Introduction
In 1971, Folkman (1) first proposed that tumor growth and
metastasis were dependent on tumor angiogenesis to a certain
extent, which is the theory of tumor angiogenesis. Angiogenesis
provides necessary oxygen and nutrients for tumor growth, and
blocking angiogenesis has become a tumor treatment approach. Recent
research has indicated that circular RNAs (circRNAs) are a noel
class of RNAs with special covalently closed loop structures
without 5′ caps and 3′ tails that can regulate angiogenesis-related
diseases through multiple mechanisms (2). circRNAs are abnormally expressed in
almost all tumor types, play a crucial role in cancer pathogenesis
as oncogenes or tumor suppressors (3), and can participate in the
proliferation, migration, invasion and apoptosis of tumor cells
through various mechanisms (4,5). As
opposed to traditional RNAs, circRNAs are more stable and conserved
in vivo due to their structural specificity and can resist
exoribonuclease RNase R-mediated degradation (6). Due to these characteristics, circRNAs
are potential clinical diagnostic markers.
The present review summarizes and discusses the
biological properties and functions of circRNAs and the related
mechanisms of tumor angiogenesis, with specific focus on the
mechanisms and roles of circRNAs in tumor angiogenesis in various
systems. In addition, the present review elaborates on the clinical
applications of circRNAs in tumor angiogenesis and their future
prospects as novel diagnostic markers and therapeutic targets in
clinical tumor angiogenesis.
Tumor angiogenesis
Angiogenesis is the process through which
endothelial cells proliferate, differentiate and migrate to form
microvessels from existing blood vessels (7). Under physiological conditions, a
variety of angiogenic factors promote or inhibit angiogenesis to
maintain the stability of the body's internal environment. Cell
proliferation, migration and vascular endothelial growth factor
(VEGF) secretion are critical for angiogenesis, and an imbalance in
these processes can lead to the development of diseases (8). The distinguishing features of tumors
are active proliferation and metabolism, and both tumor growth and
metastasis are dependent on angiogenesis. Angiogenesis is the key
to tumor development, as this process stimulates the vascular
system to grow new blood vessels by activating the ‘angiogenic
switch’ in tumors (9,10), which provides sufficient oxygen and
nutrients to promote tumor growth and proliferation (11).
With the further development of the theory of tumor
angiogenesis, studies have reported the significance of angiogenic
factors and their roles in early tumor growth, invasion and
metastasis (12–14). Angiogenic factors are divided into
pro-angiogenic and anti-angiogenic factors. Anti-angiogenic factors
include angiostatin, endostatin and thrombospondin. Pro-angiogenic
factors mainly include VEGF (15,16),
angiopoietin (17), matrix
metalloproteinases (MMPs) (18,19)
and fibroblast growth factor (FGF) (20), of which VEGF is a key angiogenic
mediator that drives continuous tumor growth (21,22).
VEGF is a family of homodimeric glycoproteins with a molecular
weight of ~45 kDa and primarily includes VEGFA, VEGFB, VEGFC, VEGFD
and placental growth factor (PIGF). The central role of VEGF in
tumorigenesis and the spread of tumor cells to other parts of the
body indicates that the inhibition of tumor angiogenesis may be an
effective strategy and target for tumor therapy.
Biological functions of circRNAs
circRNAs, as a novel type of RNA molecule, have been
found to have potential biological functions, which indicates that
some circRNAs may be involved in gene expression regulation as
transcriptional or splicing regulators (23,24).
The biological basis of tumorigenesis is the abnormal expression of
genes, and the structural specificity and abundant biological
functions of circRNAs is indicative of their critical role in
tumorigenesis and tumor progression.
It has been reported (25) that a number of different circRNAs
are expressed in eukaryotic cells at levels ~10-fold those of
linear RNAs, which suggests that circRNAs may have potential
biological functions. The present review discusses the related
biological functions of circRNAs. circRNAs can function as microRNA
(miRNA/miR) sponges, which was the earliest discovered function of
circRNAs (26) and has become a
research hotspot. Some circRNAs contain several miRNA-binding sites
that can compete with mRNAs and bind to miRNAs to sponge miRNAs and
regulate the expression of target genes (27). circRNAs can interact with
RNA-binding proteins (RBPs) to indirectly regulate gene
transcription. For example, circRNAs can compete with mRNAs to bind
to RBPs, and thus, they can affect the translation of mRNAs.
circRNAs can function as RBP super sponges to affect RBP expression
and they interact with specific proteins and enhance their
functions, and they can also regulate target gene translation
(28). circRNAs can also function
in protein translation. Of note, two mechanisms of translation have
been established: One is dependent on the internal ribosome entry
site, while the other is dependent on N6-methyladenosine
methylation modification. The post-translation products of circRNAs
can participate in multiple physiological processes in the body,
such as preventing linear translation products from being degraded
by ubiquitin proteases (29),
regulating the PI3K/AKT pathway (30) and inhibiting the transcriptional
elongation of proto-oncogenes (31).
Previous research has found that circRNAs are stable
and conserved in vivo due to their structural specificity,
and consequently, they have the potential to serve as molecular
markers (32). circRNAs have been
confirmed to be closely related to aging, cancer and other chronic
diseases, and can serve as markers for disease diagnosis and
detection. Experiments using Drosophila have demonstrated
that circRNAs are markers of aging and that circRNAs present in
human saliva are also potential disease markers (32,33).
Numerous circRNAs also exist in exosomes and can be used as
molecular diagnostic markers (34). VEGF is a key factor in the process
of tumor angiogenesis, and circRNAs can directly or indirectly
target VEGF to regulate this process.
Related mechanisms through which circRNAs
regulate tumor angiogenesis
circRNAs are involved in a variety of physiological
and pathological processes, including tumor proliferation, invasion
and metastasis (35). Previous
studies have demonstrated that circRNAs participate in tumor
angiogenesis primarily by functioning as targeting sponges for
miRNAs, either by binding to miRNAs, which blocks their expression,
or with competing endogenous RNAs (ceRNAs) (26,36,37).
miRNAs are linear non-coding RNAs that can adsorb mRNAs, promote
their degradation and thus regulate tumor angiogenesis (38). Due to their structural specificity
and diverse biological functions, circRNAs play a critical role in
tumor angiogenesis by directly or indirectly targeting VEGF or
other angiogenic factors.
Involvement of circRNAs in tumor
angiogenesis as ceRNAs
miRNAs can silence gene expression by binding to
mRNAs, and circRNAs can regulate gene expression by competitively
binding with miRNAs. ceRNAs are closely related to tumor
angiogenesis (39) and play a
crucial role in this process.
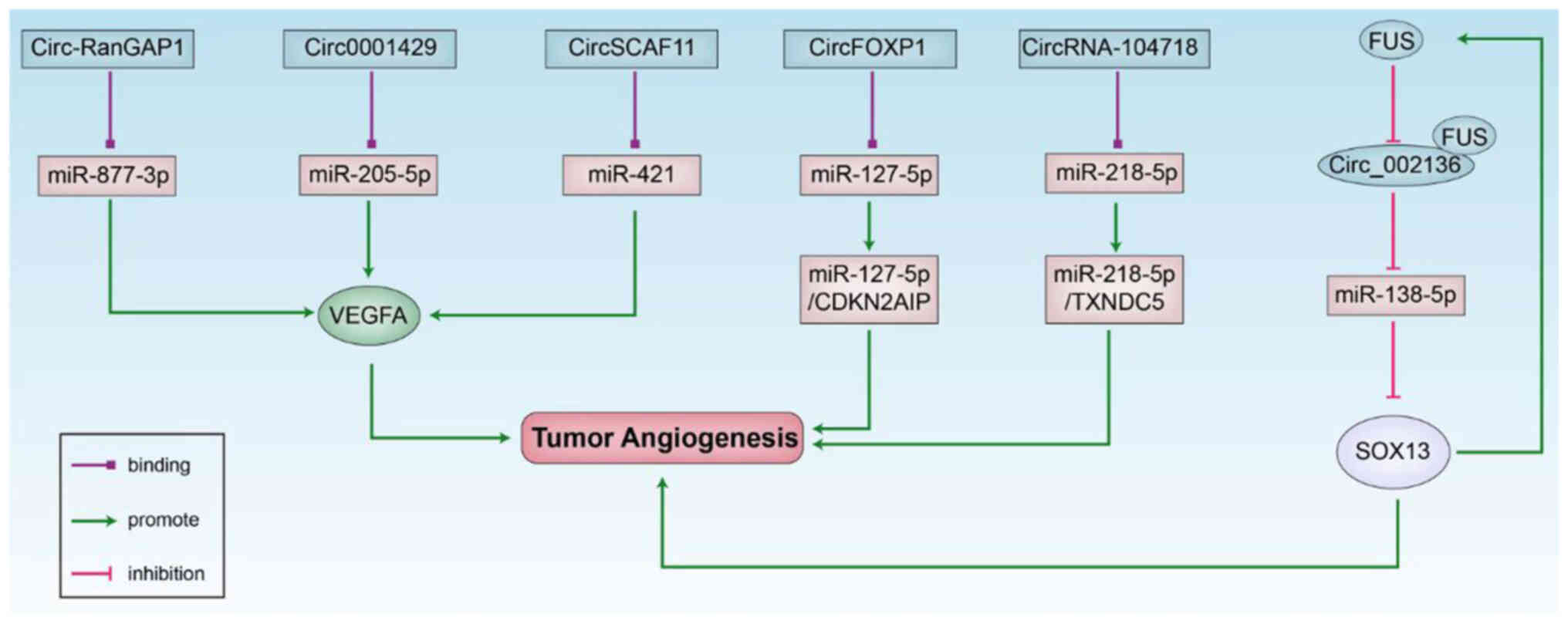
Circ-RanGAP1 can function as a ceRNA to inhibit the
activity of miR-877-3p and increase the expression of the target
gene, VEGFA (40), while
circ0001429 can target miR-205-5p and regulate VEGFA to promote
bladder cancer (41). CircSCAF11
regulates the transcription factor, SP1, through miR-421 and
activates VEGFA transcription, which accelerates glioma development
through the miR-421/SP1/VEGFA axis (42). Circ-ZNF609 downregulates miR-145
expression and upregulates stathmin 1 expression by acting as a
ceRNA to promote cell proliferation, migration and angiogenesis in
nasopharyngeal carcinoma (43),
while circRNA-104718 functions as a ceRNA and directly binds to
miR-218-5p and promotes the progression of liver cancer by
targeting the miR-218-5p/thioredoxin domain containing 5 signaling
pathway (44). CircFOXO1 is
upregulated in osteosarcoma to promote the growth of osteosarcoma
and can directly combine with miR-127-5p to promote the expression
of CDKN2AIP and promote angiogenesis by regulating the
miR-127-5p/CDKN2AIP signaling pathway (45). The fusion sarcoma (FUS) gene is a
DNA/RNA regulatory protein with a gene regulatory function. The FUS
gene is involved in the processes of intracellular RNA
transduction, mRNA synthesis, alternative splicing and
polyadenylation site selection (46). High FUS gene expression in glioma
can work synergistically with Circ_002136. Circ_002136 directly
targets miR-138-5P to inhibit the expression of the downstream
target gene SOX3 and to promote glioma-exposed endothelial cell
(GEC) angiogenesis; the knockout of circ_002136 and the
overexpression of miR-138-5P can reduce the angiogenic capacity of
gliomas (47). On the whole, the
ceRNA mechanism is a typical regulatory mechanism among non-coding
RNAs, which supports the further study of circRNAs in tumor
angiogenesis (Fig. 1).
VEGF-dependent regulation of tumor
angiogenesis
VEGF commonly refers to VEGFA, whereas the VEGF
family also includes VEGFB, VEGFC, VEGFD and PIGF. The majority of
cells in the body can produce VEGFA, which is upregulated under
hypoxic conditions (48). VEGF is
produced by tumor cells, endothelial cells and tumor-associated
macrophages under hypoxic conditions during tumor growth. It has
been found that circRNAs can affect tumor angiogenesis by
modulating VEGF, which further affects the process of tumor growth,
proliferation and metastasis (41).
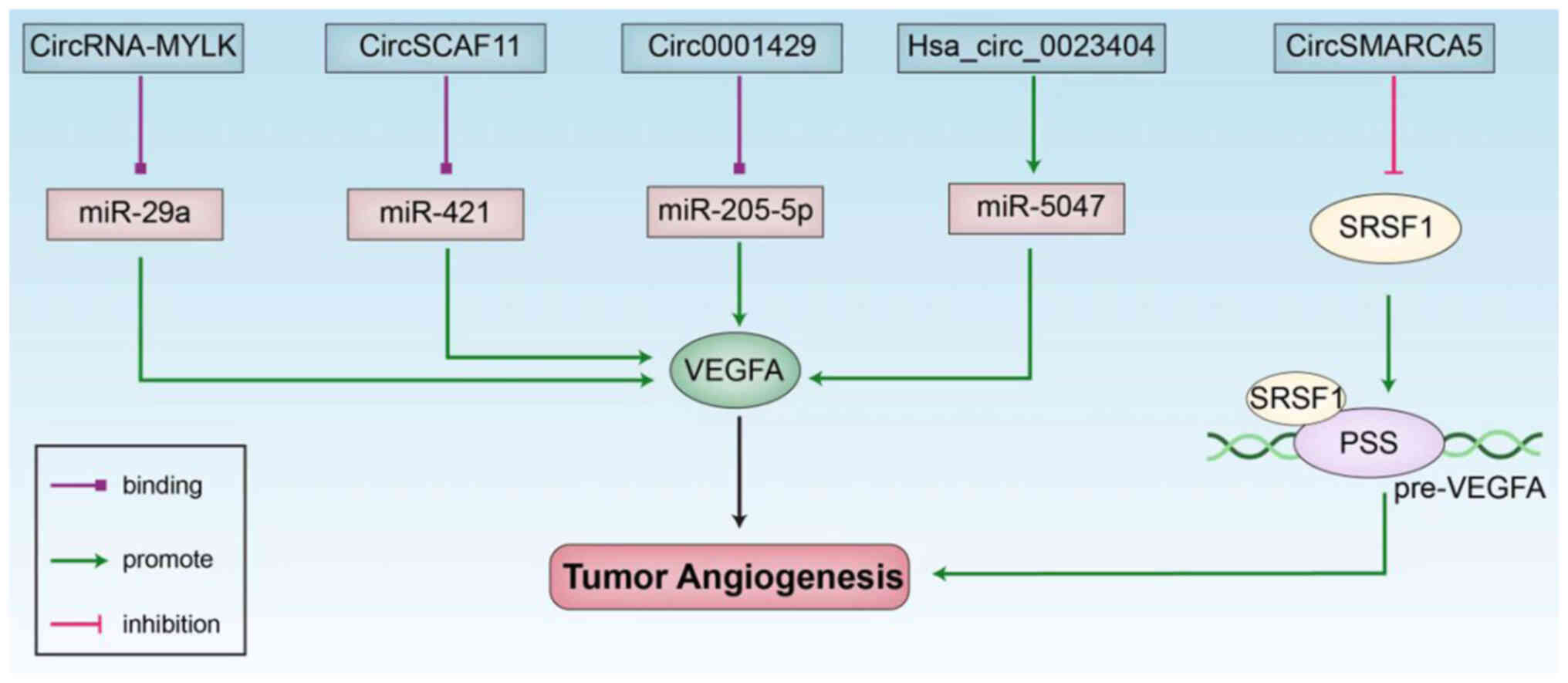
Circ-RanGAP1 can induce VEGFA expression through
miR-877-3p and enhance the invasion and metastasis of gastric
cancer (40), while
hsa_circ_0023404 directly binds to miR-5047 and regulates the
expression of the VEGFA gene (49), which enhances metastasis and
chemotherapeutic resistance in cervical cancer. CircFndc3b is
downregulated in the mouse heart following myocardial infarction,
can interact with fused RNAs in sarcoma and regulate cardiac repair
following myocardial infarction through the FUS/VEGFA signaling
axis; the regulation of circFndc3b expression has emerged as a
potential therapeutic strategy for cardiac functional remodeling
following myocardial infarction (50). Circ0001429 increases VEGFA
expression via miR-205-5p, accelerates the proliferation, invasion
and migration of bladder cancer cells, and promotes bladder cancer
progression (41). CircRNA-MYLK is
upregulated in bladder cancer and is related to clinical stage;
this circRNA can also bind to miR-29a and indirectly regulate the
expression of VEGFA, and promote the angiogenesis and metastasis of
bladder cancer (51). CircSCAF11
can bind to miR-421 to regulate the transcription factor SP1,
activate VEGFA transcription (42), and accelerate glioma development.
The expression of hsa_circ_0000096 is downregulated in gastric
cancer, and the knockout of hsa_circ_0000096 downregulates VEGFA
expression, inhibits cell proliferation and migration and further
affects tumor angiogenesis (52)
(Fig. 2).
CircRNAs not only function as ceRNAs to regulate
VEGFA to affect tumor angiogenesis, but also participate in tumor
angiogenesis by influencing the VEGF splicing process. CircSMARCA5
can regulate the splicing and angiogenesis of VEGFA mRNA in
glioblastoma multiforme (GBM) by binding to serine and
arginine-rich splicing factor 1 (SRSF1) and is therefore a
promising therapeutic target in GBM cells (53).
VEGF-independent regulation of tumor
angiogenesis
In addition to the VEGF family, angiotensin,
platelet-derived growth factor (PDGF-β), transforming growth factor
(TGF-β) and MMPs (18,19) are also closely associated with
tumor angiogenesis (54).
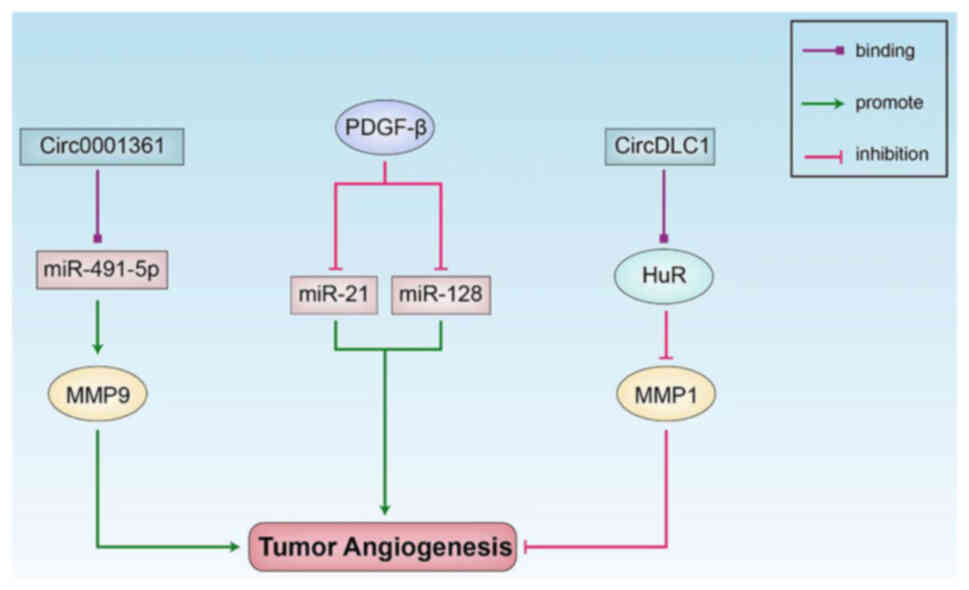
PDGF-β is an effective angiogenic factor involved in
GBM that can downregulate the tumor-promoting gene, miR-21, and the
tumor suppressor gene, miR-128, to regulate the expression of
related factors and promote tumor cell proliferation (55). It has been found that PDGF-β is
systemically upregulated in patients with age-related macular
degeneration (AMD) and in patients with diabetic macular edema
compared with healthy controls; moreover, PDGF-β may be involved in
the pathogenesis of neovascular AMD (56). In bladder cancer, circ0001361
interacts with miR-491-5p, which upregulates MMP9 expression and
promotes the migration and invasiveness of bladder cancer cells,
and thus, circ0001361 is expected to become a novel therapeutic
target for bladder cancer (57).
circDLC1 interacts with the RNA-binding protein, HuR, and inhibits
MMP1 expression by reducing the stability of MMP1 mRNA, inhibiting
the proliferation and metastasis of liver cancer cells and
attenuating the progression of liver cancer, which suggests that
circDLC1 is a promising diagnostic biomarker and potential
therapeutic target for liver cancer (58) (Fig.
3).
Effects of circRNAs on tumor angiogenesis in
various systems
Due to their structural stability and rich
biological functions, circRNAs play a criticial role in
tumorigenesis by affecting tumor growth, proliferation and
metastasis. Studies have found that circRNAs are also widely
expressed in the digestive system (40,59),
urinary system (51,60), nervous system tumors and other
related tumors (61,62), and exert marked effects on tumor
angiogenesis and development in various systems.
circRNAs and gastrointestinal tumor
angiogenesis
circRNAs are widely expressed in gastrointestinal
tumors and can affect the progression of these tumors through
angiogenesis (Table I).
Circ-RanGAP1 is upregulated in the tissues and peripheral blood of
patients with gastric cancer, where it promotes the invasion and
metastasis of gastric cancer by targeting miR-877-3p to regulate
the expression of VEGFA; moreover, the inhibition of circ-RanGAP1
can reduce the invasion and migration of gastric cancer cells
(40). Circ29 is upregulated in
gastric cancer, binds miR-29a as a ceRNA, and plays a role in
gastric cancer angiogenesis by regulating the VEGF pathway
(63). In pancreatic ductal
adenocarcinoma, circNFIB1 attenuates the carcinogenesis of
miR-486-5p and upregulates PIK3R1 expression, which further
downregulates VEGFC expression by inhibiting the PI3K/Akt pathway;
this in turn inhibits lymphangiogenesis and metastasis in
pancreatic ductal adenocarcinoma (64). CircCCT3 functions as a miR-613
sponge to promote the migration and invasiveness of pancreatic
cancer (PC) cells by regulating the VEGF/VEGFR2 signaling pathway
(59). CircADAM9 promotes PC
progression by acting as a sponge of miR-217 to upregulate serine
protease 3 expression (65). In
colorectal cancer, circ_0056618 binds to miR-206 to upregulate the
expression of CXCR4 and VEGFA, which promotes the proliferation,
migration and angiogenesis of tumor cells (66). CircRNA UBAP2 is highly expressed in
colorectal cancer tissues and cells, and promotes the further
development of colorectal cancer via the upregulation of VEGFA
expression by promoting the secretion of miR-199a (67). It has been found that the
downregulation of circSMARCC1 expression can reduce the survival of
colorectal cancer cells and inhibit the protein expression of MMP2,
MMP9 and VEGF; moreover, the binding of circSMARCC1 to miR-140-3p
can regulate the progression of colorectal cancer (68). Circ-001971 functions as a ceRNA to
reduce miR-29c-3p-induced VEGFA upregulation, and promote
colorectal cancer proliferation, invasion and angiogenesis
(69). WNT3 and VEGFA are
downstream targets of miR-613 in colorectal cancer cells, and
circCCT3 acts as a miR-61 sponge, which regulates the VEGFA and Wnt
signaling pathways to enhance colorectal cancer metastasis
(70).
 | Table I.circRNAs and gastrointestinal tumor
angiogenesis. |
Table I.
circRNAs and gastrointestinal tumor
angiogenesis.
| Origin | circRNA | Mechanism | Functions | (Refs.) |
|---|
| Gastric cancer | Circ-RanGAP1 |
miR-877-3p/VEGFA | Invasion and
metastasis | (40) |
| Gastric cancer | Circ29 | miR-29a/VEGF | Proliferation,
migration, tube formation of HUVECs | (63) |
| Pancreatic ductal
adenocarcinoma | CircNFIB1 |
miR-486-5p/PIK3R1/VEGFC | Inhibits
lymphangiogenesis and LN metastasis in pancreatic ductal
adenocarcinoma | (64) |
| Pancreatic
cancer | CircCCT3 |
miR-613/VEGFA/VEGFR2 | Promotes the
migration and invasion of pancreatic cancer | (59) |
| Pancreatic
cancer | CircADAM9 | miR-217/PRSS3 | Inhibits the
proliferation, migration and invasiveness of pancreatic cancer
cells | (65) |
| Colorectal
cancer | Circ_0056618 |
miR-206/CXCR4/VEGFA | Inhibits cell
proliferation, migration and angiogenesis | (66) |
| Colorectal
cancer | CircRNA UBAP2 | miR-199a/VEGFA | Suppresses cell
proliferation, migration and invasion | (67) |
| Colorectal
cancer | Circ-SMARCC1 |
miR-140-3p/MMPs | Inhibits cell
viability, migration, and invasion | (68) |
| Colorectal
cancer | Circ-001971 |
miR-29c-3p/VEGFA | Suppresses
proliferation, invasion and angiogenesis | (69) |
| Colorectal
cancer | CircCCT3 |
miR-613/VEGFA/Wnt | Promotes migration
and invasion, and inhibits cell apoptosis | (70) |
| Esophageal squamous
cell carcinoma | Circ_0072088 | miR-377/VEGF | Promotes
proliferation, migration, and invasion | (71) |
| Hepatocellular
carcinoma | Circ_0001178 | miR-382/VEGFA | Reduces cell
viability and suppresses migration and invasion | (72) |
In esophageal squamous cell carcinoma (ESCC),
circ_0072088 can function as a miRNA-377 sponge to upregulate VEGF
gene expression and promote the proliferation, migration and
angiogenesis of ESCC cells. Circ_0072088 is expected to become a
novel diagnostic biomarker and therapeutic target for ESCC
(71). Circ_0001178 is upregulated
in hepatocellular carcinoma (HCC) and can directly bind to miR-382.
VEGFA is a downstream target of miR-382, and it has been reported
that circ_0001178 promotes liver cancer progression by regulating
miR-382 and VEGFA; the knockdown of circ_0001178 inhibits HCC
growth in vivo (72).
circRNAs and tumor angiogenesis in the
urinary system
Common urinary system cancers include bladder
cancer, prostate cancer and renal cell carcinoma; however, research
on the role of circRNAs in angiogenesis in urinary system tumors is
mostly limited to bladder cancer. CircRNAs targeting oncogenic
pathways in urinary system tumors mainly include TGF-β, AKT, VEGF
and other pathways. It has been found that circRNAs can target
related oncogenic molecules or pathways to regulate tumor
angiogenesis in the urinary system (Table II).
| Table II.circRNAs and tumor angiogenesis in
the urinary system. |
Table II.
circRNAs and tumor angiogenesis in
the urinary system.
| Origin | circRNA | Mechanism | Functions | (Refs.) |
|---|
| Bladder cancer | CircRNA-MYLK |
miR29a/VEGFA/VEGFR2/Ras/ERK | Suppresses bladder
cancer cell proliferation and invasion | (51) |
| Bladder cancer | CircEHBP1 |
miR-130a-3p/TGF-β/SMAD3/VEGFD | Promotes
lymphangiogenesis and LN metastasis | (60) |
| Bladder cancer | Circ0001429 |
miR-205-5p/VEGFA | Promotes cell
proliferation, migration and invasiveness | (41) |
The expression of circRNA-MYLK is increased in
bladder cancer, and it can directly bind to miR-29a to promote the
expression of the target VEGFA by reducing the activity of miR-29a;
circRNA-MYLK also promotes the proliferation and
epithelial-mesenchymal transition of bladder cancer cells by
activating the VEGFA/VEGFR2 signaling pathway. CircRNA-MYLK
regulates the expression of VEGFA by binding to miR-29a (51). CircEHBP1 is significantly
upregulated in bladder cancer, where it binds to miR-130a-3p and
antagonizes the inhibitory effects of miR-130a-3p on the 3′-UTR of
TGFBR1. The upregulation of TGFβR1 expression further activates the
TGF-β/SMAD3 signaling pathway to induce VEGFD expression, thereby
promoting lymphangiogenesis and lymphatic metastasis in bladder
cancer (60). Circ0001429
expression has been found to be upregulated in bladder cancer
tissues, and in vitro experiments have demonstrated that
circ0001429 enhances the proliferation and metastasis of T24 and
5637 cells, and inhibits apoptosis (41). The results of these experiments
have revealed that circ0001429 may increase VEGFA expression
through miR-205-5p, promote the proliferation and metastasis of
cancer cells and thus regulate the progression of bladder cancer
(41).
circRNAs and angiogenesis in
gynecological tumors
Among the gynecological tumors, cervical, ovarian
and breast cancer are associated with high morbidity and mortality
rates, and pose a substantial threat to the lives and health of
women worldwide. circRNAs can bind to miRNAs through miRNA response
elements, and can play a crucial role in tumor development and
angiogenesis as tumor suppressor genes or proto-oncogenes (26). circRNAs are closely related to the
development, metastasis and prognosis of gynecological tumors
(Table III).
| Table III.circRNAs and angiogenesis in
gynecological tumors. |
Table III.
circRNAs and angiogenesis in
gynecological tumors.
| Origin | CircRNA | Mechanism | Functions | (Refs.) |
|---|
| Ovarian cancer | CircASH2L | miR-665/VEGFA | Enhances invasion,
proliferation, angiogenesis, and lymphangiogenesis | (73) |
| Ovarian cancer | CircRhoC | miR-302e/VEGFA | Enhances migration
and invasion | (74) |
| Ovarian cancer | Circ-CSPP1 |
miR-1236-3p/MMP-2/VEGFA | Promotes
proliferation, invasion and migration | (75) |
| Cervical
cancer |
hsa_circ_0023404 | miR-5047/VEGFA | Enhances lymphatic
vessel formation and promotes metastasis | (49) |
| Breast cancer |
MicroRNA-140-5p |
miR-140-5p/VEGFA | Suppresses breast
cancer metastasis and angiogenesis | (76) |
CircASH2L is highly expressed in ovarian cancer
tissues and cell lines, and functions as a ceRNA to regulate the
miR-665/VEGFA axis, which promotes VEGFA-mediated angiogenesis and
lymphangiogenesis, and enhances the proliferation and invasion of
ovarian cancer cells (73). As a
highly expressed oncogene in ovarian cancer, circRhoC can not only
act function a miR-302e sponge to regulate VEGFA protein
expression, but it can also directly bind to and regulate VEGFA,
which promotes tumorigenicity and ovarian cancer progression
(74). Circ-CSPP1 silencing can
lead to the decreased expression of MMP2 and VEGFA, and may promote
the proliferation, invasion and migration of ovarian cancer cells
by acting as a sponge of miR-1236-3p, which provides insight into
the early diagnosis and treatment of ovarian cancer (75). In cervical cancer, hsa_circ_0023404
directly interacts with miR-5047 to regulate VEGFA gene expression
and promotes cervical cancer metastasis through the miR-5047/VEGFA
and autophagy signaling pathways (49). As a tumor suppressor downregulated
in primary breast cancer tissues, miR-140-5p can inhibit the
invasiveness and angiogenesis of breast cancer cells in
vitro and in vivo by targeting VEGFA (76).
circRNAs and tumor angiogenesis in the
nervous system
Central nervous system (CNS) tumors are generally
divided into two categories: Primary tumors originating in the CNS
and secondary tumors that metastasize from other parts of the body.
CNS tumors are associated with a high morbidity and mortality
worldwide, and include GBM, medulloblastoma and oligodendroglioma.
Among the tumors of the CNS, the regulation of circRNA expression
in tumor angiogenesis is mainly observed in glioblastoma and other
gliomas.
Circ-RPL15 and VEGFA are upregulated in glioma
tissues. Circ-RPL15 can upregulate VEGFA expression by
competitively binding to miR-146b-3p, enhancing the proliferation
and migration of glioma cells, and promoting glioma progression
(77). In gliomas, circITGA7
regulates glioma proliferation and metastasis by acting as a
miR-34a-5p sponge that targets VEGFA and regulates the
miR-34a-5p/VEGFA pathway, which provides a potential target for the
early diagnosis and treatment of gliomas (61). GAUGAA, as an RNA motif, regulates
the interaction between circSMARCA5 and SRSF1 and the
circSMARCA5-mediated control of GBM cell migration and angiogenic
potential (78). CircSMARCA5 is an
upstream regulator of the ratio of proangiogenic to antiangiogenic
VEGFA isoforms in GBM cells. This circRNA also regulates VEGFA mRNA
splicing and angiogenesis in GBM by binding to SRSF1 and is
expected to become an emerging marker and effective therapeutic
target for GBM (53).
circRNAs and other related angiogenic
tumors
Due to their diverse biological functions and
specific structures, circRNAs are widely expressed in angiogenic
tumors and other related diseases in various systems. In addition
to intensive research in the gastrointestinal tract, urinary
system, CNS and gynecological tumors, circRNAs are also expressed
in diseases, such as osteosarcoma (OS), infantile hemangioma (IH),
thyroid cancer and breast cancer (Table IV).
| Table IV.circRNAs and other related angiogenic
tumors. |
Table IV.
circRNAs and other related angiogenic
tumors.
| Origin | CircRNA | Mechanism | Functions | (Refs.) |
|---|
| Osteosarcoma | Circ_001621 | miR-578/VEGF | Promotes
proliferation and migration | (62) |
| Thyroid cancer | CircPVT1 |
miR-195/Wnt/β-catenin/VEGFA | Inhibits
proliferation, migration, and invasion | (80) |
| Non-small cell lung
cancer | CircHIPK2 |
miR-1249-3p/VEGFA | By competitive
absorption of miR-1249-3p mediated VEGFA | (81) |
| Infantile
hemangioma | CircAP2A2 |
miR-382-5p/VEGFA | Promotes
proliferation and invasion | (83) |
Circ_001621 is an oncogene in OS that regulates VEGF
expression through miR-578 sponge activation, and promotes the
proliferation and migration of OS cells (62). hsa_circRNA_103801 is involved in
VEGF and angiogenesis-related pathways and in OS progression, and
is expected to become a biomarker and target for the diagnosis and
treatment of late-stage OS (79).
In papillary thyroid carcinoma (PTC), circPVT1 acts as a miR-195
sponge to mediate the ceRNA network to regulate VEGFA expression
and promote PTC progression (80).
Ren et al (81) found the
abnormal overexpression of circHIPK2 in non-small cell lung cancer
(NSCLC) tissues. The overexpression of circHIPK2 promoted the
growth of cisplatin-resistant NSCLC tumors in vivo, and the
silencing of circHIPK2 inhibited angiogenesis and drug resistance
in NSCLC through miR-1249-3p targeting VEGFA. These findings
provide new insight into the development of treatment strategies
for NSCLC (81). CircFOXP1 can
simultaneously target two key miRNAs, hsa-miR-370-3p and
hsa-miR-18a-5p, to regulate the VEGF pathway in the processes of
proliferation and metastasis and is a novel serum diagnostic marker
for NSCLC (82). CircAP2A2 is
highly expressed in IH and functions as a ceRNA to promote IH
proliferation and invasion by regulating the miR-382-5p/VEGFA axis
(83).
Potential application of circRNAs in tumor
angiogenesis
circRNAs are a novel type of RNA whose structural
stability contributes to natural advantages that allow these
factors to serve as biomarkers; notably, circRNAs in plasma and
plasma-derived exosomes are highly reliable tumor diagnostic
biomarkers (84,85). Some progress has been made in tumor
angiogenesis research. The diversity of the biological functions of
circRNAs is represented by their expression in various diseases and
their involvement in multiple tumor-related processes. circRNAs are
resistant to exonuclease (RNase) activity due to their specific and
highly conserved structures, rendering them ideal novel clinical
markers.
Biomarkers
circRNAs not only play a crucial in the occurrence
and development of tumors, but are also likely to serve as markers
for the early diagnosis of tumors and targets for tumor therapy
(34). miRNA-377 is a tumor
suppressor gene that regulates multiple tumor phenotypes (86). Using both in vitro and in
vivo experiments, Fang et al (71) found that the expression level of
circ_0072088 in esophageal squamous cell carcinoma tissues and
cells was directly proportional to the tumor size, depth of
invasion and clinical stage of patients with ESCC, and that its
mechanism of action may be that circ_072088 functions as a miR-377
sponge to regulate the expression of VEGF, thereby promoting the
development of ESCC. This suggests that circ_007 2088 may be a
novel molecular marker for the early diagnosis and treatment of
ESCC (71). The splicing mechanism
is one of the primary mechanisms through which circRNAs are
generated (87). It has been
demonstrated that circSMARCA5 is expressed at low levels in GBM,
and is directly proportional to the overall survival and
disease-free survival of patients with GBM (53). Further research has found that
circSMARCA 5 acts as a sponge for the splicing factor SRSF1 in GBM,
and thus, this circRNA regulates the splicing and angiogenesis of
VEGFA mRNA in GBM by binding to SRSF1. CircSMARC5 is therefore a
promising prognostic marker in GBM (53).
Targeted drugs that act on angiogenics
pathways
As a current clinical research hotspot, circRNAs can
not only be used as molecular markers of disease, but they also
provide a basis for the development of targeted drugs. CircFOXP1 is
upregulated in OS and promotes angiogenesis in OS by regulating the
miR-127-5p/CDKN2AIP signaling pathway, which accelerates OS
progression (45), and
circRNA-related targeted drugs may be key to future
antiangiogenesis strategies in these tumors. VEGF overexpression in
tumors has been regarded as a key factor in angiogenesis, and the
inhibition of angiogenesis and related pathways is an effective
approach for the treatment of cancer (54). Bevacizumab, the only
anti-angiogenic drug approved for metastatic colon cancer in the
USA, has been shown to effectively improve the progression-free
survival of patients with metastatic colorectal cancer who are
>65 years of age (88). In
addition, it has been demonstrated that anti-VEGFA therapy
significantly improves the prognosis of patients with advanced
renal cell carcinoma and cervical cancer (89). These studies provide insight into
the development of novel targeted drugs that block the signaling
pathways related to tumor angiogenesis, and provide targets and
strategies for anti-angiogenic therapy for glioma and OS.
Conclusions and future perspectives
In recent years, some research progress has been
made on the biological functions of circRNAs; however, research on
the mechanisms of circRNAs remains insufficient. Various circRNAs
encode proteins, which provides a basis for the study of
circRNA-related mechanisms and therapeutic targets for certain
diseases. circRNAs play a key role in the pathogenesis of
cardiovascular diseases, such as heart failure, cardiac hypertrophy
and atherosclerosis (90). In
addition, circRNAs are involved in various angiogenesis-induced
pathologies, including tumors, which offers a breakthrough for
treatment strategies (3).
According to a database search performed in the
present study, it is known that circRNAs do not have a common
target in different cancer types. It is considered that the same
circRNAs are expressed differently in various tumor tissues, which
is possibly due to the complexity and heterogeneity of these
tissues. circRNA expression is not the same in different types of
cancer, and thus, no common targets for circRNAs in different
cancer types have been reported. Therefore, in future research,
tumor pathogenesis and tumor tissue heterogeneity should be
explored in depth, and common targets of circRNAs in different
types of cancer need to be actively determined to achieve further
breakthroughs in cancer.
It is worth mentioning that miRNAs can exert a
pro-angiogenic or anti-angiogenic effect by modulating endothelial
cell migration or the cell cycle. Some of these specific miRNAs
play a role in tumor angiogenesis by regulating the response of
endothelial cells to blood flow or VEGF. As an oncogene, miR-29a is
involved in the onset of a variety of cancer types. Using in
vitro and in vivo experiments, Jia et al
(91) found that lncRNA H19
expression was upregulated in microvessels of glioma-associated
endothelial cells. It was further found that lncRNA H19 functioned
as a miR-29a sponge to downregulate miR-29a by targeting the 3′-UTR
of vasohibin 2, which promoted angiogenesis; additionally, H19 gene
knockout attenuated glioma progression by inhibiting miRNA-29a,
which prevented glioma angiogenesis (91). Microvesicles can mediate
intercellular signal transduction involved in disease progression.
Zhang et al (92) used a
mouse model of tumor implantation and revealed that microvesicles
rich in miR-29a inhibited angiogenesis and reduced the tumor growth
rate, downregulated cell-derived microvesicles containing miR-29a
to alleviate VEGFA inhibition, and promoted angiogenesis in gastric
cancer (92). Therefore, perhaps
attention should be paid to the role of specific miRNAs in tumor
angiogenesis in order to provide new insight into and targets for
the treatment of tumors.
VEGF is a key mediator of tumor angiogenesis that
promotes tumor growth, and thus, VEGF is a therapeutic target, and
its inhibition has become an important tumor treatment strategy
(21). Tumor angiogenesis relies
on the coregulation of pro-angiogenic factors and anti-angiogenic
factors; in addition to VEGF (15,16),
pro-angiogenic factors include angiopoietin (17), MMPs and FGF (18–20).
circRNAs can directly or indirectly target VEGF to regulate tumor
angiogenesis. Circ-RanGAP1 (40),
circ0001429 (41) and circRNA-MYLK
(51) function as miRNA sponges
that upregulate VEGF expression to promote angiogenesis and
accelerate tumor progression. On the contrary, as a sponge for
SRSF1, circSMARCA5 (53), can
negatively regulate VEGF and angiogenesis in GBM. However, compared
with VEGF, the circRNA regulation of angiogenesis through
angiopoietin, FGF and MMPs has not been extensively studied and
warrants further investigation.
In the process through which circRNAs regulate tumor
angiogenesis, the ceRNA mechanism is a classic regulatory mechanism
and is relatively well studied. In addition, the specificity of
circRNA structure and function allows circRNAs to be regulated by
other mechanisms; for example, RBPs directly target circRNAs to
regulate tumor angiogenesis (24,53).
He et al (93) found that
the expression of the RBPs, MOV10, circ-DICER1 and ZIC4, in
glioma-exposed endothelial cells was upregulated, while that of
miR-103a-3p/miR-382-5p expression was downregulated. The mechanism
involved the binding of MOV10 to circ-DICER1, which adsorbed
miR-103a-3p/miR-382-5p as a sponge and attenuated the negative
regulation of ZIC4 by miR-103a-3p/miR-382-5, thereby upregulating
ZIC4 expression (93). The
aforementioned research not only provides a novel mechanism for
angiogenesis in glioma, but also provides a promising target for
anti-angiogenic therapy in this type of cancer.
circRNAs are recently discovered members of the RNA
family that have potential regulatory functions; they are expressed
in various types of cancer and can affect tumor angiogenesis via
multiple mechanisms. Recent studies have demonstrated that circRNAs
participate in the regulation of tumor angiogenesis via two primary
means: They can function as miRNA sponges and participate in tumor
angiogenesis by regulating related targets or pathways; and they
can also directly target RBPs to regulate tumor angiogenesis
(24,53). The present review discussed the
biological functions of circRNAs, their roles as ceRNAs in the
regulation of tumor angiogenesis, the effects of circRNAs on tumor
angiogenesis in various systems and their potential clinical
application value. Although some progress has been made in
understanding the regulation of circRNAs involved in tumor
angiogenesis, the mechanisms are primarily centered on the sponging
effect of circRNAs, while little information is available on other
mechanisms. It is therefore necessary to further explore other
relevant mechanisms and targets of circRNAs that are involved in
tumor angiogenesis. In addition, circRNAs play a role in tumor
angiogenesis, although the specific mechanisms involved remain
unclear, and few target molecules have been reported for the
treatment of related diseases. The mechanisms of circRNAs and tumor
angiogenesis thus warrant further investigation.
In conclusion, as bioinformatics continues to
develop, it is considered that circRNAs have broad prospects as
tumor diagnostic markers and anti-angiogenic therapies, and are
worthy of further investigation.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
Not applicable.
Authors' contributions
DM wrote and revised the manuscript. RJ and SY
participated in the acquisition and analysis of data from the
literature for inclusion in the study, and designed and created the
figures. MW, XB and CZ searched the references, drafted the
manuscript, and edited and revised the figures. WW and ZL conceived
and designed the study. All authors have read and approved the
final manuscript. Data authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Arnaiz E, Sole C, Manterola L,
Iparraguirre L, Otaegui D and Lawrie CH: CircRNAs and cancer:
Biomarkers and master regulators. Semin Cancer Biol. 58:90–99.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kristensen LS, Hansen TB, Venø MT and
Kjems J: Circular RNAs in cancer: Opportunities and challenges in
the field. Oncogene. 37:555–565. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Flemming A: The enigma of circular RNA.
Nat Rev Immunol. 19:3512019. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Croce CM: Genetics: Are circRNAs involved
in cancer pathogenesis? Nat Rev Clin Oncol. 13:6582016. View Article : Google Scholar
|
|
6
|
Fischer JW and Leung AK: CircRNAs: A
regulator of cellular stress. Crit Rev Biochem Mol Biol.
52:220–233. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun LL, Li WD, Lei FR and Li XQ: The
regulatory role of microRNAs in angiogenesis-related diseases. J
Cell Mol Med. 22:4568–4587. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hanahan D and Folkman J: Patterns and
emerging mechanisms of the angiogenic switch during tumorigenesis.
Cell. 86:353–364. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Baeriswyl V and Christofori G: The
angiogenic switch in carcinogenesis. Semin Cancer Biol. 19:329–337.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Miao S, Qiu T, Zhao Y, Wang H, Sun X, Wang
Y, Xuan Y, Qin Y and Jiao W: Overexpression of S100A13 protein is
associated with tumor angiogenesis and poor survival in patients
with early-stage non-small cell lung cancer. Thorac Cancer.
9:1136–1144. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu X, Cui H, Niu H, Wang L, Li X, Sun J,
Wei Q, Dong J, Liu L and Xian CJ: Hydrocortisone suppresses early
paraneoplastic inflammation and angiogenesis to attenuate early
hepatocellular carcinoma progression in rats. Onco Targets Ther.
12:9481–9493. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Goussia A, Simou N, Zagouri F, Manousou K,
Lazaridis G, Gogas H, Koutras A, Sotiropoulou M, Pentheroudakis G,
Bafaloukos D, et al: Associations of angiogenesis-related proteins
with specific prognostic factors, breast cancer subtypes and
survival outcome in early-stage breast cancer patients. A hellenic
cooperative oncology group (HeCOG) trial. PLoS One. 13:e2003022018.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Senger DR, Van de Water L, Brown LF, Nagy
JA, Yeo KT, Yeo TK, Berse B, Jackman RW, Dvorak AM and Dvorak HF:
Vascular permeability factor (VPF, VEGF) in tumor biology. Cancer
Metastasis Rev. 12:303–324. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Senger DR, Perruzzi CA, Feder J and Dvorak
HF: A highly conserved vascular permeability factor secreted by a
variety of human and rodent tumor cell lines. Cancer Res.
46:5629–5632. 1986.PubMed/NCBI
|
|
17
|
Fagiani E and Christofori G: Angiopoietins
in angiogenesis. Cancer Lett. 328:18–26. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ebrahem Q, Chaurasia SS, Vasanji A, Qi JH,
Klenotic PA, Cutler A, Asosingh K, Erzurum S and Anand-Apte B:
Cross-talk between vascular endothelial growth factor and matrix
metalloproteinases in the induction of neovascularization in vivo.
Am J Pathol. 176:496–503. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Itoh Y: Membrane-type matrix
metalloproteinases: Their functions and regulations. Matrix Biol.
44–46. 207–223. 2015.PubMed/NCBI
|
|
20
|
Cross MJ and Claesson-Welsh L: FGF and
VEGF function in angiogenesis: Signalling pathways, biological
responses and therapeutic inhibition. Trends Pharmacol Sci.
22:201–207. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Carmeliet P: VEGF as a key mediator of
angiogenesis in cancer. Oncology. 69 (Suppl 3):S4–S10. 2005.
View Article : Google Scholar
|
|
22
|
Claffey KP and Robinson GS: Regulation of
VEGF/VPF expression in tumor cells: Consequences for tumor growth
and metastasis. Cancer Metastasis Rev. 15:165–176. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Barbagallo D, Caponnetto A, Cirnigliaro M,
Brex D, Barbagallo C, D'Angeli F, Morrone A, Caltabiano R,
Barbagallo GM, Ragusa M, et al: CircSMARCA5 inhibits migration of
glioblastoma multiforme cells by regulating a molecular axis
involving splicing factors SRSF1/SRSF3/PTB. Int J Mol Sci.
19:4802018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu WY: Roles of the circular RNA
circ-Foxo3 in breast cancer progression. Cell Cycle. 16:589–590.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xiao Y: Construction of a
circRNA-miRNA-mRNA network to explore the pathogenesis and
treatment of pancreatic ductal adenocarcinoma. J Cell Biochem.
121:394–406. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zang J, Lu D and Xu A: The interaction of
circRNAs and RNA binding proteins: An important part of circRNA
maintenance and function. J Neurosci Res. 98:87–97. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:304–315. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xia X, Li X, Li F, Wu X, Zhang M, Zhou H,
Huang N, Yang X, Xiao F, Liu D, et al: A novel tumor suppressor
protein encoded by circular AKT3 RNA inhibits glioblastoma
tumorigenicity by competing with active phosphoinositide-dependent
kinase-1. Mol Cancer. 18:1312019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Zhao K, Xu X, Yang Y, Yan S, Wei
P, Liu H, Xu J, Xiao F, Zhou H, et al: A peptide encoded by
circular form of LINC-PINT suppresses oncogenic transcriptional
elongation in glioblastoma. Nat Commun. 9:44752018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Westholm JO, Miura P, Olson S, Shenker S,
Joseph B, Sanfilippo P, Celniker SE, Graveley BR and Lai EC:
Genome-wide analysis of drosophila circular RNAs reveals their
structural and sequence properties and age-dependent neural
accumulation. Cell Rep. 9:1966–1980. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bahn JH, Zhang Q, Li F, Chan TM, Lin X,
Kim Y, Wong DT and Xiao X: The landscape of microRNA,
Piwi-interacting RNA, and circular RNA in human saliva. Clin Chem.
61:221–230. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li Y, Zheng Q, Bao C, Li S, Guo W, Zhao J,
Chen D, Gu J, He X and Huang S: Circular RNA is enriched and stable
in exosomes: A promising biomarker for cancer diagnosis. Cell Res.
25:981–984. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu H, Chen D, Bi J, Han J, Yang M, Dong
W, Lin T and Huang J: Circular RNA circUBXN7 represses cell growth
and invasion by sponging miR-1247-3p to enhance B4GALT3 expression
in bladder cancer. Aging (Albany NY). 10:2606–2623. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Memczak S, Jens M, Elefsinioti A, Torti F,
Krueger J, Rybak A, Maier L, Mackowiak SD, Gregersen LH, Munschauer
M, et al: Circular RNAs are a large class of animal RNAs with
regulatory potency. Nature. 495:333–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Taulli R, Loretelli C and Pandolfi PP:
From pseudo-ceRNAs to circ-ceRNAs: A tale of cross-talk and
competition. Nat Struct Mol Biol. 20:541–543. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Annese T, Tamma R, De Giorgis M and
Ribatti D: microRNAs biogenesis, functions and role in tumor
angiogenesis. Front Oncol. 10:5810072020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Tay Y, Rinn J and Pandolfi PP: The
multilayered complexity of ceRNA crosstalk and competition. Nature.
505:344–352. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lu J, Wang YH, Yoon C, Huang XY, Xu Y, Xie
JW, Wang JB, Lin JX, Chen QY, Cao LL, et al: Circular RNA
circ-RanGAP1 regulates VEGFA expression by targeting miR-877-3p to
facilitate gastric cancer invasion and metastasis. Cancer Lett.
471:38–48. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cao W, Zhao Y, Wang L and Huang X:
Circ0001429 regulates progression of bladder cancer through binding
miR-205-5p and promoting VEGFA expression. Cancer Biomark.
25:101–113. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Meng Q, Li S, Liu Y, Zhang S, Jin J, Zhang
Y, Guo C, Liu B and Sun Y: Circular RNA circSCAF11 accelerates the
glioma tumorigenesis through the miR-421/SP1/VEGFA axis. Mol Ther
Nucleic Acids. 17:669–677. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wang J, Lin Y, Jiang DH, Yang X and He XG:
CircRNA ZNF609 promotes angiogenesis in nasopharyngeal carcinoma by
regulating miR-145/STMN1 axis. Kaohsiung J Med Sci. 37:686–698.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yu J, Yang M, Zhou B, Luo J, Zhang Z,
Zhang W and Yan Z: CircRNA-104718 acts as competing endogenous RNA
and promotes hepatocellular carcinoma progression through
microRNA-218-5p/TXNDC5 signaling pathway. Clin Sci (Lond).
133:1487–1503. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Zhang H, Yu Z, Wu B and Sun F: Circular
RNA circFOXP1 promotes angiogenesis by regulating
microRNA-127-5p/CDKN2AIP signaling pathway in osteosarcoma.
Bioengineered. 12:9991–9999. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cassiday LA and Maher LR III: Having it
both ways: Transcription factors that bind DNA and RNA. Nucleic
Acids Res. 30:4118–4126. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
He Z, Ruan X, Liu X, Zheng J, Liu Y, Liu
L, Ma J, Shao L, Wang D, Shen S, et al:
FUS/circ_002136/miR-138-5p/SOX13 feedback loop regulates
angiogenesis in glioma. J Exp Clin Cancer Res. 38:652019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ferrara N: Vascular endothelial growth
factor: Basic science and clinical progress. Endocr Rev.
25:581–611. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Guo J, Chen M, Ai G, Mao W, Li H and Zhou
J: Hsa_circ_0023404 enhances cervical cancer metastasis and
chemoresistance through VEGFA and autophagy signaling by sponging
miR-5047. Biomed Pharmacother. 115:1089572019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Garikipati VNS, Verma SK, Cheng Z, Liang
D, Truongcao MM, Cimini M, Yue Y, Huang G, Wang C, Benedict C, et
al: Circular RNA CircFndc3b modulates cardiac repair after
myocardial infarction via FUS/VEGF-A axis. Nat Commun. 10:43172019.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhong Z, Huang M, Lv M, He Y, Duan C,
Zhang L and Chen J: Circular RNA MYLK as a competing endogenous RNA
promotes bladder cancer progression through modulating VEGFA/VEGFR2
signaling pathway. Cancer Lett. 403:305–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Li P, Chen H, Chen S, Mo X, Li T, Xiao B,
Yu R and Guo J: Circular RNA 0000096 affects cell growth and
migration in gastric cancer. Br J Cancer. 116:626–633. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Barbagallo D, Caponnetto A, Brex D,
Mirabella F, Barbagallo C, Lauretta G, Morrone A, Certo F, Broggi
G, Caltabiano R, et al: CircSMARCA5 regulates VEGFA mRNA splicing
and angiogenesis in glioblastoma multiforme through the binding of
SRSF1. Cancers (Basel). 11:1942019. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Carmeliet P and Jain RK: Molecular
mechanisms and clinical applications of angiogenesis. Nature.
473:298–307. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Costa PM, Cardoso AL, Pereira de Almeida
LF, Bruce JN, Canoll P and Pedroso de Lima MC: PDGF-B-mediated
downregulation of miR-21: New insights into PDGF signaling in
glioblastoma. Hum Mol Genet. 21:5118–5130. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Zehetner C, Kirchmair R, Neururer SB,
Kralinger MT, Bechrakis NE and Kieselbach GF: Systemic upregulation
of PDGF-B in patients with neovascular AMD. Invest Ophthalmol Vis
Sci. 55:337–344. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu F, Zhang H, Xie F, Tao D, Xiao X,
Huang C, Wang M, Gu C, Zhang X and Jiang G: Hsa_circ_0001361
promotes bladder cancer invasion and metastasis through
miR-491-5p/MMP9 axis. Oncogene. 39:1696–1709. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liu H, Lan T, Li H, Xu L, Chen X, Liao H,
Chen X, Du J, Cai Y, Wang J, et al: Circular RNA circDLC1 inhibits
MMP1-mediated liver cancer progression via interaction with HuR.
Theranostics. 11:1396–1411. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hou JP, Men XB, Yang LY, Han EK, Han CQ
and Liu LB: CircCCT3 acts as a sponge of miR-613 to promote tumor
growth of pancreatic cancer through regulating VEGFA/VEGFR2
signaling. Balkan Med J. 38:229–238. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Zhu J, Luo Y, Zhao Y, Kong Y, Zheng H, Li
Y, Gao B, Ai L, Huang H, Huang J, et al: circEHBP1 promotes
lymphangiogenesis and lymphatic metastasis of bladder cancer via
miR-130a-3p/TGFβR1/VEGF-D signaling. Mol Ther. 29:1838–1852. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Qi L, Wang W, Zhao G, Jiang H, Zhang Y,
Zhao D, Jin H, Yu H and Xu H: Circular RNA circitga7 accelerates
glioma progression via miR-34a-5p/VEGFA axis. Aging (Albany NY).
13:13138–13152. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Ji X, Shan L, Shen P and He M: Circular
RNA circ_001621 promotes osteosarcoma cells proliferation and
migration by sponging miR-578 and regulating VEGF expression. Cell
Death Dis. 11:182020. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Li S, Li J, Zhang H, Zhang Y, Wang X, Yang
H, Zhou Z, Hao X, Ying G and Ba Y: Gastric cancer derived exosomes
mediate the delivery of circRNA to promote angiogenesis by
targeting miR-29a/VEGF axis in endothelial cells. Biochem Biophys
Res Commun. 560:37–44. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kong Y, Li Y, Luo Y, Zhu J, Zheng H, Gao
B, Guo X, Li Z, Chen R and Chen C: circNFIB1 inhibits
lymphangiogenesis and lymphatic metastasis via the
miR-486-5p/PIK3R1/VEGF-C axis in pancreatic cancer. Mol Cancer.
19:822020. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Xing C, Ye H, Wang W, Sun M, Zhang J, Zhao
Z and Jiang G: Circular RNA ADAM9 facilitates the malignant
behaviours of pancreatic cancer by sponging miR-217 and
upregulating PRSS3 expression. Artif Cells Nanomed Biotechnol.
47:3920–3928. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zheng X, Ma YF, Zhang XR, Li Y, Zhao HH
and Han SG: Circ_0056618 promoted cell proliferation, migration and
angiogenesis through sponging with miR-206 and upregulating CXCR4
and VEGF-A in colorectal cancer. Eur Rev Med Pharmacol Sci.
24:4190–4202. 2020.PubMed/NCBI
|
|
67
|
Dai J, Zhuang Y, Tang M, Qian Q and Chen
JP: CircRNA UBAP2 facilitates the progression of colorectal cancer
by regulating miR-199a/VEGFA pathway. Eur Rev Med Pharmacol Sci.
24:7963–7971. 2020.PubMed/NCBI
|
|
68
|
Chen MS, Lin CH, Huang LY and Qiu XM:
CircRNA SMARCC1 sponges MiR-140-3p to regulate cell progression in
colorectal cancer. Cancer Manag Res. 12:4899–4910. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chen C, Huang Z, Mo X, Song Y, Li X, Li X
and Zhang M: The circular RNA 001971/miR-29c-3p axis modulates
colorectal cancer growth, metastasis, and angiogenesis through
VEGFA. J Exp Clin Cancer Res. 39:912020. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Li W, Xu Y, Wang X, Cao G, Bu W, Wang X,
Fang Z, Xu Y, Dong M and Tao Q: circCCT3 modulates vascular
endothelial growth factor A and Wnt signaling to enhance colorectal
cancer metastasis through sponging miR-613. DNA Cell Biol.
39:118–125. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Fang N, Shi Y, Fan Y, Long T, Shu Y and
Zhou J: Circ_0072088 promotes proliferation, migration, and
invasion of esophageal squamous cell cancer by absorbing miR-377. J
Oncol. 2020:89671262020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Gao S, Hu W, Huang X, Huang X, Chen W, Hao
L, Chen Z, Wang J and Wei H: Circ_0001178 regulates miR-382/VEGFA
axis to facilitate hepatocellular carcinoma progression. Cell
Signal. 72:1096212020. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chen J, Li X, Yang L, Li M, Zhang Y and
Zhang J: CircASH2L promotes ovarian cancer tumorigenesis,
angiogenesis, and lymphangiogenesis by regulating the miR-665/VEGFA
axis as a competing endogenous RNA. Front Cell Dev Biol.
8:5955852020. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Wang LL, Zong ZH, Liu Y, Guan X, Chen S
and Zhao Y: CircRhoC promotes tumorigenicity and progression in
ovarian cancer by functioning as a miR-302e sponge to positively
regulate VEGFA. J Cell Mol Med. 23:8472–8481. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Li QH, Liu Y, Chen S, Zong ZH, Du YP,
Sheng XJ and Zhao Y: circ-CSPP1 promotes proliferation, invasion
and migration of ovarian cancer cells by acting as a miR-1236-3p
sponge. Biomed Pharmacother. 114:1088322019. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lu Y, Qin T, Li J, Wang L, Zhang Q, Jiang
Z and Mao J: MicroRNA-140-5p inhibits invasion and angiogenesis
through targeting VEGF-A in breast cancer. Cancer Gene Ther.
24:386–392. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang B, Duan R, Li ZB and Wang L:
Circ-RPL15/miR-146b-3p/VEGFA feedback loop is responsible for
triggering proliferation and migration in glioma. Eur Rev Med
Pharmacol Sci. 24:6204–6210. 2020.PubMed/NCBI
|
|
78
|
Barbagallo D, Caponnetto A, Barbagallo C,
Battaglia R, Mirabella F, Brex D, Stella M, Broggi G, Altieri R,
Certo F, et al: The GAUGAA motif is responsible for the binding
between circSMARCA5 and SRSF1 and related downstream effects on
glioblastoma multiforme cell migration and angiogenic potential.
Int J Mol Sci. 22:16782021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Liu W, Zhang J, Zou C, Xie X, Wang Y, Wang
B, Zhao Z, Tu J, Wang X, Li H, et al: Microarray expression profile
and functional analysis of circular RNAs in osteosarcoma. Cell
Physiol Biochem. 43:969–985. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zeng L, Yuan S, Zhou P, Gong J, Kong X and
Wu M: Circular RNA Pvt1 oncogene (CircPVT1) promotes the
progression of papillary thyroid carcinoma by activating the
Wnt/β-catenin signaling pathway and modulating the ratio of
microRNA-195 (miR-195) to vascular endothelial growth factor A
(VEGFA) expression. Bioengineered. 12:11795–11810. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Ren M, Song X, Niu J, Tang G, Sun Z, Li Y
and Kong F: The malignant property of circHIPK2 for angiogenesis
and chemoresistance in non-small cell lung cancer. Exp Cell Res.
419:1132762022. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Luo Y, Zhang Q, Lv B, Shang Y, Li J, Yang
L, Yu Z, Luo K, Deng X, Min L and Zhu T: CircFOXP1: A novel serum
diagnostic biomarker for non-small cell lung cancer. Int J Biol
Markers. 37:58–65. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Yuan X, Xu Y, Wei Z and Ding Q: CircAP2A2
acts as a ceRNA to participate in infantile hemangiomas progression
by sponging miR-382-5p via regulating the expression of VEGFA. J
Clin Lab Anal. 34:e232582020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Chen B and Huang S: Circular RNA: An
emerging non-coding RNA as a regulator and biomarker in cancer.
Cancer Lett. 418:41–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Bai H, Lei K, Huang F, Jiang Z and Zhou X:
Exo-circRNAs: A new paradigm for anticancer therapy. Mol Cancer.
18:562019. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liu Y, Gao Y, Li D, He L, Iw L, Hao B,
Chen X and Cao Y: LASP1 promotes glioma cell proliferation and
migration and is negatively regulated by miR-377-3p. Biomed
Pharmacother. 108:845–851. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Montes M, Sanford BL, Comiskey DF and
Chandler DS: RNA splicing and disease: Animal models to therapies.
Trends Genet. 35:68–87. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Mody K, Baldeo C and Bekaii-Saab T:
Antiangiogenic therapy in colorectal cancer. Cancer J. 24:165–170.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Ferrara N and Adamis AP: Ten years of
anti-vascular endothelial growth factor therapy. Nat Rev Drug
Discov. 15:385–403. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Altesha MA, Ni T, Khan A, Liu K and Zheng
X: Circular RNA in cardiovascular disease. J Cell Physiol.
234:5588–5600. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jia P, Cai H, Liu X, Chen J, Ma J, Wang P,
Liu Y, Zheng J and Xue Y: Long non-coding RNA H19 regulates glioma
angiogenesis and the biological behavior of glioma-associated
endothelial cells by inhibiting microRNA-29a. Cancer Lett.
381:359–369. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zhang H, Bai M, Deng T, Liu R, Wang X, Qu
Y, Duan J, Zhang L, Ning T, Ge S, et al: Cell-derived microvesicles
mediate the delivery of miR-29a/c to suppress angiogenesis in
gastric carcinoma. Cancer Lett. 375:331–339. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
He Q, Zhao L, Liu X, Zheng J, Liu Y, Liu
L, Ma J, Cai H, Li Z and Xue Y: MOV10 binding circ-DICER1 regulates
the angiogenesis of glioma via miR-103a-3p/miR-382-5p mediated ZIC4
expression change. J Exp Clin Cancer Res. 38:92019. View Article : Google Scholar : PubMed/NCBI
|