According to the most recent figures from the
International Agency for Research on Cancer on incidence and
mortality, there will be 28.4 million new cases of cancer worldwide
in 2040, a 47% increase from 2020. Out of all cancer cases,
gastrointestinal tumors account for five of the top 10 causes of
mortality [colorectal cancer (CRC; 9.4%), liver cancer (8.3%),
stomach cancer (7.7%), esophageal cancer (5.5%), and pancreatic
cancer (4.7%)] (1). Owing to the
high metastasis and recurrence of these gastrointestinal tumors,
the 5-year overall survival rate for advanced tumors is poor
(2,3). Cancer treatment has undergone profound
changes in recent years with the continuous development of the
understanding of cancer biology at the molecular level. For
instance, a large number of targeted drugs have been approved as a
first-line treatment for numerous tumors (4). However, clinical studies revealed that
these drugs are ineffective for patients with Rictor alterations
(5). These studies suggested that
Rictor is involved in tumor resistance and may act as a therapeutic
target.
Genomic instability and mutation are important
features of cancer cells. According to The Cancer Genome Atlas
(TCGA) database (6), as determined
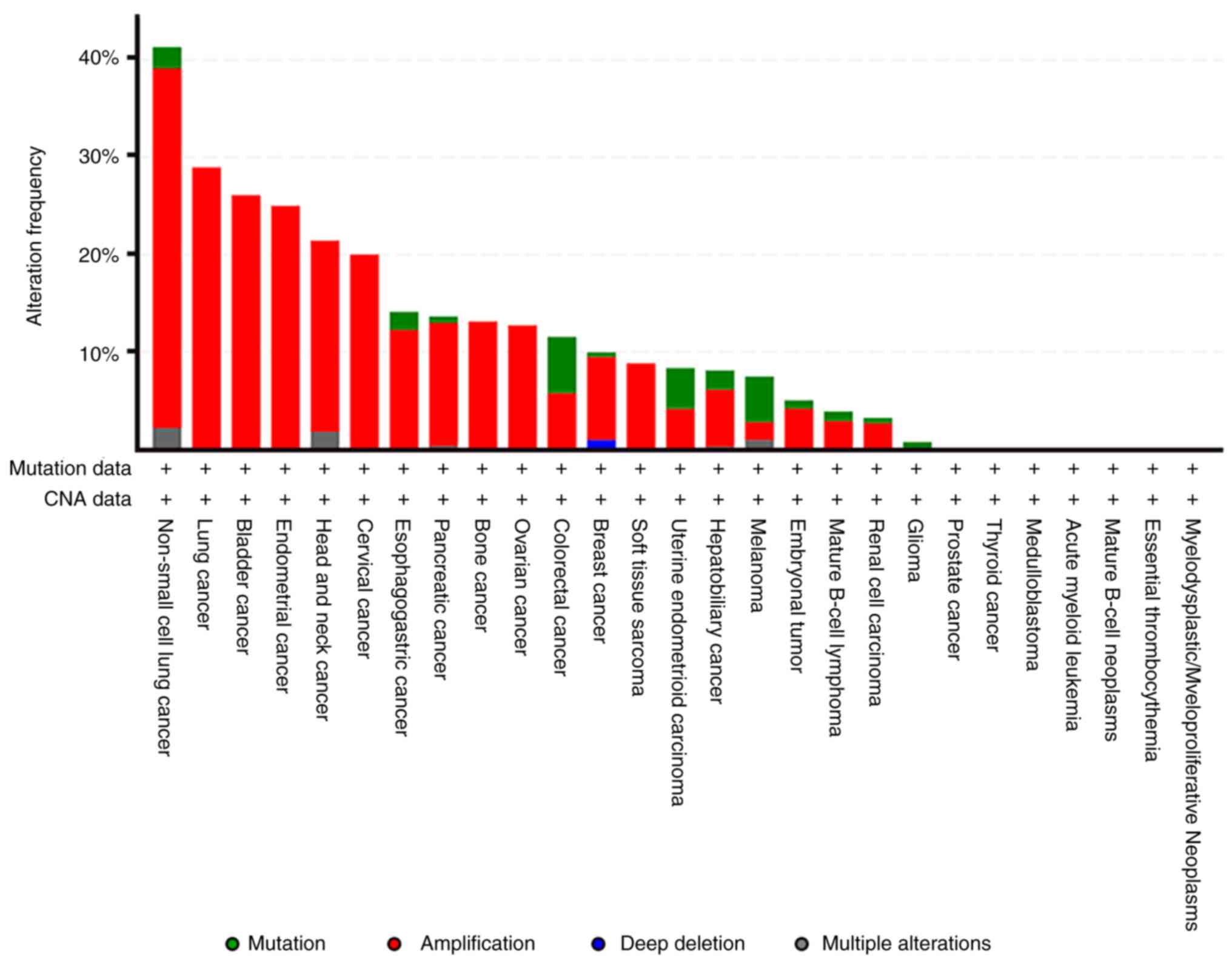
by the alterations of Rictor in respective patient samples as a
fraction of the total number of patients screened, in
non-small-cell lung cancer, 41.3% (19/46 cases) of patients had
altered Rictor levels. Similarly, the figures for altered Rictor
are 14.11% (23/163 cases) in esophagogastric cancer, 13.64% (42/308
cases) in pancreatic cancer, 11.54% (23/163 cases) in CRC and 8%
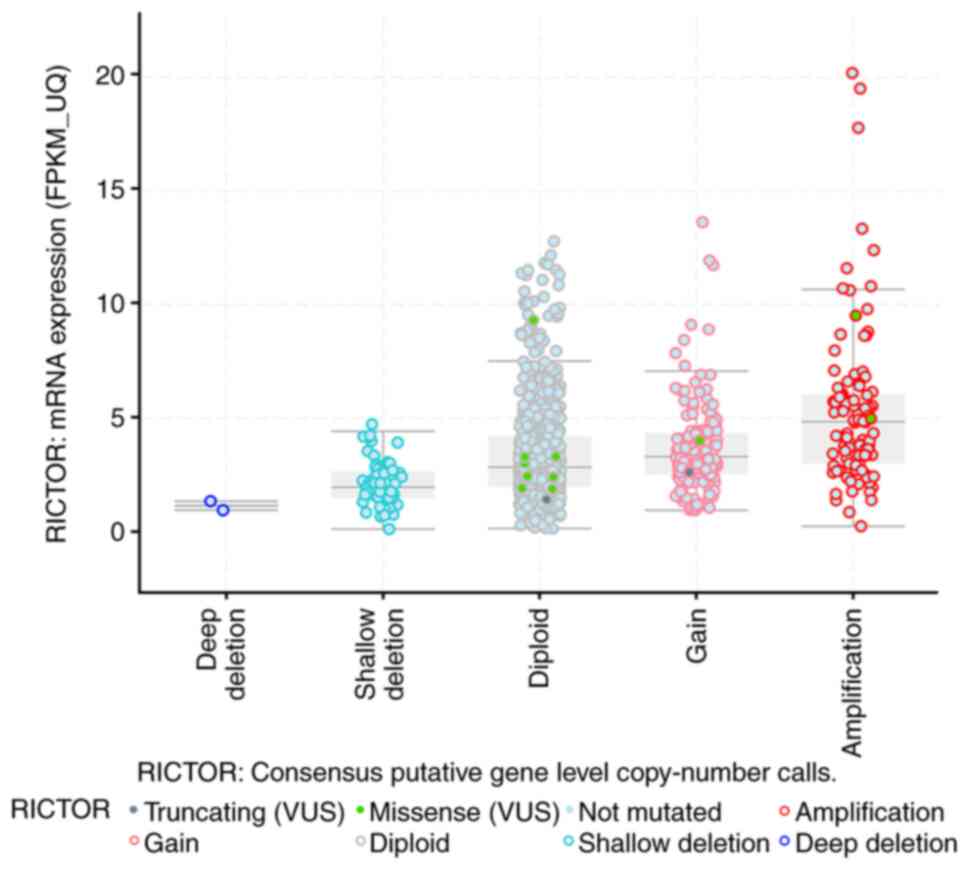
(29/358 cases) in hepatobiliary cancer (Fig. 1). Further data from the TCGA dataset
comprising 991 samples showed different types of Rictor alterations
(Fig. 2). In addition, the overall
survival rates of patients with high Rictor expression in tumor
tissues was observed to be low. For instance, Bian et al
(7) demonstrated through
immunohistochemistry that high Rictor expression is associated with
rapid tumor progression and poor prognosis in patients with gastric
cancer (GC). A study from the Southern Medical University of China
revealed that a high Rictor expression leads to poor clinical
prognosis in CRC (8). From the
analysis of 201 samples of esophageal squamous cell carcinoma
(ESCC), it was found that Rictor expression was positively
associated (P=0.011) with the cancer stage, according to the
grading by the American Joint Committee on Cancer (AJCC), and
negatively associated (P=0.007) with survival (9). These findings are sufficiently
exciting to warrant a detailed discussion of the role of Rictor in
the biology of gastrointestinal cancers.
The mTOR and its signaling pathway have important
roles in regulating protein synthesis, cell growth, apoptosis,
angiogenesis and migration. Dysfunction of the mTOR signaling
pathway is common in several human cancers (10,11).
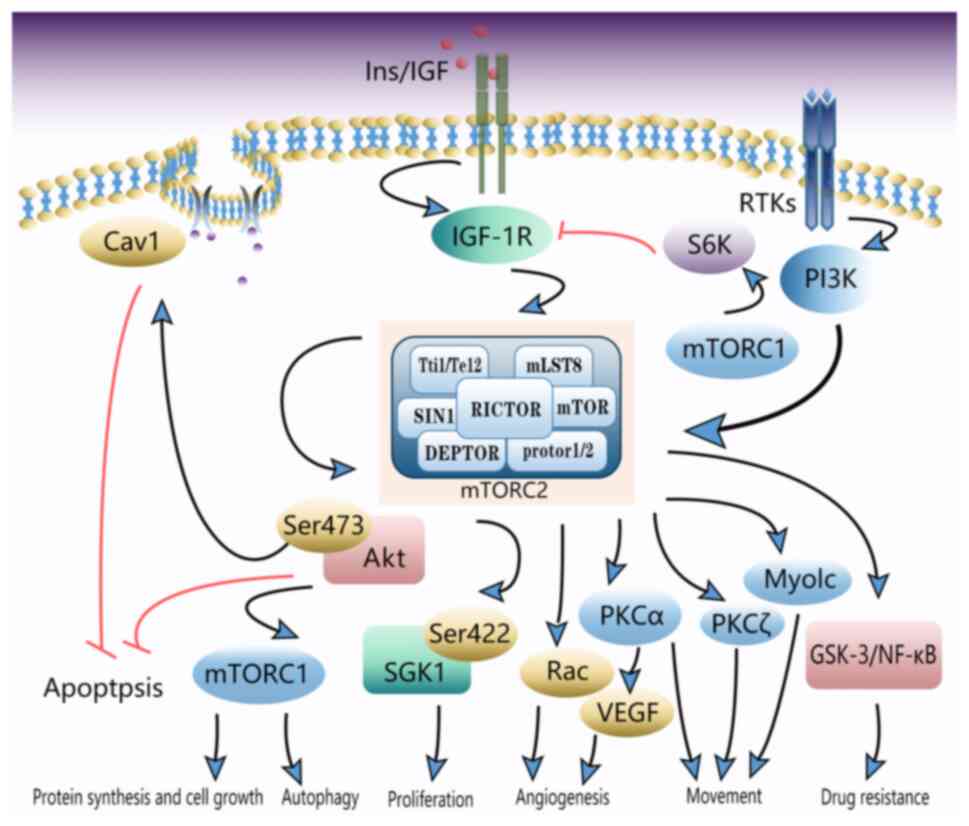
mTOR exists in two complexes: The mTOR complex 1 (mTORC1) and
mTORC2. mTORC2 consists of mTOR, mTOR-associated protein, LST8
homolog mLST8, Rictor, mSin1 and proteins associated with Rictor
1/2, which are sensitive to growth factor levels and responsible
for the regulation of cell proliferation, metabolism, survival and
cytoskeletal remodeling (12).
Rictor is a core subunit of mTORC2. The function of mTORC2 is
dependent on Rictor, which is insensitive to rapamycin.
Mutations and biological characteristics of Rictor
in tumors reported to date are presented in Fig. 3.
Eukaryotes have used autophagy as a crucial
intracellular turnover process throughout evolution. It enables
cells to keep their intracellular environment stable. However, the
influence of autophagy on specific cell functions remains
controversial. Autophagy has been linked to cell survival and death
processes under metabolic stress. Autophagy reportedly affects
tumorigenesis and treatment (15,16).
Using bioinformatics analyses, Hao et al (17) observed that Rictor was a direct
target of microRNA (miR)-let-7a. Rescue experiments in vitro
showed that miR-let-7a promoted the autophagy level by inhibiting
the expression of Rictor in GC cells. In addition, as an upstream
executor of the Akt-mTOR signaling pathway, Rictor exerted its
effect on autophagy by phosphorylating Akt and mTOR, and this
regulatory process was also mediated by miR-let-7a. miR-let-7a in
GC regulates autophagy by targeting Rictor and follows the
regulation of the Akt-mTOR signaling pathway. Seo et al
(18) reported that downregulation
of Rictor was induced after co-treatment with PP242 and curcumin in
renal cancer cells. Downregulation of Rictor increased cytosolic
calcium release from the endoplasmic reticulum, leading to
lysosomal damage in the cell, which induced autophagy. Liu et
al (19) reported that Akt is
further activated by triggering the phosphorylation of mTOR, which
regulates the growth, autophagy and apoptosis of tumor cells,
including GC cells.
The PI3K/Akt signaling pathway stimulates cell
survival and metabolism, inhibits apoptosis and regulates tumor
cell survival and proliferation. The activation of Akt depends on
the phosphorylation of PIP3 (PDK1) at Thr308 and PDK2 at Ser473,
and the phosphorylation of Ser473 promotes that of Thr308.
Sarbassov et al (20) found
that mTORC2 is PDK2 at the Ser473 site of phosphorylated Akt in
Drosophila cells. Hresko and Mueckler (21) verified the above hypothesis in
3T3-L1 cells. These studies suggested that Rictor participates in
the PI3K/Akt signaling pathway with mTORC2 and then regulates cell
survival and nutrient uptake through mTORC1 downstream of Akt, as
well as protein synthesis and cell cycle through glycogen synthase
kinase 3 (GSK-3). The PI3K/Akt/mTOR signaling pathway is frequently
altered in malignant tumors and Rictor is a key component of this
pathway (22). Resistance to the
inhibition of the adjacent PI3K pathway is usually characterized by
the feedback activation of Akt, which is related to the mechanisms
involving Rictor (23).
Serum and glucocorticoid-induced protein kinase
(SGK) is a member of the protein kinase A/protein kinase G/protein
kinase C (AGC) family and exists in three subtypes in cells: SGK1,
SGK2 and SGK3. SGK1 is usually activated by insulin or nutritional
factors and helps regulate cell nutrient uptake (24), survival, proliferation and apoptosis
(25). García-Martínez et al
(26) found that Rictor can
directly bind to SGK1 in the form of mTORC2 and phosphorylate its
Ser422 site, independent of PI3K. This finding has been verified in
293, MCF-7 and HeLa cells.
Studies have confirmed that Rictor stimulates cell
growth and proliferation by activating Akt (also known as protein
kinase B), increasing the cells' resistance to apoptosis and
promoting angiogenesis (27,28).
Rictor overexpression in GC is associated with poor prognosis. In
particular, Rictor activates caveolin 1 (Cav1) through the Akt
signaling pathway to inhibit the apoptosis of GC cells (29). Liu et al (30) reported that real-time PCR and
western blot showed that miR-153 downregulated the expression of
Rictor, and this was related to the anti-tumor effect through
increasing apoptosis and inhibiting the growth of breast cancer
cells. A recent study (27)
suggested that Rictor is a substrate for caspase-3 and is cleaved
during apoptosis. In kidney cancer cells, Rictor silencing
increases apoptosis and concomitantly enhances rasfonin-induced
autophagy (31). In ESCC, the
downregulation of Rictor expression inhibits proliferation and
migration and induces ECa-109 and EC9706 cell cycle arrest and
apoptosis (32). In vitro
experiments showed that Rictor knockdown suppressed the
proliferation, inhibited the migration and invasion, and induced
apoptosis of GC cells (33).
Rictor regulates the migration and proliferation of
vascular endothelial cells, two events that are crucial for tumor
angiogenesis. Wang et al (34) reported that Rictor deletion
drastically reduced the vascular endothelial growth factor
(VEGF)-induced proliferation and tubulogenesis of endothelial cells
in vitro by inhibiting Akt activity through PKCα
phosphorylation. Rictor/mTORC2 inhibits the prostaglandin
E2-induced proliferation and migration of vascular endothelial
cells by regulating Rac and Akt activation. The hypoxia-induced
proliferation of endothelial cells depends on the involvement of
Rictor/mTORC2 in regulating the angiogenic mimicry of melanoma
through the Akt-MMP-2/9 pathway (35).
Rictor regulates VEGF expression in addition to
controlling endothelial cell proliferation and migration (28). Guan et al (36) reported that the tumor suppressor
miR-218 specifically targets Rictor to inhibit angiogenesis in
prostate cancer, and this mechanism may be active in other cancer
tissues, including gastrointestinal cancers. mTORC2 is a key
signaling point that promotes VEGF-mediated angiogenesis in
vascular endothelial cells by regulating Akt and PKCα (37).
The actin cytoskeleton and microtubules are the
primary cellular structures that maintain cellular morphology and
stress (38). Rictor regulates
actin cytoskeleton remodeling through PKC, and PKCα is a
representative of typical PKC. Sarbassov et al (20) found that the Rictor/mTOR complex can
directly bind and phosphorylate PKCα to regulate the actin
cytoskeleton and, consequently, cell motility. Guertin et al
(39) demonstrated that Rictor
binds to PKCα and regulates its phosphorylation in Raptor-,
Rictor-, mLST8- and mTOR-knockout mice. PKCζ, a representative of
atypical PKC and has an important role in regulating actin
cytoskeletal remodeling. Rictor can directly bind to PKCζ near the
cell membrane without mTORC2 in human breast cancer. In addition,
the phosphorylation of PKCζ and its downstream F-actin binding
protein cofilin regulate actin remodeling and cell motility
(40).
Rho GTPases with a molecular weight of 21 kDa are a
family of small G proteins, including cell division control protein
42 homolog (Cdc42), Rac family small GTPase 1 (Rac1) and Ras
homolog (Rho) family member A. These proteins are responsible for
regulating actin remodeling, microtubule treadmilling and cell
migration. Rictor can maintain or enhance Rac1/Cdc42 activity by
regulating Rho GDP-dissociation inhibitor 2 (Rho GDI2), a
suppressor of Rho-GDP. Thus, Rictor regulates actin remodeling and
tumor cell motility by regulating Rho GDI2 through an
mTOR-independent pathway (41).
Rictor also regulates actin remodeling through
molecular motors, such as Myosin-1C (Myo1c). Agarwal et al
(42) found that in 3T3-L1
fibroblastic cells, Rictor can directly bind to Myo1c, form a
stable complex independent of mTORC2 and then regulate actin
reconstruction by controlling the phosphorylation of paxillin
Tyr118. This regulation and that of cell motility are not affected
by mTORC2 or PI3K inhibitors.
In patients with GC, higher expression of Rictor has
been linked to tumor growth and poor prognosis (12). mTORC1 is sensitive to rapamycin
treatment and mediates eukaryotic initiation factor 4E-binding
protein-1 (4E-BP1), as well as the phosphorylation and activation
of p70S6 ribosomal kinase (S6K) (43). In this light, treatment with
rapamycin or its analogs was observed to inhibit the mTORC1/S6K
pathway and reduce the negative feedback loop of insulin-like
growth factor-1 receptor (IGF-1R) from S6K to IGF-1, impairing
mTORC2 signaling through the complete pathway and leading to Akt
activation (44). Furthermore, the
paradoxical activation of Akt is undesirable, as it elicits drug
resistance and cell survival, both of which are harmful to the
effectiveness of mTORC1 inhibitor therapy. In other words, the mTOR
inhibitor rapamycin could inhibit mTORC1 in cancer cells and may
lead to Akt phosphorylation through mTORC2 activation. Lang et
al (45) found that rapamycin
upregulates IGF-IR and human epidermal growth factor receptor 2
(HER2) in GC and pancreatic cancer cells. Furthermore, mTORC2 has
been shown to promote the activation of IGF-IR and insulin
receptors by activating mTOR tyrosine kinase and participate in
tumorigenesis (46). Rictor
downregulation by RNA interference (RNAi) and the induction of
receptor kinase expression are mediated by Akt activation induced
by mTORC2. In addition, mTORC2 inhibition reduces the motility of
cancer cells by suppressing GSK-3/NF-κB activity (45).
More than 1.9 million new cases of CRC, including
anal cancer, and 935,000 deaths are expected in the coming years
(1). In general, CRC ranks third in
incidence and second in mortality; the higher mortality is likely
due to the development of drug resistance (47). Bellier et al (48) used methylglyoxal (MGO), a metabolite
of glycolysis that promotes tumor growth and metastasis, to induce
Akt activation and analyzed CRC resistance. The study found that
MGO induces Akt activation by regulating PI3K/mTORC2 and heat shock
protein (Hsp)27. The premise of that study was that cancers with
Kirsten rat sarcoma viral oncogene homologue (KRAS) mutations
exhibit poor response rates to therapies and that cells with
mutated KRAS under MGO stress rely on Akt for their survival,
particularly when compared to the cells with wild-type KRAS. Akt is
activated through PI3K/mTORC2 and Hsp27. An important finding was
that MGO scavengers can inhibit Akt, which may result in the
re-sensitization of KRAS-mutant cells to cetuximab. In another
study, the autophagy-related genes Beclin 1, Raptor and Rictor were
shown to be associated with the development and progression of CRC
and multidrug resistance (MDR) (49). All three genes were selected due to
their association with autophagy. Immunohistochemistry and reverse
transcription-quantitative PCR-based evaluation was performed in
279 patients with CRC. These three autophagy-related genes, as well
as light chain 3 (LC3) and MDR-1, were significantly upregulated in
CRC tissues as compared with the adjacent control tissues.
Furthermore, their expression in patients with lymph node
metastasis was higher than that in patients without. LC3 was found
to be positively correlated with Beclin 1 and Rector and negatively
correlated with Raptor and mTORC in patients with CRC. Furthermore,
it was revealed that the five-year survival rate of patients with
CRC without lymph node metastasis, positive/high expression of
Rictor, Beclin 1 and LC3, and negative Raptor and mTOR expression,
was higher than that of patients with lymph node metastasis, high
Rictor, Beclin 1 and LC3 expression, and high Raptor and mTOR
expression.
Hepatocellular carcinoma (HCC) is a leading cause of
cancer-related mortality worldwide, due to the lack of precise
therapeutic targets (50).
mTORC2/Rictor is involved in the pretumor state of HCC and
participates in the malignant transformation of liver HCC.
Reyes-Gordillo et al (51)
showed that Akt isomers are activated in alcoholic liver disease by
increasing the expression levels of mTORC2 and genes associated
with inflammation, proliferation and fibrosis. In addition, mTORC2
affects HCC tumorigenesis by regulating metabolic reprogramming.
mTORC2 triggers the synthesis of fatty acids and lipids, resulting
in liver steatosis and tumorigenesis (52). Xin et al (53) identified that Rictor interacted with
Homo sapiens actin binding LIM protein 1 (ABLIM1) and
regulated its serine phosphorylation in HCC cells. ABLIM1, as a
previously unknown phosphorylation target of Rictor, induced by
Rictor, was indicated to have an important role in controlling
actin polymerization in HCC cells. Rictor knockdown significantly
suppressed cell migration and actin polymerization.
Immunohistochemical results showed that mTORC2 is activated in 60%
of HCC cases (54). High mTORC2
expression was found to be associated with poor prognosis of
patients with HCC.
Cholangiocarcinoma (CCC) is a highly malignant
tumor. In a previous study, HCC and CCC cells were treated with
sorafenib, a multikinase inhibitor of the
RAF/extracellular-signal-regulated kinase (ERK) kinase (MEK)/ERK
pathway, to study the differences in signaling pathways among cell
lines. Sorafenib inhibited the growth of HCC cells significantly
more than that of CCC cells but minimally suppressed ERK
phosphorylation. Correspondingly, sorafenib decreased Akt
phosphorylation at Ser473 in HCC cells but increased Akt
phosphorylation at Ser473 and mTORC2 in CCC cells. Rictor
downregulation by small inhibitory RNA in RBE cells (a CCC-derived
cell line) disrupted mTORC2 and inhibited Akt phosphorylation at
Ser473, which promoted apoptosis and inhibited RBE cell
proliferation by increasing Forkhead box protein O1. Inhibition of
mTORC2 activity in the Akt/mTOR signaling pathway during sorafenib
treatment to prevent the escape of the RAF/MEK/ERK pathway may lead
to promising treatments for CCC (59).
The TCGA database shows that the mutation rate of
Rictor in patients with esophageal GC is ~10.5%. Rictor knockdown
by short hairpin RNA enhanced the inhibitory effect of LY294002 on
the in vitro proliferation, migration and colony formation
of ECa109 and EC9706 cells, which also caused cell cycle arrest and
apoptosis in these cells. Furthermore, stable knockdown of Rictor
in vivo enhanced the antitumor effect of LY294002 by
promoting apoptosis and inhibiting tumor growth (60). A previous study identified 70.6%
(142/201) Rictor positivity in ESCC samples (14). Furthermore, the American Joint
Committee on Cancer staging was found to be positively correlated
with Rictor expression and negatively associated with survival.
mTOR overexpression is common in GC. Wang et al (33) found that Rictor protein
overexpression and Rictor and Helicobacter pylori status may
have a prognostic role in GC. A previous study by our group showed
that Rictor inhibits apoptosis of GC cells by activating Cav1
through the Akt signaling pathway (29). Seo et al (18) reported that miR-let-7A regulates
autophagy by targeting Rictor in GC cells. In other words, Rictor
is involved in the autophagy of GC cells. Bian et al
(7) analyzed 396 GC tissue samples
and found that patients who were positive for Rictor and p-Akt
(Ser473) expression had lower overall and relapse-free survival
rates than those negative for Rictor expression. Rictor
amplification is also related to tumor size, invasion depth, tumor
thrombosis and tumor stage. In line with these observations,
another study showed that targeting Rictor inhibited the
proliferation and promoted the apoptosis of GC cells (12). Furthermore, Kim et al
(61) reported that AZD2014, a dual
mTORC1/2 inhibitor, significantly inhibited the proliferation of a
Rictor-amplified patient-derived cell (PDC) line. Rictor knockdown
can reverse the sensitivity of AZD2014 to PDCs. These results
supported the need for further preclinical and clinical
investigations with AZD2014 in Rictor-amplified GC and highlighted
the importance of genomic profiling.
Pancreatic cancer is a devastating disease with the
worst outcomes among human cancers (62). Rictor blockers reportedly inhibit
tumor growth by reducing AGC kinase activation and
hypoxia-inducible factor 1-α and VEGF-A expression (63). Everolimus, a Food and Drug
Administration-approved mTOR inhibitor, can act in conjunction with
KPT-9274, a dual inhibitor of p21-activated kinase 4
(PAK4)-nicotinamide phosphoribosyltransferase. In vitro
synergy with everolimus was supported by mTORC2 modification
through the downregulation of Rictor, as revealed by molecular
analysis. By inhibiting PAK4, KPT-9274 reduced β-catenin activity,
indicating the interaction between Rho GTPases and Wnt signalling
in metastatic pancreatic neuroendocrine tumors (64). Zhao et al (65) found that mTORC1 and mTORC2 have dual
but not redundant regulatory roles in acinar-to-ductal metaplasia
and early pancreatic cancer by promoting the function of the
actin-related protein 2/3 (ARP2/3) complex. The ARP2/3 complex, as
a co-effector of mTORC1 and mTORC2, bridges the gap between
oncogenic signaling and actin dynamics of pancreatic ductal
adenocarcinoma initiation. In addition, miR-155 exacerbates
impaired autophagy in pancreatic acinar cells treated with
caerulein by targeting Rictor (66). Gemcitabine in combination with the
pro-apoptotic cytokine TNF-related apoptosis-inducing ligand
inhibited the survival and induced apoptosis of pancreatic cancer
cells. This combination therapy significantly increased the levels
of the low phosphorylated form of tumor suppressor protein 4E-BP1.
This phenomenon can be attributed to the mTOR inhibition resulting
from the caspase-mediated cleavage of the Raptor and Rictor
components of mTOR (67).
A previous study showed that rapamycin, a
first-generation mTOR inhibitor, is significantly less toxic than
other drugs in the effective dose range (68). The protective effect of
5-fluorouracil-rapamycin-cyclophosphamide sequential therapy is
stronger than that of 5-fluorouracil-adriamycin-cyclophosphamide
sequential therapy in 38 mice with colon tumors. Rapamycin has been
administered for tumor treatment, but an increasing number of
studies have confirmed that it was not as successful as expected in
clinical trials, likely due to two reasons: First, mTOR complexes
have different degrees of sensitivity to rapamycin. Since rapamycin
is sensitive to mTORC1, the drug primarily inhibits the mTORC1/S6K
pathway and lowers IGF-1R, which then activates mTORC2 to activate
Akt (45). The activation of Akt
promotes cell survival and drug resistance; thus, mTORC1 inhibitor
therapy may not be beneficial. Furthermore, inhibition of mTORC2
may eliminate the adverse signaling effects of mTOR1 inhibitors.
Further studies are warranted to identify potential therapeutic
targets of mTORC2 and explore its related molecular mechanisms in
tumors (69–71).
The results of multiple clinical trials showed that
second-generation ‘rapalogs’ possess effective pharmacokinetic
properties and exert anticancer effects (72). Table
I provides a list of different types of mTORC2 inhibitors to
treat CRC (73–84), liver cancer (85–99),
gallbladder cancer (100–102), GC (61,103–106), esophageal cancer (32,107–110), pancreatic cancer (111–119) and biliary tract cancer (120–123). The therapeutic efficacy of
rapalogs may be diminished by the pro-survival feedback loops that
may be induced by the rapalogs' mTORC1-specific inhibition, such as
the PI3K-Akt and PI3K-RAS-ERK pathways. Therefore, some of the
drawbacks of rapalogs may be resolved and a higher antitumor
activity may be achieved by combination therapy or through the use
of second-generation mTOR inhibitors, such as dual mTOR/PI3K
(124) and selective mTORC1/2
inhibitors (125). However, no
particular mTORC2 inhibitor has so far been identified. Therefore,
it is critical to discover a specific medication that blocks
Rictor. According to an in vitro study, CID613034 prevents
the phosphorylation of mTORC2 substrates, such as Akt (Ser-473),
N-myc downstream-regulated gene 1 (TR-346) and PKC; however, the
phosphorylation state of the mTORC1 substrate S6K (Thr-389) or the
mTORC1-dependent negative feedback loop are unaffected (126).
Rictor, a key effector of the PI3K/Akt/mTOR pathway,
has an important role in tumor development and invasion. It causes
tumor resistance through Akt-dependent and -independent pathways,
severely limiting the efficacy of targeted drugs. Therefore, Rictor
is an important potential target for addressing drug
resistance.
Rictor/mTORC2 alterations are more frequent in a
variety of tumor types. However, the mechanisms of Rictor/oncogenic
mTORC2 remain to be further clarified. It is essential to
understand how Rictor/oncogenic mTORC2 relates to other PI3K/mTOR
signalling pathways. Currently available Rictor/mTORC2 inhibitors
are second-generation mTOR inhibitors, and their inhibitory effects
on Rictor/mTORC2 comprise dual mTOR/PI3K, selective mTORC1/2
inhibition and ATP-competitive mTOR inhibition. The impacts of the
second-generation mTOR inhibitors on gastrointestinal cancers
showed better treatment efficacy than monotherapies in in
vitro cell experiments and preclinical studies. However, no
special inhibitors of Rictor/mTORC2 have been identified (138).
At present, PI3K/mTOR inhibitors cannot serve as
effective treatment agents. The therapeutic benefits of select
molecular inhibitors may be useful if patients are classified based
on their Rictor alteration status.
Not applicable.
Funding: No funding was received.
Not applicable.
All authors contributed to the article and have read
and approved the submitted version. RC and LM reviewed the
literature and collated and analysed the information. SG and PL
conceived and designed the study. RC, SG and PL drafted the
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare they have no competing
interests.
|
1
|
Sung H, Ferlay J, Siegel RL, Laversanne M,
Soerjomataram I, Jemal A and Bray F: Global Cancer Statistics 2020:
GLOBOCAN estimates of incidence and mortality worldwide for 36
cancers in 185 countries. CA Cancer J Clin. 71:209–249. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yang L, Ying X, Liu S, Lyu G, Xu Z, Zhang
X, Li H, Li Q, Wang N and Ji J: Gastric cancer: Epidemiology, risk
factors and prevention strategies. Chin J Cancer Res. 32:695–704.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lu L, Mullins CS, Schafmayer C, Zeißig S
and Linnebacher M: A global assessment of recent trends in
gastrointestinal cancer and lifestyle-associated risk factors.
Cancer Commun (Lond). 41:1137–1151. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hou J, He Z, Liu T, Chen D, Wang B, Wen Q
and Zheng X: Evolution of molecular targeted cancer therapy:
Mechanisms of drug resistance and novel opportunities identified by
CRISPR-Cas9 Screening. Front Oncol. 12:7550532022. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhao D, Jiang M, Zhang X and Hou H: The
role of Rictor amplification in targeted therapy and drug
resistance. Mol Med. 26:202020. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bian Y, Wang Z, Xu J, Zhao W, Cao H and
Zhang Z: Elevated Rictor expression is associated with tumor
progression and poor prognosis in patients with gastric cancer.
Biochem Biophys Res Commun. 464:534–540. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang LF, Chen HJ, Yu JL, Qi J, Lin XH and
Zou ZW: Expression of Rictor and mTOR in colorectal cancer and
their clinical significance. Nan Fang Yi Ke Da Xue Xue Bao.
36:396–400. 2016.(In Chinese). PubMed/NCBI
|
|
9
|
Jiang WJ, Feng RX, Liu JT, Fan LL, Wang H
and Sun GP: RICTOR expression in esophageal squamous cell carcinoma
and its clinical significance. Med Oncol. 34:322017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Beauchamp EM and Platanias LC: The
evolution of the TOR pathway and its role in cancer. Oncogene.
32:3923–3932. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Murugan AK: mTOR: Role in cancer,
metastasis and drug resistance. Semin Cancer Biol. 59:92–111. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gaubitz C, Prouteau M, Kusmider B and
Loewith R: TORC2 structure and function. Trends Biochem Sci.
41:532–545. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sarbassov DD, Ali SM, Kim DH, Guertin DA,
Latek RR, Erdjument-Bromage H, Tempst P and Sabatini DM: Rictor, a
novel binding partner of mTOR, defines a rapamycin-insensitive and
raptor-independent pathway that regulates the cytoskeleton. Curr
Biol. 14:1296–1302. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou P, Zhang N, Nussinov R and Ma B:
Defining the domain arrangement of the mammalian target of
rapamycin complex component rictor protein. J Comput Biol.
22:876–886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang MC, Wu AG, Huang YZ, Shao GL, Ji SF,
Wang RW, Yuan HJ, Fan XL, Zheng LH and Jiao QL: Autophagic
regulation of cell growth by altered expression of Beclin 1 in
triple-negative breast cancer. Int J Clin Exp Med. 8:7049–7058.
2015.PubMed/NCBI
|
|
16
|
Sui H, Shi C, Yan Z and Li H: Combination
of erlotinib and a PARP inhibitor inhibits growth of A2780 tumor
xenografts due to increased autophagy. Drug Des Devel Ther.
9:3183–3190. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Fan H, Jiang M, Li B, He Y, Huang C, Luo
D, Xu H, Yang L and Zhou J: MicroRNA-let-7a regulates cell
autophagy by targeting Rictor in gastric cancer cell lines MGC-803
and SGC-7901. Oncol Rep. 39:1207–1214. 2018.PubMed/NCBI
|
|
18
|
Seo SU, Woo SM, Lee HS, Kim SH, Min KJ and
Kwon TK: mTORC1/2 inhibitor and curcumin induce apoptosis through
lysosomal membrane permeabilization-mediated autophagy. Oncogene.
37:5205–5220. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu Y, Sun Y and Zhao A: MicroRNA-134
suppresses cell proliferation in gastric cancer cells via targeting
of GOLPH3. Oncol Rep. 37:2441–2448. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sarbassov DD, Guertin DA, Ali SM and
Sabatini DM: Phosphorylation and regulation of Akt/PKB by the
rictor-mTOR complex. Science. 307:1098–1101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hresko RC and Mueckler M: mTOR. Rictor is
the Ser473 kinase for Akt/protein kinase B in 3T3-L1 adipocytes. J
Biol Chem. 280:40406–40416. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yuan TL and Cantley LC: PI3K pathway
alterations in cancer: Variations on a theme. Oncogene.
27:5497–5510. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Treins C, Warne PH, Magnuson MA, Pende M
and Downward J: Rictor is a novel target of p70 S6 kinase-1.
Oncogene. 29:1003–1016. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lang F, Strutz-Seebohm N, Seebohm G and
Lang UE: Significance of SGK1 in the regulation of neuronal
function. J Physiol. 588:3349–3354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Leong ML, Maiyar AC, Kim B, O'Keeffe BA
and Firestone GL: Expression of the serum- and
glucocorticoid-inducible protein kinase, Sgk, is a cell survival
response to multiple types of environmental stress stimuli in
mammary epithelial cells. J Biol Chem. 278:5871–5882. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
García-Martínez Juan M and Alessi Dario R:
mTOR complex 2 (mTORC2) controls hydrophobic motif phosphorylation
and activation of serum- and glucocorticoid-induced protein kinase
1 (SGK1). Biochem J. 416:375–385. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhao L, Zhu L, Oh YT, Qian G, Chen Z and
Sun SY: Rictor, an essential component of mTOR complex 2, undergoes
caspase-mediated cleavage during apoptosis induced by multiple
stimuli. Apoptosis. 26:338–347. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wen FF, Li XY, Li YY, He S, Xu XY, Liu YH,
Liu L and Wu SH: Expression of Raptor and Rictor and their
relationships with angiogenesis in colorectal cancer. Neoplasma.
67:501–508. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cao RZ, Min L, Liu S, Tian RY, Jiang HY,
Liu J, Shao LL, Cheng R, Zhu ST, Guo SL and Li P: Rictor activates
Cav 1 through the Akt signaling pathway to inhibit the apoptosis of
gastric cancer cells. Front Oncol. 11:6414532021. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu H, Zang H, Kong J and Gong L: In vivo
and impact of miRNA-153 on the suppression of cell growth apoptosis
through mTORC2 signaling pathway in breast cancer. J Recept Signal
Transduct Res. 42:390–398. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hou B, Liu S, Li E and Jiang X: Different
role of raptor and rictor in regulating Rasfonin-Induced autophagy
and apoptosis in renal carcinoma cells. Chem Biodivers.
17:e20007432020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lu Z, Shi X, Gong F, Li S, Wang Y, Ren Y,
Zhang M, Yu B, Li Y, Zhao W, et al: Rictor/mTORC2 affects
tumorigenesis and therapeutic efficacy of mTOR inhibitors in
esophageal squamous cell carcinoma. Acta Pharm Sin B. 10:1004–1019.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang F, Lou X, Zou Y, Hu D, Liu J, Ning J,
Jiao Y, Zhang Z, Yang F, Fan L, et al: Overexpression of Rictor
protein and Rictor-H. pylori interaction has impact on tumor
progression and prognosis in patients with gastric cancer. Folia
Histochem Cytobiol. 58:96–107. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang S, Amato KR, Song W, Youngblood V,
Lee K, Boothby M, Brantley-Sieders DM and Chen J: Regulation of
endothelial cell proliferation and vascular assembly through
distinct mTORC2 signaling pathways. Mol Cell Biol. 35:1299–1313.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liang X, Sun R, Zhao X, Zhang Y, Gu Q,
Dong X, Zhang D, Sun J and Sun B: Rictor regulates the vasculogenic
mimicry of melanoma via the Akt-MMP-2/9 pathway. J Cell Mol Med.
21:3579–3591. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Guan B, Wu K, Zeng J, Xu S, Mu L, Gao Y,
Wang K, Ma Z, Tian J, Shi Q, et al: Tumor-suppressive microRNA-218
inhibits tumor angiogenesis via targeting the mTOR component Rictor
in prostate cancer. Oncotarget. 8:8162–8172. 2027. View Article : Google Scholar
|
|
37
|
Dormond O, Contreras AG, Meijer E, Datta D
and Flynn E: CD40-induced signaling in human endothelial cells
results in mTORC2- and Akt-dependent expression of vascular
endothelial growth factor in vitro and in vivo. J Immunol.
181:8088–8095. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Alizadeh AM, Shiri S and Farsinejad S:
Metastasis review: From bench to bedside. Tumour Biol.
35:8483–8523. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Guertin DA, Stevens DM, Thoreen CC, Burds
AA, Kalaany NY, Moffat J, Brown M, Fitzgerald KJ and Sabatini DM:
Ablation in mice of the mTORC components raptor, Rictor, or mLST8
reveals that mTORC2 is required for signaling to Akt-FOXO and
PKCalpha, but not S6K1. Dev Cell. 11:859–871. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hua H, Kong Q, Zhang H, Wang J, Luo T and
Jiang Y: Targeting mTOR for cancer therapy. J Hematol Oncol.
12:712019. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Agarwal NK, Chen CH, Cho H, Boulbès DR,
Spooner E and Sarbassov DD: Rictor regulates cell migration by
suppressing RhoGDI2. Oncogene. 32:2521–2526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Agarwal NK, Kazyken D and Sarbassov dos D:
Rictor encounters RhoGDI2: The second pilot is taking a lead. Small
GTPases. 4:102–105. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Savukaitytė A, Gudoitytė G, Bartnykaitė A,
Ugenskienė R and Juozaitytė E: siRNA knockdown of REDD1 facilitates
aspirin-mediated dephosphorylation of mTORC1 target 4E-BP1 in
MDA-MB-468 human breast cancer cell line. Cancer Manag Res.
13:1123–1133. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Wei F, Zhang Y, Geng L, Zhang P, Wang G
and Liu Y: mTOR inhibition induces EGFR feedback activation in
association with its resistance to human pancreatic cancer. Int J
Mol Sci. 16:3267–3282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Lang SA, Hackl C, Moser C, Fichtner-Feigl
S, Koehl GE, Schlitt HJ, Geissler EK and Stoeltzing O: Implication
of Rictor in the mTOR inhibitor-mediated induction of insulin-like
growth factor-I receptor (IGF-IR) and human epidermal growth factor
receptor-2 (HER2) expression in gastrointestinal cancer cells.
Biochim Biophys Acta. 1803:435–442. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yin Y, Hua H, Li M, Liu S, Kong Q, Shao T,
Wang J, Luo Y, Wang Q, Luo T, et al: mTORC2 promotes type I
insulin-like growth factor receptor and insulin receptor activation
through the tyrosine kinase activity of Mtor. Cell Res. 26:46–65.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Hossain MS, Karuniawati H, Jairoun AA,
Urbi Z, Ooi J, John A, Lim YC, Kibria KMK, Mohiuddin AKM, Ming LC,
et al: Colorectal cancer: A review of carcinogenesis, global
epidemiology, current challenges, risk factors, preventive and
treatment strategies. Cancers (Basel). 14:17322022. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Bellier J, Nokin MJ, Caprasse M, Tiamiou
A, Blomme A, Scheijen JL, Koopmansch B, MacKay GM, Chiavarina B,
Costanza B, et al: Methylglyoxal scavengers resensitize
KRAS-Mutated colorectal tumors to cetuximab. Cell Rep.
30:1400–1416.e6. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Shuhua W, Chenbo S, Yangyang L, Xiangqian
G, Shuang H, Tangyue L and Dong T: Autophagy-related genes Raptor,
Rictor, and Beclin 1 expression and relationship with multidrug
resistance in colorectal carcinoma. Hum Pathol. 46:1752–1759. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wei Y, Tang X, Ren Y, Yang Y, Song F, Fu
J, Liu S, Yu M, Chen J, Wang S, et al: An RNA-RNA crosstalk network
involving HMGB1 and RICTOR facilitates hepatocellular carcinoma
tumorigenesis by promoting glutamine metabolism and impedes
immunotherapy by PD-L1+ exosomes activity. Signal Transduct Target
Ther. 6:4212021. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Reyes-Gordillo K, Shah R,
Arellanes-Robledo J, Cheng Y, Ibrahim J and Tuma PL: Akt1 and Akt2
isoforms play distinct roles in regulating the development of
inflammation and fibrosis associated with alcoholic liver disease.
Cells. 8:13372019. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guri Y, Colombi M, Dazert E, Hindupur SK,
Roszik J, Moes S, Jenoe P, Heim MH, Riezman I, Riezman H and Hall
MN: mTORC2 promotes tumorigenesis via lipid synthesis. Cancer Cell.
32:807–823.e12. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Dong X, Feng M, Yang H, Liu H, Guo H, Gao
X, Liu Y, Liu R, Zhang N, Chen R and Kong R: Rictor promotes cell
migration and actin polymerization through regulating ABLIM1
phosphorylation in Hepatocellular Carcinoma. Int J Biol Sci.
16:2835–2852. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Hu J, Che L, Li L, Pilo MG, Cigliano A,
Ribback S, Li X, Latte G, Mela M, Evert M, et al: Co-activation of
Akt and c-Met triggers rapid hepatocellular carcinoma development
via the mTORC1/FASN pathway in mice. Sci Rep. 6:204842016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Villanueva A, Chiang DY, Newell P, Peix J,
Thung S, Alsinet C, Tovar V, Roayaie S, Minguez B, Sole M, et al:
Pivotal role of mTOR signaling in hepatocellular carcinoma.
Gastroenterology. 135:1972–1983. 1983.e1–e11. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Xu Z, Xu M, Liu P, Zhang S, Shang R, Qiao
Y, Che L, Ribback S, Cigliano A, Evert K, et al: The mTORC2-Akt1
Cascade Is Crucial for c-Myc to Promote Hepatocarcinogenesis in
Mice and Humans. Hepatology. 70:1600–1613. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Lin XM, Hu L, Gu J, Wang RY, Li L, Tang J,
Zhang BH, Yan XZ, Zhu YJ, Hu CL, et al: Choline Kinase α mediates
interactions between the epidermal growth factor receptor and
mechanistic target of rapamycin complex 2 in hepatocellular
carcinoma cells to promote drug resistance and xenograft tumor
progression. Gastroenterology. 152:1187–1202. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Joechle K, Guenzle J, Hellerbrand C,
Strnad P, Cramer T, Neumann UP and Lang SA: Role of mammalian
target of rapamycin complex 2 in primary and secondary liver
cancer. World J Gastrointest Oncol. 13:1632–1647. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Yokoi K, Kobayashi A, Motoyama H, Kitazawa
M, Shimizu A, Notake T, Yokoyama T, Matsumura T, Takeoka M and
Miyagawa SI: Survival pathway of cholangiocarcinoma via Akt/mTOR
signaling to escape RAF/MEK/ERK pathway inhibition by sorafenib.
Oncol Rep. 39:843–850. 2018.PubMed/NCBI
|
|
60
|
Hou G, Zhao Q, Zhang M, Fan T, Liu M, Shi
X, Ren Y, Wang Y, Zhou J and Lu Z: Down-regulation of Rictor
enhances cell sensitivity to PI3K inhibitor LY294002 by blocking
mTORC2-medicated phosphorylation of Akt/PRAS40 in esophageal
squamous cell carcinoma. Biomed Pharmacother. 106:1348–1356. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Kim ST, Kim SY, Klempner SJ, Yoon J, Kim
N, Ahn S, Bang H, Kim KM, Park W, Park SH, et al:
Rapamycin-insensitive companion of mTOR (RICTOR) amplification
defines a subset of advanced gastric cancer and is sensitive to
AZD2014-mediated mTORC1/2 inhibition. Ann Oncol. 28:547–554. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Schmidt KM, Hellerbrand C, Ruemmele P,
Michalski CW, Kong B, Kroemer A, Hackl C, Schlitt HJ, Geissler EK
and Lang SA: Inhibition of mTORC2 component Rictor impairs tumor
growth in pancreatic cancer models. Oncotarget. 8:24491–24505.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Mpilla GB, Uddin MH, Al-Hallak MN,
Aboukameel A, Li Y, Kim SH, Beydoun R, Dyson G, Baloglu E,
Senapedis WT, et al: PAK4-NAMPT dual inhibition sensitizes
pancreatic neuroendocrine tumors to everolimus. Mol Cancer Ther.
20:1836–1845. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Zhao Y, Schoeps B, Yao D, Zhang Z, Schuck
K, Tissen V, Jäger C, Schlitter AM, van der Kammen R, Ludwig C, et
al: mTORC1 and mTORC2 Converge on the Arp2/3 complex to promote
Kras-induced Acinar-to-ductal metaplasia and early pancreatic
carcinogenesis. Gastroenterology. 160:1755–1770.e17. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhang X, Chu J, Sun H, Zhao D, Ma B, Xue
D, Zhang W and Li Z: MiR-155 aggravates impaired autophagy of
pancreatic acinar cells through targeting Rictor. Acta Biochim
Biophys Sin (Shanghai). 52:192–199. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Elia A, Henry-Grant R, Adiseshiah C,
Marboeuf C, Buckley RJ, Clemens MJ, Mudan S and Pyronnet S:
Implication of 4E-BP1 protein dephosphorylation and accumulation in
pancreatic cancer cell death induced by combined gemcitabine and
TRAIL. Cell Death Dis. 8:32042017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Eng CP, Sehgal SN and Vézina C: Activity
of rapamycin (AY-22,989) against transplanted tumors. J Antibiot
(Tokyo). 37:1231–1237. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Chiarini F, Evangelisti C, McCubrey JA and
Martelli AM: Current treatment strategies for inhibiting Mtor in
cancer. Trends Pharmacol Sci. 36:124–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wu SH, Bi JF, Cloughesy T, Cavenee WK and
Mischel PS: Emerging function of mTORC2 as a core regulator in
Glioblastoma: Metabolic reprogramming and drug resistance. Cancer
Biol Med. 11:255–263. 2014.PubMed/NCBI
|
|
71
|
Masui K, Harachi M, Cavenee WK, Mischel PS
and Shibata N: mTOR Complex 2 is an integrator of cancer metabolism
and epigenetics. Cancer Lett. 478:1–7. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Zhou HY and Huang SL: Current development
of the second generation of mTOR inhibitors as anticancer agents.
Chin J Cancer. 31:8–18. 2012.PubMed/NCBI
|
|
73
|
Hu Y, Zhang K, Zhu X, Zheng X, Wang C, Niu
X, Jiang T, Ji X, Zhao W, Pang L, et al: Synergistic inhibition of
drug-resistant colon cancer growth with PI3K/mTOR dual inhibitor
BEZ235 and Nano-emulsioned paclitaxel via reducing multidrug
resistance and promoting apoptosis. Int J Nanomedicine.
16:2173–2186. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Hermanowicz JM, Kalaska B, Pawlak K,
Sieklucka B, Miklosz J, Mojzych M and Pawlak D: Preclinical
toxicity and safety of MM-129-First-in-Class BTK/PD-L1 inhibitor as
a potential candidate against colon cancer. Pharmaceutics.
13:12222021. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Foley TM, Payne SN, Pasch CA, Yueh AE, Van
De Hey DR, Korkos DP, Clipson L, Maher ME, Matkowskyj KA, Newton MA
and Deming DA: APC dual PI3K/mTOR inhibition in colorectal cancers
with and mutations. Mol Cancer Res. 15:317–327. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Lou J, Lv JX, Zhang YP and Liu ZJ: OSI-027
inhibits the tumorigenesis of colon cancer through mediation of
c-Myc/FOXO3a/PUMA axis. Cell Biol Int. 46:1204–1214. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Wang H, Liu Y, Ding J, Huang Y, Liu J, Liu
N, Ao Y, Hong Y, Wang L, Zhang L, et al: Targeting mTOR suppressed
colon cancer growth through 4EBP1/eIF4E/PUMA pathway. Cancer Gene
Ther. 27:448–460. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Chang GR, Kuo CY, Tsai MY, Lin WL, Lin TC,
Liao HJ, Chen CH and Wang YC: Anti-cancer effects of zotarolimus
combined with 5-fluorouracil treatment in HCT-116 colorectal
cancer-bearing BALB/c Nude Mice. Molecules. 26:46832021. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Rashid MM, Lee H and Jung BH: Evaluation
of the antitumor effects of PP242 in a colon cancer xenograft mouse
model using comprehensive metabolomics and lipidomics. Sci Rep.
10:175232020. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Wang L, Zhu YR, Wang S and Zhao S:
Autophagy inhibition sensitizes WYE-354-induced anti-colon cancer
activity in vitro and in vivo. Tumor Biol. 37:11743–11752. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Chen Y, Lee CH, Tseng BY, Tsai YH, Tsai
HW, Yao CL and Tseng SH: AZD8055 exerts antitumor effects on colon
cancer cells by inhibiting mTOR and Cell-cycle Progression.
Anticancer Res. 38:1445–1454. 2018.PubMed/NCBI
|
|
82
|
Jin ZZ, Wang W, Fang DL and Jin YJ: mTOR
inhibition sensitizes ONC201-induced anti-colorectal cancer cell
activity. Biochem Biophys Res Commun. 478:1515–1520. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Nguyen DQ, Hoang DH, Nelson M, Nigam L,
Nguyen VTT, Zhang L, Pham TKT, Ho HD, Nguyen DDT, Lam TQ, et al:
Requirement of GTP binding for TIF-90-regulated ribosomal RNA
synthesis and oncogenic activities in human colon cancer cells. J
Cell Physiol. 235:7567–7579. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Reita D, Bour C, Benbrika R, Groh A,
Pencreach E, Guérin E and Guenot D: Synergistic Anti-tumor effect
of mTOR inhibitors with irinotecan on colon cancer cells. Cancers
(Basel). 11:15812019. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Wang Y, Miao X, Jiang Y, Wu Z, Zhu X, Liu
H, Wu X, Cai J, Ding X and Gong W: The synergistic antitumor effect
of IL-6 neutralization with NVP-BEZ235 in hepatocellular carcinoma.
Cell Death Dis. 13:1462022. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Narahara S, Watanabe T, Nagaoka K,
Fujimoto N, Furuta Y, Tanaka K, Tokunaga T, Kawasaki T, Yoshimaru
Y, Setoyama H, et al: Clusterin and related scoring index as
potential early predictors of response to sorafenib in
hepatocellular carcinoma. Hepatol Commun. 6:1198–1212. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Cao W, Liu X, Zhang Y, Li A, Xie Y, Zhou
S, Song L, Xu R, Ma Y, Cai S and Tang X: BEZ235 increases the
sensitivity of hepatocellular carcinoma to sorafenib by inhibiting
PI3K/Akt/mTOR and inducing autophagy. Biomed Res Int.
2021:55563062021. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Liang Y, Xie C, Li A, Huo Z, Wu B, Cai S,
Cao W, Ma Y, Xu R, Jiang Z, et al: Anti-GPC3 Antibody-Conjugated
BEZ235 loaded polymeric nanoparticles (Ab-BEZ235-NP) enhances
radiosensitivity in hepatocellular carcinoma cells by inhibition of
DNA double-strand break repair. J Biomed Nanotechnol. 16:446–455.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Xie Z, Wang J, Liu M, Chen D, Qiu C and
Sun K: CC-223 blocks mTORC1/C2 activation and inhibits human
hepatocellular carcinoma cells in vitro and in vivo. PLoS One.
12:e01732522017. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Choi HJ, Park JH, Kim OH, Kim KH, Hong HE,
Seo H and Kim SJ: Combining Everolimus and Ku0063794 Promotes
apoptosis of hepatocellular carcinoma cells via reduced autophagy
resulting from diminished expression of miR-4790-3p. Int J Mol Sci.
22:28592021. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Yongxi T, Haijun H, Jiaping Z, Guoliang S
and Hongying P: Autophagy inhibition sensitizes KU-0063794-mediated
anti-HepG2 hepatocellular carcinoma cell activity in vitro and in
vivo. Biochem Biophys Res Commun. 465:494–500. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Zhen MC, Wang FQ, Wu SF, Zhao YL, Liu PG
and Yin ZY: Identification of mTOR as a primary resistance factor
of the IAP antagonist AT406 in hepatocellular carcinoma cells.
Oncotarget. 8:9466–9475. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Kaneya Y, Takata H, Wada R, Kure S, Ishino
K, Kudo M, Kondo R, Taniai N, Ohashi R, Yoshida H and Naito Z:
Inhibitor for protein disulfide-isomerase family A member 3
enhances the antiproliferative effect of inhibitor for mechanistic
target of rapamycin in liver cancer: An study on combination
treatment with everolimus and 16F16. Oncol Lett. 21:282021.
View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Navarro-Villarán E, de la Cruz-Ojeda P,
Contreras L, González R, Negrete M, Rodríguez-Hernández MA,
Marín-Gómez LM, Álamo-Martínez JM, Calvo A, Gómez-Bravo MA, et al:
Molecular pathways leading to induction of cell death and
anti-proliferative properties by tacrolimus and mTOR inhibitors in
liver cancer cells. Cell Physiol Biochem. 54:457–473. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Zhang S, Song X, Cao D, Xu Z, Fan B, Che
L, Hu J, Chen B, Dong M, Pilo MG, et al: Pan-mTOR inhibitor MLN0128
is effective against intrahepatic cholangiocarcinoma in mice. J
Hepatol. 67:1194–1203. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Jee HY, Lee YG, Lee S, Elvira R, Seo HE,
Lee JY, Han J and Lee K: Activation of ERK and p38 reduces
AZD8055-mediated inhibition of protein synthesis in hepatocellular
carcinoma HepG2 cell line. Int J Mol Sci. 22:118242021. View Article : Google Scholar : PubMed/NCBI
|
|
97
|
Patra T, Meyer K, Ray RB, Kanda T and Ray
R: Akt inhibitor augments anti-proliferative efficacy of a dual
mTORC1/2 inhibitor by FOXO3a activation in p53 mutated
hepatocarcinoma cells. Cell Death Dis. 12:10732021. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Liu M, Gu P, Guo W and Fan X: C6 ceramide
sensitizes the anti-hepatocellular carcinoma (HCC) activity by
AZD-8055, a novel mTORC1/2 dual inhibitor. Tumor Biol.
37:11039–11048. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Peng X, Zhang D, Li Z, Fu M and Liu H:
mTOR inhibition sensitizes human hepatocellular carcinoma cells to
resminostat. Biochem Biophys Res Commun. 477:556–562. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Weber H, Leal P, Stein S, Kunkel H, García
P, Bizama C, Espinoza JA, Riquelme I, Nervi B, Araya JC, et al:
Rapamycin and WYE-354 suppress human gallbladder cancer xenografts
in mice. Oncotarget. 6:31877–31888. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Li Q, Mou LJ, Tao L, Chen W, Sun XT, Xia
XF, Wu XY and Shi XL: Inhibition of mTOR suppresses human
gallbladder carcinoma cell proliferation and enhances the
cytotoxicity of 5-fluorouracil by downregulating MDR1 expression.
Eur Rev Med Pharmacol Sci. 20:1699–1706. 2016.PubMed/NCBI
|
|
102
|
Mohri D, Ijichi H, Miyabayashi K,
Takahashi R, Kudo Y, Sasaki T, Asaoka Y, Tanaka Y, Ikenoue T,
Tateishi K, et al: A potent therapeutics for gallbladder cancer by
combinatorial inhibition of the MAPK and mTOR signaling networks. J
Gastroenterol. 51:711–721. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
103
|
Yokoyama D, Hisamori S, Deguchi Y,
Nishigori T, Okabe H, Kanaya S, Manaka D, Kadokawa Y, Hata H,
Minamiguchi S, et al: PTEN is a predictive biomarker of trastuzumab
resistance and prognostic factor in HER2-overexpressing
gastroesophageal adenocarcinoma. Sci Rep. 11:90132021. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Gao F, Li R, Wei PF, Ou L, Li M, Bai Y,
Luo WJ and Fan Z: Synergistic anticancer effects of everolimus
(RAD001) and Rhein on gastric cancer cells via
phosphoinositide-3-kinase (PI3K)/protein kinase B (Akt)/mammalian
target of rapamycin (mTOR) pathway. Bioengineered. 13:6332–6342.
2022. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Xu E, Zhu H, Wang F, Miao J, Du S, Zheng
C, Wang X, Li Z, Xu F, Xia X and Guan W: OSI-027 alleviates
Oxaliplatin Chemoresistance in gastric cancer cells by suppressing
P-gp induction. Curr Mol Med. 21:922–930. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Xing X, Zhang L, Wen X, Wang X, Cheng X,
Du H, Hu Y, Li L, Dong B, Li Z and Ji J: PP242 suppresses cell
proliferation, metastasis, and angiogenesis of gastric cancer
through inhibition of the PI3K/Akt/mTOR pathway. Anticancer Drugs.
25:1129–1140. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Zaidi AH, Kosovec JE, Matsui D, Omstead
AN, Raj M, Rao RR, Biederman RWW, Finley GG, Landreneau RJ, Kelly
RJ and Jobe BA: PI3K/mTOR dual inhibitor, LY3023414, demonstrates
potent antitumor efficacy against esophageal adenocarcinoma in a
rat model. Ann Surg. 266:91–98. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Du W, Gao A, Herman JG, Wang L, Zhang L,
Jiao S and Guo M: Methylation of NRN1 is a novel synthetic lethal
marker of PI3K-Akt-mTOR and ATR inhibitors in esophageal cancer.
Cancer Sci. 112:2870–2883. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Hou H, Zhao H, Yu X, Cong P, Zhou Y, Jiang
Y and Cheng Y: METTL3 promotes the proliferation and invasion of
esophageal cancer cells partly through Akt signaling pathway.
Pathol Res Pract. 216:1530872020. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Lu Z, Zhang Y, Xu Y, Wei H, Zhao W, Wang
P, Li Y and Hou G: mTOR inhibitor PP242 increases antitumor
activity of sulforaphane by blocking Akt/mTOR pathway in esophageal
squamous cell carcinoma. Mol Biol Rep. 49:451–461. 2022. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Chen B, Xu M, Zhang H, Xu MZ, Wang XJ,
Tang QH and Tang JY: The Antipancreatic cancer activity of OSI-027,
a potent and selective inhibitor of mTORC1 and mTORC2. DNA Cell
Biol. 34:610–617. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Huang B, Wang J, Chen Q, Qu C, Zhang J,
Chen E, Zhang Y, Wang Y, Ni L and Liang T: Gemcitabine enhances
OSI-027 cytotoxicity by upregulation of miR-663a in pancreatic
ductal adenocarcinoma cells. Am J Transl Res. 11:473–485.
2019.PubMed/NCBI
|
|
113
|
Zhi X, Chen W, Xue F, Liang C, Chen BW,
Zhou Y, Wen L, Hu L, Shen J, Bai X and Liang T: OSI-027 inhibits
pancreatic ductal adenocarcinoma cell proliferation and enhances
the therapeutic effect of gemcitabine both in vitro and in vivo.
Oncotarget. 6:26230–26241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Soares HP, Ni Y, Kisfalvi K, Sinnett-Smith
J and Rozengurt E: Different patterns of Akt and ERK feedback
activation in response to rapamycin, active-site mTOR inhibitors
and metformin in pancreatic cancer cells. PLoS One. 8:e572892013.
View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Peng T and Dou QP: Everolimus inhibits
growth of gemcitabine-resistant pancreatic cancer cells via
induction of caspase-dependent apoptosis and G2/M
arrest. J Cell Biochem. 118:2722–2730. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Hofmann BT, Picksak AS, Kwiatkowski M,
Grupp K, Jücker M, Bachmann K, Mercanoglu B, Izbicki JR, Kahlert C,
Bockhorn M, et al: Truncated O-GalNAc glycans impact on fundamental
signaling pathways in pancreatic cancer. Glycobiology. Aug
18–2021.(Epub ahead of print). doi: 10.1093/glycob/cwab088.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Zhu J, Lv J, Chen J, Zhang X and Ji Y:
Down-regulated microRNA-223 or elevated ZIC1 inhibits the
development of pancreatic cancer via inhibiting PI3K/Akt/mTOR
signaling pathway activation. Cell Cycle. 19:2851–2865. 2020.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Lewis CS, Elnakat Thomas H, Orr-Asman MA,
Green LC, Boody RE, Matiash K, Karve A, Hisada YM, Davis HW, Qi X,
et al: mTOR kinase inhibition reduces tissue factor expression and
growth of pancreatic neuroendocrine tumors. J Thromb Haemost.
17:169–182. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Conway JRW, Warren SC, Herrmann D, Murphy
KJ, Cazet AS, Vennin C, Shearer RF, Killen MJ, Magenau A, Mélénec
P, et al: Intravital imaging to monitor therapeutic response in
moving hypoxic regions resistant to PI3K pathway targeting in
pancreatic cancer. Cell Rep. 23:3312–3326. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Sakamoto Y, Yamagishi S, Tanizawa Y,
Tajimi M, Okusaka T and Ojima H: PI3K-mTOR pathway identified as a
potential therapeutic target in biliary tract cancer using a newly
established patient-derived cell panel assay. Jpn J Clin Oncol.
48:396–399. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Joechle K, Jumaa H, Thriene K, Hellerbrand
C, Kulemann B, Fichtner-Feigl S, Lang SA and Guenzle J: Dual
inhibition of mTORC1/2 reduces migration of cholangiocarcinoma
cells by regulation of matrixmetalloproteinases. Front Cell Dev
Biol. 9:7859792021. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Buzzoni R, Pusceddu S, Bajetta E, De Braud
F, Platania M, Iannacone C, Cantore M, Mambrini A, Bertolini A,
Alabiso O, et al: Activity and safety of RAD001 (everolimus) in
patients affected by biliary tract cancer progressing after prior
chemotherapy: A phase II ITMO study. Ann Oncol. 25:1597–1603. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Ewald F, Grabinski N, Grottke A, Windhorst
S, Nörz D, Carstensen L, Staufer K, Hofmann BT, Diehl F, David K,
et al: Combined targeting of Akt and mTOR using MK-2206 and RAD001
is synergistic in the treatment of cholangiocarcinoma. Int J
Cancer. 133:2065–2076. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Rodon J, Dienstmann R, Serra V and
Tabernero J: Development of PI3K inhibitors: Lessons learned from
early clinical trials. Nat Rev Clin Oncol. 10:143–153. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Stuttfeld E, Aylett CH, Imseng S,
Boehringer D, Scaiola A, Sauer E, Hall MN, Maier T and Ban N:
Architecture of the human mTORC2 core complex. Elife. 7:e331012018.
View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Benavides-Serrato A, Lee J, Holmes B,
Landon KA, Bashir T, Jung ME, Lichtenstein A and Gera J: Specific
blockade of Rictor-mTOR association inhibits mTORC2 activity and is
cytotoxic in glioblastoma. PLoS One. 12:e01765992017. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Werfel TA, Wang S, Jackson MA, Kavanaugh
TE, Joly MM, Lee LH, Hicks DJ, Sanchez V, Ericsson PG, Kilchrist
KV, et al: Selective mTORC2 inhibitor therapeutically blocks breast
cancer cell growth and survival. Cancer Res. 78:1845–1858. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Waldner M, Fantus D, Solari M and Thomson
AW: New perspectives on mTOR inhibitors (rapamycin, rapalogs and
TORKinibs) in transplantation. Br J Clin Pharmacol. 82:1158–1170.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Yang C and Malarkannan S: Transcriptional
regulation of NK cell development by mTOR complexes. Front Cell Dev
Biol. 8:5660902020. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Yang W, Gorentla B, Zhong XP and Shin J:
mTOR and its tight regulation for iNKT cell development and
effector function. Mol Immunol. 68:536–545. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Singh Y, Garden OA, Lang F and Cobb BS:
MicroRNA-15b/16 enhances the induction of regulatory T cells by
regulating the expression of Rictor and mTOR. J Immunol.
195:5667–5677. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Moore KN, Hong DS, Patel MR, Pant S,
Ulahannan SV, Jones S, Meric-Bernstam F, Wang JS, Aljumaily R,
Hamilton EP, et al: A Phase 1b trial of prexasertib in combination
with Standard-of-Care agents in advanced or metastatic cancer.
Target Oncol. 16:569–589. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Zhu AX, Kudo M, Assenat E, Cattan S, Kang
YK, Lim HY, Poon RT, Blanc JF, Vogel A, Chen CL, et al: Effect of
everolimus on survival in advanced hepatocellular carcinoma after
failure of sorafenib: The EVOLVE-1 randomized clinical trial. JAMA.
312:57–67. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Geissler EK, Schnitzbauer AA, Zülke C,
Lamby PE, Proneth A, Duvoux C, Burra P, Jauch KW, Rentsch M, Ganten
TM, et al: Sirolimus use in liver transplant recipients with
hepatocellular carcinoma: A randomized, multicenter, open-label
phase 3 trial. Transplantation. 100:116–125. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Chung V, Frankel P, Lim D, Yeon C, Leong
L, Chao J, Ruel N, Luevanos E, Koehler S, Chung S, et al: Phase Ib
trial of mFOLFOX6 and Everolimus (NSC-733504) in patients with
metastatic gastroesophageal adenocarcinoma. Oncology. 90:307–312.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Joka M, Boeck S, Zech CJ, Seufferlein T,
Wichert Gv, Licht T, Krause A, Jauch KW, Heinemann V and Bruns CJ:
Combination of antiangiogenic therapy using the mTOR-inhibitor
everolimus and low-dose chemotherapy for locally advanced and/or
metastatic pancreatic cancer: A dose-finding study. Anticancer
Drugs. 25:1095–1101. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Yu K, Toral-Barza L, Shi C, Zhang WG,
Lucas J, Shor B, Kim J, Verheijen J, Curran K, Malwitz DJ, et al:
Biochemical, cellular, and in vivo activity of novel
ATP-competitive and selective inhibitors of the mammalian target of
rapamycin. Cancer Res. 69:6232–6240. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Xiao Y, Liu P, Wei J, Zhang X, Guo J and
Lin Y: Recent progress in targeted therapy for non-small cell lung
cancer. Front Pharmacol. 14:11255472023. View Article : Google Scholar : PubMed/NCBI
|