motility is
indispensable for tumor metastasis, wound healing and Transwell
invasion assays were performed. Ectopic expression of RASSF10
significantly inhibited cell migration capability in the ECA-109
and TE-10 cells compared with the vector-transfected cells
(Fig. 3A and B). The
RASSF10-expressing cells exhibited a marked decrease in reduction
in wound closure in comparison with the control groups by
quantitative analyses at 48 h. By comparison, knockdown of RASSF10
significantly facilitated the migration of ECA-109 cells (Fig. 3C). Transwell invasion assays were
also conducted. As expected, it was demonstrated that RASSF10
overexpression markedly impaired the invasive capability of the
ECA-109 and TE-10 cells compared with that of the control groups
while RASSF10 knockdown increased the invasive ability of the cells
(Fig. 3D-F). Taken together, our
results indicated the capability of RASSF10 to inhibit ESCC cell
migration and invasion.
Correlation between RASSF10 and EMT
biomarkers in ESCC tissues and cell lines
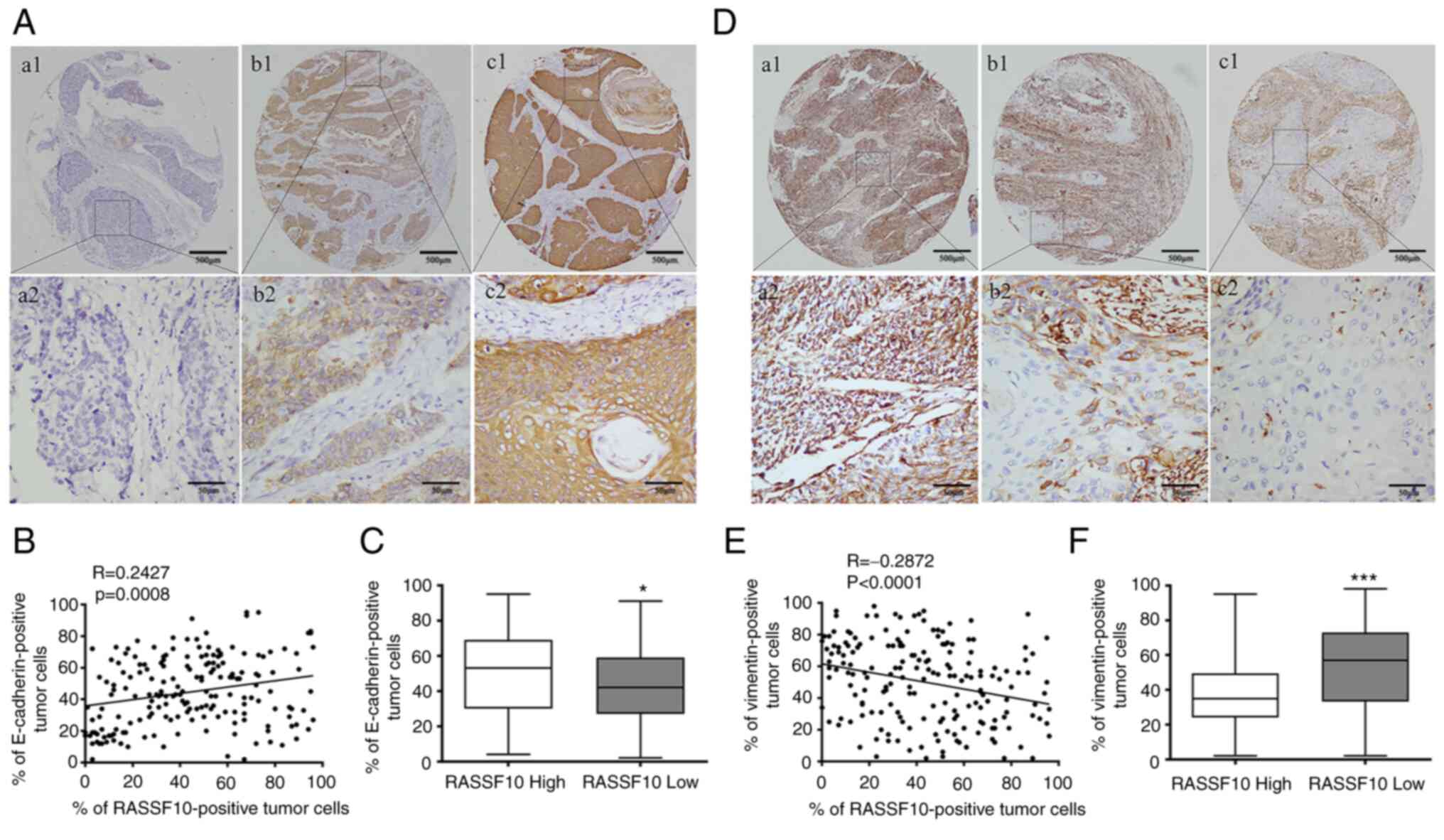
A growing body of studies have revealed that EMT
plays a crucial part in ESCC invasion and metastasis cascade, and
our above-mentioned results suggested that decreased expression of
RASSF10 was correlated with tumor metastasis in ESCC patients
(20–22). Therefore, we investigated the protein
expression levels of EMT markers, E-cadherin (epithelial phenotype)
and vimentin (mesenchymal phenotype), using the same TMA samples to
further explore the role of RASSF10 in the EMT process in ESCC. IHC
analysis demonstrated that tumors with lower RASSF10 expression
exhibited lower E-cadherin expression levels and higher vimentin
expression levels (Fig. 4A and D).
According to the Spearman rank correlations test, a positive
correlation was observed between RASSF10 and E-cadherin (R=0.2427,
P=0.0008) (Fig. 4B), and a negative
correlation was observed between RASSF10 and vimentin (R=−0.2872,
P<0.0001) (Fig. 4E). ESCC
patients with lower RASSF10 expression had significantly lower
E-cadherin expression than those with higher RASSF10 expression
(P=0.0295, Fig. 4C). We also found
the ESCC patients with RASSF10 low status exhibited a higher
vimentin-positive rate compared with that of the patients with high
RASSF10 expression (P=0.0003, Fig.
4F).
To further explore the effect of RASSF10 on the EMT
process, western blotting was employed to detect the protein levels
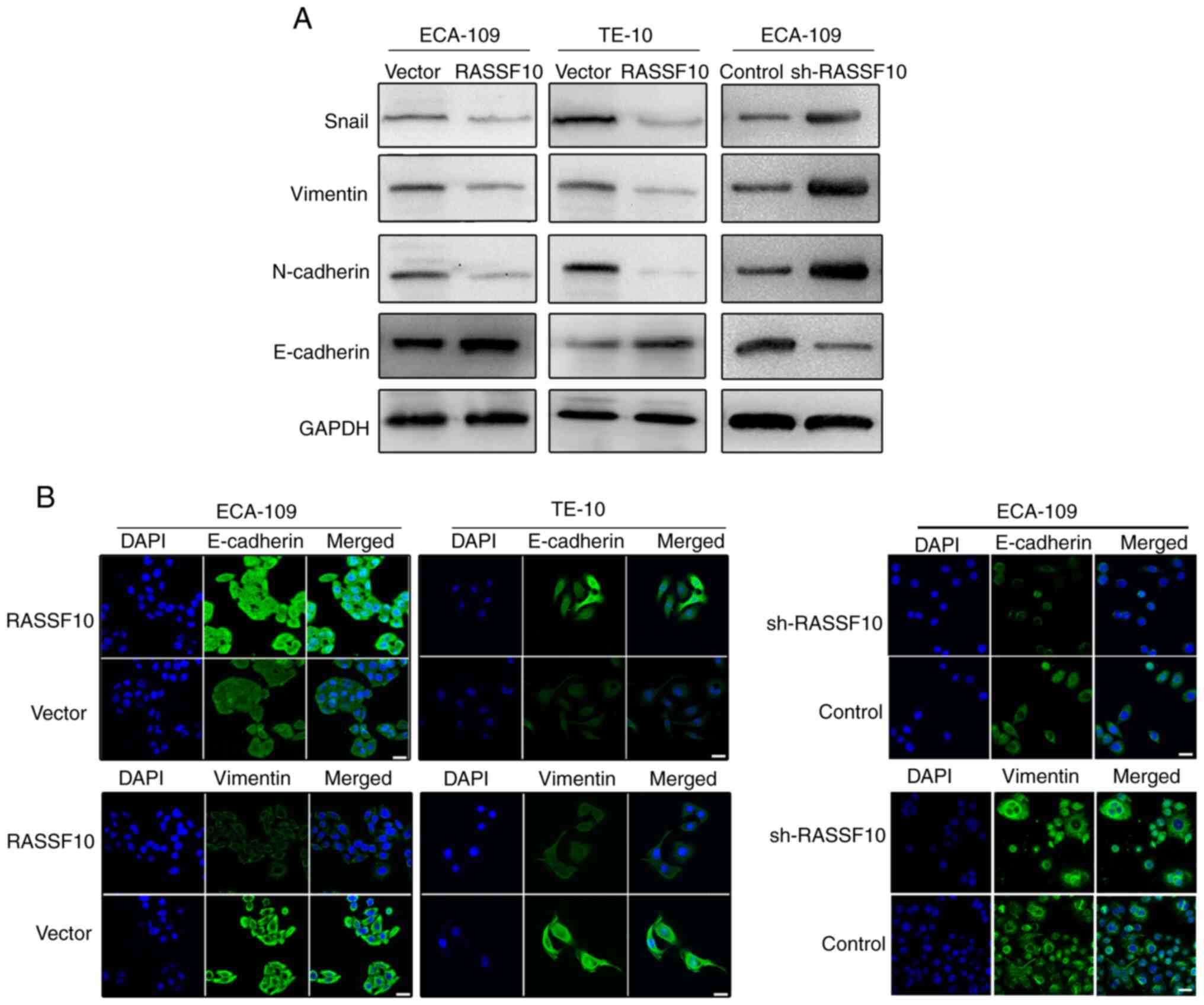
of EMT biomarkers. The results indicated that the protein level of
E-cadherin was markedly upregulated, while Snail, vimentin and
N-cadherin levels were significantly downregulated in the
RASSF10-overexpressing cells compared with those in the control
group (Fig. 5A). In contrast,
knockdown of RASSF10 decreased the expression of E-cadherin and
increased the levels of Snail, vimentin and N-cadherin in the
ECA-109 cells. These findings suggest that RASSF10 may be involved
in suppression of the EMT process. Subsequently, our results using
immunofluorescence analysis indicated that the
RASSF10-overexpressing cells exhibited higher level of E-cadherin
and lower level of vimentin when compared with the control groups
(Fig. 5B). Knockdown of RASSF10
reversed this phenotype (Fig. 5B).
Our results indicate that RASSF10 may play a crucial part in the
regulation of tumor EMT and may control tumor invasion and
metastasis by inhibiting the EMT process.
Overexpression of RASSF10 attenuates
the Wnt/β-catenin signaling pathway
Previous studies suggest that RASSF10 suppresses
gastric cancer cell progression by inhibiting the Wnt/β-catenin
signaling pathway (23). Therefore,
we hypothesized that inactivation of the Wnt/β-catenin signaling
pathway may be involved in the RASSF10-mediated inhibition of ESCC
cell progression. In general, the Wnt/β-catenin pathway can be
mediated by stabilized accumulation and relocalization of
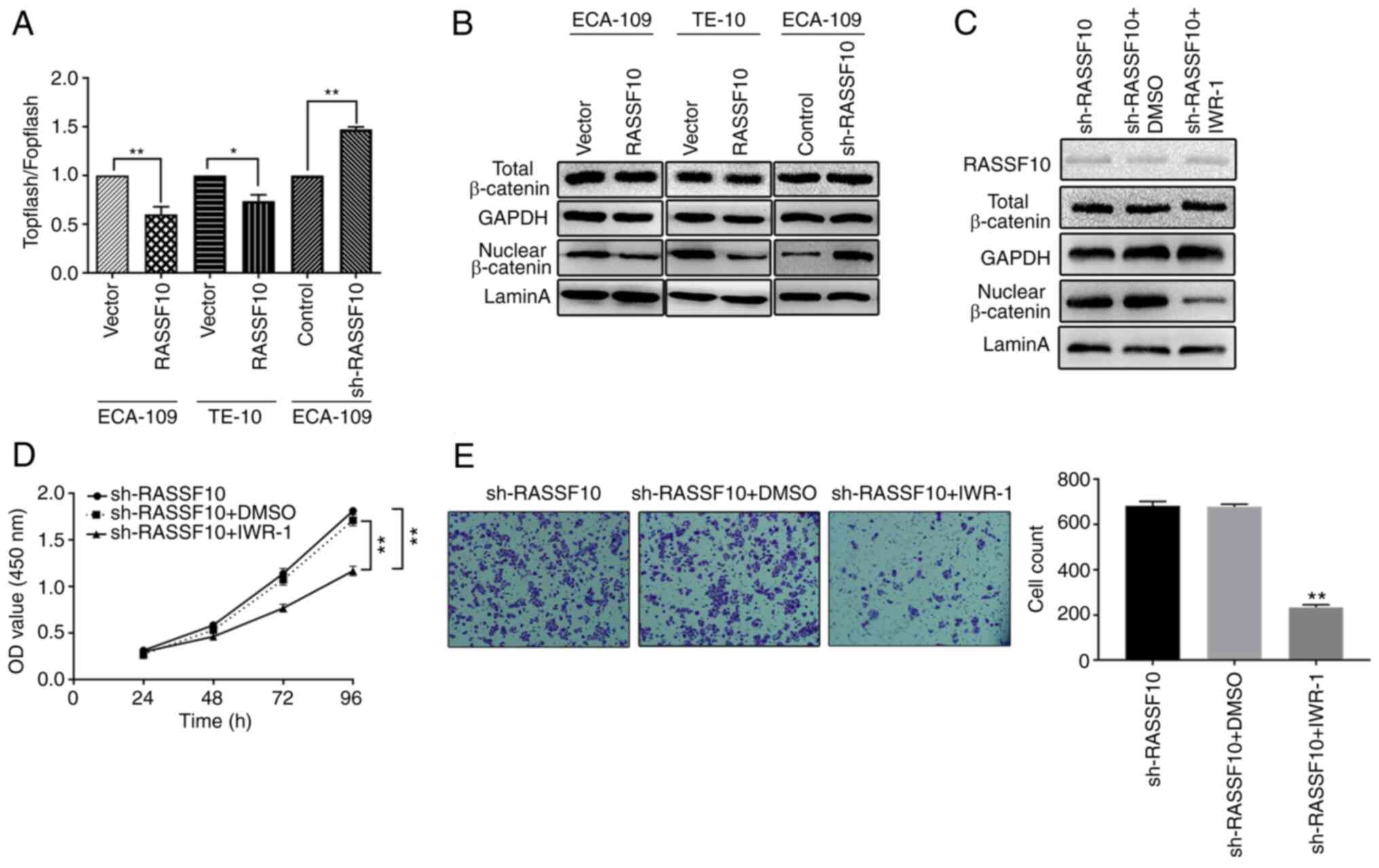
Wnt/β-catenin. We first verified the activities of the
Wnt/β-catenin pathway employing TOPflash/FOPflash luciferase assay
in ECA-109 and TE-10 cells overexpressing RASSF10. As shown in
Fig. 6A, we observed that relative
TOPflash/FOPflash luciferase activity was decreased in the ECA-109
and TE-10 cells overexpressing RASSF10 compared with that in the
control group; while knockdown of RASSF10 increased the relative
TOPflash/FOPflash luciferase activity, which suggested that the
Wnt/β-catenin pathway is inactivated by overexpression of RASSF10.
Western blot analysis was employed to evaluate the protein levels
of β-catenin to further verify the nuclear translocation of
β-catenin by separation of the cytoplasmic and nuclear fractions of
transfected ESCC cells. Our results indicated that RASSF10
overexpression markedly decreased β-catenin nuclear accumulation
(Fig. 6B). We then treated ECA-109
RASSF10-kncokdown cells with IWR-1 inhibitor and observed that the
levels of nuclear β-catenin were decreased, although RASSF10 levels
remained unchanged (Fig. 6C). We
also found that treatment with IWR-1 inhibited cell growth and the
migration activity of the ECA109 RASSF10-knockdown cells (Fig. 6D and E). These data demonstrated that
RASSF10 overexpression inactivated the Wnt/β-catenin signaling
pathway in ESCC.
Discussion
The mechanisms underlying esophageal squamous cell
carcinoma (ESCC) occurrence, progression and relapse are intricate.
Although the diagnosis and treatment for patients with ESCC have
made significant progress, the majority of ESCC patients usually
lose the chance of surgery due to diagnosis at advanced or
metastatic stage which leads to an unsatisfactory prognosis
(2). Metastatic outgrowths from
tumor invasion and recurrence usually lead to rapid resistance to
therapy (24,25); therefore, it is urgent and beneficial
to ESCC patients that we understand the molecular mechanisms
correlated with ESCC invasion and metastasis and confirm preventive
strategies and therapeutic targets to improve precise prognoses and
treatment of the diseases.
In the present study, we found a novel potential
biomarker, RASSF10, involved in ESCC invasion and metastasis.
RASSF10, belonging to the RASSF family, is a
tumor-suppressor gene, which has been revealed to be play crucial
roles in inhibition of hepatocellular carcinoma cell invasion and
metastasis (21). We found decreased
mRNA expression levels of RASSF10 in cancerous tissues when
compared with that noted in para-cancerous tissues of ESCC
patients, suggesting that RASSF10 functions as a tumor suppressor
in ESCC. Based on IHC results, we further confirmed that the
protein expression of RASSF10 in ESCC tissues was lower than that
in the adjacent tissues. By comparing the immunoreaction scores of
RASSF10 in ESCC tissues of various T-tumor and metastatic stages,
we found increased RASSF10 in tumors with lower scores of invasion
(T1-T2) or in non-metastatic individuals (N0), which suggest that
RASSF10 serves as a promising tumor detector. These studies support
the anticancer role of RASSF10. ESCC patients with low expression
levels of RASSF10 had a markedly worse prognosis in comparison with
that of patients with high expression of RASSF10. Moreover,
Multivariate Cox regression analysis suggested that RASSF10 is a
marked independent prognostic factor. Previous studies have shown
that low RASSF10 status contributes to an unfavorable prognosis
with malignant carcinomas such as gastric and colorectal cancer
(23,26). Our results are in keeping with those
of previous reports.
Based on the above-mentioned results, we
hypothesized that increased expression of RASSF10 may inhibit ESCC
cell proliferation, migration and invasion. Immortalization is one
of the most significant phenotypes of cancer cells, which is also
vital for ESCC progression (19).
RASSF10 might be involved in inhibition of the proliferation of
ESCC cells. A series of assays indicated that tumor cell growth and
clonogenic capacity were markedly inhibited by overexpression of
RASSF10. As determined by the CCK-8 assay, the growth of ESCC cells
was markedly inhibited by RASSF10 overexpression. The colony
formation ability in the RASSF10-transfected TE-10 and ECA-109
cells was reduced observably when compared with the control group.
Knockdown of RASSF10 reversed this phenotype. Taken together, these
results demonstrated that RASSF10 participates in the malignant
progression of ESCC via inhibition of ESCC cell proliferation,
which is in agreement with the findings of Lu et al
(27).
The major prognostic factors in ESCC patients are
largely relevant to metastasis and recurrence. Metastasis is the
most common feature of the fundamental biologic behaviors of
malignant tumor cells, which refers to the motility and migration
of primary malignant tumor cells to other parts of the body, and
spread and proliferation in a new site (24). At the molecular level, many
alterations, such as genetic and/or epigenetic alterations induced
by specific mutations or microenvironmental error signals, can act
as contributors of metastasis (28).
Cell migration and invasion are central for the process of the
metastasis of tumors. Our results indicated that the migration and
invasion abilities of the ESCC cells was suppressed by RASSF10
overexpression, suggesting that RASSF10 induced the alteration of
metastasis-related genes in the treated ESCC ECA109 and TE-10 cell
lines.
RASSF10 exerts anti-metastatic function by
obstructing epithelial-mesenchymal transition (EMT) (29). EMT is a dynamic process by which
tumor cells of epithelial origin show loss of polarity and
cell-cell adhesion and reduction in contact with the surrounding
cells and mesenchymal cells are easily transformed into spindle
fibrocyte cells with interstitial characteristics. In the meantime,
the tumor cells acquire specific biological characteristics, such
as a strong ability to migrate, the ability to infiltrate normal
surrounding tissues and to undergo distant metastasis (30,31).
Mesenchymal phenotype cells exhibit decreased expression of
epithelial-related markers such as E-cadherin, and increased
expression of mesenchymal-related markers, such as vimentin, Snail
and N-cadherin (32,33). In the present study, E-cadherin and
vimentin expression levels were assessed using the same TMA samples
to further determine the function of RASSF10 in ESCC. IHC analysis
demonstrated that tumors with lower RASSF10 expression exhibited
lower E-cadherin expression levels and higher vimentin expression
levels. It was also found that there was a positive association
between RASSF10 and E-cadherin expression and a negative
association between RASSF10 and vimentin expression. Moreover, the
protein levels of EMT-related markers were also investigated by
western blotting and immunofluorescence analysis. The results
suggested that EMT may occur in the development of ESCC, and
RASSF10 may be involved in the regulation of EMT and inhibit the
invasion and metastasis of tumor by suppressing EMT.
Based on the potentially crucial role of RASSF10 in
inhibiting ESCC metastasis, we identified the possible pathway
through which RASSF10 participates in ESCC development. Activation
of the Wnt/β-catenin signaling pathway is common in many malignant
tumors and is involved in the carcinogenesis consisting of ESCC
proliferation and migration processes (34). RASSF10 has also been proved to play a
role in blocking activation of the Wnt/β-catenin pathway to inhibit
the progression of gastric cancer (23).
Whether RASSF10 regulates the Wnt/β-catenin
signaling pathway in ESCC still remains unknown. In the present
study, the TOPflash/FOPflash luciferase assay was explored to
verify that the Wnt/β-catenin signaling pathway was inactivated by
overexpression of RASSF10, and this was also confirmed by western
blot analysis as overexpression of RASSF10 was found to decrease
nuclear accumulation of β-catenin. Conversely, compared with the
control group, sh-RASSF10-transfected cells showed increased
nuclear accumulation of β-catenin. IWR-1, a Wnt/β-catenin signaling
pathway inhibitor, was employed to confirm this possibility. Our
results indicated that the protein levels of nuclear β-catenin were
decreased, followed by inhibition of ESCC cell proliferative and
migrative capability, and it was also found that RASSF10 levels
were unchanged after treatment with IWR-1. Our results suggest that
inactivation of Wnt/β-catenin may be responsible for RASSF10
overexpression-mediated inhibitory effects on ESCC progression.
In conclusion, our results elucidated that the
tumor-suppressive role of RASSF10 is negatively correlated with
cell proliferation, metastasis and the EMT process in ESCC. These
findings help us to understand the inevitable and valuable role of
RASSF10 in the progression and development of ESCC, suggesting that
RASSF10 may serve as an independent candidate for a novel
therapeutic target in the treatment of ESCC.
Acknowledgements
Not applicable.
Funding
The present study was funded by grants from the Six Talent Peaks
Project in Jiangsu Province, China (no. WSN-059), the Science
Foundation of Nantong City, Jiangsu, China (no. MS22016032 and
MS12017008-5), the Scientific Research Topic of Jiangsu Provincial
Health and Family Planning Commission, China (no. H201626), the Key
Talents of Medical Science in Jiangsu Province, China (no.
QNRC2016682), the Nantong Science and Technology Program (no.
GJZ17050), the National Natural Science Foundation of China
(81770266), ‘Six-one’ Project for High-level Health Talents
(LGY2016037), and the Nantong Key Laboratory of Translational
Medicine in Cardiothoracic Diseases in Jiangsu, China.
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
YL, JS and JF conceived and designed the
experiments. XZ, WZ, HQ and TB carried out the experiments. ZW, JZ
and YH collected the clinical samples and corresponding clinical
data and confirmed their accuracy. YL, JF and WZ revised the
manuscript. YL wrote this manuscript. All authors read and approved
the final manuscript for publication.
Ethics approval and consent to
participate
This project was authorized by permission of the
Ethics Committee of the Affiliated Hospital of Nantong University
(Nantong, China) (2014-025, March 5, 2014), and signed
informed-consent forms were collected from all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Cowan AJ, Allen C, Barac A, Basaleem H,
Bensenor I, Curado MP, Foreman K, Gupta R, Harvey J, Hosgood HD, et
al: Global burden of multiple myeloma: A systematic analysis for
the global burden of disease study 2016. JAMA Oncol. 4:1221–1227.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Alsop BR and Sharma P: Esophageal cancer.
Gastroenterol Clin North Am. 45:399–412. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lou F, Sima CS, Adusumilli PS, Bains MS,
Sarkaria IS, Rusch VW and Rizk NP: Esophageal cancer recurrence
patterns and implications for surveillance. J Thorac Oncol.
8:1558–1562. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tran GD, Sun XD, Abnet CC, Fan JH, Dawsey
SM, Dong ZW, Mark SD, Qiao YL and Taylor PR: Prospective study of
risk factors for esophageal and gastric cancers in the Linxian
general population trial cohort in China. Int J Cancer.
113:456–463. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Arnold M, Soerjomataram I, Ferlay J and
Forman D: Global incidence of oesophageal cancer by histological
subtype in 2012. Gut. 64:381–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zou S, Yang J, Guo J, Su Y, He C, Wu J, Yu
L, Ding WQ and Zhou J: RAD18 promotes the migration and invasion of
esophageal squamous cell cancer via the JNK-MMPs pathway. Cancer
Lett. 417:65–74. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Richter AM, Pfeifer GP and Dammann RH: The
RASSF proteins in cancer; From epigenetic silencing to functional
characterization. Biochim Biophys Acta. 1796:114–128.
2009.PubMed/NCBI
|
|
8
|
Sherwood V, Recino A, Jeffries A, Ward A
and Chalmers AD: The N-terminal RASSF family: A new group of
Ras-association-domain-containing proteins, with emerging links to
cancer formation. Biochem J. 425:303–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Underhill-Day N, Hill V and Latif F:
N-terminal RASSF family: RASSF7-RASSF10. Epigenetics. 6:284–292.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Global Burden of Disease Cancer
Collaboration, . Fitzmaurice C, Allen C, Barber RM, Barregard L,
Bhutta ZA, Brenner H, Dicker DJ, Chimed-Orchir O, Dandona R, et al:
Global, regional, and national cancer incidence, mortality, years
of life lost, years lived with disability, and disability-adjusted
life-years for 32 cancer groups, 1990 to 2015: A systematic
analysis for the global burden of disease study. JAMA Oncol.
3:524–548. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Helmbold P, Richter AM, Walesch S,
Skorokhod A, Marsch WCh, Enk A and Dammann RH: RASSF10 promoter
hypermethylation is frequent in malignant melanoma of the skin but
uncommon in nevus cell nevi. J Invest Dermatol. 132(3 Pt 1):
687–694. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hesson LB, Dunwell TL, Cooper WN,
Catchpoole D, Brini AT, Chiaramonte R, Griffiths M, Chalmers AD,
Maher ER and Latif F: The novel RASSF6 and RASSF10 candidate tumour
suppressor genes are frequently epigenetically inactivated in
childhood leukaemias. Mol Cancer. 8:422009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hill VK, Underhill-Day N, Krex D, Robel K,
Sangan CB, Summersgill HR, Morris M, Gentle D, Chalmers AD, Maher
ER and Latif F: Epigenetic inactivation of the RASSF10 candidate
tumor suppressor gene is a frequent and an early event in
gliomagenesis. Oncogene. 30:978–989. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Schagdarsurengin U, Richter AM, Wöhler C
and Dammann RH: Frequent epigenetic inactivation of RASSF10 in
thyroid cancer. Epigenetics. 4:571–576. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu X, Yu AX, Qi BW, Fu T, Wu G, Zhou M,
Luo J and Xu JH: The expression and significance of IDH1 and p53 in
osteosarcoma. J Exp Clin Cancer Res. 29:432010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kimura M: A simple method for estimating
evolutionary rates of base substitutions through comparative
studies of nucleotide sequences. J Mol Evol. 16:111–120. 1980.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Q, Yao YT, Xu H, Chen YB, Gu M, Cai
ZK and Wang Z: SPOCK1 promotes tumor growth and metastasis in human
prostate cancer. Drug Des Devel Ther. 10:2311–2321. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheung CT, Singh R, Kalra RS, Kaul SC and
Wadhwa R: Collaborator of ARF (CARF) regulates proliferative fate
of human cells by dose-dependent regulation of DNA damage
signaling. J Biol Chem. 289:18258–18269. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang J, Zhi X, Shi S, Tao R, Chen P, Sun
S, Bian L, Xu Z and Ma L: SPOCK1 is up-regulated and promotes tumor
growth via the PI3K/AKT signaling pathway in colorectal cancer.
Biochem Biophys Res Commun. 482:870–876. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao Y, Yi J, Zhang K, Bai F, Feng B, Wang
R, Chu X, Chen L and Song H: Downregulation of MiR-31 stimulates
expression of LATS2 via the hippo pathway and promotes
epithelial-mesenchymal transition in esophageal squamous cell
carcinoma. J Exp Clin Cancer Res. 36:1612017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang F, Feng Y, Li P, Wang K, Feng L, Liu
YF, Huang H, Guo YB, Mao QS and Xue WJ: RASSF10 is an
epigenetically inactivated tumor suppressor and independent
prognostic factor in hepatocellular carcinoma. Oncotarget.
7:4279–4297. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhu R, Liu Y, Zhou H, Li L, Li Y, Ding F,
Cao X and Liu Z: Deubiquitinating enzyme PSMD14 promotes tumor
metastasis through stabilizing SNAIL in human esophageal squamous
cell carcinoma. Cancer Lett. 418:125–134. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei Z, Chen X, Chen J, Wang W, Xu X and
Cai Q: RASSF10 is epigenetically silenced and functions as a tumor
suppressor in gastric cancer. Biochem Biophys Res Commun.
432:632–637. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tarapore RS, Yang Y and Katz JP: Restoring
KLF5 in esophageal squamous cell cancer cells activates the JNK
pathway leading to apoptosis and reduced cell survival. Neoplasia.
15:472–480. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang F, Li P, Feng Y, Hu YL, Liu YF, Guo
YB, Jiang XL, Mao QS and Xue WJ: Low expression of RASSF10 is
associated with poor survival in patients with colorectal cancer.
Hum Pathol. 62:108–114. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lu D, Ma J, Zhan Q, Li Y, Qin J and Guo M:
Epigenetic silencing of RASSF10 promotes tumor growth in esophageal
squamous cell carcinoma. Discov Med. 17:169–178. 2014.PubMed/NCBI
|
|
28
|
Chaffer CL and Weinberg RA: A perspective
on cancer cell metastasis. Science. 331:1559–1564. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan C, Wang W, Jin J, Yu Z and Xin X:
RASSF10 is epigenetically inactivated and suppresses cell
proliferation and induces cell apoptosis by activating the p53
signalling pathway in papillary thyroid carcinoma cancer. Cell
Physiol Biochem. 41:1229–1239. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Figiel S, Vasseur C, Bruyere F, Rozet F,
Maheo K and Fromont G: Clinical significance of
epithelial-mesenchymal transition markers in prostate cancer. Hum
Pathol. 61:26–32. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Fukuda K, Takeuchi S, Arai S, Katayama R,
Nanjo S, Tanimoto A, Nishiyama A, Nakagawa T, Taniguchi H, Suzuki
T, et al: Epithelial-to-mesenchymal transition is a mechanism of
ALK inhibitor resistance in lung cancer independent of ALK mutation
status. Cancer Res. 79:1658–1670. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Das V, Bhattacharya S, Chikkaputtaiah C,
Hazra S and Pal M: The basics of epithelial-mesenchymal transition
(EMT): A study from a structure, dynamics, and functional
perspective. J Cell Physiol. Feb 5–2019.(Epub ahead of print).
View Article : Google Scholar
|
|
33
|
Zhang C and Wang Y: Metformin attenuates
cells stemness and epithelial-mesenchymal transition in colorectal
cancer cells by inhibiting the Wnt3a/β-catenin pathway. Mol Med
Rep. 19:1203–1209. 2019.PubMed/NCBI
|
|
34
|
He J, Zhou M, Chen X, Yue D, Yang L, Qin
G, Zhang Z, Gao Q, Wang D, Zhang C, et al: Inhibition of SALL4
reduces tumorigenicity involving epithelial-mesenchymal transition
via Wnt/β-catenin pathway in esophageal squamous cell carcinoma. J
Exp Clin Cancer Res. 35:982016. View Article : Google Scholar : PubMed/NCBI
|