Introduction
Coronary artery disease (CAD), also termed coronary
arteriosclerosis, is one of the most common types of heart disease
(1). Morbidity and mortality of
CAD has increased in recent years, reducing the quality of life of
patients and continuing to present an important socioeconomic
problem (2). Early diagnosis of
CAD is difficult, and the mechanism of its onset and progression is
complicated (3). Coronary artery
bypass graft surgery and drug treatments are the primary treatment
strategies for CAD (4). Although
advances have been achieved with regards to CAD treatment, CAD is a
health burden that remains to be solved. Therefore, it is important
to investigate the mechanisms of CAD progression, and explore
potential methods of CAD diagnosis and treatment.
A previous study demonstrated that CAD progression
may be driven by immune factors, traditional risk factors, such as
high blood pressure, diabetes, hyperlipidemia and smoking, and
other novel risk factors; for example, high blood pressure is
involved in the cardiovascular outcome of patients with diabetes
and CAD (5). Apolopoprotein B is a
novel CAD-associated protein that has been identified to stimulate
the proliferation of coronary artery smooth muscle cells and
promote their movement into the subendocardial layer to enhance the
progression of CAD (6).
Interleukin-18 (IL-18) is an independent predictor of the
cardiovascular events in patients with CAD (7). Additionally, increasing evidence
demonstrated that microRNAs (miRNAs/miRs) are crucial in CAD
progression (8). The
overexpression of miR-1 downregulates B-cell lymphoma 2 (Bcl-2)
expression levels by targeting the 3′-untranslated region of Bcl-2
in cardiac muscles and is, thus, closely associated with ischemic
injury (9). miR-210 expression
targets caspase-8-associated protein 2 in ischemic preconditioning
and may contribute to the survival of stem cells; therefore,
protecting from ischemic injury (10). Although numerous risk miRNAs and
crucial genes have been associated with CAD progression, the
mechanism of CAD remains largely unknown.
In previous studies, the molecules in different cell
types were investigated. Due to its interactive and dynamic
properties, blood composition it is often closely associated with
alterations that occur during the progression of disease and
responses to injury (11).
Therefore, whole blood cells may be useful samples for CAD disease
research, and may be an alternative to tissue biopsy (12). Additionally, adhesion of
circulating leukocytes was confirmed to be an important step for
the development of CAD (13).
Furthermore, the platelets of CAD patients may be easily activated
when coronary blood flow is increased (14). These samples are important for CAD
research. The use of computational methods may allow for a more
thorough investigation of the interactions between the molecules in
different cell types, and thus lead to the identification of novel
factors that contribute to CAD progression. Zhang et al
(15) used GSE12288 microarrays to
identify growth factor receptor-bound protein 2 and heat shock
protein family A (Hsp 70) member 8 as the key genes for CAD
development. Chen et al (16) used GSE28858 microarrays to analyze
the key miRNAs (miR-545 and miR-585) associated with CAD. In
addition, Hua et al (17)
identified the CAD-associated miRNA clusters using the same
microarray data. The present study aimed to elucidate the key genes
and miRNAs associated with CAD progression.
Materials and methods
Data resources and preprocessing
The gene expression profiles of GSE20680 (18) and GSE12288 (19) were downloaded from the gene
expression omnibus (GEO) database in NCBI (National Center for
Biotechnology Information; http://www.ncbi.nlm.nih.gov/geo/) based on the
platforms of GPL4133 Agilent-014850 Whole Human Genome Microarray
4×44K G4112F (Feature Number version) and GPL96 [HG-U133A]
Affymetrix Human Genome U133A Array, respectively. The dataset of
GSE20680 contained 143 CAD and 52 control samples, and that of
GSE12288 included 110 CAD and 112 control samples. In addition, the
miRNA expression profile data of GSE28858 (20) was comprised of 12 samples from
patients with premature CAD and 12 age- and gender-matched healthy
control samples. It was downloaded from the GEO database in NCBI
based on the platform of GPL8179 Illumina Human v2 MicroRNA
expression beadchip.
The gene profile data of GSE20680 was preprocessed
using Agilent Feature Extraction software (version 9.5.3.1; Aglient
Technologies, Inc. Santa Clara, CA, USA) (21). The CEL file data of GSE12288 was
preprocessed using the robust multi-array analysis method from the
affy package in R (22). If a gene
had several probes the mean expression value was selected.
Additionally, miRNA IDs from the preprocessed expression matrix of
GSE28858 were transformed into the miRNA symbols.
Differentially expressed gene (DEG)
screening and enrichment analysis
The DEGs in CAD samples were compared with the
control samples from the two gene expression profile datasets using
a t-test in the limma package in R software (23). P<0.05 and a log2
fold-change of 0.1 were selected as thresholds to indicate a
statistically significant difference.
In addition, the significant biological functions
and pathways of the screened DEGs in GSE20680 and GSE12288 were
analyzed using Database for Annotation, Visualization, and
Integrated Discovery (DAVID) (24)
from the Gene Ontology (GO) (25)
and Kyoto Encyclopedia of Genes and Genomes (KEGG) (26) databases with P<0.05.
Protein-protein interaction (PPI) network
construction and module selection
Search Tool for the Retrieval of Interacting
Genes/Proteins (STRING) is a database of known and predicted
protein interactions that may aid in the comprehensive description
of cellular mechanisms and functions (27). The PPI network of the selected DEGs
was constructed using the STRING database. Interaction pairs with a
PPI score of 0.7 were selected for the construction of the final
network.
The modules from the constructed PPI network were
selected using the ClusterOne plugin in Cytoscape software (version
2.8) (28). In addition, the
significant interaction pathways of DEGs with
P<2.727×10−9 were analyzed using DAVID with P<0.05
indicating a statistically significant difference.
Regulatory network construction
The transcriptional associations between
transcription factors (TFs) and target genes are of great
biological significance, and may aid in the analysis of numerous
physiological activities (29).
The TFs and target genes from the selected DEGs in the two profile
datasets were analyzed based on the information of TF-target genes
stored in the UCSC database (30).
Also, the regulatory network of TFs-target genes was constructed
using the Cytoscape software (version 2.8) (31).
Enrichment analysis of common DEGs
The screened DEGs that appeared in the two datasets
(GSE20680 and GSE12288) were considered common DEGs. The
significant biological functions and pathways of the selected
common DEGs were analyzed using DAVID (24) in GO (25) and KEGG (26) database, respectively. P<0.05 was
selected as the cut-off criteria for including a statistically
significant difference.
miRNAs screening and regulatory network
construction of miRNA-targets
The differentially expressed miRNAs in CAD samples
from the GSE28858 dataset were screened and compared with the
control samples using the t-test in the limma package in R
(23). P<0.05 and a
log2 fold-change of 0.1 were selected as the thresholds
for indicating statistically significant differences.
In addition, miRecords (32) and MirWalk (33) are two databases that integrate
miRNA-target interactions with the experimental validated target
genes of miRNAs. The target genes that are regulated by the
selected differentially expressed miRNAs were predicted based on
the miRecords and MirWalk databases. Genes that are present in one
of the two or in both databases were selected for inclusion in the
current study.
CAD-associated miRNA-target selection and
enrichment analysis
Genes that appeared in the predicted miRNA-target
interactions and in the CAD-associated dataset from the Comparative
Toxicogenomics Database (CTD) (34) were confirmed to be the
CAD-associated genes. The significant biological functions and
pathways of the predicted miRNA-targets were analyzed using DAVID
(24) in GO (25) and KEGG (26) databases, respectively with
P<0.05 indicating a statistically significant difference.
Analysis of CAD-associated DEGs and
miRNAs
To investigate the associations between DEGs
(GSE20680 and GSE12288) and differentially expressed miRNAs
(GSE28858) in the CAD samples, the total genes and miRNAs were
integrated to screen for CAD-associated differentially expressed
miRNA-target genes. Additionally, the significant functions of
miRNAs were analyzed using DAVID (24) in the GO (25) database with P<0.05 indicating a
statistically significant difference. A crosstalk network of miRNAs
involved in the same biological processes was constructed,
P<0.0001 indicating a statistically significant difference.
Results
DEG screening and enrichment
analysis
The GSE20680 dataset was comprised of 270 DEGs (167
upregulated and 103 downregulated), and the GSE12288 dataset was
comprised of 2,268 DEGs (534 upregulated and 1,734
downregulated).
The enriched significant GO terms and KEGG pathways
of DEGs in GSE20680 and GSE12288 are indicated in Tables I and II, respectively. The upregulated DEGs in
GSE20680 were involved in cell chemotaxis and positive regulation
of the defense response, while the downregulated DEGs were involved
in the positive regulation of B cell activation (Table IA). Additionally, the significant
pathways of upregulated DEGs included oxidative phosphorylation,
cardiac muscle contraction and metabolic pathways, while the
downregulated genes were enriched in primary immunodeficiency and
gap junction pathways (Table
IIA). Conversely, the upregulated DEGs in the GSE12288 dataset
were involved in system and multicellular organismal processes,
while the downregulated DEGs were involved in cell activation,
activation of immune response and the immune system process
(Table IB). In addition, the
significant pathways of upregulated DEGs included neuroactive
ligand-receptor interaction and dilated cardiomyopathy, while the
downregulated genes were enriched in the phagosome and Fc γ
R-mediated phagocytosis pathways (Table IIB).
 | Table IEnriched GO terms of DEGs in the two
datasets. |
Table I
Enriched GO terms of DEGs in the two
datasets.
A, GSE20680 dataset
|
|---|
| DEG | ID | Name | Count | P-value |
|---|
| Upregulated | GO:0060326 | Cell
chemotaxis | 10 |
4.15×10−7 |
| GO:0031349 | Positive regulation
of defense response | 10 |
2.76×10−5 |
| GO:0045087 | Innate immune
response | 18 |
3.44×10−5 |
| GO:0006952 | Defense
response | 25 |
3.46×10−5 |
| GO:0002376 | Immune system
process | 32 |
4.13×10−5 |
| Downregulated | GO:0002460 | Adaptive immune
response based on somatic recombination of immune receptors built
from immunoglobulin superfamily domains | 8 |
5.42×10−6 |
| GO:0050871 | Positive regulation
of B cell activation | 5 |
5.84×10−6 |
| GO:0042100 | B-cell
proliferation | 5 |
9.01×10−6 |
| GO:0002250 | Adaptive immune
response | 8 |
1.07×10−5 |
| GO:0030890 | Positive regulation
of B cell proliferation | 4 |
1.89×10−5 |
B, GSE12288 dataset
|
|---|
| DEG | DEG | DEG | DEG | DEG |
|---|
| Upregulated | GO:0003008 | System process | 112 |
1.11×10−15 |
| GO:0032501 | Multicellular
organismal process | 255 |
5.66×10−15 |
| GO:0044707 |
Single-multicellular organism process | 248 |
8.22×10−15 |
| GO:0050877 | Neurological system
process | 80 |
7.50×10−11 |
| GO:0048731 | System
development | 155 |
6.31×10−8 |
| Downregulated | GO:0001775 | Cell
activation | 161 | 0 |
| GO:0002253 | Activation of
immune response | 105 | 0 |
| GO:0002376 | Immune system
process | 379 | 0 |
| GO:0002474 | Antigen processing
and presentation of peptide antigen via MHC class I | 43 | 0 |
| GO:0002682 | Regulation of
immune system process | 203 | 0 |
| Table IIEnriched Kyoto Encyclopedia of Genes
and Genomes pathways of DEGs in the two datasets. |
Table II
Enriched Kyoto Encyclopedia of Genes
and Genomes pathways of DEGs in the two datasets.
A, GSE20680 dataset
|
|---|
| DEG | ID | Name | Count | P-value |
|---|
| Upregulated | hsa0190 | Oxidative
phosphorylation | 8 |
4.47×10−5 |
| hsa0:5010 | Alzheimer's
disease | 7 |
1.31×10−3 |
| hsa0:5012 | Parkinson's
disease | 6 |
1.81×10−3 |
| hsa0:3050 | Proteasome | 3 |
9.54×10−3 |
| hsa0:5016 | Huntington's
disease | 6 |
9.68×10−3 |
| hsa0:3018 | RNA
degradation | 3 |
3.40×10−2 |
| hsa0:4260 | Cardiac muscle
contraction | 3 |
4.18×10−2 |
| hsa0:1100 | Metabolic
pathways | 17 |
4.88×10−2 |
| Downregulated | hsa0:5340 | Primary
immunodeficiency | 3 |
1.21×10−3 |
| hsa0:4540 | Gap junction | 4 |
2.09×10−3 |
| hsa0:5130 | Pathogenic
Escherichia coli infection | 3 |
4.70×10−3 |
| hsa0:260 | Glycine, serine and
threonine metabolism | 2 |
1.62×10−2 |
| hsa0:4916 | Melanogenesis | 3 |
2.34×10−2 |
| hsa0:4114 | Oocyte meiosis | 3 |
3.06×10−2 |
| hsa0:4672 | Intestinal immune
network for IgA production | 2 |
3.46×10−3 |
B, GSE12288 dataset
|
|---|
| DEG | ID | Name | Count | P-value |
|---|
| Upregulated | hsa0:4080 | Neuroactive
ligand-receptor interaction | 21 |
5.45×10−5 |
| hsa0:5414 | Dilated
cardiomyopathy | 10 |
3.26×10−4 |
| hsa0:5412 | Arrhythmogenic
right ventricular cardiomyopathy | 8 |
1.55×10−3 |
| hsa0:4610 | Complement and
coagulation cascades | 7 |
4.35×10−3 |
| hsa0:4970 | Salivary
secretion | 8 |
4.97×10−3 |
| hsa0:4976 | Bile secretion | 7 |
5.10×10−3 |
| hsa0:4972 | Pancreatic
secretion | 8 |
1.05×10−2 |
| hsa0:5410 | Hypertrophic
cardiomyopathy | 7 |
1.18×10−2 |
| Downregulated | hsa0:4145 | Phagosome | 54 |
4.07×10−12 |
| hsa0:4666 | Fc γ R-mediated
phagocytosis | 38 |
6.91×10−11 |
| hsa0:5131 | Shigellosis | 29 |
1.21×10−10 |
| hsa0:5130 | Pathogenic
Escherichia coli infection | 26 |
2.13×10−9 |
| hsa0:4722 | Neurotrophin
signaling pathway | 43 |
2.96×10−9 |
| hsa0:4142 | Lysosome | 41 |
6.79×10−9 |
| hsa0:4144 | Endocytosis | 56 |
3.98×10−8 |
| hsa0:4380 | Osteoclast
differentiation | 41 |
4.25×10−8 |
PPI network construction and module
selection
The PPI networks of DEGs in the two datasets were
constructed. The PPI network of DEGs in GSE20680 contains 68 nodes
and 60 interaction pairs (Fig. 1A)
while the PPI network of DEGs in GSE12288 includes 1,558 nodes and
7,695 pairs. The four modules from the PPI network of DEGs in
GSE12288 were selected for further analysis. The significant
pathways of DEGs in the selected four modules are indicated in
Table III. Genes in module 1
were involved in the spliceosome and RNA polymerase pathway, genes
in module 2 were enriched in proteasome and oocyte meiosis
pathways, and genes in module 4 participated in oxidative
phosphorylation and cardiac muscle contraction (Table III). Module 3 did not contain any
enriched pathway genes.
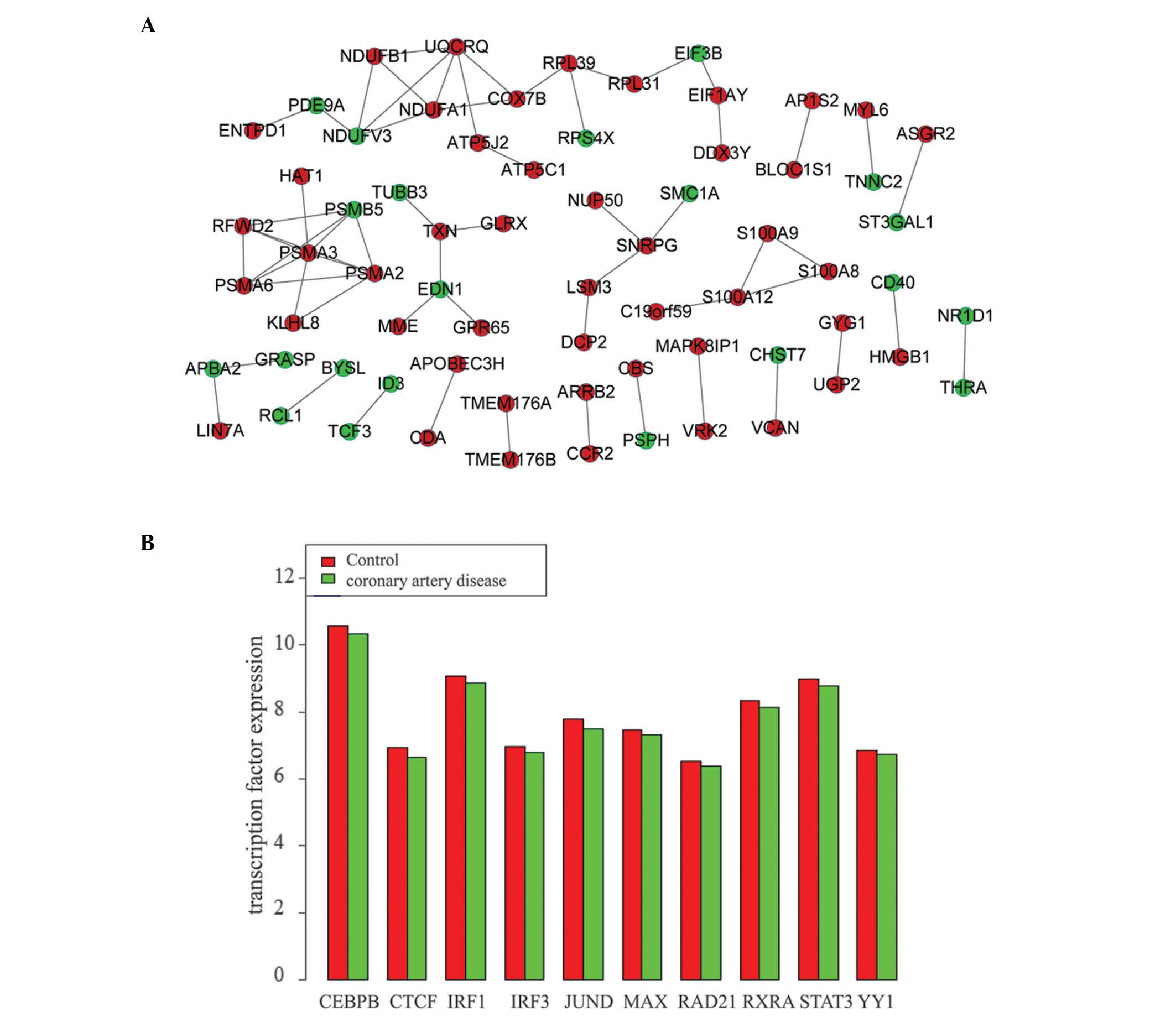 | Figure 1Interactions among DEGs in the two
datasets. (A) Protein-protein interaction network of DEGs in the
GSE20680 dataset. (B) Expression values of the ten transcription
factors in GSE12288 dataset. DEGs, differentially expressed genes;
CEBPB, CCAAT enhancer-binding protein β; CTCF, CCCTC-binding factor
(zinc finger protein); IRF1/2, interferon regulatory factor 1/3;
JUND; jun D proto-oncogene, MAX, MYC-associated factor X; RAD21,
RAD21 cohesin complex component; RXRA, retinoid X receptor α;
STAT3, signal transducer and activator of transcription 3; YY1, YY1
transcription factor. |
| Table IIIEnriched Kyoto Encyclopedia of Genes
and Genomes pathways of differentially expressed genes in the
selected significant modules in GSE12288 dataset. |
Table III
Enriched Kyoto Encyclopedia of Genes
and Genomes pathways of differentially expressed genes in the
selected significant modules in GSE12288 dataset.
| Module | ID | Name | Count | P-value |
|---|
| Module 1 | hsa0:3040 | Spliceosome | 24 | 0 |
| hsa0:3020 | RNA polymerase | 5 |
6.00×10−7 |
| hsa0:3015 | mRNA surveillance
pathway | 6 |
7.79×10−6 |
| hsa0:240 | Pyrimidine
metabolism | 5 |
2.68×10−4 |
| hsa0:3013 | RNA transport | 5 |
1.83×10−3 |
| hsa0:230 | Purine
metabolism | 5 |
2.50×10−3 |
| hsa0:5016 | Huntington's
disease | 5 |
4.23×10−3 |
| hsa0:3420 | Nucleotide excision
repair | 2 |
2.79×10−2 |
| Module 2 | hsa0:3050 | Proteasome | 17 | 0 |
| hsa0:4114 | Oocyte meiosis | 3 |
7.99×10−3 |
| hsa0:4110 | Cell cycle | 3 |
1.06×10−2 |
| hsa0:4120 | Ubiquitin mediated
proteolysis | 3 |
1.33×10−2 |
| hsa0:4914 |
Progesterone-mediated oocyte
maturation | 2 |
4.05×10−2 |
| Module 4 | hsa0:190 | Oxidative
phosphorylation | 24 | 0 |
| hsa0:5010 | Alzheimer's
disease | 23 | 0 |
| hsa0:5012 | Parkinson's
disease | 23 | 0 |
| hsa0:5016 | Huntington's
disease | 23 | 0 |
| hsa0:1100 | Metabolic
pathways | 22 |
2.84×10−14 |
| hsa0:4260 | Cardiac muscle
contraction | 8 |
3.76×10−10 |
Regulatory network construction
No TF target genes were obtained from the GSE20680
dataset. However, a total of 3,400 TF target genes were obtained
from the GSE12288 dataset, including 10 TFs [CCAAT enhancer-binding
protein β; CCCTC-binding factor (zinc finger protein); interferon
regulatory factor 1 and 3 (IRF1 and 3), jun D
proto-oncogene, MYC-associated factor X (MAX); RAD21 cohesin
complex component; retinoid X receptor α; signal transducer and
activator of transcription 3 (STAT3); YY1 transcription factor] and
1,747 target genes. The expression level of these 10 TFs in CAD
samples were analyzed compared with the control samples (Fig. 1B). The expression levels of these
TFs in CAD samples were lower than that in control samples,
indicating that they were all downregulated genes.
In addition, regulatory networks between the 10 TFs
and their target genes [IRF2 and cell death-inducing
DFFA-like effector b (CIDEB)] were constructed. The results
indicated that the number of downregulated genes regulated by the
TFs was greater than the number of upregulated genes.
Enrichment analysis of common DEGs
A total of 41 common DEGs between GSE20680 and
GSE12288 dataset were identified, including IRF2,
fibrinogen-like 2 (FGL2), CIDEB and ribosomal protein
S4, Y-linked 1 (RPS4Y1). The enriched GO terms and KEGG
pathways of these common DEGs are indicated in Table IV. The significant GO terms of
common DEGs included polysaccharide and carbohydrate derivative
metabolic processes, as well as leukocyte mediated immunity
(Table IV), while the enriched
pathways of common genes were amino and nucleotide sugar
metabolism, and protein digestion and absorption (Table IV).
| Table IVEnriched GO terms and Kyoto
Encyclopedia of Genes and Genomes pathways of the common DEGs. |
Table IV
Enriched GO terms and Kyoto
Encyclopedia of Genes and Genomes pathways of the common DEGs.
| ID | Name | Count | P-value |
|---|
| GO:0005976 | Polysaccharide
metabolic process | 4 |
7.93×10−5 |
| GO:1901135 | Carbohydrate
derivative metabolic process | 11 |
2.50×10−4 |
| GO:0002443 | Leukocyte mediated
immunity | 5 |
2.86×10−4 |
| GO:0044710 | Single-organism
metabolic process | 17 |
4.38×10−4 |
| GO:1901564 | Organonitrogen
compound metabolic process | 12 |
5.02×10−4 |
| GO:0005975 | Carbohydrate
metabolic process | 8 |
5.58×10−4 |
| GO:0044281 | Small molecule
metabolic process | 15 |
5.83×10−4 |
| GO:0006022 | Aminoglycan
metabolic process | 4 |
6.57×10−4 |
| GO:1901566 | Organonitrogen
compound biosynthetic process | 7 |
8.45×10−4 |
| GO:0042269 | Regulation of
natural killer cell mediated cytotoxicity | 2 |
8.54×10−4 |
| hsa0:520 | Amino sugar and
nucleotide sugar metabolism | 2 |
1.48×10−2 |
| hsa0:4974 | Protein digestion
and absorption | 2 |
3.95×10−2 |
miRNA screening and regulatory network
construction of miRNA-targets
A total of 214 differentially expressed miRNAs (102
upregulated and 112 downregulated) in CAD samples were screened.
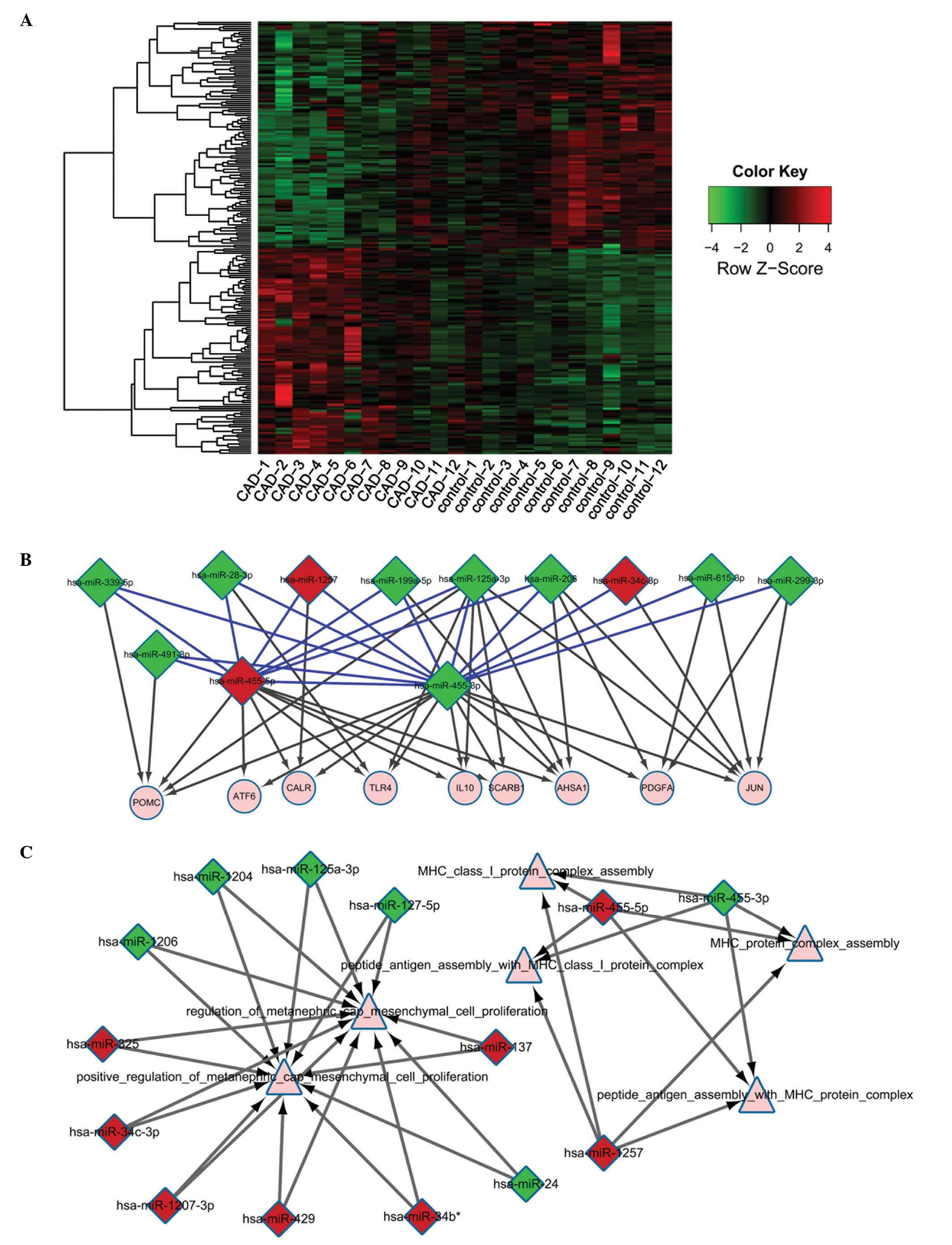
Fig. 2A represents a heat map of
miRNA expression levels. A total of 71 miRNAs were confirmed to
regulate 455 target genes based on the miRRecords and MirWalk
databases. Finally, 640 interactions between the 71 miRNAs and
their target genes were determined.
CAD-associated miRNA target selection and
enrichment analysis of miRNA targets
A total of 402 common genes involved in the
expression of 69 miRNAs were identified based on a comparison
between the 455 target genes that are regulated by the 71 miRNAs
and the genes stored in the CTD database.
In addition, the significant GO terms and KEGG
pathways of the 69 miRNAs are indicated in Table V. GO terms, including major
histocompatibility complex (MHC) protein assembly, MHC class I
protein complex assembly and peptide antigen assembly with MHC
protein complex, were identified as significantly enriched in the
miRNAs tested (Table V). Only one
miRNA was identified to participate in the allograft rejection
pathway (Table V).
| Table VEnriched GO terms and Kyoto
Encyclopedia of Genes and Genomes pathways of microRNAs. |
Table V
Enriched GO terms and Kyoto
Encyclopedia of Genes and Genomes pathways of microRNAs.
| ID | Name | Count | P-value |
|---|
| GO:0002396 | MHC protein complex
assembly | 3 |
1.66×10−3 |
| GO:0002397 | MHC class I protein
complex assembly | 3 |
1.66×10−3 |
| GO:0002501 | Peptide antigen
assembly with MHC protein complex | 3 |
1.66×10−3 |
| GO:0002502 | Peptide antigen
assembly with MHC class I protein complex | 3 |
1.66×10−3 |
| GO:0033139 | Regulation of
peptidyl-serine phosphorylation of STAT protein | 9 |
2.44×10−3 |
| GO:0033141 | Positive regulation
of peptidyl-serine phosphorylation of STAT protein | 9 |
2.44×10−3 |
| GO:0002689 | Negative regulation
of leukocyte chemotaxis | 7 |
4.11×10−3 |
| GO:0090095 | Regulation of
metanephric cap mesenchymal cell proliferation | 11 |
5.52×10−3 |
| GO:0090096 | Positive regulation
of metanephric cap mesenchymal cell proliferation | 11 |
5.52×10−3 |
| GO:0045651 | Positive regulation
of macrophage differentiation | 13 |
1.14×10−2 |
| hsa0:5330 | Allograft
rejection | 14 |
3.80×10−2 |
The regulatory network between miRNAs associated
with CAD and their target genes was also constructed (Fig. 2B). It was identified that
miR-455-5p, miR-455-3p and miR-1257 may regulate numerous miRNAs
and target genes, including pro-opiomelanocortin (POMC),
toll-like receptor 4 (TLR4), IL10, activating
transcription factor 6 (ATF6), and calreticulin
(CALR).
Analysis of CAD associated DEGs and
miRNAs
A total of 5 and 138 miRNA-target interaction pairs
were obtained from the GSE20680 and GSE12288 dataset, respectively.
However, crosstalk analysis of the 5 miRNA-target interaction pairs
was not significant (Table VI).
Additionally, the crosstalk network of miRNAs indicated that miRNAs
were predominantly involved in MHC class I protein complex
assembly, peptide antigen assembly with MHC protein complex,
peptide antigen assembly with MHC class I protein complex,
regulation of metanephric cap mesenchymal cell proliferation,
positive regulation of metanephric cap mesenchymal cell
proliferation, and MHC protein complex assembly (Fig. 2C).
| Table VIEnriched GO terms of microRNAs in
coronary artery disease. |
Table VI
Enriched GO terms of microRNAs in
coronary artery disease.
| ID | Name | Count | P-value |
|---|
| GO:0090095 | Regulation of
metanephric cap mesenchymal cell proliferation | 11 |
2.08×10−4 |
| GO:0090096 | Positive regulation
of metanephric cap mesenchymal cell proliferation | 11 |
2.08×10−4 |
| GO:0002396 | MHC protein complex
assembly | 3 |
5.49×10−4 |
| GO:0002397 | MHC class I protein
complex assembly | 3 |
5.49×10−4 |
| GO:0002501 | Peptide antigen
assembly with MHC protein complex | 3 |
5.49×10−4 |
| GO:0002502 | Peptide antigen
assembly with MHC class I protein complex | 3 |
5.49×10−4 |
| GO:0072185 | Metanephric cap
development | 11 |
2.72×10−3 |
| GO:0072186 | Metanephric cap
morphogenesis | 11 |
2.72×10−3 |
| GO:0090094 | Metanephric cap
mesenchymal cell proliferation involved in metanephros
development | 11 |
2.72×10−3 |
| GO:0072131 | Kidney mesenchyme
morphogenesis | 11 |
7.54×10−3 |
Discussion
CAD is one of the most common types of heart
disease, exhibiting increasing morbidity and mortality rates. CAD
may reduce the quality of life of the patients and is an important
socioeconomic problem (1,2). The mechanism of CAD progression is
complicated; thus, it is important to investigate the disease
mechanism, in addition to exploring different methods for CAD
diagnosis and treatment. In the present study, microarrays were
used to screen for CAD-associated genes and miRNAs. The results
indicated that IRF2 and CIDEB, which were regulated
by STAT3 and MAX, were common DEGs for CAD. In
addition, miR-455-5p, miR-455-3p and miR-1257, which are involved
in the MHC protein complex assembly pathway and peptide antigen
assembly with MHC class I protein complex pathway, may regulate
numerous miRNAs and target genes, including POMC,
TLR4, IL10, ATF6 and CALR.
The results of the current study indicated that the
CAD-associated gene POMC was a target of the upregulated
miR-455-5p, which enriched in the MHC protein complex assembly
pathway. POMC is a polypeptide hormone precursor that functions as
a feeding suppressant and is similar to leptin (35). It has been determined that POMC
neurons were targeted by leptin in the hypothalamus to promote the
synthesis of α-MSH from POMC (36). Additionally, α-MSH acts on the
melanocortin 4 receptor to induce seeding suppression, thus
protecting the body from obesity (37). Logue et al (38) reported that obesity frequently led
to fatal CAD. Therefore, POMC may be associated with CAD
progression. Conversely, miR-455-5p has been identified to target
scavenger receptor class BI and reduce high density lipoprotein
cholesterol (HDL-C) uptake (39).
Lower HDL-C is a predictor for CAD risk (40). Based on the current study,
upregulation of miR-455-5p may reduce the progression of CAD by
targeting the POMC gene via the MHC protein complex assembly
pathway.
The present study demonstrated that the TLR4
gene was the common target of the downregulated miR-455-3p and the
upregulated miR-455-5p. TLR4 is a member of the Toll-like receptor
family, which is important for pathogen recognition and the
activation of innate immunity (41). Otsui et al (42) determined that TLR4 was highly
expressed in smooth muscle cells in patients with atherosclerotic
arteries, and TLR4-mediated inflammatory activation of human
coronary artery endothelial cells via lipopolysaccharide (43). Therefore, TLR4 may be involved in
CAD progression. It has been demonstrated that upregulated
miR-455-3p was involved in the acute myocardial infarction
(44), while myocardial infarction
was the pathological basis for ventricular remodeling in CAD
(45). Therefore, miR-455-3p may
be associated with CAD progression via myocardial infarction. Based
on the results of the present study, downregulated miR-455-3p may
inhibit CAD progression by targeting the TLR4 gene.
The current study indicates that CALR was the
only target gene for the upregulated miR-1257, implying their
respective importance in CAD progression. CALR is a multifunctional
protein that acts as a major Ca2+-binding protein in the
lumen of the endoplasmic reticulum (46). CALR is also associated with the
myocardial hypertrophy. Overexpression of CALR may induce the
dilated cardiomyopathy (47).
Additionally, myocardial hypertrophy is one of the
pathophysiological alterations that occur during CAD (48). Therefore, CALR may be involved in
CAD development. Notably, the role of miR-1257 in CAD remains to be
fully investigated. However, Kamiński et al (49) reported that miR-1257 is a
cardiovascular disease-associated miRNA that has an A binding site.
Based on the observations of the present study, it is possible that
miR-1257 may be a key regulator of CAD progression by regulating
the CALR gene.
The current study also indicated that the
downregulated IRF2 and CIDEB genes were the common
DEGs for CAD. IRF2 was regulated by STAT3 while
CIDEB was regulated by MAX. IRF2 is a member of the
interferon regulatory transcription factor family of proteins that
have a transcriptional binding site for STAT3 (50). The roles of IRF2 and CIDEB in CAD
have not been fully elucidated in previous studies. However,
co-operative IRF1 (the homologue of IRF2) and IL-6 expression was
associated with myocardial infarction (51). In addition, IRF1 inhibited the
differentiation of T helper cells from CD4+ T cells in
the peripheral blood in cases of acute coronary syndrome,
indicating their involvement in the development of this syndrome
(52). STAT3 was also reported to
contribute to heart failure, which is associated with CAD (53). Therefore, based on the results of
the current study IRF-2 may be important in CAD development
regulated by STAT3 while CIDEB may be a novel factor that is
regulated by MAX in CAD.
Additionally, the observations of the present study
indicate that the selected significant miRNAs (miR-455-5p,
miR-455-3p and miR-1257) were involved in the MHC protein complex
assembly pathway and peptide antigen assembly with MHC class I
protein complex pathway. A previous study revealed that the T cell
receptor may only recognize and bind to the peptide fragments of
MHC (54). Higher numbers of
CD4+ T cells may promote the progression of
atherosclerosis (55). In
addition, the interaction between dendritic cells and T cells
contributed towards the process of atherosclerosis (56). The gathered dendritic cells may
secrete tumor necrosis factor-α to induce CD4+ T cells
to produce tumor necrosis factor superfamily member 10 (TNFSF10,
also known as TRAIL) (57). The
TRAIL may combine with its receptors (TRAIL-R1 or TRAIL-R2), which
are located on the vascular smooth muscle cells surface, and then
induce the apoptosis of smooth muscle cells (58). Therefore, it is possible that
miR-455-5p, miR-455-3p and miR-1257, may be important for the
progression of CAD by participating in the MHC protein complex
assembly pathway and peptide antigen assembly with MHC class I
protein complex pathway.
The screened DEGs and TFs were enriched in various
GO terms, including carbohydrate metabolic process, and KEGG
pathways including cardiac muscle contraction and protein digestion
and absorption. Therefore, free fatty acid metabolism was the key
factor in CAD patients, which may regulate the coupling between
carbohydrate oxidation and glycolysis (59). In addition, Fichtlscherer et
al (60) also determined that
certain critical miRNAs, such as miR-133 and miR-208a, were
significantly enriched in cardiac muscle, and further participated
in CAD disease. Therefore, the screened target genes and their
associated TFs may participate in CAD development by being enriched
in the aforementioned pathways.
In conclusion, the present study suggests that
miR-455-5p reduces the progression of CAD by targeting POMC while
miR-455-3p inhibits CAD by targeting TLR4. miR-1257 may be a key
regulator for CAD by targeting CALR. Additionally, IRF-2, which is
regulated by STAT3, may be important in CAD development while
CIDEB, which is regulated by MAX, may be a novel factor in CAD
progression. The current study may provide a basis for future
research on the mechanism of CAD progression. There were however
limitations to the present study. For example, the expression of
the identified molecules and the machinery of the disease process
should be verified in patients with CAD using western blot analyses
and reverse transcription-quantitative polymerase chain reaction.
Therefore, further experimental and clinical studies are required
to confirm the results presented of the present study.
Acknowledgments
The current study was supported by Shanghai City
Jiading District Construction Projects of Medical Subjects (grant
no. TS02).
References
|
1
|
Sun JL, Huang WM, Guo JH, Li XY, Ma XL and
Wang CY: Relationship between myocardial bridging and coronary
arteriosclerosis. Cell Biochem Biophys. 65:485–489. 2013.
View Article : Google Scholar
|
|
2
|
Ahmadi N, Hajsadeghi F, Mirshkarlo HB,
Budoff M, Yehuda R and Ebrahimi R: Post-traumatic stress disorder,
coronary atherosclerosis, and mortality. Am J Cardiol. 108:29–33.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dean JC and Ilvento CC: Improved cancer
detection using computer-aided detection with diagnostic and
screening mammography: Prospective study of 104 cancers. AJR Am J
Roentgenol. 187:20–28. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Velazquez EJ, Lee KL, Deja MA, Jain A,
Sopko G, Marchenko A, Ali IS, Pohost G, Gradinac S, Abraham WT, et
al: STICH Investigators: Coronary-artery bypass surgery in patients
with left ventricular dysfunction. N Engl J Med. 364:1607–1616.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cooper-DeHoff RM, Gong Y, Handberg EM,
Bavry AA, Denardo SJ, Bakris GL and Pepine CJ: Tight blood pressure
control and cardiovascular outcomes among hypertensive patients
with diabetes and coronary artery disease. JAMA. 304:61–68. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Walldius G and Jungner I: The IL/apoA-I
ratio: A strong, new risk factor for cardiovascular disease and a
target for lipid-lowering therapy - a review of the evidence. J
Intern Med. 259:493–519. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hartford M, Wiklund O, Hultén LM, Persson
A, Karlsson T, Herlitz J, Hulthe J and Caidahl K: Interleukin-18 as
a predictor of future events in patients with acute coronary
syndromes. Arterioscler Thromb Vasc Biol. 30:2039–2046. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Contu R, Latronico MV and Condorelli G:
Circulating microRNAs as potential biomarkers of coronary artery
disease A promise to be fulfilled? Circ Res. 107:573–574. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tang Y, Zheng J, Sun Y, Wu Z, Liu Z and
Huang G: MicroRNA-1 regulates cardiomyocyte apoptosis by targeting
Bcl-2. Int Heart J. 50:377–387. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kim HW, Haider HK, Jiang S and Ashraf M:
Ischemic preconditioning augments survival of stem cells via
miR-210 expression by targeting caspase-8-associated protein 2. J
Biol Chem. 284:33161–33168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Taurino C, Miller WH, McBride MW, McClure
JD, Khanin R, Moreno MU, Dymott JA, Delles C and Dominiczak AF:
Gene expression profiling in whole blood of patients with coronary
artery disease. Clin Sci (Lond). 119:335–343. 2010. View Article : Google Scholar
|
|
12
|
Liew CC, Ma J, Tang HC, Zheng R and
Dempsey AA: The peripheral blood transcriptome dynamically reflects
system wide biology: A potential diagnostic tool. J Lab Clin Med.
147:126–132. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Blankenberg S, Rupprecht HJ, Bickel C,
Peetz D, Hafner G, Tiret L and Meyer J: Circulating cell adhesion
molecules and death in patients with coronary artery disease.
Circulation. 104:1336–1342. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Diodati JG, Cannon RO III, Hussain N and
Quyyumi AA: Inhibitory effect of nitroglycerin and sodium
nitroprusside on platelet activation across the coronary
circulation in stable angina pectoris. Am J Cardiol. 75:443–448.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang X, Cheng X, Liu H, Zheng C, Rao K,
Fang Y, Zhou H and Xiong S: Identification of key genes and crucial
modules associated with coronary artery disease by bioinformatics
analysis. Int J Mol Med. 34:863–869. 2014.PubMed/NCBI
|
|
16
|
Chen F, Zhao X, Peng J, Bo L, Fan B and Ma
D: Integrated microRNA-mRNA analysis of coronary artery disease.
Mol Biol Rep. 41:5505–5511. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hua L, Yang Z and Liu H: Explore coronary
artery disease related microRNA clusters by combing single
nucleotide polymor phisms with microRNA microar ray. Fifth
International Conference on Biomedical Engineering and Infromatics
(BMEI). 1193–1197. 2012.
|
|
18
|
Elashoff MR, Wingrove JA, Beineke P,
Daniels SE, Tingley WG, Rosenberg S, Voros S, Kraus WE, Ginsburg
GS, Schwartz RS, et al: Development of a blood-based gene
expression algorithm for assessment of obstructive coronary artery
disease in non-diabetic patients. BMC Medical Genomics. 4:262011.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sinnaeve PR, Donahue MP, Grass P, Seo D,
Vonderscher J, Chibout SD, Kraus WE, Sketch M Jr, Nelson C,
Ginsburg GS, et al: Gene expression patterns in peripheral blood
correlate with the extent of coronary artery disease. PLoS One.
4:e70372009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Sondermeijer BM, Bakker A, Halliani A, de
Ronde MW, Marquart AA, Tijsen AJ, Mulders TA, Kok MG, Battjes S,
Maiwald S, et al: Platelets in patients with premature coronary
artery disease exhibit upregulation of miRNA340* and miRNA624*.
PLoS One. 6:e259462011. View Article : Google Scholar
|
|
21
|
Patterson TA, Lobenhofer EK,
Fulmer-Smentek SB, Collins PJ, Chu T-M, Bao W, Fang H, Kawasaki ES,
Hager J, Tikhonova IR, et al: Performance comparison of one-color
and two-color platforms within the MicroArray Quality Control
(MAQC) project. Nat Biotechnol. 24:1140–1150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
McCall MN and Irizarry RA: Thawing frozen
robust multi-array analysis (fRMA). BMC Bioinformatics. 12:3692011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Efron B and Tibshirani R: On testing the
significance of sets of genes. Ann Appl Stat. 1:107–129. 2007.
View Article : Google Scholar
|
|
24
|
Dennis G Jr, Sherman BT, Hosack DA, Yang
J, Gao W, Lane HC and Lempicki RA: DAVID: Database for annotation,
visualization, and integrated discovery. Genome Biol. 4:32003.
View Article : Google Scholar
|
|
25
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Altermann E and Klaenhammer TR:
PathwayVoyager: Pathway mapping using the Kyoto Encyclopedia of
Genes and Genomes (KEGG) database. BMC Genomics. 6:602005.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering C
and Jensen LJ: STRING v9.1: Protein-protein interaction networks,
with increased coverage and integration. Nucleic Acids Res.
41(Database Issue): D808–D815. 2013. View Article : Google Scholar :
|
|
28
|
Spinelli L, Gambette P, Chapple CE,
Robisson B, Baudot A, Garreta H, Tichit L, Guénoche A and Brun C:
Clust&See: A Cytoscape plugin for the identification,
visualization and manipulation of network clusters. Biosystems.
113:91–95. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Weinmann AS and Farnham PJ: Identification
of unknown target genes of human transcription factors using
chromatin immunoprecipitation. Methods. 26:37–47. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Strauer BE, Brehm M, Zeus T, Bartsch T,
Schannwell C, Antke C, Sorg RV, Kögler G, Wernet P, Müller HW and
Köstering M: Regeneration of human infarcted heart muscle by
intracoronary autologous bone marrow cell transplantation in
chronic coronary artery disease: The IACT Study. J Am Coll Cardiol.
46:1651–1658. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Smoot ME, Ono K, Ruscheinski J, Wang PL
and Ideker T: Cytoscape 2.8: New features for data integration and
network visualization. Bioinformatics. 27:431–432. 2011. View Article : Google Scholar :
|
|
32
|
Xiao F, Zuo Z, Cai G, Kang S, Gao X and Li
T: miRecords: An integrated resource for microRNA-target
interactions. Nucleic Acids Res. 37(Database Issue): D105–D110.
2009. View Article : Google Scholar :
|
|
33
|
Dweep H, Sticht C, Pandey P and Gretz N:
miRWalk - database: Prediction of possible miRNA binding sites by
'walking' the genes of three genomes. J Biomed Inform. 44:839–847.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Davis AP, King BL, Mockus S, Murphy CG,
Saraceni-Richards C, Rosenstein M, Wiegers T and Mattingly CJ: The
comparative toxicogenomics database: Update 2011. Nucleic Acids
Res. 9(Database Issue): D1067–D1072. 2011. View Article : Google Scholar
|
|
35
|
Balthasar N, Coppari R, McMinn J, Liu SM,
Lee CE, Tang V, Kenny CD, McGovern RA, Chua SC Jr, Elmquist JK and
Lowell BB: Leptin receptor signaling in POMC neurons is required
for normal body weight homeostasis. Neuron. 42:983–991. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schwartz MW, Seeley RJ, Woods SC, Weigle
DS, Campfield LA, Burn P and Baskin DG: Leptin increases
hypothalamic pro-opiomelanocortin mRNA expression in the rostral
arcuate nucleus. Diabetes. 46:2119–2123. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
McDaniel FK, Molden BM, Mohammad S and
Baldini G, McPike L, Narducci P, Granell S and Baldini G:
Constitutive cholesterol-dependent endocytosis of melanocortin-4
receptor (MC4R) is essential to maintain receptor responsiveness to
α-melanocyte-stimulating hormone (α-MSH). J Biol Chem.
287:21873–21890. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Logue J, Murray HM, Welsh P, Shepherd J,
Packard C, Macfarlane P, Cobbe S, Ford I and Sattar N: Obesity is
associated with fatal coronary heart disease independently of
traditional risk factors and deprivation. Heart. 97:564–568. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vickers KC, Rye K-A and Tabet F: MicroRNAs
in the onset and development of cardiovascular disease. Clin Sci
(Lond). 126:183–194. 2014. View Article : Google Scholar
|
|
40
|
Cannon CP, Shah S, Dansky HM, Davidson M,
Brinton EA, Gotto AM Jr, Stepanavage M, Liu SX, Gibbons P, Ashraf
TB, et al: Determining the Efficacy and Tolerability Investigators:
Safety of anacetrapib in patients with or at high risk for coronary
heart disease. N Engl J Med. 363:2406–2415. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
O'Neill LA and Bowie AG: The family of
five: TIR-domain-containing adaptors in Toll-like receptor
signalling. Nat Rev Immunol. 7:353–364. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Otsui K, Inoue N, Kobayashi S, Shiraki R,
Honjo T, Takahashi M, Hirata K, Kawashima S and Yokoyama M:
Enhanced expression of TLR4 in smooth muscle cells in human
atherosclerotic coronary arteries. Heart Vessels. 22:416–422. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zeuke S, Ulmer AJ, Kusumoto S, Katus HA
and Heine H: TLR4-mediated inflammatory activation of human
coronary artery endothelial cells by LPS. Cardiovasc Res.
56:126–134. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Meder B, Keller A, Vogel B, Haas J,
Sedaghat-Hamedani F, Kayvanpour E, Just S, Borries A, Rudloff J,
Leidinger P, et al: MicroRNA signatures in total peripheral blood
as novel biomarkers for acute myocardial infarction. Basic Res
Cardiol. 106:13–23. 2011. View Article : Google Scholar
|
|
45
|
Nabel EG and Braunwald E: A tale of
coronary artery disease and myocardial infarction. N Engl J Med.
366:54–63. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Király R, Demény M and Fésüs L: Protein
transamidation by transglutaminase 2 in cells: A disputed
Ca2+-dependent action of a multifunctional protein. FEBS
J. 278:4717–4739. 2011. View Article : Google Scholar
|
|
47
|
Lee D, Oka T, Hunter B, Robinson A, Papp
S, Nakamura K, Srisakuldee W, Nickel BE, Light PE, Dyck JR, et al:
Calreticulin induces dilated cardiomyopathy. PLoS One.
8:e563872013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Basso C, Thiene G, Corrado D, Buja G,
Melacini P and Nava A: Hypertrophic cardiomyopathy and sudden death
in the young: Pathologic evidence of myocardial ischemia. Hum
Pathol. 31:988–998. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kamiński MJ, Kamińska M, Skorupa I,
Kazimierczyk R, Musiał WJ and Kamiński KA: In-silico identification
of cardiovascular disease-related SNPs affecting predicted microRNA
target sites. Pol Arch Med Wewn. 123:355–363. 2013.
|
|
50
|
Yamagata T, Nishida J, Tanaka S, Sakai R,
Mitani K, Yoshida M, Taniguchi T, Yazaki Y and Hirai H: A novel
interferon regulatory factor family transcription factor,
ICSAT/Pip/LSIRF, that negatively regulates the activity of
interferon-regulated genes. Mol Cell Biol. 16:1283–1294. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ridker PM, Rifai N, Stampfer MJ and
Hennekens CH: Plasma concentration of interleukin-6 and the risk of
future myocardial infarction among apparently healthy men.
Circulation. 101:1767–1772. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Guo M, Mao X, Ji Q, Lang M, Li S, Peng Y,
Zhou W, Xiong B and Zeng Q: Inhibition of IFN regulatory factor-1
downregulate Th1 cell function in patients with acute coronary
syndrome. J Clin Immunol. 30:241–252. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Fischer P and Hilfiker-Kleiner D: Survival
pathways in hypertrophy and heart failure: The gp130-STAT3 axis.
Basic Res Cardiol. 102:279–297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Zhang W, Young AC, Imarai M, Nathenson SG
and Sacchettini JC: Crystal structure of the major
histocompatibility complex class I H-2Kb molecule containing a
single viral peptide: Implications for peptide binding and T-cell
receptor recognition. Proc Natl Acad Sci USA. 89:8403–8407. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Mor A, Planer D, Luboshits G, Afek A,
Metzger S, Chajek-Shaul T, Keren G and George J: Role of naturally
occurring CD4+ CD25+ regulatory T cells in
experimental atherosclerosis. Arterioscler Thromb Vasc Biol.
27:893–900. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yilmaz A, Reiss C, Tantawi O, Weng A,
Stumpf C, Raaz D, Ludwig J, Berger T, Steinkasserer A, Daniel WG
and Garlichs CD: HMG-CoA reductase inhibitors suppress maturation
of human dendritic cells: New implications for atherosclerosis.
Atherosclerosis. 172:85–93. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Chen X, Du Y and Huang Z:
CD4+CD25+ Treg derived from hepatocellular
carcinoma mice inhibits tumor immunity. Immunol Lett. 148:83–89.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Döring Y, Manthey HD, Drechsler M, Lievens
D, Megens RT, Soehnlein O, Busch M, Manca M, Koenen RR, Pelisek J,
et al: Auto-antigenic protein-DNA complexes stimulate plasmacytoid
dendritic cells to promote atherosclerosis. Circulation.
125:1673–1683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rosano GM, Vitale C and Fragasso G:
Metabolic therapy for patients with diabetes mellitus and coronary
artery disease. Am J Cardiol. 98:14J–18J. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Fichtlscherer S, De Rosa S, Fox H,
Schwietz T, Fischer A, Liebetrau C, Weber M, Hamm CW, Roxe T, Röxe
T, Müller-Ardogan M, et al: Circulating microRNAs in patients with
coronary artery disease. Circ Res. 107:677–684. 2010. View Article : Google Scholar : PubMed/NCBI
|