Introduction
Formation of new blood vessels via angiogenesis
supports the continued growth of a solid tumor and promotes
hematogenous metastasis, thus it is critical for cancer progression
(1–5). The process of angiogenesis is
mediated by multiple intracellular signaling pathways, including
signal transducer and activator of transcription 3 (STAT3) and
sonic hedgehog (SHH) transduction cascades (6–15).
Aberrant activation of these pathways promotes tumor angiogenesis
by inducing the expression of various critical angiogenic
stimulators (16–20), including vascular endothelial
growth factor A (VEGF-A) and basic fibroblast growth factor (bFGF).
Thus, inhibition of tumor angiogenesis via suppression of the STAT3
and SHH signaling pathways may be a promising strategy for future
cancer therapeutic strategies.
Natural products, including traditional Chinese
medicines, have been used as anti-tumor treatments in China for
thousands of years. They provide an alternative therapeutic
strategy for a variety of diseases and result in relatively few
adverse effects when compared with modern chemotherapeutic agents
(21–23). Oleanolic acid
(3β-hydroxy-olea-12-en-28-oic acid, OA) is the principal active
compound present in various traditional Chinese medicinal herbs
(including Hedyotis diffusa and Patrinia
scabiosaefolia) that have been used to clinically treat various
types of human malignancies (24–27).
Previous studies have reported that OA may suppress tumor growth
via inhibition of proliferation and promotion of apoptosis of
cancer cells (28–30). To further elucidate the underlying
mechanism of anti-cancer activity, the present study used a
colorectal cancer (CRC) mouse xenograft model and human umbilical
vein endothelial cells (HUVECs) to evaluate the effect of OA on
tumor angiogenesis and the activation of the STAT3 and SHH
signaling pathways.
Materials and methods
Materials and reagents
Oleanolic acid (OA) was purchased from Sigma-Aldrich
(St. Louis, MO, USA). Matrigel was provided by BD Biosciences (San
Jose, CA, USA). Dulbecco's modified Eagle's medium (DMEM), RPMI
1640, fetal bovine serum (FBS), penicillin-streptomycin,
trypsin-EDTA, and TRIzol reagent were purchased from Thermo Fisher
Scientific, Inc. (Waltham, MA, USA). PrimeScript RT Reagent kit was
purchased from Takara Bio, Inc. (Otsu, Japan). Goldview Nucleic
Acid Gel Stain (cat. no. G8142) was purchased from Beijing Solarbio
Science & Technology Co., Ltd. (Beijing, China). The In
Vitro Angiogenesis assay kit was purchased from EMD Millipore
(Billerica, MA, USA). BCA Protein assay kit was purchased from
Tiangen Biotech Co., Ltd. (Beijing, China).
Cell culture
HT-29 human colon carcinoma cells were obtained from
the Cell Bank of Chinese Academy of Sciences (Shanghai, China).
Human umbilical vein endothelial cells (HUVECs) were purchased from
Xiangya Cell Center, University of Zhongnan (Hunan, China). HT-29
cells and HUVECs were respectively cultured in DMEM or RPMI 1640,
at 37°C and 5% CO2 in a humidified environment. DMEM and
RPMI 1640 were supplemented with 10% (v/v) FBS, 100 U/ml
penicillin, and 100 μg/ml streptomycin.
In vivo nude mice xenograft study
A total of 20 six-week-old male BALB/c athymic
(nude) mice (weight, 20–22 g) were obtained from Shanghai SLAC
Laboratory Animal Co., Ltd. (Shanghai, China) and housed in a
specific pathogen-free controlled environment (temperature, 22°C;
12 h light/dark cycle) with ad libitum access to food and
water. The HT-29 xenograft mouse model was generated as previously
described (25). Following
xenograft implantation, mice were placed in two treatment groups
(n=10) and treated with 12.5 mg/kg OA [dissolved in
phosphate-buffered saline (PBS)] or saline by daily intraperitoneal
injections, 6 days a week for 16 days. Body weight and tumor size
were assessed. Tumor size was determined by measuring the major (L)
and minor (W) tumor diameter with calipers. The tumor volume (T)
was calculated according to the following formula: T = π/6× L ×
W2. At the end of the experiment, animals were
sacrificed using pelltobarbitalum natricum overdose
(Sigma-Aldrich), and tumors were excised. A portion of each tumor
(0.125 cm3) was fixed in 10% buffered formalin (China
National Medicines Corporation, Ltd., Beijing, China) and the
remaining tissue was snap-frozen in liquid nitrogen and stored at
−80°C. All animal treatments were performed in accordance with
international ethical guidelines and the National Institutes of
Health Guide for the Care and Use of Laboratory Animals (31). The experiments were approved by the
Institutional Animal Care and Use Committee of Fujian University of
Traditional Chinese Medicine (Fuzhou, China).
Immunohistochemical analysis
Slides of paraffin-embedded tumor tissue (n=8; 4
μm; RM2235; Leica Microsystems GmbH, Wetzlar, Germany) were
randomly selected from OA-treated or control groups, and analyzed
by immunohistochemistry as previously described (25). Samples were blocked using normal
goat serum in PBS with 0.1% Tween 20 (Shanghai Sangon Biological
Engineering Technology Co., Ltd., Shanghai, China), then incubated
with rabbit polyclonal antibodies against cluster of
differentiation (CD)31 (cat. no. 3528S; Cell Signaling Technology,
Danvers, MA, USA), VEGF-A (cat. no. sc-7269; Santa Cruz
Biotechnology, Inc.), bFGF (cat. no. 8910LC; Cell Signaling
Technology), SHH (cat. no. sc-1194; Santa Cruz Biotechnology, Inc.)
and GLI-Kruppel family member GLI1 (Gli-1; cat. no. sc-6152; Santa
Cruz Biotechnology, Inc.) (all 1:200) were used to detect the
relevant proteins by incubation overnight at 4°C. Five fields
(magnification, ×400; DM4000; Leica Microsystems GmbH) were
randomly selected from each slide and the proportion of positive
cells in each field was determined using a true color
multi-functional cell image analysis management system (Image-Pro
Plus version 6.0; Media Cybernetics, Rockville, MD, USA). PBS
served as a negative control by replacing the primary antibody.
Cell viability evaluation by MTT
assay
HUVECs were seeded into 96-well plates at a density
of 1×104 cells/well in 0.1 ml RPMI 1640 and were
incubated for 24 h at 37°C. In order to test dose-dependent effects
of OA, the cells were treated with 0, 25, 50 and 100 μM OA
[dissolved in 0.1% dimethyl sulfoxide (DMSO); Sigma-Aldrich] for 24
h. In order to analyze time-dependent effects, with 40 μM of
OA for 0, 1, 3, 6, 12, 8 and 24 h. At the end of the treatment, 10
μl of 5 mg/ml MTT (Sigma-Aldrich) was added to each well,
the samples were then incubated for an additional 4 h. The formazan
precipitate was dissolved in 100 μl DMSO. Absorbance was
measured at 570 nm using an single channel filter-based absorbance
reader (EL×800; BioTek Instruments, Inc., Winooski, VT, USA).
Migration assay of HUVECs
Investigation into the migration of HUVECs was
performed using the wound healing method. HUVECs were seeded into
12-well plates at a density of 2×105 cells/well in 1 ml
medium. After 24 h of incubation, cells were scraped away
vertically in each well using a P100 pipette tip. Three randomly
selected views along the scraped line were photographed in each
well using phase-contrast inverted microscopy at a magnification of
×100 (FMIL/DFC295; Leica Microsystems GmbH). Following treatments
with OA at various concentrations for 24 h, a second set of images
was captured using the same method. The reduction in the size of
the scraped region is indicative of cell migration.
Tube formation assay of HUVECs
Tube formation in HUVECs was investigated using the
In Vitro Angiogenesis assay kit (ECMatrix assay kit; EMD
Millipore) following the manufacturer's protocol. Confluent HUVECs
were harvested and diluted (1×104 cells) in 50 ml RPMI
1640 containing various concentrations of OA. The harvested cells
were seeded into 96-well plates with ECMatrix gel (1:1 v/v) and
incubated for 9 h at 37°C. The cells were photographed using
phase-contrast inverted microscopy at a magnification of ×100
(DMIL/DFC295; Leica Microsystems GmbH). The level of HUVEC tube
formation was quantified by calculating the length of the tubes in
three randomly selected fields from each well.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was isolated from tumor tissues (3 tumors
were randomly selected from OA-treated or control groups) or HT-29
cells with TRIzol reagent. Oligo(dT)-primed RNA (1 μg,
isolated from tumor tissues or cells) was reverse transcribed using
the PrimeScript RT Reagent kit according to the manufacturer's
protocol. Briefly, 1 μl gDNA Eraser was used to remove the
genomic DNA following incubation with total RNA for 2 min at 42°C.
Then, the PrimeScript RT Enzyme Mix and RT Primer Mix were added to
perform the reverse transcription, by incubation for 15 min at
37°C. The obtained cDNA was used to determine the mRNA expression
levels of VEGF-A, bFGF, SHH and Gli-1 by PCR with Taq polymerase
(Fermentas; Thermo Fisher Scientific, Inc.). The primers used for
amplification of VEGF-A, bFGF, SHH, Gli-1, and GAPDH transcripts
were as follows: VEGF-A forward, 5′-CAT CCTGGCCTCGCTGTC-3′ and
reverse, 5′-CTCGCTCCAACCGACTGC-3′; bFGF forward,
5′-CGGCTGTACTGCAAAAACGG-3′ and reverse,
5′-GATGTGAGGGTCGCTCTTCTCC-3′; SHH forward,
5′-CGGAGCGAGGAAGGGAAAG-3′ and reverse,
5′-TTGGGGATAAACTGCTTGTAGGC-3′; Gli-1 forward,
5′-TCTGCCCCCATTGCCCACTTG-3′ and reverse,
5′-TACATAGCCCCCAGCCCATACCTC-3′; and GAPDH forward,
5′-GTCATCCATGACAACTTTGG-3′ and reverse, 5′-GAGCTTGACAAAGTGGTCGT-3′.
The thermal cycling conditions were as follows: Denaturation at
95°C for 30 sec, annealing at the appropriate temperature (VEGF-A
and GAPDH at 58°C, bFGF at 56°C, and SHH and Gli-1 at 54°C) for 30
sec, and extension at 60°C for 30 sec for 30 cycles. GAPDH was used
as an internal control. A negative control with no DNA and an RT
control with no reverse transcription were used as the experimental
controls. The PCR was repeated in 3 independent times. A BIO-RAD
S1000 Thermal Cycler (Bio-Rad Laboratories, Inc., Hercules, CA,
USA) was used to perform the experiment. Samples were analyzed by
1.5% agarose gel electrophoresis. The DNA bands were examined using
a gel documentation system (Gel Doc 2000; Bio-Rad Laboratories,
Inc.).
Western blotting
Protein from tumor tissues (3 tumors were randomly
selected from OA-treated or control groups) and HT-29 cells was
extracted using radioimmunoprecipitation assay protein extraction
kit (Tiangen Biotech Co., Ltd.) containing protease and phosphatase
inhibitor cocktails. For the interleukin (IL)-6 stimulation
experiment, HT-29 cells were grown in complete DMEM (10% FBS) until
~70% confluency, then cultured in FBS-free medium overnight. In
complete DMEM, cells were pre-treated with various concentrations
of OA for 1 h followed by stimulation with 10 ng/ml of IL-6 for 15
min. The concentration of proteins was determined using the BCA
Protein Assay Reagent kit. Proteins (50 μg) were separated
by 10% SDS-PAGE and transferred onto polyvinylidene fluoride
membranes. The membranes were blocked for 1 h with 5% nonfat milk
and incubated with the relevant primary antibody against STAT3
(cat. no. 9132), CD31 (cat. no. 3528S), phosphorylated STAT3 (cat.
no. 9131), β-actin (cat. no. 4967) (all purchased from Cell
Signaling Technology), SHH (sc-1194), VEGF-A (cat. no. sc-7269) or
Gli-1 (cat. no. sc-6152) (all purchased from Santa Cruz
Biotechnology, Inc.) overnight at 4°C (all in 1:1,000 dilutions).
β-actin served as an internal control. The horseradish
peroxidase-conjugated goat anti-rabbit secondary antibody (1:2,000
dilution; cat. no. E030120-01; EarthOx Life Sciences, Millbrae, CA,
USA) were added for 1 h at room temperature. The levels of protein
expression were detected with Enhanced Chemiluminescence (Beyotime
Institute of Biotechnology, Haimen, China) detection. The
chemiluminescence signals were visualized using the SuperSignal
West Pico Chemiluminescent Substrate (Thermo Fisher Scientific,
Inc.). Image Lab Software, version 3.0, was used for densitometric
analysis of the western blotting (Bio-Rad Laboratories, Inc.).
Statistical analysis
Data were presented as mean ± standard deviation for
the indicated number of independently performed experiments. The
data were analyzed using SPSS (version 17.0; SPSS, Inc., Chicago,
IL, USA). Statistical analysis was performed on the data using
Student's t-test and analysis of variance and P<0.05 was
considered to indicate a statistically significant difference.
Results
OA inhibits CRC angiogenesis
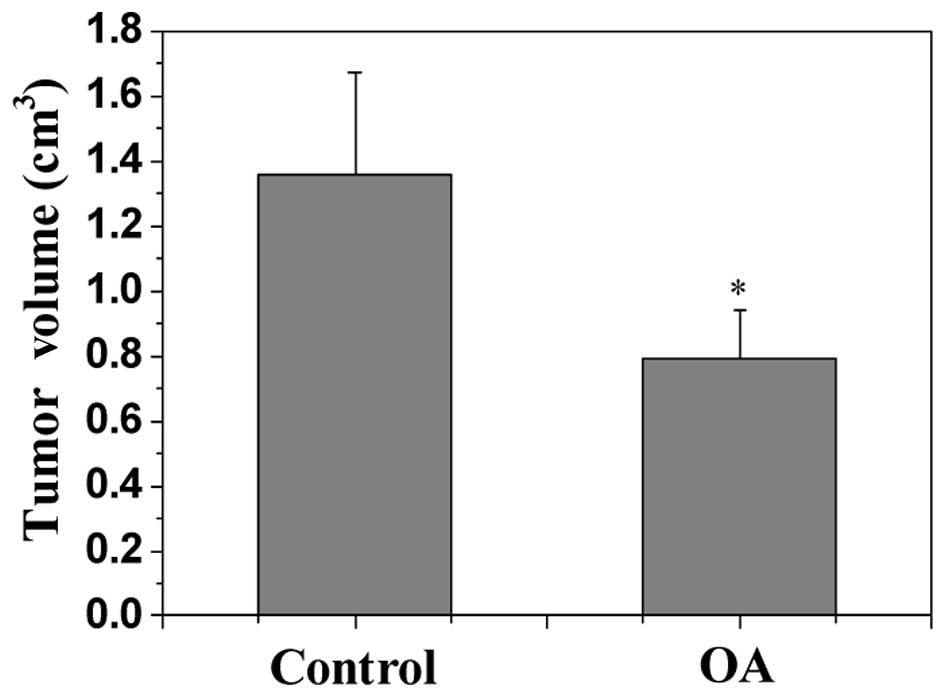
Tumor growth in CRC xenograft mice was evaluated by
determining the volume of the tumor in the mice. As indicated by
Fig. 1, OA treatment resulted in a
significant reduction in tumor volume in CRC mice compared with the
control (P<0.05), demonstrating the in vivo anti-tumor
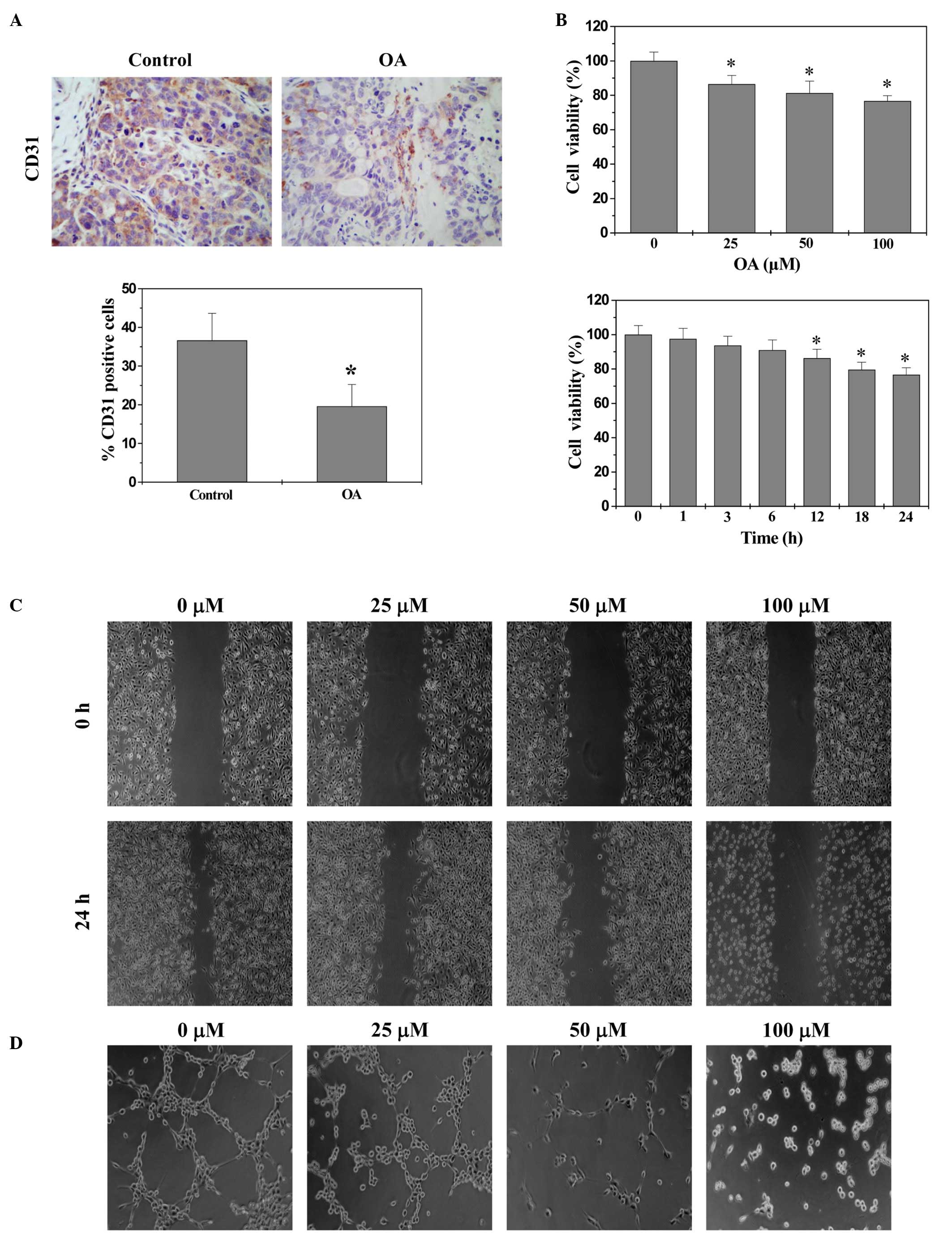
effect of OA. The in vivo tumor angiogenesis [intratumoral
microvessel density (MVD)] was assessed by the expression of CD31,
an endothelial cell-specific marker, using immunohistochemical
staining. As indicated by Fig. 2A,
OA significantly reduced the percentage of CD31-positive cells in
CRC xenograft tumor tissues compared with the control (P<0.01),
indicating its in vivo anti-angiogenic activity. The effect
of OA on in vitro angiogenesis was also evaluated. As
indicated by Fig. 2B–D, OA
treatment significantly decreased the proliferation (viability)
compared with the control (P<0.01), and decreased migration and
capillary tube formation of HUVECs in dose- and/or time-dependent
manner.
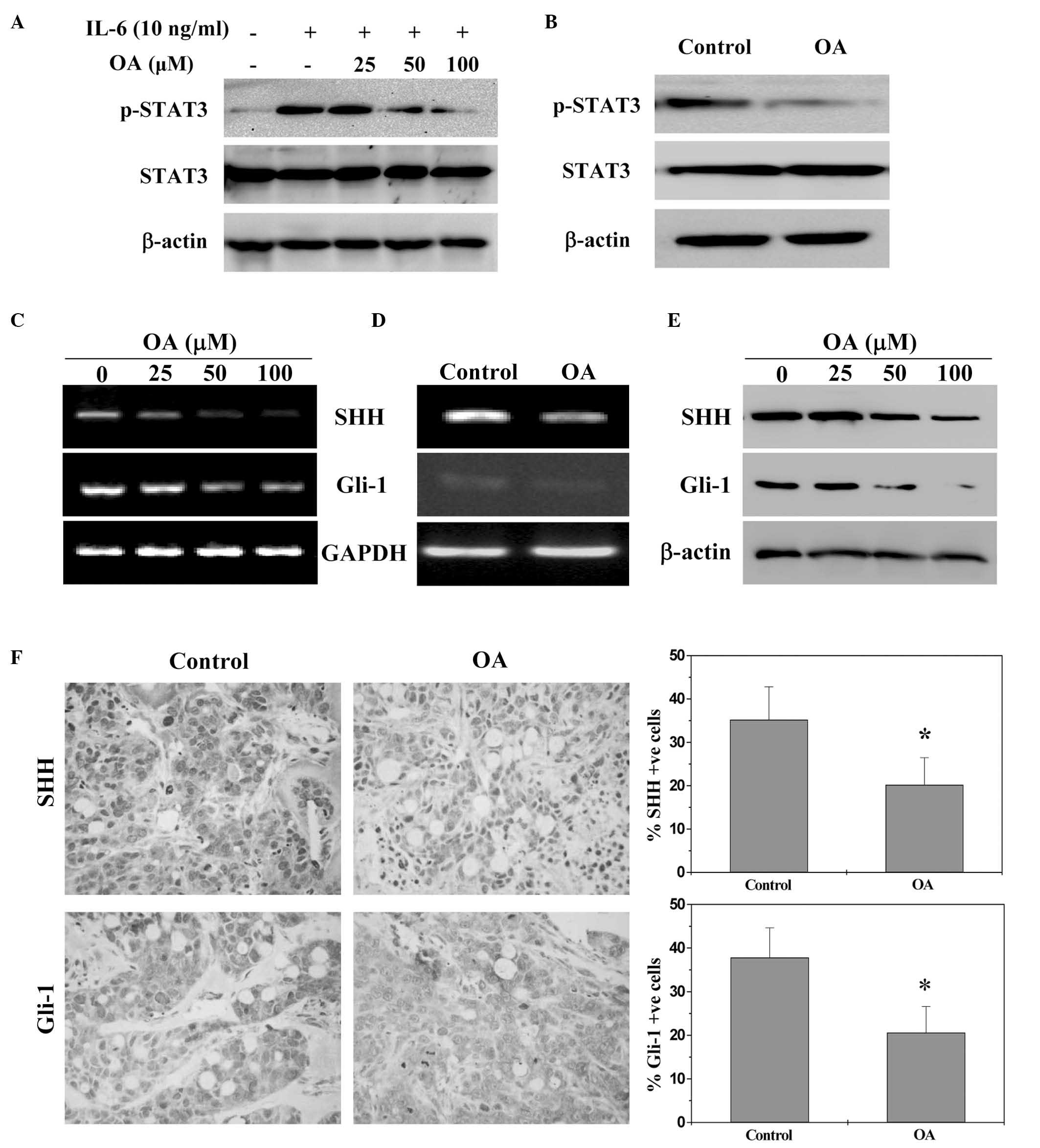
OA suppresses STAT3 and SHH signalling
pathways in vivo and in vitro
STAT3 activation was determined by western blotting
using an antibody that targets STAT3 phosphorylation at
Tyr705. As indicated by Fig. 3A and B, OA reduced IL-6-induced
phosphorylation of STAT3 in HT-29 cells and in tumors of CRC
xenograft mice. The expression levels of non-phosphorylated STAT3
remained unchanged. The activation of SHH pathway was investigated
by examining the expression levels of the key mediators in CRC
xenograft tumors and HT-29 cells. As indicated by Fig. 3C–F, OA treatment significantly
reduced expression of SHH and Gli-1 in vitro and in
vivo compared with the control (P<0.05), at the
transcriptional and translational level.
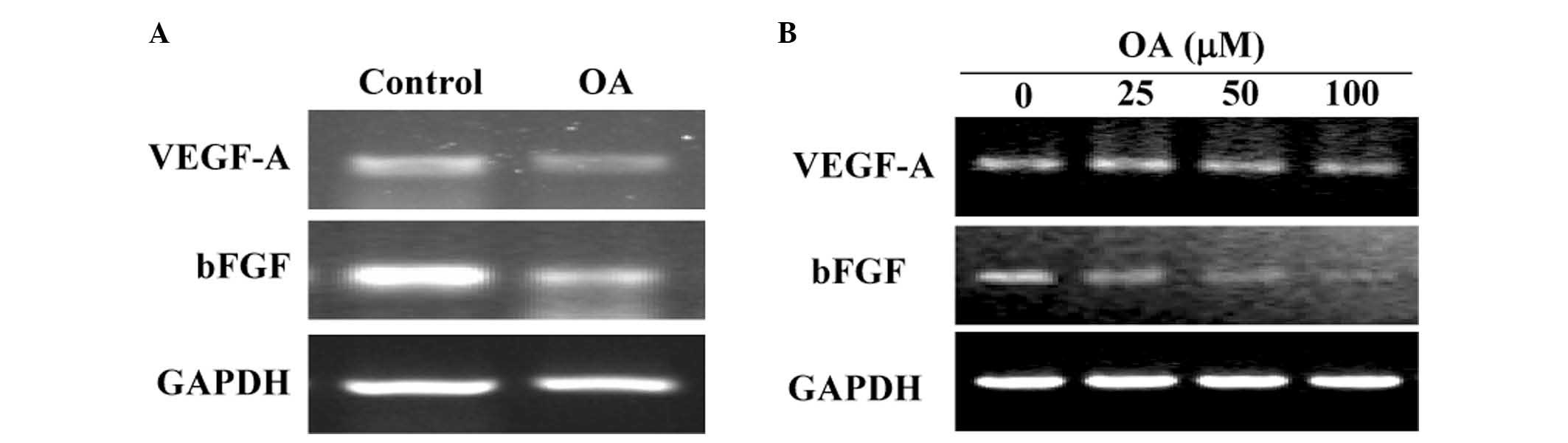
OA inhibits the expression of VEGF-A and
Bfgf
The mRNA expression levels of VEGF-A and bFGF were
determined by RT-PCR analysis. As indicated by Fig. 4, OA treatment decreased VEGF-A and
bFGF mRNA expression levels in CRC xenograft tumor tissues and in
HT-29 cells.
Discussion
Colorectal cancer (CRC) is one of the most common
human malignant cancers. There are over 1 million newly diagnosed
cases of CRC worldwide per year and >500,000 cancer-associated
mortalities (32,33). Chemotherapy is the predominant
non-surgical therapeutic strategy for patients with advanced CRC.
The majority of patients undergoing chemotherapy experience severe
and debilitating side effects, which may be lethal in certain cases
and may considerably outweigh the benefits for the patient
(34,35). In addition, long-term
administration of the current chemotherapeutic agents often results
in drug resistance (36). Thus,
natural products may be an alternative as they produce fewer side
effects and have been used extensively as viable alternative
remedies for a variety of cancers (21–23).
Various traditional Chinese medicinal herbs, including Hedyotic
diffusa, Spica prunellae and Scutellaria barbata, have
been used in clinical cancer treatment in China. OA is a bioactive
compound present in these herbs and has been indicated to exert
anti-cancer effects. However, the underlying mechanisms for these
anti-tumor effects remain to be elucidated.
The present study determined that OA may
significantly reduce intratumoral MVD in tumor tissue compared with
the control (P<0.01), possibly via its inhibitory effect on
multiple critical processes of angiogenesis, including
proliferation, migration and capillary tube formation of
endothelial cells. Therefore, OA may be an effective
anti-angiogenic treatment in vivo and in vitro.
Induction of angiogenesis is mediated by a variety of molecules
released by tumor cells. VEGF-A and bFGF are considered to be
strong stimulators of angiogenesis. Overexpression of VEGF-A and
bFGF has frequently been observed in various types of human cancer,
which is associated with tumor progression (16–20).
Upon binding to their specific receptors, VEGF-A and bFGF exert
their biological function and trigger tyrosine kinase signaling
cascades, resulting in endothelial cell proliferation and
migration, and eventually capillary tube formation. Using RT-PCR
analysis, the present study determined that OA treatment reduced
mRNA expression levels of VEGF-A and bFGF in vivo in the CRC
xenograft tumor tissues and in vitro in HT-29 cells.
The expression of VEGF-A and bFGF is regulated by
multiple signal transduction cascades, including STAT3 and SHH
pathways. STAT3 is a transcription factor, which is essential for
numerous cellular processes. Constitutive activation of STAT3 is
often associated with the development of numerous types of cancer
and frequently indicates a poor prognosis (6–10).
SHH is an extensively investigated member of the Hedgehog family
and its aberrant activation has been associated with numerous types
of human cancer, including CRC (11–15).
OA treatment inhibited STAT3 phosphorylation and the expression of
various key mediators of SHH signaling in CRC tumors and in HT-29
cells, indicating its suppressive effects on the activation of the
STAT3 and SHH signaling pathways.
In conclusion, the present study demonstrated that
inhibition of tumor angiogenesis via suppression of multiple
signaling pathways may be one of the mechanisms by which OA exerts
its anti-cancer function.
Abbreviations:
|
OA
|
oleanolic acid
|
|
CRC
|
colorectal cancer
|
|
STAT3
|
signal transducer and activator of
transcription 3
|
|
SHH
|
sonic hedgehog
|
|
MVD
|
microvessel density
|
|
VEGF-A
|
vascular endothelial growth factor
A
|
|
bFGF
|
basic fibroblast growth factor
|
|
IHS
|
immunohistochemical staining
|
Acknowledgments
The current study was sponsored by the Research Fund
for the Doctoral Program of Higher Education of China (grant no.
20133519110003) and the Developmental Fund of Chen Keji Integrative
Medicine (grant nos. CKJ2014013 and CKJ2015007).
References
|
1
|
Folkman J: Angiogenesis. Annu Rev Med.
57:1–18. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cook KM and Figg WD: Angiogenesis
inhibitors: Current strategies and future prospects. CA Cancer J
Clin. 60:222–243. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mantovani A, Allavena P, Sica A and
Balkwill F: Cancer related inflammation. Nature. 454:436–444. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kerbel RS: Tumor angiogenesis. N Engl J
Med. 358:2039–2049. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Aggarwal BB, Kunnumakkara AB, Harikumar
KB, Gupta SR, Tharakan ST, Koca C, Dey S and Sung B: Signal
transducer and activator of transcription-3, inflammation and
cancer: How intimate is the relationship? Ann NY Acad Sci.
1171:59–76. 2009. View Article : Google Scholar
|
|
7
|
Bromberg J and Wang TC: Inflammation and
cancer: IL-6 and STAT3 complete the link. Cancer Cell. 15:79–80.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kusaba T, Nakayama T, Yamazumi K, Yakata
Y, Yoshizaki A, Inoue K, Nagayasu T and Sekine I: Activation of
STAT3 is a marker of poor prognosis in human colorectal cancer.
Oncol Rep. 15:1445–1451. 2006.PubMed/NCBI
|
|
9
|
Lin Q, Lai R, Chirieac LR, Li C, Thomazy
VA, Grammatikakis I, Rassidakis GZ, Zhang W, Fujio Y, Kunisada K,
et al: Constitutive activation of JAK3/STAT3 in colon carcinoma
tumors and cell lines: Inhibition of JAK3/STAT3 signaling induces
apoptosis and cell cycle arrest of colon carcinoma cells. Am J
Pathol. 167:969–980. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiong H, Zhang ZG, Tian XQ, Sun DF, Liang
QC, Zhang YJ, Lu R, Chen YX and Fang JY: Inhibition of JAK1,
2/STAT3 signaling induces apoptosis, cell cycle arrest and reduces
tumor cell invasion in colorectal cancer cells. Neoplasia.
10:287–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ingham PW, Nakano Y and Seger C:
Mechanisms and functions of Hedgehog signalling across the metazoa.
Nat Rev Genet. 12:393–406. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Theunissen JW and de Sauvage FJ: Paracrine
Hedgehog signaling in cancer. Cancer Res. 69:6007–6010. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yoshikawa K, Shimada M, Miyamoto H,
Higashijima J, Miyatani T, Nishioka M, Kurita N, Iwata T and Uehara
H: Sonic hedgehog relates to colorectal carcinogenesis. J
Gastroenterol. 44:1113–1117. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Varnat F, Duquet A, Malerba M, Zbinden M,
Mas C, Gervaz P and Ruiz i Altaba A: Human colon cancer epithelial
cells harbour active HEDGEHOG-GLI signalling that is essential for
tumour growth, recurrence, metastasis and stem cell survival and
expansion. EMBO Mol Med. 1:338–351. 2009. View Article : Google Scholar
|
|
15
|
Mazumdar T, DeVecchio J, Shi T, Jones J,
Agyeman A and Houghton JA: Hedgehog signaling drives cellular
survival in human colon carcinoma cells. Cancer Res. 71:1092–1102.
2011. View Article : Google Scholar :
|
|
16
|
Kaya M, Wada T, Akatsuka T, Kawaguchi S,
Nagoya S, Shindoh M, Higashino F, Mezawa F, Okada F and Ishii S:
Vascular endothelial growth factor expression in untreated
osteosarcoma is predictive of pulmonary metastasis and poor
prognosis. Clin Cancer Res. 6:572–577. 2000.PubMed/NCBI
|
|
17
|
Maeda K, Chung YS, Ogawa Y, Takatsuka S,
Kang SM, Ogawa M, Sawada T and Sowa M: Prognostic value of vascular
endothelial growth factor expression in gastric carcinoma. Cancer.
77:858–863. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ishigami SI, Arii S, Furutani M, Niwano M,
Harada T, Mizumoto M, Mori A, Onodera H and Imamura M: Predictive
value of vascular endothelial growth factor (VEGF) in metastasis
and prognosis of human colorectal cancer. Br J Cancer.
78:1379–1384. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gille H, Kowalski J, Li B, LeCouter J,
Moffat B, Zioncheck TF, Pelletier N and Ferrara N: Analysis of
biological effects and signaling properties of Flt-1 (VEGFR-1) and
KDR (VEGFR-2). A reassessment using novel receptor-specific
vascular endothelial growth factor mutants. Biol Chem.
276:3222–3230. 2001. View Article : Google Scholar
|
|
21
|
Gordaliza M: Natural products as leads to
anticancer drugs. Clin Transl Oncol. 9:767–776. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Newman DJ, Cragg GM and Snader KM: The
influence of natural products upon drug discovery. Nat Prod Rep.
17:215–234. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang G, Li X, Li X, Wang L, Li J, Song X,
Chen J, Guo Y, Sun X, Wang S, et al: Traditional chinese medicine
in cancer care: A review of case series published in the Chinese
literature. Evid Based Complement Alternat Med. 2012:7510462012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wei L, Lin J, Wu G, Xu W, Li H, Hong Z and
Peng J: Scutellaria barbata D. Don induces G1/S arrest via
modulation of p53 and Akt pathways in human colon carcinoma cells.
Oncol Rep. 29:1623–1628. 2013.PubMed/NCBI
|
|
25
|
Lin J, Wei L, Shen A, Cai Q, Xu W, Li H,
Zhan Y, Hong Z and Peng J: Hedyotis diffusa Willd extract
suppresses Sonic hedgehog signaling leading to the inhibition of
colorectal cancer angiogenesis. Int J Oncol. 42:651–656. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai Q, Lin J, Wei L, Zhang L, Wang L, Zhan
Y, Zeng J, Xu W, Shen A, Hong Z and Peng J: Hedyotis diffusa Willd
inhibits colorectal cancer growth in vivo via inhibition of STAT3
signaling pathway. Int J Mol Sci. 13:6117–6128. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Peng J, Chen Y, Lin J, Zhuang Q, Xu W,
Hong Z and Sferra TJ: Patrinia scabiosaefolia extract suppresses
proliferation and promotes apoptosis by inhibiting the STAT3
pathway in human multiple myeloma cells. Mol Med Rep. 4:313–318.
2011.PubMed/NCBI
|
|
28
|
Wang X, Bai H, Zhang X, Liu J, Cao P, Liao
N, Zhang W, Wang Z and Hai C: Inhibitory effect of oleanolic acid
on hepatocellular carcinoma via ERK-p53-mediated cell cycle arrest
and mitochondrial-dependent apoptosis. Carcinogenesis.
34:1323–1330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lúcio KA, Rocha Gda G, Monção-Ribeiro LC,
Fernandes J, Takiya CM and Gattass CR: Oleanolic acid initiates
apoptosis in non-small cell lung cancer cell lines and reduces
metastasis of a B16F10 melanoma model in vivo. PLoS One.
6:e285962011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wei J, Liu M, Liu H, Wang H, Wang F, Zhang
Y, Han L and Lin X: Oleanolic acid arrests cell cycle and induces
apoptosis via ROS-mediated mitochondrial depolarization and
lysosomal membrane permeabilization in human pancreatic cancer
cells. J Appl Toxicol. 33:756–765. 2013. View Article : Google Scholar
|
|
31
|
Institute of Laboratory Animal Resources
(US); Committee on Care, Use of Laboratory Animals, and National
Institutes of Health (US); Division of Research Resources: Guide
for the care and use of laboratory animals. 8th edition. National
Academies Press; Washington, DC: 2011
|
|
32
|
Siegel R, Desantis C and Jemal A:
Colorectal cancer statistics, 2014. CA Cancer J Clin. 64:104–117.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Markowitz SD and Bertagnolli MM: Molecular
origins of cancer: Molecular basis of colorectal cancer. N Engl J
Med. 361:2449–2460. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hanahan D and Weinberg RA: The hallmarks
of cancer. Cell. 100:57–70. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lippman SM: The dilemma and promise of
cancer chemoprevention. Nat Clin Pract Oncol. 3:5232006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Longley DB, Allen WL and Johnston PG: Drug
resistance, predictive markers and pharmacogenomics in colorectal
cancer. Biochim Biophys Acta. 1766:184–196. 2006.PubMed/NCBI
|