Introduction
Vascular neointimal remodeling is a crucial
pathological process in proliferative cardiovascular diseases, such
as atherosclerosis and restenosis following angioplasty (1). Vascular smooth muscle cells (VSMCs)
are not terminally differentiated; their phenotypic transition from
the differentiated state to the dedifferentiated state induces
alterations in vascular biology and subsequently leads to vascular
neointimal remodeling (2).
However, the molecular mechanisms behind this pathological process
have not been fully determined.
Endogenous, non-coding microRNAs (miRNAs) negatively
regulate the expression of greater than one-third of human genes at
the post-transcriptional level by degrading or inhibiting the
translation of their specific target genes (3). Certain miRNAs determine cell fate and
tissue homeostasis, including VSMCs biology and vascular
remodeling. Identification of these specific miRNAs may aid in the
discovery of novel targets in the prevention and treatment of
proliferative cardiovascular diseases (4,5).
miRNA (miR)-146a was the first small RNA to be
discovered that was demonstrated to be involved in the regulation
of the immune system, mainly associated with the development of
rheumatoid arthritis, cancer and sepsis (6). A previous study demonstrated that the
level of miR-146a expression was significantly higher in the
peripheral blood mononuclear cells of patients with acute coronary
syndrome (7). miR-146a may promote
differentiation and functions of type 1 T helper (Th1) cells, and
may be involved in immune regulation in patients with coronary
heart disease (7). The level of
miR-146a expression was reported to be significantly upregulated in
the carotid artery following carotid artery balloon injury, which
suggested that miR-146a may be involve in the pathophysiology of
restenosis following balloon injury (8). A recent study also revealed that
miR-146a was involved in atherosclerosis in apolipoprotein
E-deficient (ApoE−/−) rats (9). Our previous study demonstrated that
the level of miR-146a was significantly increased during VSMC
proliferation, and that miR-146a promoted the proliferation and
migration of VSMCs and inhibited their apoptosis, but its
mechanisms remains unclear (10).
The present study aimed to explore the target genes
of miR-146a and the signaling pathways that may be involve in
promoting VSMC proliferation to gain a better understanding of its
role in cardiovascular diseases.
Materials and methods
Ethics statement
This study was carried out in strict accordance with
the recommendations in the Guide for the Care and Use of Laboratory
Animals of the National Institutes of Health. The protocol was
approved by the Animal Ethics Committee of the Shenzhen People's
Hospital (Shenzhen, China); all efforts were made to minimize
suffering.
Rat VSMC culture and miR-146a
interference
Male Sprague-Dawley rats (8 weeks-old; 150±20 g;
n=3) were purchased from Guangdong Medical Laboratory Animal Center
(Foshan, China) and housed in specific pathogen-free animal room on
12-h light/dark cycle at 40–70% humidity and 22±2°C with access to
food and water ad libitum. Primary VSMCs were obtained and
isolated from the thoracic aortic media of SD rats (10). The VSMCs were divided into four
groups: i) miR-146a mimics group; ii) miR-146a inhibitor group;
iii) miR-Scramble negative control group; and iv) untransfected
VSMC control group. miR-146a inhibitors were labeled with
6-carboxyfluorescein. Cells (1×105 cells/ml) in miR-146a
mimics group, miR-146a inhibitor group and negative control group
were transfected with miR-146a mimics (50 nmol/l; Shanghai
GenePharma Co., Ltd., Shanghai, China), miR-146a inhibitor (siRNA)
(50 nmol/l; Shanghai GenePharma Co., Ltd.) or miR-Scramble negative
control (50 nmol/l; Shanghai GenePharma Co., Ltd.), respectively,
using Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA) in an incubator containing 5%
CO2 at 37°C for 5 h. PBS (Gibco; Thermo Fisher
Scientific, Inc.) was added at the same equivalent dosage to normal
VSMC group. The medium was replaced with Dulbecco's modified
Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.) 5 h
post-transfection, and transfection efficiency was observed under a
fluorescence microscope. VSMCs were subsequently incubated for 48 h
in the incubator containing 5% CO2 at 37°C prior to
further experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following incubation at 37°C for 48 h
post-transfection, total RNA of VSMCs (3×105 cells/ml)
was extracted using the miRNeasy Mini Kit (Qiagen GmbH, Hilden,
Germany). The cDNA was synthesized using a PrimeScript Reagent Kit
(Takara Bio, Inc., Otsu, Japan). RT-qPCR was accomplished using the
ABI PRISM 7900 Sequence Detection System (Applied Biosystems;
Thermo Fisher Scientific, Inc.) with the SYBR Premix Ex Taq Kit
(Takara Bio, Inc.). Primer sequences were as follows: miR-146a,
forward 5′-TGAGAACTGAATTCCATGGGTT-3′, reverse
5′-GGCCAACCGCGAGAAGATGTTTTTTTTT-3′; caspase-3, forward
5′-GCTGGACTGCGGTATTGAGA-3′, reverse 5′-CCATGACCCGTCCCTTGA-3′;
phosphatase and tensin homology (PTEN), forward
5′-AGACCATAACCCACCACAGC-3′, reverse 5′-TTACACCAGTCCGTCCTTTCC−3′;
p53, forward 5′-AGATTGGGGAATGGGTTGG-3′, reverse
5′-GATAGAATCTTACAGGCGGTGG-3′; cyclin D1, forward
5′-GCGTACCCTGACACCAATCTC-3′, reverse 5′-CTCTTCGCACTTCTGCTCCTC−3′;
U6, forward 5′-CTCGCTTCGGCAGCACA-3′, reverse
5′-ACGCTTCACGAATTTGCGT-3′. PCR conditions were as follows: One
cycle at 95°C for 30 sec, 40 cycles at 95°C for 5 sec and one cycle
at 60°C for 30 sec. Relative expression levels of miR-146a,
caspase-3, PTEN, p53, cyclin D1 and U6 (internal control) were
calculated by using SDS 2.1.1 software (Applied Biosystems; Thermo
Fisher Scientific, Inc.). All primers were synthesized by Sangon
Biotech Co., Ltd. (Shanghai, China). All Experiments were performed
according to the manufacturer's protocol.
VSMC proliferation
VSMCs seeded (1×104 cells/well) in the
96-well plate were cultured in the incubator containing 5%
CO2 at 37°C for 24 h. The cells were transfected with
miR-146a inhibitor (50 nM), miR-Scramble control (50 nM) or PBS for
5 h at 37°C. The proliferative properties of the VSMCs were
measured using the Cell Counting Kit-8 (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) and cell counting was
performed 48 h after VSMC transfection. The absorbance was measured
at 450 nm with a reference wavelength at 650 nm using microplate
reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The number
of VSMCs was counted using a hemocytometer under an IX51 light
microscope (Olympus Corporation, Tokyo, Japan). All experiments
were performed six times.
Microarray expression profiling
analysis
VSMCs (1×105 cells/ml) cultured in an
incubator containing 5% CO2 at 37°C were divided into
miR-146a inhibitor group and miR-146a scramble group (n=3
samples/group). VSMCs were transfected with miR-146a inhibitor (50
nmol/l) or miR-146a scramble (50 nmol/l) in 5% CO2 at
37°C for 5 h at room temperature prior to further incubation in 5%
CO2 at 37°C for 48 h. Total RNA was isolated from VSMCs
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to manufacturer's protocol. Gene profile was scanned
using an Agilent DNA Microarray Scanner (Agilent Technologies,
Inc., Santa Clara, CA, USA) and data were collected using Agilent
Feature Extraction software (v11.0.1.1; Agilent Technologies,
Inc.). Gene Ontology (http://www.geneontology.org) analysis was carried out
to detect different genes and Kyoto Encyclopedia of Genes and
Genomes (http://www.genome.jp/KEGG) analysis
was carried out to detect different pathways. Enrichment scores
were calculated to rank distinct pathways according to the P-values
of genes detected in these pathways.
Western blot analysis
Following incubated at 5% CO2, 37°C for
48 h post-transfection, cultured cells were lysed on ice for 30 min
in 1× RIPA Lysis Buffer (Sangon Biotech Co., Ltd.) supplemented
with a protease inhibitor cocktail (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany). Protein concentrations of the cell lysates
were measured with the BCA Protein Assay kit (Sangon Biotech Co.,
Ltd.). Denatured proteins (20 µg/lane) were electrophoresed on 10%
SDS-polyacrylamide gels and were electrotransferred from the gel to
PDVF membranes (EDM Millipore, Billerica, MA, USA). Following
blocking with 5% non-fat milk in a Tris buffered saline with 0.05%
Tween-20 for 2 h at room temperature, membranes were incubated
overnight at 4°C with polyclonal rabbit anti-rat Caspase-3
(1:1,000; ab217; Abcam, Cambridge, MA, USA), polyclonal rabbit
anti-rat PTEN (1:1,000; ab32199; Abcam), polyclonal rabbit anti-rat
P53 (1:200; ab1431; Abcam), monoclonal rabbit anti-rat cyclinD1
(1:1,000; ab134175; Abcam) and GAPDH (1:1,000; ab8245; Abcam). The
blots were subsequently incubated in 1:6,000 dilutions of goat
anti-rabbit horseradish peroxidase-conjugated secondary antibodies
for 1 h at room temperature. ECL kit (Thermo Fisher Scientific
Inc.) developing solution (100 µl) was added to each sample to
detect the protein signal with a Quant RT ECL cold CCD imaging
system (GE Healthcare Life Sciences, Little Chalfont, UK). GAPDH
was used as the internal control.
Statistical analysis
Data are presented as the mean ± standard deviation
and were analyzed using SPSS 12.0 statistical software (SPSS, Inc.,
Chicago, IL, USA), one-way analysis of variance was used to analyze
the data, with a Student-Newman-Keuls-q test used for pairwise
comparison. P<0.05 was considered to indicate a statistically
significant difference.
Results
miR-146a promotes VSMC
proliferation
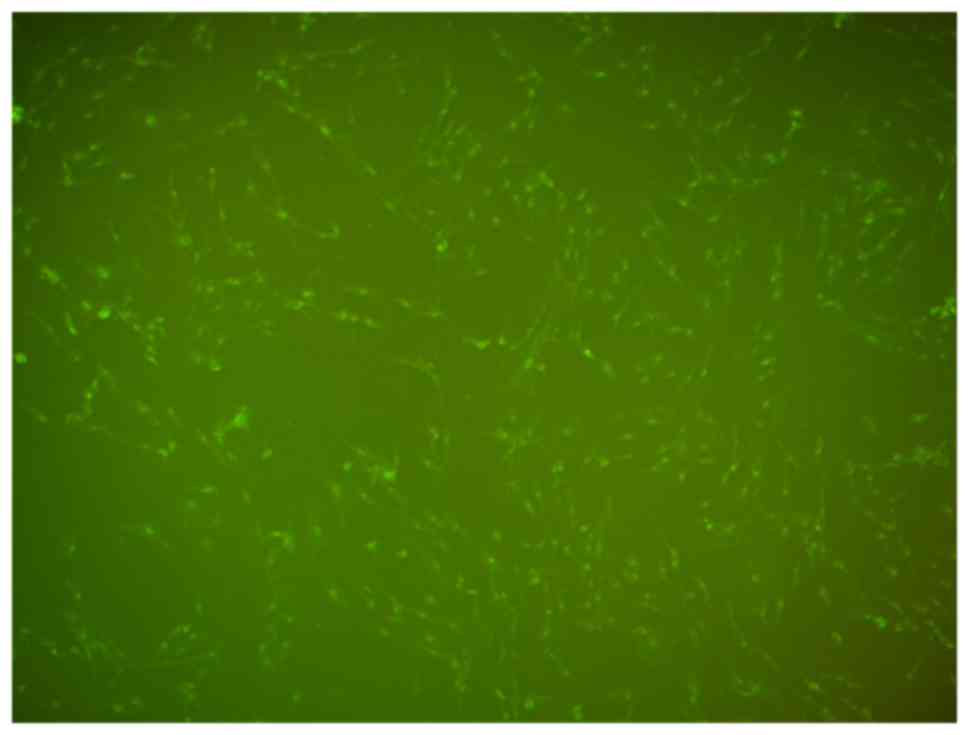
Following transfection with the miR-146a inhibitor
for 5 h, a large number of fluorescent granules were visible in the
cytoplasm of VSMCs under fluorescence microscopy, which indicated
that miR-146a inhibitor was successfully transfected into the VSMCs
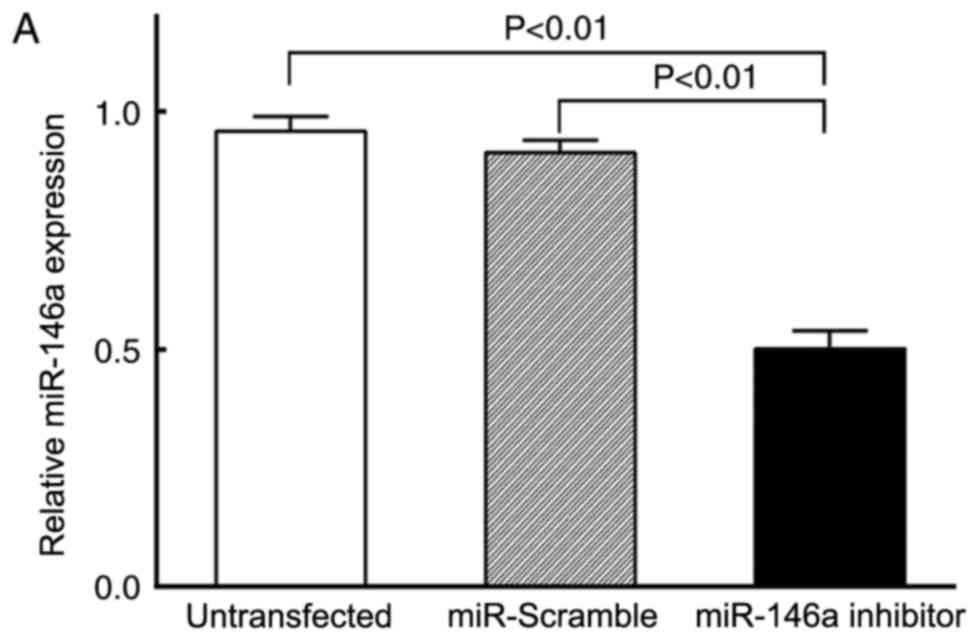
(Fig. 1). The level of miR-146a
expression was detected by RT-qPCR. Compared with cells transfected
with the miR-Scramble negative control and the untransfected VSMCs,
the relative expression level of miR-146a in the miR-146a inhibitor
group was significantly decreased (P<0.01; Fig. 2A). No significant differences were
identified between the miR-Scramble and the untransfected control
cells (P>0.05; Fig. 2A). VSMC
proliferation was detected 48 h post-transfection by CCK-8 analysis
and cell counting. No significant differences were identified for
the OD values and the cell numbers between the miR-Scramble
negative control group and the untransfected VSMC group (P>0.05;
Fig. 2B and C); however, the OD
values and the cell numbers were significantly decreased in cells
transfected with the miR-146a inhibitor (P<0.01; Fig. 2B and C).
p53 signaling pathway is a miR-146a
target
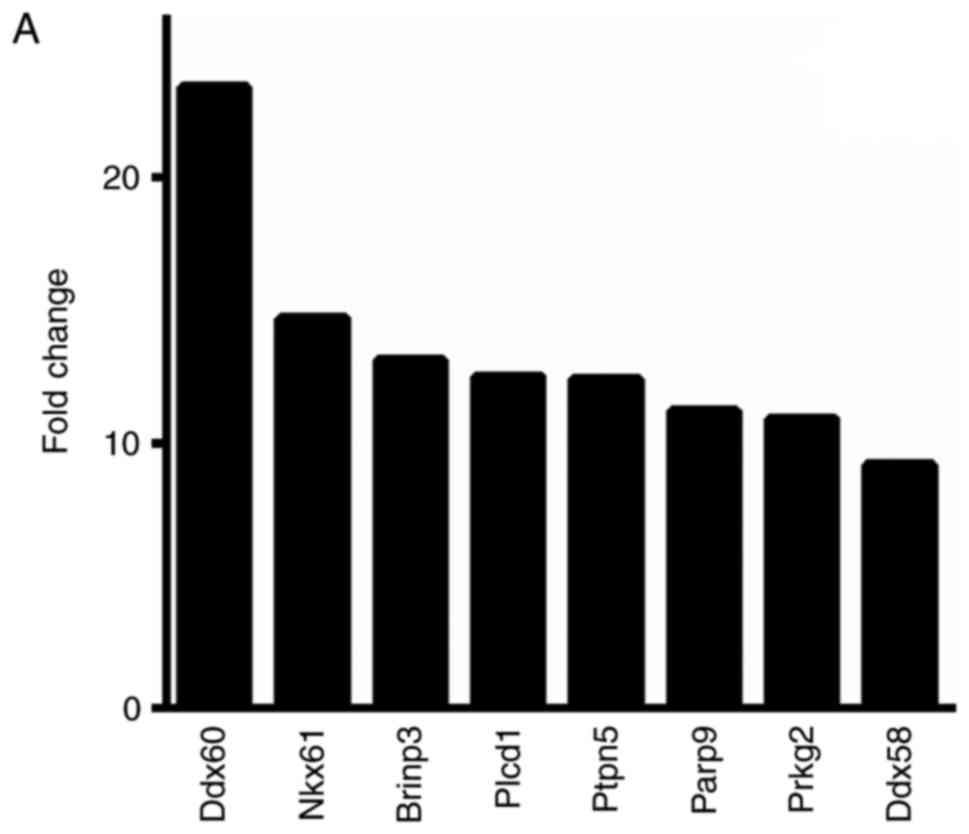
Microarray expression profiling analysis revealed
that, among the total 16,802 target genes identified, 8,547 (50.9%)
genes were upregulated in cells treated with the miR-146a inhibitor
and 8,255 (49.1%) genes were downregulated. A fold-change cut-off
value >2.0 was used to select for differential expression, which
identified 806 genes as upregulated in cells treated with the
miR-146a inhibitor, and of these, 8 genes exhibited a fold change
>10.0 (Fig. 3A). Among the
1,026 genes that were identified as downregulated in cells treated
with the miR-146a inhibitor, >2.0-fold change, 9 genes exhibited
a >10.0-fold change (Fig. 3B).
Kyoto Encyclopedia of Genes and Genome analysis identified that the
p53 signaling pathway was upregulated by miR-146a inhibitor
(P=0.019; enrichment score, 1.72).
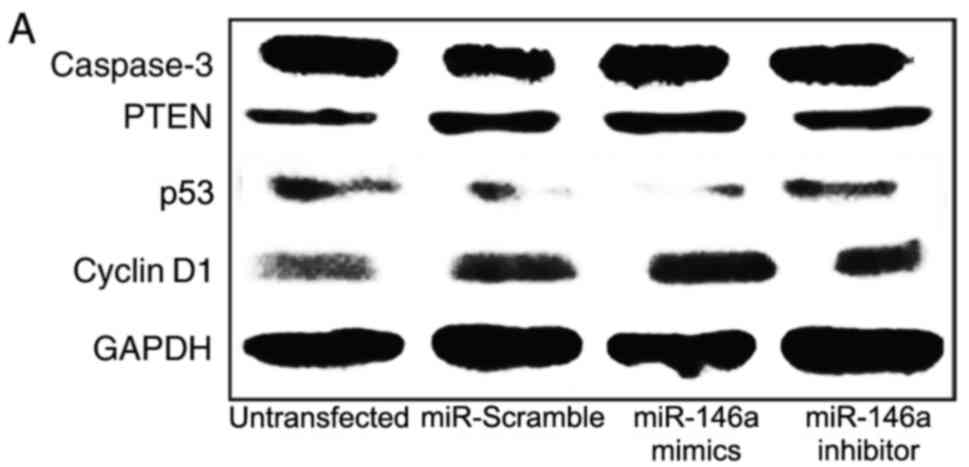
miR-146a downregulates p53
expression
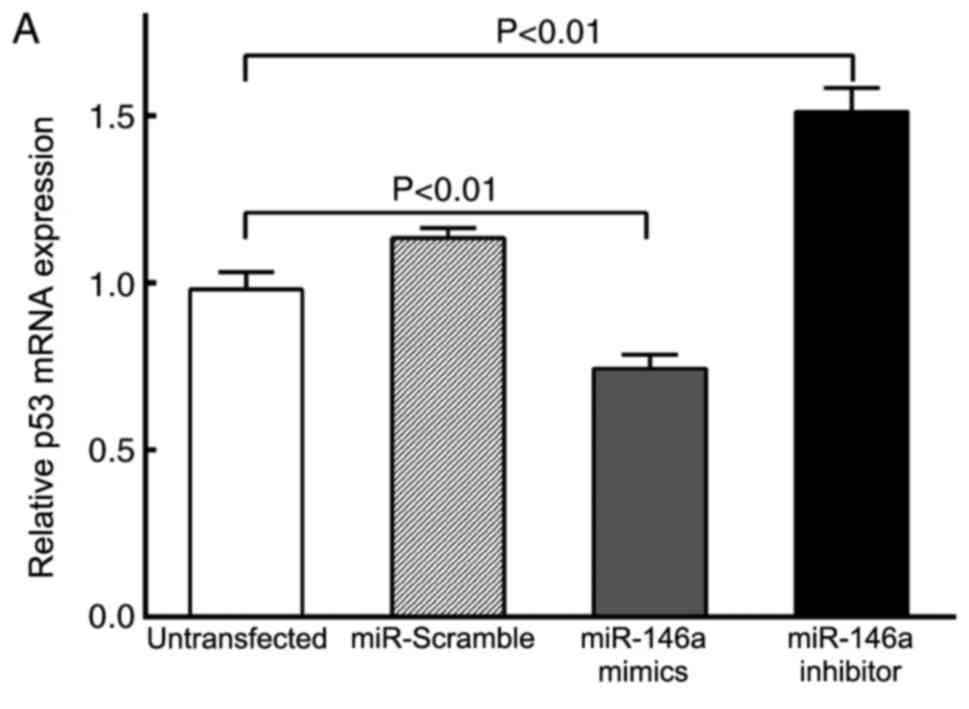
The key molecules in the p53 signaling pathway,
including p53, cyclin D1, caspase-3 and PTEN, were detected by
RT-qPCR and Western blot analysis (Figs. 4 and 5). The results demonstrated that there
were no significant differences in mRNA and protein expression
levels of caspase-3 and PTEN between the four experimental groups
(P>0.05; Figs. 4C, D and
5A). However, p53 mRNA and protein
expression levels were decreased in cells transfected with the
miR-146a mimics compared with the untransfected VSMC group
(P<0.01; Figs. 4A, 5A and B); p53 mRNA and protein expression
levels were increased in cells transfected with the miR-146a
inhibitor compared with untransfected VSMC cells (P<0.01).
Compared with the untransfected VSMC group, cyclin D1 mRNA and
protein expression levels were increased in miR-146a mimics
transfected cells (P<0.01; Figs.
4B, 5A and C), and were
decreased in the miR-146a inhibitor group (P<0.05).
Discussion
miRNAs are important and widespread gene regulators,
and serve important roles in physiological and pathological
processes, including cell growth, development, differentiation,
signal transduction, disease and death (3). Previous studies reported that miRNAs
may not only be used as diagnostic biomarkers of disease, but may
also be used as therapeutic targets for treating diseases (11,12).
miR-146a was reported as one of the major miRNAs
involved in the regulation of immune functions (13). It was also previously confirmed
that stimulation of monocytes by lipopolysaccharide caused an
increase in the expression of miR-146a. Tumor necrosis factor
(TNF-α) and interleukin (IL)-1β have been reported to promote
miR-146a expression through nuclear factor (NF)-κB-dependent
pathway (14). miR-146a was
revealed to negatively regulate inflammation and the immune
response by downregulating the expression levels of TNF
receptor-associated factor 6 and IL-1 receptor-associated kinase 1,
to avoid excessive inflammatory response (15). miR-146a was also reported to serve
an important role in the occurrence and development of rheumatoid
arthritis, cancer and sepsis, and may be used as a biomarker of the
diseases (16–18). A previous study reported that the
level of miR-146a in the peripheral blood mononuclear cells was
significantly increased in patients with acute coronary syndrome
(7), miR-146a expression
upregulated the function of Th1 cells, promoted Th1 cell
differentiation and NF-κB p65 synthesis, which indicated that
miR-146a may be associated with immune function regulation in
patients with coronary heart disease. The expression level of
miR-146a was revealed to be significantly upregulated in animal
models of restenosis following the balloon injury (8). ApoE serves an important role in lipid
metabolism and negatively regulates atherosclerosis (19). ApoE proteins were revealed to
inhibit NF-κB activation, inflammation and atherosclerosis by
promoting miR-146a expression in monocytes and macrophages
(9). miR-146a may aid in
predicting ventricular remodeling in patients with acute
ST-elevation myocardial infarction (20). Results from these previous studies
suggested that there may be a close correlation between miR-146a
expression and the development of atherosclerosis, coronary heart
disease and stenosis. However, these findings are not consistent,
as Guo et al (7) reported
that miR-146a was able to induce Th1 cells to synthesize NF-κB,
whereas Li et al (9)
demonstrated that in monocytes and macrophages, miR-146a inhibited
NF-κB expression.
Our previous study indicated that the levels of
miR-146a expression were significantly increased during VSMC
proliferation (10); following
miR-146a expression inhibition by RNA interference, VSMC
proliferation and migration were significantly reduced, and the
apoptosis was increased, but the underlying mechanism is unclear.
The dual-luciferase reporter assay is the classical method for
studying miRNA target genes; however, systematized study of
signaling pathways by this method is still lacking. p53 is a
classic tumor suppressor gene and a pro-apoptotic gene, p53 may not
only serve an important role in tumor growth and invasion, but is
also closely related to atherosclerosis and ventricular remodeling
(19). Induction of p53 expression
may inhibit VSMC proliferation (21) and p53-knockout may lead to
atherosclerosis (22). p53 may
directly or indirectly inhibit the expression of miR-146a in R6/2
rats (23). The cyclins and
cyclin-dependent kinase (cyclin/CDK) complex serves an important
role in the regulation of the G1/S transition phase; cyclin D1/CDK4
is an important regulatory protein (24). Cyclin D1 activation is closely
associated with VSMC proliferation and intima reconstruction, and
is an important target to promote VSMC proliferation (25). The present study demonstrated that
miR-146a expression may be able to induce VSMC proliferation by
downregulating the expression of the key tumor suppressor gene p53,
and by upregulating the expression of cyclin D1.
In summary, miR-146a may promote the proliferation
of rat VSMCs by downregulating p53 and upregulating cyclin D1
expression, which is related to vascular neointimal remodeling and
proliferative cardiovascular diseases. Further work is required to
better understand the role of miR-146a in the pathophysiological
processes of cardiovascular diseases and its possible use as
therapeutic agent.
Acknowledgements
The present study was supported by The Shenzhen
Health and Family Planning Commission foundation (grant no.
201505001).
References
|
1
|
Pasterkamp G, de Kleijn DP and Borst C:
Arterial remodeling in atherosclerosis, restenosis and after
alteration of blood flow: Potential mechanisms and clinical
implications. Cardiovasc Res. 45:843–852. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Davis-Dusenbery BN, Wu C and Hata A:
Micromanaging vascular smooth muscle cell differentiation and
phenotypic modulation. Arterioscler Thromb Vasc Biol. 31:2370–2377.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Small EM and Olson EN: Pervasive roles of
microRNAs in cardiovascular biology. Nature. 469:336–342. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fazi F and Nervi C: MicroRNA: Basic
mechanisms and transcriptional regulatory networks for cell fate
determination. Cardiovasc Res. 79:553–561. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chan EK, Ceribelli A and Satoh M:
MicroRNA-146a in autoimmunity and innate immune responses. Ann
Rheum Dis. 72 Suppl 2:ii90–ii95. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Guo M, Mao X, Ji Q, Lang M, Li S, Peng Y,
Zhou W, Xiong B and Zeng Q: MiR-146a in PBMCs modulates Th1
function in patients with acute coronary syndrome. Immunol Cell
Biol. 88:555–564. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ji R, Cheng Y, Yue J, Yang J, Liu X, Chen
H, Dean DB and Zhang C: MicroRNA expression signature and
antisense-mediated depletion reveal an essential role of microRNA
in vascular neointimal lesion formation. Circ Res. 100:1579–1588.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li K, Ching D, Luk FS and Raffai RL:
Apolipoprotein E enhances microRNA-146a in monocytes and
macrophages to suppress nuclear factor-κB-driven inflammation and
atherosclerosis. Circ Res. 117:e1–e11. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Dong S, Xiong W, Yuan J, Li J, Liu J and
Xu X: MiRNA-146a regulates the maturation and differentiation of
vascular smooth muscle cells by targeting NF-κB expression. Mol Med
Rep. 8:407–412. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Creemers EE, Tijsen AJ and Pinto YM:
Circulating microRNAs: Novel biomarkers and extracellular
communicators in cardiovascular disease? Circ Res. 110:483–495.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
van Rooij E and Olson EN: MicroRNA
therapeutics for cardiovascular disease: Opportunities and
obstacles. Nat Rev Drug Discov. 11:860–872. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li L, Chen XP and Li YJ: MicroRNA-146a and
human disease. Scand J Immunol. 71:227–231. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Taganov KD, Boldin MP, Chang KJ and
Baltimore D: NF-kappaB-dependent induction of microRNA miR-146, an
inhibitor targeted to signaling proteins of innate immune
responses. Proc Natl Acad Sci USA. 103:12481–12486. 2006;
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bhaumik D, Scott GK, Schokrpur S, Patil
CK, Campisi J and Benz CC: Expression of microRNA-146 suppresses
NF-kappaB activity with reduction of metastatic potential in breast
cancer cells. Oncogene. 27:5643–5647. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Pauley KM, Satoh M, Chan AL, Bubb MR,
Reeves WH and Chan EK: Upregulated miR-146a expression in
peripheral blood mononuclear cells from rheumatoid arthritis
patients. Arthritis Res Ther. 10:R1012008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pacifico F, Crescenzi E, Mellone S,
Iannetti A, Porrino N, Liguoro D, Moscato F, Grieco M, Formisano S
and Leonardi A: Nuclear factor-{kappa}B contributes to anaplastic
thyroid carcinomas through up-regulation of miR-146a. J Clin
Endocrinol Metab. 95:1421–1430. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang JF, Yu ML, Yu G, Bian JJ, Deng XM,
Wan XJ and Zhu KM: Serum miR-146a and miR-223 as potential new
biomarkers for sepsis. Biochem Biophys Res Commun. 394:184–188.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Curtiss LK and Boisvert WA: Apolipoprotein
E and atherosclerosis. Curr Opin Lipidol. 11:243–251. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu X, Dong Y, Chen S, Zhang G, Zhang M,
Gong Y and Li X: Circulating MicroRNA-146a and microRNA-21 predict
left ventricular remodeling after ST-elevation myocardial
infarction. Cardiology. 132:233–241. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wassmann S1, Wassmann K, Jung A, Velten M,
Knuefermann P, Petoumenos V, Becher U, Werner C, Mueller C and
Nickenig G: Induction of p53 by GKLF is essential for inhibition of
proliferation of vascular smooth muscle cells. J Mol Cell Cardiol.
43:301–307. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Guevara NV, Kim HS, Antonova EI and Chan
L: The absence of p53 accelerates atherosclerosis by increasing
cell proliferation in vivo. Nat Med. 5:335–339. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ghose J, Sinha M, Das E, Jana NR and
Bhattacharyya NP: Regulation of miR-146a by RelA/NFκB and p53 in
STHdh(Q111)/Hdh(Q111) cells, a cell model of Huntington's disease.
PLoS One. 6:e238372011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pestell RG: New roles of cyclin D1. Am J
Pathol. 183:3–9. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim MH, Ham O, Lee SY, Choi E, Lee CY,
Park JH, Lee J, Seo HH, Seung M, Choi E, et al: MicroRNA-365
inhibits the proliferation of vascular smooth muscle cells by
targeting cyclin D1. J Cell Biochem. 115:1752–1761. 2014.
View Article : Google Scholar : PubMed/NCBI
|