Introduction
The Helicase POLQ-like (HELQ) gene, also
referred to as Hel308/holliday junction migration protein,
maps to human chromosome 4q21 and was first cloned by its homology
to Drosophila melanogaster (D. melanogaster)
mutagen-sensitive 308 (Mus308) (1).
In humans, HELQ is typical superfamily-2 (SF2) ATPase-dependent 3′
to 5′ helicase as a functions to unwind DNA (2,3). As
HELQ has ATPase and helicase functions, it has been classified as a
member of the Ski2-like subfamily of SF2 helicases (4), which is a small subfamily (5,6)
involved in RNA processing, splicing and degradation pathways
(7). Similar to other SF2
helicases, HELQ is a regulator of genome stability (2), DNA recombination (2), DNA binding (8), DNA unwinding (9), DNA replication (10), DNA strand annealing (9) and DNA repair (11,12).
HELQ is widely expressed in normal human tissues,
particularly of the reproductive system, and plays an important
role in maintaining germ cells by promoting DNA repair protein
RecA-like ATP-dependent recombinase 51 (RAD51) homolog 1 (RAD51)
paralog-dependent repair (11).
Furthermore, HELQ is abnormally expressed in multiple tumors, and
contributes to cell proliferation, metastasis and chemotherapy
resistance through various molecular pathways, such as nucleotide
excision repair (NER), as well as the checkpoint kinase 1
(CHK1)-RAD51 and ataxia telangiectasia mutated protein/ataxia
telangiectasia and Rad3-related protein (ATM/ATR) signaling
pathways (12,13). HELQ can also contribute to germ cell
maintenance and tumorigenesis in mammals by regulating its target
genes, which include CHK1, fanconi anemia group D2
(FANCD2), RAD51 and replication protein A (RPA),
among others (3,4,9,14). Of
note, the low expression or loss of HELQ can have a significant
impact on clinical features and prognosis in patients with
cancer.
In the present review, the structural features,
function, target genes, roles in reproduction, molecular mechanisms
associated with tumorigenesis, and potential applications of HELQ
were summarized. It was concluded that HELQ is an attractive future
clinical target in conditions involving germ cell maintenance and
in molecular targeted cancer therapies.
Structural features of HELQ
SF2 helicase family
The DNA helicases are classified into five different
superfamilies (SF1-SF5) (15). SF1
and SF2 helicases encompass a large group of DNA and RNA helicases
found in eubacteria, archaea, viruses and eukaryotes. They possess
an ATP-dependent translocation module consisting of two RecA-fold
domains responsible for nucleic acid and ATP binding (15,16).
HELQ was first described by Marini in 2002, which belongs to the
SF2 protein family (1). SF2 family
proteins have a pair of RecA-like domains that provide motion
associated with helicase activity (6,17–19)
and play key roles in chromatin rearrangement, DNA repair and
transcription (20–22) and RNA metabolism (17,23).
SF2 family proteins are divided into Ski2-like, RecQ-like,
RecG-like, RecA-like ATP-dependent recombinase 3 (Rad3)/XPD, type I
restriction enzymes, DEAD-box and NS3/NPH-II subfamilies, Swi/Snf,
DEAH/RNA helicase A, RIG-I-like, based on sequence homology
(6,24,25).
In archaea, in all Ski2-like helicases including Mtr4, Ski2, and
Bad Response to Refrigeration 2 homolog, the molecular ‘core’ of
them is a ring-like assembly of four domains comprising a ratchet
domain, a winged helix domain and two RecA domains (25).
Structural and functional
characteristics of HELQ
HELQ maps to chromosome 4q21 (1) and generates a full-length mRNA
(NM_133636.5) comprising 3,543 base pairs and encoding a protein of
1,101 amino acids (aa) (1). A total
of 7 transcribed splice isoforms of HELQ have been
described, encoding six HELQ protein variants (26). HELQ is a conserved protein that
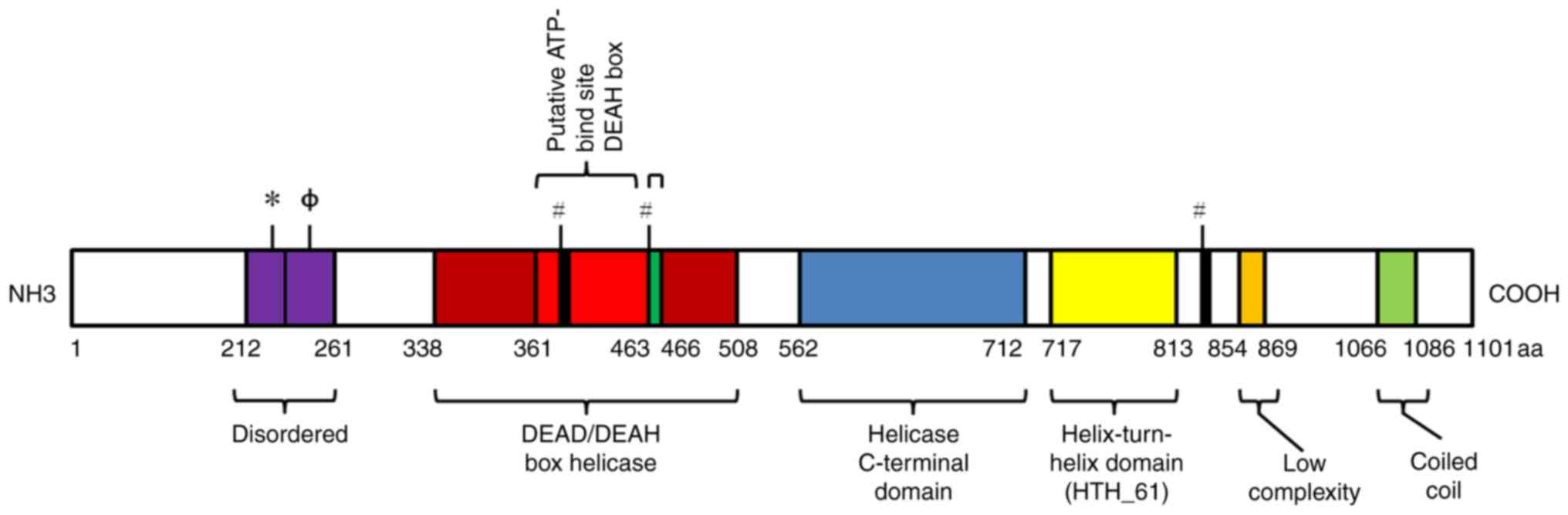
possesses three main protein domains: The DEAD/DEAH box helicase
domain, a helix-turn-helix (HTH_61) domain and a helicase
C-terminal domain (Fig. 1). The
domain of DEAD/DEAH box helicase contains a DEAH box and an
ATP-binding region (putative ATP binding site), and is involved in
unwinding nucleic acids. In addition, human HELQ has a disordered
domain (212–261 aa), containing two compositionally biased regions,
comprising basic and acidic (214–228 aa) and polar (229–253 aa)
residues. A structural diagram of the human HELQ protein is
presented in Fig. 1.
The DEAD/DEAH box helicase class of proteins, which
share eight conserved motifs (I, Ia, Ib, II, III, IV, V and VI)
(27). ATP is bound to motifs I and
II, while RNA is bound to motifs Ia and Ib. The DEAD/DEAH box is
named after the Walker B motif (motif III) consensus sequence
(28) and functions in ATP
hydrolysis. Motifs IV and V perform similar functions to those of
motifs Ia and Ib, respectively, in RNA binding (27). DEAD/DEAH box helicases are found in
various prokaryotes and eukaryotes (28), and are involved in several aspects
of RNA metabolism, such as pre-mRNA splicing, RNA decay, nuclear
transcription, editing, nucleocytoplasmic transport, ribosome
biogenesis, translation and organellar gene expression (29–31).
The helicase C-terminal domain (HelicC) is present
in various SF1 and SF2 helicases and helicase-related proteins. The
HelicC domain does not fold autonomously, but rather as a component
of the helicase, which participates in the ATP-dependent unwinding
of DNA or RNA.
The α-helical protein domain family includes winged
helix-turn-helix (HTH) domains that have characteristic folds,
which function as sequence-specific DNA-binding domains (32). The HTH plays an important role in
DNA binding and protein interactions (33,34)
and consists of two α-helices (α-helix 22 and 23) and a β-sheet
turn, where a double-stranded DNA (dsDNA) major groove can be
recognized by α-helix 23 (14,2).
HELQ expression pattern
HELQ is highly conserved across a wide range of
species from archaea through to mammals (https://www.ncbi.nlm.nih.gov/homologene/?term=HELQ).
For example, Homo sapiens HELQ shares a >97, 78 and 60%
DNA similarity with the homologous Macaca mulatta, Mus
musculus, and Danio rerio genes, respectively (Table I). It is a conserved gene in
eukaryotes that HELQ (HomoloGene ID: 14667) has important molecular
functions (i.e., ATP binding, DNA binding, single-stranded 3′-5′DNA
helicase activity) and cellular component classifications (i.e.,
nucleus, site of DNA damage), and acts in biological processes
(i.e., DNA duplex unwinding, rRNA processing) (1,9,35–37)
(https://www.alliancegenome.org/gene/HGNC:18536)
(Table II). Since HELQ is
expressed in a wide variety of species, it was likely present in a
common ancestor of vertebrates.
 | Table I.Conserved gene HELQ homology in
Eukaryota (HomoloGene: 14667)
[https://www.ncbi.nlm.nih.gov/homologene/?term=HELQ]. |
Table I.
Conserved gene HELQ homology in
Eukaryota (HomoloGene: 14667)
[https://www.ncbi.nlm.nih.gov/homologene/?term=HELQ].
| HomoloGene: 14667.
Gene conserved in Eukaryota |
|---|
|
|---|
| Species | Symbol | Genetic
location | Protein Acc. | Protein length | Identity (%)*
protein | DNA |
|---|
| H.
sapiens | HELQ | Chr4 q21.23 | NP_598375.2 | 1101 aa |
|
|
| P.
troglodytes | HELQ | Chr4 | XP_003310356.1 | 1101 aa | 99.1 | 99.4 |
| M.
mulatta | HELQ | Chr5 |
XP-_001104832.1 | 1101 aa | 97.0 | 97.7 |
| C.
lupus | HELQ | Chr32 | XP_544959.2 | 1072 aa | 85.9 | 88.4 |
| B.
taurus | HELQ | Chr6 | XP_002688448.3 | 1094 aa | 83.7 | 86.3 |
| M.
musculus | Helq | Chr5 E4 | NP_001074576.1 | 1069 aa | 78.6 | 79.3 |
| R.
norvegicus | Helq | Chr14 p22 | NP_001014156.2 | 1065 aa | 79.1 | 79.0 |
| G.
gallus | HELQ | Chr4 | XP_420565.1 | 1048 aa | 68.8 | 69.7 |
| X.
tropicalis | helq | Chr1 | XP_002939628.2 | 1000 aa | 66.0 | 66.8 |
| D.
rerio | helq | Chr21 | XP_691411.3 | 1010 aa | 60.9 | 61.6 |
| D.
melanogaster | mus301 | Chr3-22cM | NP_648178.1 | 1051 aa | 45.8 | 50.5 |
| A.
gambiae |
AgaP-AGAP012297 | Chr3L | XP_551895.3 | 914
aa | 44.4 | 48.3 |
| C.
elegans | helq-1 | ChrIII | NP_001022911.1 | 923
aa | 41.2 | 49.9 |
| Table II.Function-Gene Ontology Annotations of
HELQ (https://www.alliancegenome.org/gene/HGNC:18536). |
Table II.
Function-Gene Ontology Annotations of
HELQ (https://www.alliancegenome.org/gene/HGNC:18536).
| Category | Classification
term | Gene Ontology
IDa | Reference |
|---|
| Molecular
function | ATP binding | GO:0005524 | GO_REF:0000043 |
|
| ATP hydrolysis
activity | GO:0016887 | GO_REF:0000116 |
|
| DNA binding | GO:0003677 | GO_REF:0000043 |
|
| Single-stranded
3′-5′ DNA helicase activity | GO:1990518 | (1,9,35) |
| Cellular
component | Nucleus | GO:0005634 | (9,36) |
|
| Site of DNA
damage | GO:0090734 | (9) |
|
| DNA double-strand
break processing involved in repair via single-strand
annealing | GO:0010792 | (9) |
|
| DNA duplex
unwinding | GO:0032508 | GO_REF:0000108 |
|
| Double-strand break
repair via alternative non-homologous end joining | GO:0097681 | (9) |
| Biological
process | Double-strand break
repair via homologous recombination | GO:0000724 | (37) |
|
| Double-strand break
repair via synthesis-dependent strand annealing | GO:0045003 | (9) |
|
| Positive regulation
of double-strand break repair via Homologous recombination | GO:1905168 | GO_REF:0000024 |
|
| rRNA
processing | GO:0006364 | (36) |
Physiological functions of HELQ
DNA helicases are ubiquitous in living organisms,
where they facilitate processes involved in DNA metabolism through
unwinding the DNA double helix (3).
It is essential for the maintenance of genome stability for DNA
helicases to function in the replication and repair of DNA
(38). HELQ is a single-stranded
3′-5′ DNA helicase with critical roles in DNA repair, binding,
unwinding, replication and strand annealing (2,8,9).
DNA repair
DNA damage is a common event that may have either
endogenous or exogenous causes and can lead to mutations,
cell/organ death and cancer (39,40).
It is crucial to repair damaged DNA after it has been damaged, as
this allows damaged DNA to regain its original structure and
function normally (34,41). Various pathways recognize and repair
different types of DNA lesions, including direct repair, NER, base
excision repair, inter-strand cross-link (ICL) repair and
double-strand break (DSB) repair (39). HELQ maintains genomic stability and
avoids tumorigenesis through its involvement in different repair
pathways, including NER, DSB repair and ICL repair (9,11–13).
A NER pathway is involved in the removal of DNA
damage after certain types of DNA damage have occurred (13). The expression levels of NER pathway
proteins [e.g., xeroderma pigmentosum complementation group A
(XPA), XPC, replication protein A (RPA) and ERCC excision repair 1,
endonuclease non-catalytic subunit] are important mediators for
responses to platinum-based chemotherapy and influence DNA repair
activity (13,42). HELQ is crucial for cellular
responses to cisplatin through its role in regulating the
expression of NER pathway proteins (13).
DSBs are major DNA lesions deleterious to cell
survival and genetic stability (43). When DSBs are not repaired, they can
result in chromosome loss and rearrangements and even
carcinogenesis (44). Two major
pathways can repair DSBs: Homologous recombination (HR) and
non-homologous end joining (NHEJ) (45,46),
as well as alternative pathways, such as microhomology-mediated end
joining (MMEJ) and single strand annealing (SSA) (45,46).
HR is essential for DSB repair in post-replicative chromatin
following replication fork collapse (47,48),
and requires the loading of the RAD51 recombinase onto
single-stranded DNA (ssDNA) through DNA ends or at post-replicative
ssDNA gaps (45). The function of
HELQ in HR is to capture RPA-bound ssDNA and then to displace it to
speed up the annealing of complementary DNA strands (9). HELQ immediately interacts with BCDX2,
a paralog of RAD51, to accelerate effective HR in damaged
replication forks (11). RFS-1
binds to HELQ and plays a complementary role in facilitating the
breakdown of RAD51 dsDNA filaments in postsynaptic HR
intermediates, which is required to complete meiotic DSB repair in
Caenorhabditis elegans (47). In addition, the HR factor with
OB-fold (HROB)-mini-chromosome maintenance 8 (MCM8)-MCM9 pathway
functions redundantly with HELQ, supporting a postsynaptic step of
HR in a parallel pathway (49).
HELQ accelerates HR-efficiency at compromised replication forks by
working in parallel with the FA pathway in HELQΔC/ΔC and
FANCD2−/− double mutant mice (11). Furthermore, HELQ is essential for
the function of the synthesis-dependent strand annealing modes of
HR, MMEJ of G4-induced DSBs and SSA in genome stability and tumor
avoidance (9).
ICLs are also deleterious DNA lesions caused by
endogenous (malondialdehyde) or exogenous [mitomycin C (MMC);
cis-platinum and psoralens] sources (50,51),
which induce mutations and chromosomal rearrangements by inhibiting
DNA replication and transcription (1,34). ICL
repair is complex and proteins from the NER, translation synthesis
and HR pathways are involved in ICL repair (40). With its association with RAD51
paralogs, HELQ prevents germ cell loss and reduces cancer
susceptibility through ICL repair (11,52).
HELQ deficiency leads to germ cell attrition and ICL repair
sensitivity in mouse and human cells (11,12,53).
The FA pathway plays a critical role in recruiting RAD51-mediated
HR during ICL repair (40). In
parallel with HELQ, HROB participates in ICL repair as epistatic
with MCM8-MCM9 (49), which also
directly facilitates the repair of ICL-induced damage downstream of
FANCD2 ubiquitylation (49,54).
DNA binding and unwinding
The HELQ HTH domain is associated with DNA strand
binding and protein interactions (2,34). The
dissociation constant values of wild-type HELQ for ssDNA and dsDNA
were reported as 0.14 and 5.3 µM, respectively, indicating a strong
preference for ssDNA binding and suggesting that the protein must
track along, displacing the dsDNA strand (2). Furthermore, HELQ mutations reduce
dsDNA binding, but ssDNA binding is not affected (55). The HELQ-ssDNA interaction is
essential for the translocation mechanism. Mechanistically, HELQ
interacts with RPA, and RPA coordinates the loading of HELQ onto
ssDNA. HELQ helicase core is activated by ATP-Mg2+
binding and translocates along ssDNA as a dimer when loaded onto
ssDNA (35).
In addition, HELQ is an ssDNA-activated ATPase,
which is important for unwinding forked DNA (10,56).
As well as unwinding ssDNA and dsDNA junctions, and HELQ is capable
of unwinding 3′ overhangs, 3′ lagging strand forks, Y-structures,
and D-loops (3); however, HELQ
cannot unwind using ATPγS or 5′ overhang substrates (9). The HELQ unwinding of 3′ overhang
substrates is inhibited by RPA, whereas RAD51 stimulates the
unwinding activity of its D-loops by forming a complex with HELQ
(9).
DNA replication
HELQ is an ATP-dependent enzyme involved in the
recovery of replication forks that have stalled or collapsed
following DNA damage (3). HELQ can
act on damaged replication forks where the leading strand template
for DNA replication has stopped, causing the polymerase to uncouple
and continue DNA synthesis from the lagging strand template
(10). Following treatment with
camptothecin (CPT), an agent that stalls and collapses replication
forks, HELQ is recruited to stalled replication sites that are
associated with replication resumption. HELQ can facilitate
replication resumption, possibly through colocalizing with IdU
incorporation sites and RPA foci, by unwinding the nascent lagging
strand, or alternatively through HELQ co-localization with RAD51
and FANCD2 at sites of stalled replication (3,12). In
addition, HELQ has an important role in rescuing stalled forks
during the normal S phase, and the loss of HELQ results in
increased stalled forks, a role which is not epistatic with that of
FA complementation group C (53).
DNA strand annealing
Previous studies of HELQ have focused more on its
DNA repair and DNA unwinding functions, while its role in DNA
strand annealing has been underappreciated. Tafel et al
(3) reported that HELQ does not
exhibit a strong annealing activity, and that RPA can suppress
separated strand reannealing by binding to unwound ssDNA generated
by HELQ. By contrast, Anand et al (9) reported that RPA strongly accelerated
the DNA strand annealing activity of HELQ. Mechanistically, in
addition to capturing RPA-bound DNA strands, HELQ is capable of
superseding RPA and stimulating complementary DNA strands.
Furthermore, it was found that ATP binding and hydrolysis are
essential for HELQ DNA annealing activity, whereas the RAD51
addition was unaffected by HELQ-dependent DNA annealing activity
(9).
Gene functions regulated by HELQ
One important question is ‘How are HELQ, its
downstream mediators, and their mechanisms related in normal and
cancer cells?’ Further, there are various studies reporting that
HELQ can contribute to both germ cell maintenance and tumorigenesis
in mammals by regulating its target genes: CHK1, FANCD2,
RAD51 and RPA (3,4,9,14).
CHK1
CHK1 is a primary effector of DNA lesion and
replication checkpoint responses, and its inhibition promotes DNA
damage and reduces HR repair (57,58).
CHK1 physically interacts with RAD51, while CHK1/RAD51 disruption
or inactivation induces defective HR, aberrant replication
dynamics, and chromosome instability (59). In osteosarcoma cells, CHK1
activation is promoted by HELQ, and HELQ colocalizes with RAD51 to
participate in the repair of damaged forks by HR (3).
FANCD2
FA is a rare genetic disease, and patients with FA
are hypersensitive to DNA ICL-inducing agents such as mitomycin C
and cisplatin (60). FA signal
transduction involves 22 proteins, which share a common pathway
that is activated on DNA damage (61). FANCD2 is currently the focus of
research into the FA pathway, and is crucial in cellular responses
to DNA lesions (62). HR may be
regulated by the FA pathway, where activated FANCD2 is translocated
to chromatin-associated foci, where it colocalizes with HR proteins
[RAD51 and breast invasive carcinoma (BRCA)2] (63). The strong negative effect of HELQ
knockdown on single-stranded templated repair via the HR repair
pathway may be explained by its physical interaction with FANCD2
and RAD51 (64). Following CPT
treatment, HELQ is involved in fork repair and restart by
localizing to replication forks, unwinding lagging strand
structures, and co-localizing with RAD51 and FANCD2 (3,14). In
addition, although the FA pathway, ICL sensitivities, and HELQ are
additive, there is no epistasis of HELQ for downstream targets of
FANCD2, and HELQ plays an independent role from FA pathway in ICL
processing (12).
RAD51
RAD51 is a DNA-binding protein that can bind to both
ssDNA and dsDNA and maintain genome stability in DNA replication
(65,66). RAD51 is also an ATPase that forms
nucleoprotein filaments on ssDNA and facilitates the search for
homologous repair templates, such as homologous chromosomes, sister
chromatids or ectopic homologous sequences (67). Hence, RAD51 is essential for
regulating fork reversal by discovering and invading homologous DNA
sequences during DSB repair by HR (65,67).
In addition, five RAD51 paralogs [RAD51B/C/D and X-ray repair cross
complementing (XRCC)2/3] that form two main complexes, BCDX2
(RAD51B/C/D-XRCC2) and CX3 (RAD51C-XRCC3), are involved in the HR
repair of collapsed replication forks and maintenance of genome
stability (68–70). The BCDX2 complex preferentially
binds to ssDNA and accelerates HELQ ATPase activity (71), which plays a central role in
recognizing damage and stimulating fork remodeling in response to
fluctuating dNTP pools (72).
Adelman et al (11) found
that HELQ plays a critical role in replication-coupled HR by
interacting with complexes of the RAD51 paralog, BCDX2, to avoid
germ cell loss and tumorigenesis. Anand et al (9) also demonstrated that HELQ binds to
BCDX2 complexes, and that the latter strongly stimulates the
translocation of HELQ during DNA unwinding. Takata et al
(12) reported that HELQ interacts
with ATR and HR-related RAD51 paralogs and participates in DNA LCL
resistance in human cells.
RPA
As a major ssDNA binding protein, RPA is abundant in
eukaryotic cells (66,73). RPA comprises three subunits, RPA14,
RPA32 and RPA70, which are first responders in the event of the
disruption of DNA metabolism. RPA is essential for replication,
recombination and repair by binding to ssDNA to maintain genome
duplication and stability (74).
RPA and HELQ physically interact to form a supershift complex with
HELQ on ssDNA (4). Furthermore, RPA
stimulates HELQ helicase activity by binding to unwound regions
generated by HELQ and inhibiting reannealing (4). Similarly, in mammals, RPA can restrain
DNA unwinding while promoting DNA strand annealing by HELQ.
Mechanistically, HELQ is recruited to ssDNA by interacting with
RPA, followed by RPA displacement to stimulate complementary DNA
strand annealing (9).
The roles of HELQ in reproduction
HELQ was first identified in the mouse and human
genomes through its homology to D. melanogaster Mus308
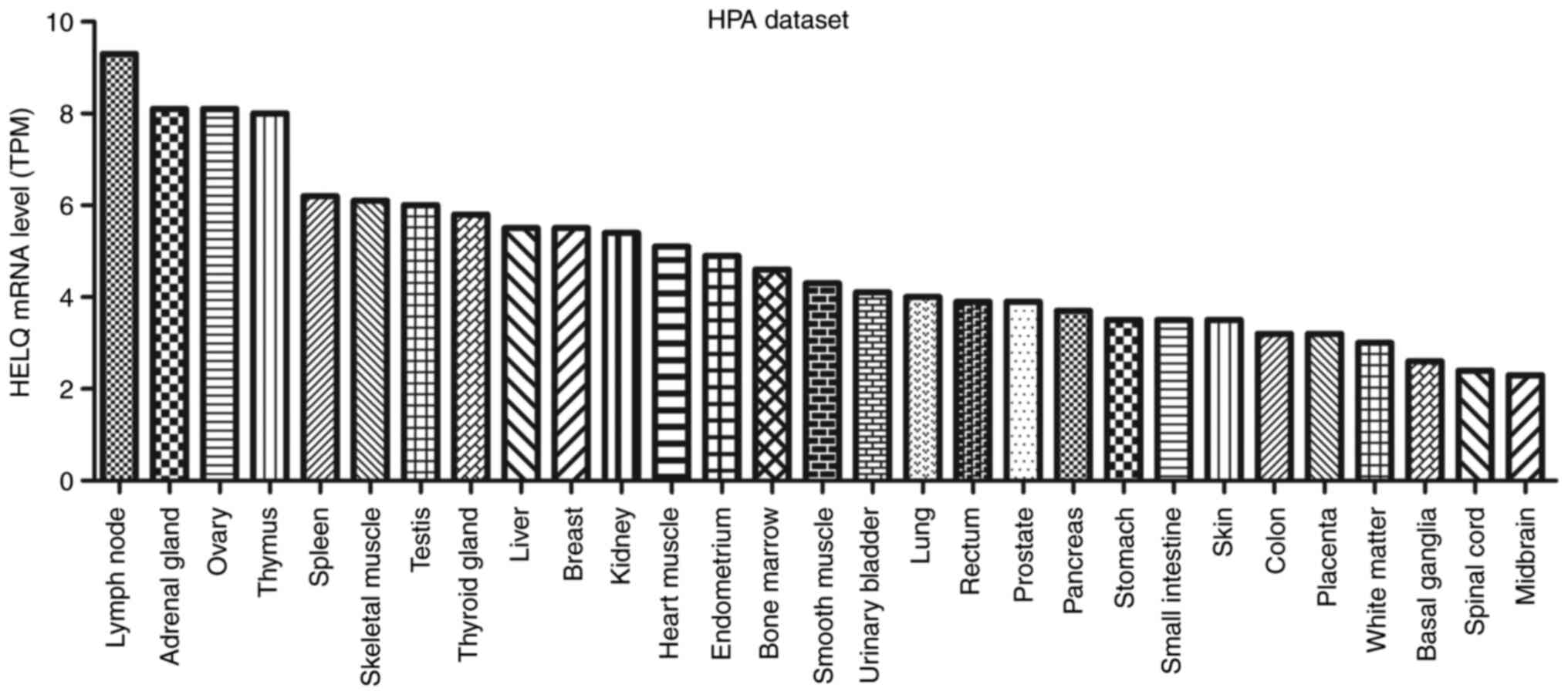
(1). HELQ is expressed at different
levels in various tissues, including the testes, ovaries, skeletal
muscle and heart (75) in humans,
and has also been detected in several other tissues (https://www.proteinatlas.org/ENSG00000163312-HELQ/tissue)
(Fig. 2).
HELQ plays a crucial role in germ cell maintenance,
and its loss results in subfertility. A high level of HELQ
expression is found in the reproductive system, including the
ovaries, testes, cervix, breast, epididymis, endometrium and
prostate. In HELQ-deficient mice, HELQ facilitates RAD51
paralog-dependent repair, thereby preventing germ cell attrition
(11). It is disputed whether
mutant HELQ has an adverse effect on the male reproductive system.
HELQgt/gt male mice have significantly smaller testes,
suffer from seminiferous tubule atrophy, and lack spermatocytes and
spermatogonia, as compared with wild-type male mice, indicating
that HELQ deficiency leads to a mild form of hypogonadism (53); however, two heterozygous mutant
mouse models (HELQ+/M5 and HELQ+/M6) did not
show any spermatogenic defects, indicating that heterozygous HELQ
variants alone do not cause development of the sertoli cell-only
syndrome phenotype in mice (76).
Furthermore, Wang et al (77) found no evidence that HELQ mutations
are associated with premature ovarian failure in Chinese women.
Potential roles of HELQ in tumorigenesis and
underlying mechanisms
In addition to its physiological functions, HELQ is
often implicated in tumorigenesis. Genetic alterations, such as
deletions and mismatches, are frequently associated with
tumorigenesis. Single-nucleotide polymorphism variants of
HELQ have been associated with an increased risk of various
cancers, including upper aerodigestive tract cancers (78,79),
esophageal squamous cell carcinoma (80,81),
head and neck cancers (82),
gastric adenocarcinoma (81), and
breast and ovarian cancer (OV) (83–85)
through genome-wide association studies (Table III). Furthermore, there is
accumulating evidence indicating that HELQ may act as a tumor
suppressor for several cancers, such as osteosarcoma (86), OV (12,13,34,77,87),
chronic lymphocytic leukemia (CLL) (88), non-small cell lung cancer (NSCLC)
(89) and endometrial stromal
sarcoma (ESS) (Table IV) (90). HELQ is an important regulator of
cancer proliferation, invasion, migration and can contribute to
poor patient prognosis and platinum resistance through several
mechanisms (13,86,87) in
different types of tumors (Fig.
3).
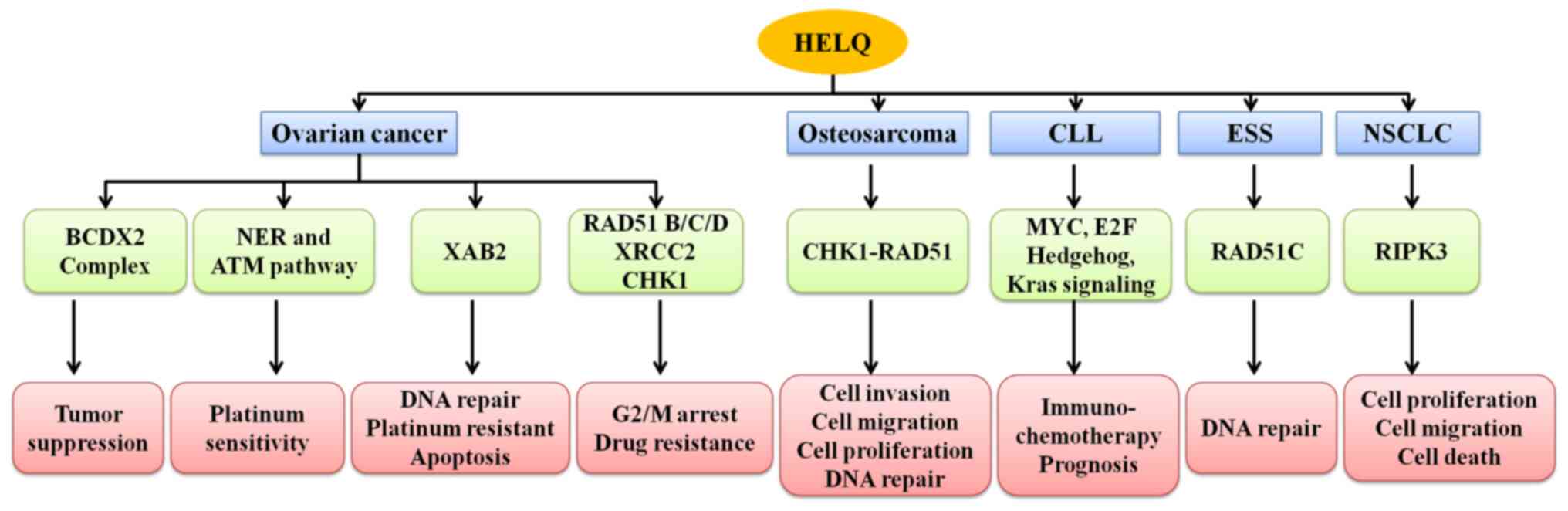 | Figure 3.Mechanisms of HELQ in tumorigenesis.
HELQ inhibits DNA repair and drug resistance, induces G2/M arrest
and apoptosis through XAB2, NER/ATM, BCDX2 pathway in ovarian
cancer. HELQ inhibits osteosarcoma cell proliferation, migration,
invasion and DNA repair by CHK1-RAD51 pathway. HELQ was associated
with improved immuno-chemotherapy response in patients with CLL
associated with activation of MYC signaling, E2F signaling, and
suppression of Hedgehog and Kras signaling. DNA repair involving
HELQ and RAD51C may participate in ESS occurrence and development.
HELQ inhibits NSCLC cell proliferation and migration through
suppressing DNA damage repair, and promotes cell death through
inducing necrosis by interacting with RIPK3. HELQ, helicase
POLQ-like; NER, nucleotide excision repair; ATM, ataxia
telangiectasia mutated protein; CHK1, checkpoint kinase 1; RAD51,
RecA-like ATP-dependent recombinase 51; CLL, chronic lymphocytic
leukemia; ESS, endometrial stromal sarcoma; NSCLC, non-small cell
lung cancer. |
| Table III.Associations between SNPs within HELQ
and cancer risk. |
Table III.
Associations between SNPs within HELQ
and cancer risk.
|
|
| Alleles |
|
|
|
|
|
|---|
|
|
|
|
|
|
|
|
|
|---|
| SNP | Genotype | Major | Minor | Reason for
replication attempt | Cancer type | Risk factors | Significant
association | (Refs.) |
|---|
| rs1494961 | / | T | C | Non-synonymous | Upper
aero-digestive tract (UADT) cancers | Age and sex | Yes,
P=1×10−8 | (78) |
| rs1494961 | / | / | C | Missense mutation
V306I |
| Smoking and/or
alcohol | Yes,
P=2.65×10−4 | (79) |
| rs1494961 | C/C + C/T and
T/T | / | / | / | Head and neck
squamous cell carcinoma | Alcohol consumption
and smoking pack-years | Yes,
P<0.001 | (82) |
| rs1494961 | TT, TC, CC | T | C | / | ESCC | Age, Smoking,
Drinking | Yes, P=0.032 | (80) |
| rs13115704 | / | T | C | / | ESCC | / | Yes,
P=8.07×10−3 | (81) |
| rs1494961 | / | T | C | / | Gastric cancer
(GC) | / | Yes, P=0.035 |
|
| rs13141136 | / | / | / | Synonymous and | Breast and | / | No, P>0.05 | (83) |
| rs7665103 |
|
|
| non-synonymous | ovarian cancer |
|
|
|
| rs141700135 |
|
|
| missense
variants |
|
|
|
|
| rs138939487 |
|
|
|
|
|
|
|
|
| rs1494961 |
|
|
|
|
|
|
|
|
| Rs11099601 |
| A | G | / | Breast cancer | / | Yes,
P=5.62×10−6 | (84) |
| Rs4693089 | GG GA/AA |
| G | / | Breast cancer | / | Yes,
P=2.38×10−19 | (85) |
| Table IV.Summary of the current literature on
HELQ dysregulation in different cancers. |
Table IV.
Summary of the current literature on
HELQ dysregulation in different cancers.
| Cancer | Deregulation | Downstream
targets | Phenotypic
effect | Studies | (Refs.) |
|---|
| Osteosarcoma | Low | CHK1↓, RAD51↓ | DNA repair↓, cell
invasion↑, cell migration↑, cell proliferation↑ | In
vitro | (86) |
| CLL | Low (CLL vs. normal
B cells) | MYC↓, E2F1↓, DNA
repair pathway↓, Hedgehog signaling↑, Kras signaling pathway↑ | response to
immuno-chemotherapy ↓, Richter transformation↓, unfavorable OS | GEO dataset | (88) |
| Ovarian cancer | Loss/mutation | RAD51B/C/D↓,
XRCC2↓, CHK1↓ | G2/M arrest↓,
cellular sensitivity↑, chromosome radial formation↑ | In
vitro | (12) |
| High-grade serous
ovarian cancer | High in platinum
resistance | XAB2↑ | Poor prognosis, DNA
damage repair↑, platinum resistance↑, apoptosis↓ | In vitro and
TCGA database | (87) |
| Cisplatin-resistant
ovarian cancer | High in
cisplatin-resistant | NER pathway
proteins (RPA32, RAP70, XPA, XPC, ERCC1)↑, ATM/ATR pathway
protein↑ | The cellular
resistance to cisplation↑, DNA repair activity↑, Nucleotide
excision repair pathway↑ | In
vitro | (13) |
| Non-small cell lung
cancer | Low | RIPK3↓ | Cell
proliferation↑, cell migration↑, chemotactic response↑, DNA damage
repair↓, cell necrosis↓, cell death↓ | In vitro and
in vivo | (89) |
| Endometrial stromal
sarcoma | Low | RAD51C↓ | DNA repair↓ | Clinical
tissues | (90) |
Osteosarcoma
HELQ plays a critical role in tumor suppression in
mammals through interacting with the BCDX2 complex (11). HELQ overexpression inhibited
osteosarcoma cell proliferation, migration, invasion and DNA damage
repair (86). In addition, the
antitumor activity of HELQ may be associated with the upregulation
of the DNA damage-related proteins RAD51 and CHK1expression, and
HELQ modulates an anti-invasive phenotype and DNA damage repair in
osteosarcoma cells by activation of the CHK1-RAD51 signaling
pathway (86).
NSCLC
HELQ is downregulated in NSCLC tissues and cells,
while the malignancy of lung cancer cells was enhanced by HELQ
depletion. HELQ overexpression inhibits cell proliferation and
migration by suppressing DNA damage repair, and promotes cell death
by inducing necrosis through its interaction with
receptor-interacting serine/threonine kinase 3 (RIPK3) (89). HELQ is a favorable prognostic factor
for patients with NSCLC, and low HELQ levels in patients with NSCLC
were associated with a reduced overall survival (OS) through a
Kaplan-Meier plotter.
ESS
HELQ and RAD51C expression levels are decreased in
ESS compared with normal endometrial tissues. HELQ expression was
found to be correlated with the size and type of ESS. Neither HELQ
nor RAD51C expression were correlated with age, FIGO stage or lymph
node metastasis status. The occurrence and development of ESS may
be affected by DNA repair involving HELQ and RAD51C (90).
OV
In addition, HELQ plays a role in cellular
resistance in response to ICLs and G2/M arrest, through its
association with the RAD51 paralogs RAD51B/C/D and XRCC2, and
promotes the activation of the ATR substrate, CHK1 (12). HELQ and XPA binding protein 2 are
associated with platinum resistance, poor prognosis, decreased
apoptosis and increased DNA damage repair in ascites from
high-grade serous OV cells (87).
In addition, HELQ decreases cisplatin sensitivity in OV cells by
activating the NER and ATM/ATR pathways (13). OV patients with HELQ-low expression
had improved overall and disease-free survival than those with a
high HELQ level. There is evidence that HELQ can be used
independently as a prognostic marker to predict survival in
patients with OV (13).
CLL
CLL patients with low HELQ levels exhibited a
significantly unfavorable OS compared with patients with high HELQ
levels. HELQ may be useful in predicting patients at high risk for
CLL based on the Richter transformation. Higher HELQ expression was
also associated with an improved response to immuno-chemotherapy in
patients with CLL. HELQ represented a prognostic marker for CLL
associated with the activation of MYC signaling, E2 factor
signaling and DNA repair pathways, as well as the suppression of
Hedgehog and Kras signaling (88).
Pan-cancer analysis of HELQ
Pan-cancer analysis of HELQ was conducted using The
Cancer Genome Atlas (TCGA) (https://tcga-data.nci.nih.gov/tcga/) and
Genotype-Tissue Expression (GTEx) project (https://gtexportal.org/) datasets. In TCGA dataset,
HELQ expression was significantly lower in bladder urothelial
carcinoma (BLCA), BRCA, cervical squamous cell carcinoma and
endocervical adenocarcinoma (CESC), glioblastoma multiforme (GBM),
colon adenocarcinoma (COAD), kidney chromophobe (KICH), kidney
renal clear cell carcinoma (KIRC), kidney renal papillary cell
carcinoma (KIRP), lung squamous cell carcinoma (LUSC), lung
adenocarcinoma (LUAD), pancreatic adenocarcinoma (PAAD),
pheochromocytoma and paraganglioma (PCPG), rectum adenocarcinoma
(READ), prostate adenocarcinoma (PRAD), stomach adenocarcinoma
(STAD), thyroid carcinoma (THCA), and uterine corpus endometrial
carcinoma (UCEC) than that in normal tissues, while HELQ expression
in cholangiocarcinoma (CHOL), esophageal carcinoma (ESCA), head and
neck squamous cell carcinoma (HNSC) and liver hepatocellular
carcinoma (LIHC) were significantly higher (Fig. 4A). In combined TCGA and GTEx
datasets, HELQ expression levels in adrenocortical carcinoma, BRCA,
BLCA, COAD, CESC, ESCA, KIRC, KICH, KIRP, LUSC, LUAD, PAAD, OV,
PRAD, PCPG, READ, THCA, testicular germ cell tumors, uterine
carcinosarcoma and UCEC were significantly lower than those in
normal tissues, while in CHOL, GBM, lymphoid neoplasm diffuse large
B-cell lymphoma, HNSC, LIHC, brain lower grade glioma, PAAD, STAD,
skin cutaneous melanoma and thymoma were significantly higher
(Fig. 4B).
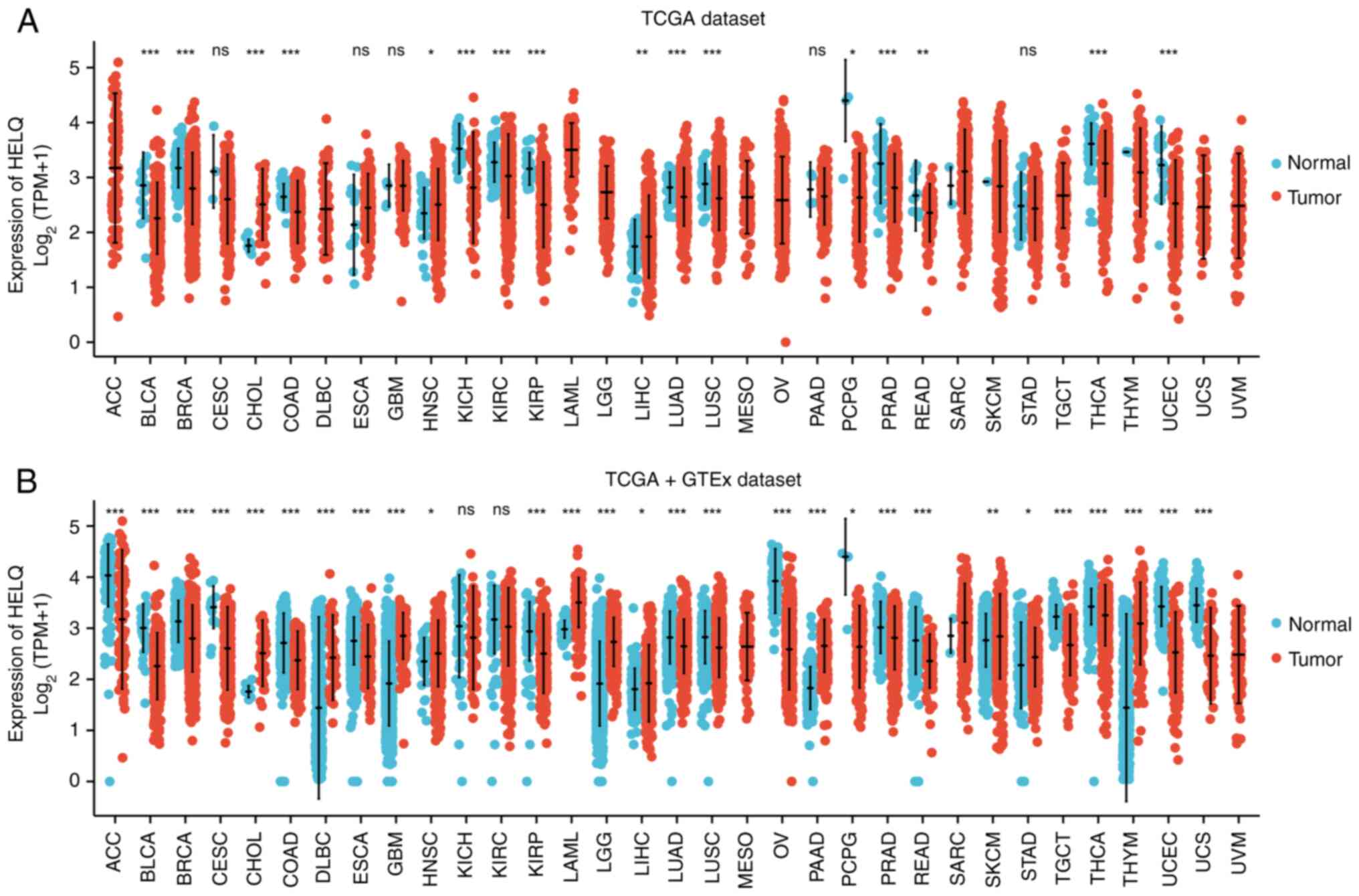 | Figure 4.Pan-cancer analysis of HELQ
expression in (A) TCGA dataset and (B) TCGA + GTEx combined
dataset. (A) The expression of HELQ in BLCA, BRCA, CESC, GBM, COAD,
KICH, KIRC, KIRP, LUSC, LUAD, PAAD, PCPG, READ, PRAD, STAD, THCA
and UCEC cancer types was significantly lower than that in normal
tissues, while levels in CHOL, ESCA, HNSC and LIHC were
significantly higher in TCGA dataset. (B) HELQ expression levels in
ACC, BRCA, BLCA, COAD, CESC, ESCA, KIRC, KICH, KIRP, LUSC, LUAD,
PAAD, OV, PRAD, PCPG, READ, THCA, TGCT, UCS and UCEC were
significantly lower than those in normal tissues, while levels in
CHOL, GBM, DLBC, HNSC, LIHC, LGG, PAAD, STAD, SKCM and THYM were
significantly higher in combined TCGA and GTEx datasets. HELQ,
helicase POLQ-like; TCGA, The Cancer Genome Atlas; GTEx, BLCA,
bladder urothelial carcinoma; BRCA: breast invasive carcinoma;
CESC, cervical squamous cell carcinoma and endocervical
adenocarcinoma; GBM, glioblastoma multiforme; COAD, colon
adenocarcinoma; KICH, kidney chromophobe; KIRC, kidney renal clear
cell carcinoma; KIRP, kidney renal papillary cell carcinoma; LUSC,
lung squamous cell carcinoma; LUAD, lung adenocarcinoma; PAAD,
pancreatic adenocarcinoma; PCPG, pheochromocytoma and
paraganglioma; READ, rectum adenocarcinoma; PRAD: prostate
adenocarcinoma; STAD, stomach adenocarcinoma; THCA: thyroid
carcinoma; UCEC, uterine carcinosarcoma; CHOL: cholangiocarcinoma;
ESCA, esophageal carcinoma; HNSC, head and neck squamous cell
carcinoma; LIHC, liver hepatocellular carcinoma; OV, ovarian serous
cystadenocarcinoma; TGCT, testicular germ cell tumors; DLBC,
lymphoid neoplasm diffuse large B-cell lymphoma; SKCM: skin
cutaneous melanoma; THYM, thymoma. *P<0.05, **P<0.01,
***P<0.001. |
Targeting HELQ for potential treatment
HELQ, a superfamily II DNA helicase, is a tumor
suppressor that can mediate tumor-inhibiting activity. Furthermore,
HELQ is pivotal in tumor proliferation, metastasis, platinum
resistance, cell-cycle checkpoint regulation and DNA damage
response. These important roles of HELQ in tumorigenesis highlight
its potential as a target for the development of novel cancer
therapeutics. Nevertheless, a few small compounds or drugs that
target HELQ have been identified for potential treatment (Table V). Kras signaling is negatively
associated with HELQ expression in patients with CLL, and
ulixertinib, a Kras signaling inhibitor, may offer a new
therapeutic option for patients with HELQ-low CLL (88,91).
Furthermore, topotecan, a topoisomerase I inhibitor, is sensitive
in HELQ mutants that induce single-ended DNA DSBs in replicating
cells (92). In additionally, a
RAD51 inhibitor, Cam833, which disrupts the interaction between
RAD51 and BRCA2, synergizes with the poly (ADP-ribose) polymerase
(PARP) inhibitors (93). HR
efficacy was reduced by 50–60% in HELQ knockout mice with increased
sensitivity to PARP inhibitors (11). It was hypothesized that PARP
inhibitors may also play a role in patients with HELQ deficiency,
but further studies are required to test this hypothesis.
HELQ-deficient mice and cells show hypersensitivity to the ICL
agent MMC through more chromatid breaks and radial chromosomes
(11). HELQ-deficient cells were
also hypersensitive to CPT, a replication inhibiting agent
(11).
| Table V.Description of HELQ as therapeutic
target at present. |
Table V.
Description of HELQ as therapeutic
target at present.
| Compound |
Drugs/Inhibitors | Target HELQ | Mechanism | (Refs.) |
|---|
| Ulixertinib | A Kras signaling
inhibitor | HELQ-low CLL | Inhibit the Kras
signaling, negatively associated HELQ | (88) |
| Topotecan | A topoisomerase I
inhibitor | HELQ mutants | induce one-ended
DNA DSBs | (92) |
| Cam833 | A RAD51
inhibitor | HELQ
deficiency | Prevent
RAD51-mediated homologous recombination | (93) |
| PARPi | A PARP
inhibitor | HELQ
deficiency | NA | (11) |
| Mitomycin C | ICL agent | HELQ
deficiency | Inhibit
replication | (11) |
| Camptothecin | Replication
blocking agent | HELQ
deficiency | Inhibit
replication | (11) |
Conclusions
HELQ is a DNA helicase with multiple biological
functions under normal and pathological conditions. The diverse
functions of HELQ, including DNA binding, DNA unwinding and DNA
repair, among others, involve three HELQ protein domains: a
DEAD/DEAH box helicase domain, a helicase C-terminal domain, and a
HTH (HTH_61) domain. The DEAD/DEAH box helicase domain is
responsible for ATP binding-mediated RNA metabolism, the helicase
C-terminal domain is involved in ATP-dependent DNA or RNA
unwinding, and the HTH domain is important for DNA strand binding
and protein interactions. Under normal conditions, HELQ is
abundantly expressed in healthy human tissues, especially in the
reproductive system, where it plays an important role in
maintaining germ cell viability through RAD51 paralog-dependent
repair. In addition, recent studies have reported that HELQ is
abnormally expressed in various cancers (86–90).
HELQ is important for cell proliferation, metastasis and
chemotherapy resistance by regulating various molecular signaling
pathways (for example, the NER, CHK1-RAD51 and ATM/ATR pathways).
HELQ deficiency is also associated with the clinical
characteristics and prognosis of patients with cancer, and is
considered a novel prognostic biomarker and potentially critical
target in cancer therapy.
Of note, multiple biological processes involving
HELQ are associated with tumorigenesis, and the protein plays a
particular role in the reproductive system. Therefore, further
studies focusing on the physiological roles of HELQ, as well as the
detailed mechanisms underlying the relationship between HELQ and
tumor occurrence under different pathological conditions, are
warranted.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National Natural Science
Foundation of China (grant no. 81902802), the National Science
Foundation of Guangdong (grant no. 2019A1515010633), the Guangzhou
Science and Technology Project (grant nos. 202102010060,
202102020766 and 202201010020), and the Guangzhou Science and
Technology Project of Traditional Chinese Medicine and Integrated
Traditional and Western Medicine (grant no. 20232A011007).
Availability of data and materials
Not applicable.
Authors' contributions
XX and NT wrote original draft preparation. NT and
WW prepared figures and tables; ZL and YW wrote review and
revision; NT, XX, ZL and YW were responsible for conceptual design;
YW carried out supervision and management. All authors have read
and approved the final version of the manuscript. Data
authentication is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
ATM
|
ataxia telangiectasia mutated
protein
|
|
ATR
|
ataxia telangiectasia and
Rad3-related protein
|
|
BLCA
|
bladder urothelial carcinoma
|
|
BRCA
|
breast invasive carcinoma
|
|
CESC
|
cervical squamous cell carcinoma and
endocervical adenocarcinoma
|
|
CHK1
|
checkpoint kinase 1
|
|
CHOL
|
cholangiocarcinoma
|
|
COAD
|
colon adenocarcinoma
|
|
CPT
|
camptothecin
|
|
dsDNA
|
double-stranded DNA
|
|
DSBs
|
DNA double-stranded breaks
|
|
ESCA
|
esophageal carcinoma
|
|
ESS
|
endometrial stromal sarcoma
|
|
FA
|
Fanconi anemia
|
|
FANCD2
|
Fanconi anemia group D2
|
|
GBM
|
glioblastoma multiforme
|
|
GTEx
|
genotype-tissue expression
|
|
HELQ
|
helicase POLQ-like
|
|
HelicC
|
helicase C-terminal domain
|
|
HNSC
|
head and neck squamous cell
carcinoma
|
|
HR
|
homologous recombination
|
|
HROB
|
homologous recombination factor with
OB-fold
|
|
HTH
|
helix-turn-helix
|
|
ICL
|
inter-strand cross-link
|
|
KICH
|
kidney chromophobe
|
|
KIRC
|
kidney renal clear cell carcinoma
|
|
KIRP
|
kidney renal papillary cell
carcinoma
|
|
LIHC
|
liver hepatocellular carcinoma
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
MCM
|
mini-chromosome maintenance
|
|
MMEJ
|
microhomology mediated end
joining
|
|
NER
|
nucleotide excision repair
|
|
NHEJ
|
non-homologous end joining
|
|
OS
|
overall survival
|
|
OV
|
ovarian cancer
|
|
PAAD
|
pancreatic adenocarcinoma
|
|
PARP
|
poly ADP ribose polymerase
|
|
PCPG
|
pheochromocytoma and
paraganglioma
|
|
PRAD
|
prostate adenocarcinoma
|
|
RAD51
|
RecA-like ATP-dependent recombinase
51
|
|
READ
|
rectum adenocarcinoma
|
|
RIPK3
|
receptor-interacting serine/threonine
kinase 3
|
|
RPA
|
replication protein A
|
|
SF2
|
superfamily-2
|
|
SSA
|
single strand annealing
|
|
STAD
|
stomach adenocarcinoma
|
|
ssDNA
|
single-stranded DNA
|
|
TCGA
|
The Cancer Genome Atlas
|
|
THCA
|
thyroid carcinoma
|
|
UCEC
|
uterine corpus endometrial
carcinoma
|
References
|
1
|
Marini F and Wood RD: A human DNA helicase
homologous to the DNA cross-link sensitivity protein Mus308. J Biol
Chem. 277:8716–8723. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Richards JD, Johnson KA, Liu H, McRobbie
AM, McMahon S, Oke M, Carter L, Naismith JH and White MF: Structure
of the DNA repair helicase hel308 reveals DNA binding and
autoinhibitory domains. J Biol Chem. 283:5118–5126. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tafel AA, Wu L and McHugh PJ: Human HEL308
localizes to damaged replication forks and unwinds lagging strand
structures. J Biol Chem. 286:15832–15840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Woodman IL and Bolt EL: Winged helix
domains with unknown function in Hel308 and related helicases.
Biochem Soc Trans. 39:140–144. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Singleton MR, Dillingham MS and Wigley DB:
Structure and mechanism of helicases and nucleic acid translocases.
Ann Rev Biochem. 76:23–50. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fairman-Williams ME, Guenther UP and
Jankowsky E: SF1 and SF2 helicases: Family matters. Curr Opin
Struct Biol. 20:313–324. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bleichert F and Baserga SJ: The long
unwinding road of RNA helicases. Mol Cell. 27:339–352. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Woodman IL, Briggs GS and Bolt EL:
Archaeal Hel308 domain V couples DNA binding to ATP hydrolysis and
positions DNA for unwinding over the helicase ratchet. J Mol Biol.
374:1139–1144. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Anand R, Buechelmaier E, Belan O, Newton
M, Vancevska A, Kaczmarczyk A, Takaki T, Rueda DS, Powell SN and
Boulton SJ: HELQ is a dual-function DSB repair enzyme modulated by
RPA and RAD51. Nature. 601:268–273. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Guy CP and Bolt EL: Archaeal Hel308
helicase targets replication forks in vivo and in vitro and unwinds
lagging strands. Nucleic Acids Res. 33:3678–3690. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Adelman CA, Lolo RL, Birkbak NJ, Murina O,
Matsuzaki K, Horejsi Z, Parmar K, Borel V, Skehel JM, Stamp G, et
al: HELQ promotes RAD51 paralogue-dependent repair to avert germ
cell loss and tumorigenesis. Nature. 502:381–384. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Takata K, Reh S, Tomida J, Person MD and
Wood RD: Human DNA helicase HELQ participates in DNA interstrand
crosslink tolerance with ATR and RAD51 paralogs. Nat Commun.
4:23382013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Long J, Zhu JY, Liu YB, Fu K, Tian Y, Li
PY, Yang WQ, Yang SY, Yin JY, Yin G and Zhang Y: Helicase POLQ-like
(HELQ) as a novel indicator of platinum-based chemoresistance for
epithelial ovarian cancer. Gynecol Oncol. 149:341–349. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Song X, Ni J and Shen Y: Structure-based
genetic analysis of Hel308a in the Hyperthermophilic Archaeon
Sulfolobus islandicus. J Genet Genomics. 43:405–413. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Gorbalenya AE, Koonin EV, Donchenko AP and
Blinov VM: Two related superfamilies of putative helicases involved
in replication, recombination, repair and expression of DNA and RNA
genomes. Nucleic Acids Res. 17:4713–4730. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Caruthers JM and McKay DB: Helicase
structure and mechanism. Curr Opin Struct Biol. 12:123–133. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pyle AM: Translocation and unwinding
mechanisms of RNA and DNA helicases. Ann Rev Biophys. 37:317–336.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jankowsky E: RNA helicases at work:
Binding and rearranging. Trends Biochem Sci. 36:19–29. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Johnson SJ and Jackson RN: Ski2-like RNA
helicase structures: Common themes and complex assemblies. RNA
Biol. 10:33–43. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bennett RJ and Keck JL: Structure and
function of RecQ DNA helicases. Crit Rev Biochem Mol Biol.
39:79–97. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fuller-Pace FV: DExD/H box RNA helicases:
Multifunctional proteins with important roles in transcriptional
regulation. Nucleic Acids Res. 34:4206–4215. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lusser A and Kadonaga JT: Chromatin
remodeling by ATP-dependent molecular machines. Bioessays.
25:1192–1200. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jankowsky E and Fairman ME: RNA
helicases-one fold for many functions. Curr Opin Struct Biol.
17:316–324. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Jankowsky E and Bowers H: Remodeling of
ribonucleoprotein complexes with DExH/D RNA helicases. Nucleic
Acids Res. 34:4181–4188. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Byrd AK and Raney KD: Superfamily 2
helicases. Front Bioscience. 17:2070–2088. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Han X, Zhao L and Li X: HELQ in cancer and
reproduction. Neoplasma. 63:825–835. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Cordin O, Banroques J, Tanner NK and
Linder P: The DEAD-box protein family of RNA helicases. Gene.
367:17–37. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Heung LJ and Del Poeta M: Unlocking the
DEAD-box: A key to cryptococcal virulence? J Clin Invest.
115:593–595. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
de la Cruz J, Kressler D and Linder P:
Unwinding RNA in Saccharomyces cerevisiae: DEAD-box proteins and
related families. Trends Biochem Sci. 24:192–198. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Aubourg S, Kreis M and Lecharny A: The
DEAD box RNA helicase family in Arabidopsis thaliana. Nucleic Acids
Res. 27:628–636. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tanner NK and Linder P: DExD/H box RNA
helicases: From generic motors to specific dissociation functions.
Mol Cell. 8:251–262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huffman JL and Brennan RG: Prokaryotic
transcription regulators: More than just the helix-turn-helix
motif. Curr Opin Struct Biol. 12:98–106. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kitano K, Kim SY and Hakoshima T:
Structural basis for DNA strand separation by the unconventional
winged-helix domain of RecQ helicase WRN. Structure. 18:177–187.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li YP, Yang JJ, Xu H, Guo EY and Yu Y:
Structure-function analysis of DNA helicase HELQ: A new diagnostic
marker in ovarian cancer. Oncol Lett. 12:4439–4444. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Jenkins T, Northall SJ, Ptchelkine D,
Lever R, Cubbon A, Betts H, Taresco V, Cooper CDO, McHugh PJ,
Soultanas P and Bolt EL: The HelQ human DNA repair helicase
utilizes a PWI-like domain for DNA loading through interaction with
RPA, triggering DNA unwinding by the HelQ helicase core. NAR
Cancer. 3:zcaa0432021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Gaudet P, Livstone MS, Lewis SE and Thomas
PD: Phylogenetic-based propagation of functional annotations within
the Gene Ontology consortium. Brief Bioinform. 12:449–462. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Moldovan GL, Madhavan MV, Mirchandani KD,
McCaffrey RM, Vinciguerra P and D'Andrea AD: DNA polymerase POLN
participates in cross-link repair and homologous recombination. Mol
Cell Biol. 30:1088–1096. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu L and Hickson ID: DNA helicases
required for homologous recombination and repair of damaged
replication forks. Ann Rev Genet. 40:279–306. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Sancar A, Lindsey-Boltz LA, Unsal-Kacmaz K
and Linn S: Molecular mechanisms of mammalian DNA repair and the
DNA damage checkpoints. Ann Rev Biochem. 73:39–85. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Li X and Heyer WD: Homologous
recombination in DNA repair and DNA damage tolerance. Cell Res.
18:99–113. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Roy S: Maintenance of genome stability in
plants: Repairing DNA double strand breaks and chromatin structure
stability. Front Plant Sci. 5:4872014. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Martin LP, Hamilton TC and Schilder RJ:
Platinum resistance: The role of DNA repair pathways. Clin Cancer
Res. 14:1291–1295. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Kamp JA, Lemmens B, Romeijn RJ, Changoer
SC, van Schendel R and Tijsterman M: Helicase Q promotes
homology-driven DNA double-strand break repair and prevents tandem
duplications. Nat Commun. 12:71262021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hoeijmakers JH: Genome maintenance
mechanisms for preventing cancer. Nature. 411:366–374. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Scully R, Panday A, Elango R and Willis
NA: DNA double-strand break repair-pathway choice in somatic
mammalian cells. Nat Rev Mol Cell Biol. 20:698–714. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gartner A and Engebrecht J: DNA repair,
recombination, and damage signaling. Genetics. 220:iyab1782022.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Ward JD, Muzzini DM, Petalcorin MI,
Martinez-Perez E, Martin JS, Plevani P, Cassata G, Marini F and
Boulton SJ: Overlapping mechanisms promote postsynaptic RAD-51
filament disassembly during meiotic double-strand break repair. Mol
Cell. 37:259–272. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
San Filippo J, Sung P and Klein H:
Mechanism of eukaryotic homologous recombination. Ann Rev Biochem.
77:229–257. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Hustedt N, Saito Y, Zimmermann M,
Alvarez-Quilon A, Setiaputra D, Adam S, McEwan A, Yuan JY, Olivieri
M, Zhao Y, et al: Control of homologous recombination by the
HROB-MCM8-MCM9 pathway. Genes Dev. 33:1397–1415. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Niedernhofer LJ, Daniels JS, Rouzer CA,
Greene RE and Marnett LJ: Malondialdehyde, a product of lipid
peroxidation, is mutagenic in human cells. J Biol Chem.
278:31426–31433. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
McHugh PJ, Spanswick VJ and Hartley JA:
Repair of DNA interstrand crosslinks: Molecular mechanisms and
clinical relevance. Lancet Oncol. 2:483–490. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Adelman CA and Boulton SJ: Metabolism of
postsynaptic recombination intermediates. FEBS Lett. 584:3709–3716.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Luebben SW, Kawabata T, Akre MK, Lee WL,
Johnson CS, O'Sullivan MG and Shima N: Helq acts in parallel to
Fancc to suppress replication-associated genome instability.
Nucleic Acids Res. 41:10283–10297. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Garcia-Higuera I, Taniguchi T, Ganesan S,
Meyn MS, Timmers C, Hejna J, Grompe M and D'Andrea AD: Interaction
of the Fanconi anemia proteins and BRCA1 in a common pathway. Mol
Cell. 7:249–262. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Northall SJ, Buckley R, Jones N, Penedo
JC, Soultanas P and Bolt EL: DNA binding and unwinding by Hel308
helicase requires dual functions of a winged helix domain. DNA
Repair. 57:125–132. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Fujikane R, Shinagawa H and Ishino Y: The
archaeal Hjm helicase has recQ-like functions, and may be involved
in repair of stalled replication fork. Genes Cells. 11:99–110.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Smith J, Tho LM, Xu N and Gillespie DA:
The ATM-Chk2 and ATR-Chk1 pathways in DNA damage signaling and
cancer. Adv Cancer Res. 108:73–112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Montano R, Thompson R, Chung I, Hou H,
Khan N and Eastman A: Sensitization of human cancer cells to
gemcitabine by the Chk1 inhibitor MK-8776: Cell cycle perturbation
and impact of administration schedule in vitro and in vivo. BMC
Cancer. 13:6042013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Krajewska M, Fehrmann RS, Schoonen PM,
Labib S, de Vries EG, Franke L and van Vugt MA: ATR inhibition
preferentially targets homologous recombination-deficient tumor
cells. Oncogene. 34:3474–3481. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Bagby GC Jr: Genetic basis of Fanconi
anemia. Curr Opin Hematol. 10:68–76. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Seki S, Ohzeki M, Uchida A, Hirano S,
Matsushita N, Kitao H, Oda T, Yamashita T, Kashihara N, Tsubahara
A, et al: A requirement of FancL and FancD2 monoubiquitination in
DNA repair. Genes Cells. 12:299–310. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Nepal M, Che R, Ma C, Zhang J and Fei P:
FANCD2 and DNA Damage. Int J Mol Sci. 18:18042017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Muzzini DM, Plevani P, Boulton SJ, Cassata
G and Marini F: Caenorhabditis elegans POLQ-1 and HEL-308 function
in two distinct DNA interstrand cross-link repair pathways. DNA
Repair. 7:941–950. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Richardson CD, Kazane KR, Feng SJ, Zelin
E, Bray NL, Schäfer AJ, Floor SN and Corn JE: CRISPR-Cas9 genome
editing in human cells occurs via the Fanconi anemia pathway. Nat
Genet. 50:1132–1139. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zellweger R, Dalcher D, Mutreja K, Berti
M, Schmid JA, Herrador R, Vindigni A and Lopes M: Rad51-mediated
replication fork reversal is a global response to genotoxic
treatments in human cells. J Cell Biol. 208:563–579. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Bhat KP and Cortez D: RPA and RAD51: Fork
reversal, fork protection, and genome stability. Nat Struct Mol
Biol. 25:446–453. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Bonilla B, Hengel SR, Grundy MK and
Bernstein KA: RAD51 gene family structure and function. Ann Rev
Genet. 54:25–46. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Somyajit K, Subramanya S and Nagaraju G:
RAD51C: A novel cancer susceptibility gene is linked to Fanconi
anemia and breast cancer. Carcinogenesis. 31:2031–2038. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Suwaki N, Klare K and Tarsounas M: RAD51
paralogs: Roles in DNA damage signalling, recombinational repair
and tumorigenesis. Seminars Cell Dev Biol. 22:898–905. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Somyajit K, Saxena S, Babu S, Mishra A and
Nagaraju G: Mammalian RAD51 paralogs protect nascent DNA at stalled
forks and mediate replication restart. Nucleic Acids Res.
43:9835–9855. 2015.PubMed/NCBI
|
|
71
|
Masson JY, Stasiak AZ, Stasiak A, Benson
FE and West SC: Complex formation by the human RAD51C and XRCC3
recombination repair proteins. Proc Natl Acad Sci USA.
98:8440–8446. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Rein HL, Bernstein KA and Baldock RA:
RAD51 paralog function in replicative DNA Damage and tolerance.
Curr Opin Genet Dev. 71:86–91. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Chen R and Wold MS: Replication protein A:
Single-stranded DNA's first responder: Dynamic DNA-interactions
allow replication protein A to direct single-strand DNA
intermediates into different pathways for synthesis or repair.
Bioessays. 36:1156–1161. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Oakley GG and Patrick SM: Replication
protein A: Directing traffic at the intersection of replication and
repair. Front Bioscience. 15:883–900. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Marini F, Kim N, Schuffert A and Wood RD:
POLN, a nuclear PolA family DNA polymerase homologous to the DNA
cross-link sensitivity protein Mus308. J Biol Chem.
278:32014–32019. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Murtaza G, Yang L, Khan I, Unar A, Khan M,
Huan Z, Khan R and Shi Q: Identification and functional
investigation of novel heterozygous HELQ mutations in patients with
Sertoli Cell-only Syndrome. Genet Test Mol Biomarkers. 25:654–659.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang W, Zhao S, Zhuang L, Li W, Qin Y and
Chen ZJ: The screening of HELQ gene in Chinese patients with
premature ovarian failure. Reprod Biomed Online. 31:573–576. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
McKay JD, Truong T, Gaborieau V, Chabrier
A, Chuang SC, Byrnes G, Zaridze D, Shangina O, Szeszenia-Dabrowska
N, Lissowska J, et al: A genome-wide association study of upper
aerodigestive tract cancers conducted within the INHANCE
consortium. PLoS Genet. 7:e10013332011. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Babron MC, Kazma R, Gaborieau V, McKay J,
Brennan P, Sarasin A and Benhamou S: Genetic variants in DNA repair
pathways and risk of upper aerodigestive tract cancers: Combined
analysis of data from two genome-wide association studies in
European populations. Carcinogenesis. 35:1523–1527. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Gao Y, He Y, Xu J, Xu L, Du J, Zhu C, Gu
H, Ma H, Hu Z, Jin G, et al: Genetic variants at 4q21, 4q23 and
12q24 are associated with esophageal squamous cell carcinoma risk
in a Chinese population. Hum Genet. 132:649–656. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Li WQ, Hu N, Hyland PL, Gao Y, Wang ZM, Yu
K, Su H, Wang CY, Wang LM, Chanock SJ, et al: Genetic variants in
DNA repair pathway genes and risk of esophageal squamous cell
carcinoma and gastric adenocarcinoma in a Chinese population.
Carcinogenesis. 34:1536–1542. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Liang C, Marsit CJ, Houseman EA, Butler R,
Nelson HH, McClean MD and Kelsey KT: Gene-environment interactions
of novel variants associated with head and neck cancer. Head Neck.
34:1111–1118. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Pelttari LM, Kinnunen L, Kiiski JI, Khan
S, Blomqvist C, Aittomaki K and Nevanlinna H: Screening of HELQ in
breast and ovarian cancer families. Fam Cancer. 15:19–23. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Hamdi Y, Soucy P, Adoue V, Michailidou K,
Canisius S, Lemacon A, Droit A, Andrulis IL, Anton-Culver H, Arndt
V, et al: Association of breast cancer risk with genetic variants
showing differential allelic expression: Identification of a novel
breast cancer susceptibility locus at 4q21. Oncotarget.
7:80140–80163. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Homer MV, Charo LM, Natarajan L,
Haunschild C, Chung K, Mao JJ, DeMichele AM and Su HI: Genetic
variants of age at menopause are not related to timing of ovarian
failure in breast cancer survivors. Menopause. 24:663–668. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Liu DN, Zhou YF, Peng AF, Long XH, Chen
XY, Liu ZL and Xia H: HELQ reverses the malignant phenotype of
osteosarcoma cells via CHK1-RAD51 signaling pathway. Oncol Rep.
37:1107–1113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Zhu F, Yang S, Lei M, He Q, Wu L and Zhang
Y: DNA repair protein HELQ and XAB2 as chemoresponse and prognosis
biomarkers in ascites tumor cells of high-grade serous ovarian
cancer. J Oncol. 2022:75219342022. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Guo C, Gao YY, Ju QQ, Zhang CX, Gong M and
Li ZL: HELQ and EGR3 expression correlate with IGHV mutation status
and prognosis in chronic lymphocytic leukemia. J Transl Med.
19:422021. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Zhong NS, Tong WL, Zhang Y, Xiao SN, Liu
JM, Li AA, Yao GL, Lin Q and Liu ZL: HELQ suppresses migration and
proliferation of non-small cell lung cancer cells by repairing DNA
damage and inducing necrosis. Cell Biol Int. 47:188–200. 2023.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Liu Y, Zhang Y and Tian Y: Expressions of
HELQ and RAD51C in endometrial stromal sarcoma and their clinical
significance. Nan Fang Yi Ke Da Xue Xue Bao. 40:936–941. 2020.(In
Chinese). PubMed/NCBI
|
|
91
|
Gimenez N, Martinez-Trillos A, Montraveta
A, Lopez-Guerra M, Rosich L, Nadeu F, Valero JG, Aymerich M,
Magnano L, Rozman M, et al: Mutations in the RAS-BRAF-MAPK-ERK
pathway define a specific subgroup of patients with adverse
clinical features and provide new therapeutic options in chronic
lymphocytic leukemia. Haematologica. 104:576–586. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Thomas A, Cox J, Wolfe KB, Mingalone CH,
Yaspan HR and McVey M: Division of Labor by the HELQ, BLM, and
FANCM Helicases during Homologous Recombination Repair in
Drosophila melanogaster. Genes (Basel). 13:4742022.
View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Scott DE, Francis-Newton NJ, Marsh ME,
Coyne AG, Fischer G, Moschetti T, Bayly AR, Sharpe TD, Haas KT,
Barber L, et al: A small-molecule inhibitor of the BRCA2-RAD51
interaction modulates RAD51 assembly and potentiates DNA
damage-induced cell death. Cell Chem Biol. 28:835–847.e5. 2021.
View Article : Google Scholar : PubMed/NCBI
|