Introduction
Cervical cancer is a disease with poor prognosis
among women worldwide (1–3). Surgery assessment of local disease
state, including tumor size assessment and vaginal and parametrial
involvement, is good during early stages of the disease; however,
the evaluation of tumor size and vaginal and parametrial
involvement are difficult in advanced- and late-stages, such as in
those patients who do not qualify for up-front surgery (4,5).
Owing to such limitations, alternative approaches and more
efficient remedies are necessary to treat cervical cancer other
than surgical operation, chemotherapy and vaccination.
Previous studies have reported on the use of
traditional herbal medicines as alternative cancer therapies
(6–8), including a number of studies on HeLa
cervical cancer cells that used herbal medicines as therapeutic
agents and compounds for potentially treating cervical cancer
(9–11). Our previous study developed a new
herbal medicine, called SH003, which is composed of Astragalus
membranaceus (Am), Angelica gigas (Ag) and
Trichosanthes kirilowii Maximowicz (Tk), based on the
principles of traditional Chinese medicine (12,13).
Am and Ag have been reported to be effective treatments for a
number of diseases, including hematologic diseases, endocrine
disorders and cancers (14–19).
Therapeutic effects of Tk have also been indicated for various
cancer types, such as leukemia, hepatocellular carcinoma, colon
cancer, non-small cell lung cancer and gastric cancer (18,20–25).
Several previous studies have demonstrated that a mixture of those
components named SH003 was better therapeutic effect than each
component in other types of cancer (12,13,17,26–30).
Thus, SH003 is likely to be used for cancer treatment as novel
herbal mixture.
Apoptosis, or programmed cell death, is
characterized by cell shrinkage, membrane blebbing, maintenance of
organelle integrity, and DNA condensation and fragmentation
(31). There are two subtypes of
the apoptotic pathway: The extrinsic, or receptor-mediated, pathway
and the intrinsic, or mitochondria-mediated, pathway (32). The extrinsic apoptotic pathway
involves the so-called ‘death receptors’, such as Fas receptor,
tumor necrosis factor receptor and death receptor, and their
associated extracellular ligands (33–35).
These apoptotic factors lead to the activation of caspase (casp)-3
and casp8 (35,36). Conversely, the intrinsic apoptotic
pathway is activated by a number of different factors, such as
genomic and metabolic stresses, and the presence of unfolded
proteins, which may cause permeabilization of the outer
mitochondrial membrane and the release apoptotic proteins into the
cytosol (34). Concurrently, cell
cycle checkpoints are triggered and apoptosis is induced in cells
exhibiting abnormal cell cycle. In particular, the G1
phase is regulated by cyclin D proteins (including cyclin
D1, D2 and D3), cyclin E proteins
which associate with cyclin-dependent kinase (CDK) 2 to regulate
thr progression from G1 into S phase, CDK2, CDK4 and
CDK6 (37). The CDKs are a family
of serine/threonine protein kinases that are activated at the
G1 checkpoint; Cyclin D proteins interact with
CDK4/CDK6, and these complexes directly regulate the G1
phase of cell cycle.
The present study investigated the anticancer
effects of SH003 in the HeLa cervical cancer cell line. The results
demonstrated that SH003 treatment reduced the viability of HeLa
cells without affecting that of normal cells. Rat intestinal
epithelial cells were used as the control group in the present
study, as previously described (38). SH003 treatment led to extrinsic
apoptosis and induced the cleavage of casp3, casp8 and
poly(ADP-ribose) polymerase 1. SH003 treatment also increased the
production of reactive oxygen species (ROS) and induced G1 cell
cycle arrest in HeLa cells. Results from the present study
suggested that SH003 may be a potential treatment for cervical
cancer.
Materials and methods
SH003 preparation
The production of SH003 and its components, Am, Ag
and Tk, was commissioned to Hanpoong Pharm and Foods Company
(Jeonju, Korea), and the extraction procedures were performed as
previously reported (12).
Briefly, extracts (~333 g each) of Am, Ag and Tk in 10 l 30%
ethanol were obtained by boiling for 3 h at temperature ranging
between 80 and 100°C. The ethanol extract was filtered,
concentrated using vacuum evaporation and freeze-dried at <60°C
(average yield=35.5%). Dried extract was then dissolved in 30%
ethanol to prepare a stock solution of 100 mg/ml. The stock
solution was stored at −80°C until used.
Cell culture
HeLa human cervical cancer cells (Korean Cell Line
Bank, Seoul, Korea) and rat intestinal epithelium (RIE; provided by
Dr Joon Woo Lee; Seoul National University, Seoul, Korea) were
cultured in Dulbecco's Modified Eagle's Medium (DMEM; Welgene,
Gyeongsan, Korea) containing 10% fetal bovine serum (Welgene) and
1% penicillin. Cells were maintained at 37°C in a 5% CO2
atmosphere.
Reagents
Methyl thiazolyl tetrazolium (MTT),
2′,7′-dichlorofluorescin diacetate (DCFH-DA), N-acetyl L-cysteine
(NAC) and 7-aminoactinomycin D (7-AAD) were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
Acrylamide/bis-acrylamide solution (30%) was purchased from Bio-Rad
Laboratories, Inc. (Hercules, CA, USA). Annexin V-fluorescein
isothiocyanate (FITC) was purchased from BD Biosciences (San Jose,
CA, USA), and
5,5′,6,6′-tetrachloro-1,1′,3,3′tetraethylbenzimidazolylcarbocyanine
(JC-1) was purchased from Biotium Inc. (Freemont, CA, USA).
Western blotting
Radioimmunoprecipitation assay lysis buffer
containing protease inhibitors (Dithiothreitol 3 mg/ml,
Phenylmethylsulfonyl fluoride 3 µg/ml, Na3VO4
5 µg/ml, NaF 5 µg/ml and protease inhibitor 4 µg/ml) was used for
cell lysis for 20 min at 4°C, and supernatants were collected by
centrifugation at 16,600 × g for 20 min at 4°C. Protein
concentration was measured using a Bradford assay kit (Bio-Rad
Laboratories, Inc.). Proteins (15 µg) were separated by 8–12%
SDS-PAGE and transferred to nitrocellulose membranes. Membranes
were blocked at room temperature for 1 h in 2% skim milk containing
3% bovine serum albumin (Sigma-Aldrich; Merck KGaA) and probed with
the primary antibodies raised against the following proteins at 4°C
overnight: Cleaved (C)-casp3 (cat no. 9661; 1:2,000), C-casp8 (cat
no. 9661; 1:2,000), casp9 (cat no. 9502; 1:50,000),
poly(ADP-ribose) polymerase 1 (PARP1; cat no. 9542; 1:50,000),
phosphorylated (p)-p53 (cat no. 9284; 1:50,000), cyclin-dependent
kinase (CDK) 4 (cat no. 12790s; 1:50,000) and CDK6 (cat no. 3136;
1:50,000), all from Cell Signaling Technology, Inc. (Danvers, MA,
USA); β-actin (cat no. sc47778; 1:50,000), Bcl-2 (cat no. sc7382;
1:10,000), Bcl-2-like protein (Bax; cat no. sc-7480; 1:100,000),
CDK2 (cat no. sc163; 1:100,000) and cyclin E (cat no. sc-247;
1:5,000) all from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA);
cyclin D1 (cat no. 556470; 1:50,000) and retinoblastoma-associated
protein (pRb; cat no. 554140; 1:50,000; from BD Biosciences.
Membranes were then incubated in room temperature for 1 h with the
following horseradish peroxidase-conjugated secondary antibodies
(1:1,000): Anti-rabbit immunoglobulin (Ig) G (cat no. 7074) and
anti-mouse IgG (cat no. 7076) from Cell Signaling Technology, Inc.
Protein bands were visualized using enhanced chemiluminescence (cat
no. DG-W250; DoGEN, Seoul, Korea). Blots were semi-quantified by
densitometry using ImageJ software version 1.4.3.67 (National
Institutes of Health, Bethesda, MD, USA) and normalized to
β-actin.
Cell viability assays
HeLa (5×103) and RIE (4×103)
cells were seeded in DMEM in 96-well plates and treated with
various concentrations (0, 100, 200 or 400 µg/ml) of either SH003
or one of the individual components (Am, Ag or Tk) for 72 h at
37°C. Subsequently, the medium was discarded and 10 µl MTT solution
(Sigma-Aldrich; Merck KGaA) in 90 µl medium was added to each well.
Cells were incubated for 2 h at 37°C, and then 100 µl dimethyl
sulfoxide (DMSO) was added to dissolve the formazan crystals. The
absorbance of each sample was measured at 570 nm using an ELISA
microplate reader (Molecular Devices, LLC, Sunnyvale, CA, USA).
Mitochondrial membrane potential
assay
HeLa cells (3×105) were seeded on 60 mm
dishes and treated with 100, 200 or 400 µg/ml SH003 for 24 h.
Control cells received no treatment. Briefly, 100X JC-1 solution
was diluted in to 1X in DMSO. Cells were stained with 4 µg/ml JC-1
solution for 15 min at 37°C. In healthy cells, JC-1 accumulates as
J-aggregates (FL-2) in the mitochondria and stains them red,
whereas in apoptotic cells, the mitochondrial membrane potential
collapses and JC-1 remains in the cytoplasm as a monomer (FL-1),
emitting green fluorescence. Cells were analyzed using a
FACSCalibur Flow Cytometry System (BD Biosciences). Histograms and
dot plots were analyzed with CellQuestPro software version 5.2 (BD
Biosciences). The M1 gate was calculated based on the control
values (no treatment).
ROS generation assay
ROS generation was detected using the dye DCFH-DA.
Briefly, HeLa cells (3×105) were plated on 60-mm plates
and incubated in DMEM at 37°C overnight. Cells were treated with
various concentrations of SH003 (0, 100, 200 and 400 µg/ml), with
or without NAC (2 mM). Subsequently, 1 µl/ml DCFH-DA was added and
cells were incubated for 1 h at 37°C and samples were analyzed
using the FACSCalibur Flow Cytometry System (BD Biosciences).
Histograms and dot plots were analyzed using CellQuestPro Software
version 5.2 (BD Biosciences). NAC (2 mM) was used for inhibition of
ROS generation.
Annexin V-FITC/7-AAD apoptosis
assay
Rates of apoptosis were measured by annexin
V-FITC/7-AAD assay. HeLa cells (3×105) were treated with
400 µg/ml SH003 for 48 h and 2 mM NAC for 2 h at 37°C, and then
stained with 1.25 µl annexin V-FITC in 1X binding buffer and 1 µl
7-AAD for 15 min at room temperature in the dark. Apoptotic cell
population was analyzed by FACSCalibur Flow Cytometry System,
measuring the signal by FL-1 (green, 525 nm) and FL-3 (red, 620–767
nm). Cells were analyzed using a FACSCalibur Flow Cytometry System
(BD Biosciences). Histograms and dot plots were analyzed with
CellQuestPro software version 5.2 (BD Biosciences). The M1 gate was
calculated based on the control values (no treatment).
Cell cycle assay
HeLa cells were plated in DMEM in 60-mm dishes at a
density of 7×105 cells/dish and treated with various
concentrations of SH003 (0, 100, 200 and 400 µg/ml) for 24 h at
37°C. Subsequently, cells were harvested, centrifuged at 221 × g
for 10 min at 4°C, fixed with 95% ethanol at 4°C overnight, and
washed twice in PBS. Subsequently, cells were stained with 50 µl
Muse Cell Cycle Assay kit (Merck KGaA) in the dark at room
temperature for 30 min. Cell cycle plots were analyzed using the
Muse Cell Analyzer (Merck KGaA).
Statistical analysis
Statistical analysis was performed using SPSS
software version 22.0 (IBM Corp., Armonk, NY, USA). Data are
presented as the mean ± standard deviation of 3 independent
experiments. The statistical significance of the differences
between groups was assessed using one-way analysis of variance
followed by a post hoc Tukey test for multiple comparisons.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effects of SH003 and its components on
RIE and HeLa cell viability
An MTT assay was performed to evaluate the cytotoxic
effects of Am, Ag, Tk and SH003 on normal RIE cells and HeLa
cervical cancer cells. RIE cells were treated with varying
concentrations (100, 200 and 400 µg/ml) of each extract or SH003
(Fig. 1A). While SH003 and Am
treatment exhibited lower toxicity on RIE cells compared with Ag
and Tk, SH003 appeared to be more effective at inducing cell death
in HeLa cells compared with Am (Fig.
1A). Conversely, SH003 treatment reduced HeLa cell viability at
all concentrations (Fig. 1B),
which suggested that SH003 may induce toxicity in cervical cancer
cells without affecting normal cell viability.
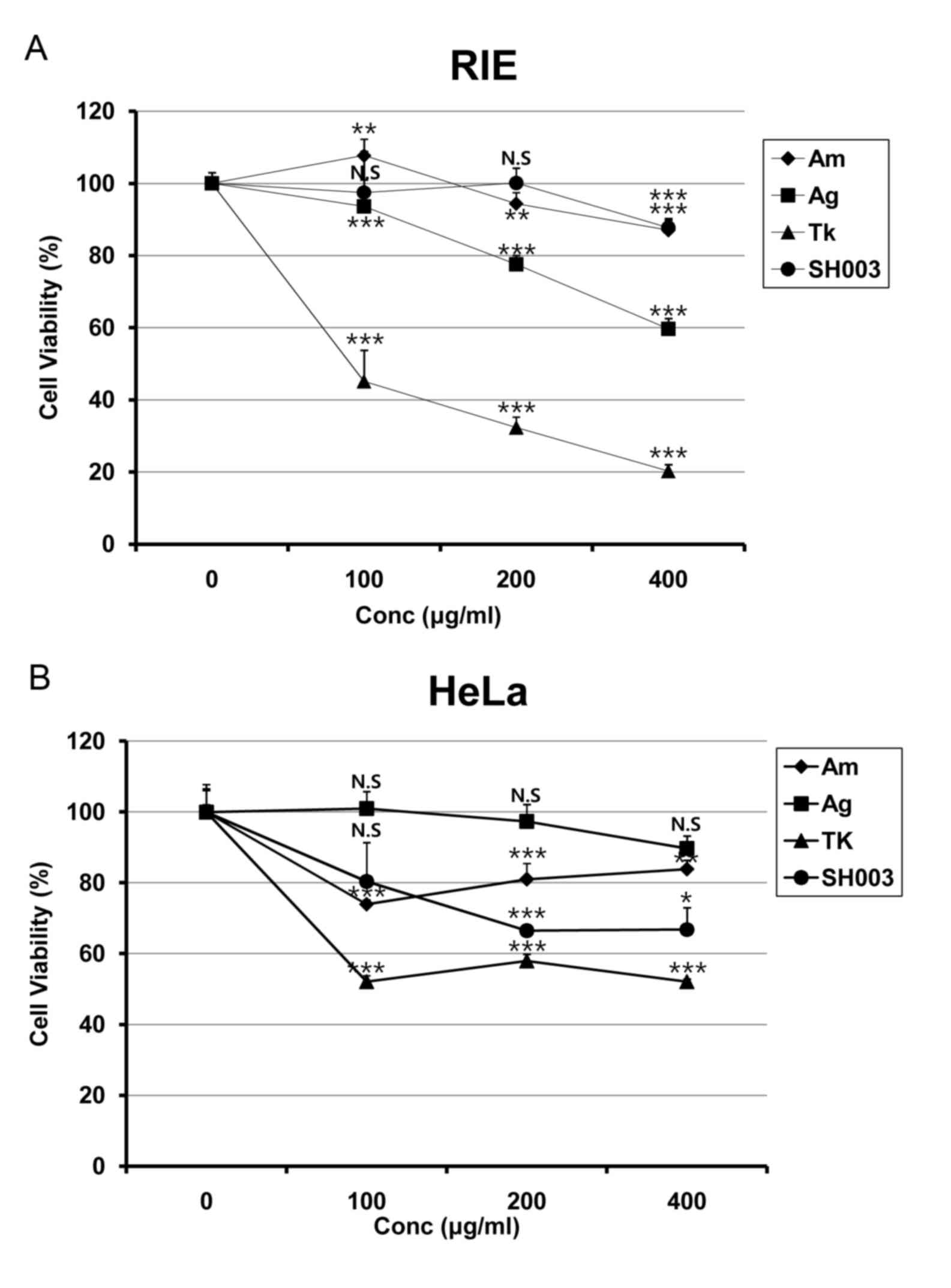 | Figure 1.SH003 treatment reduces viability of
HeLa cells. (A) RIE and (B) and HeLa cells were seeded in 96-well
plates. Following 24-h incubation, cells were treated with various
concentrations (0, 100, 200 and 400 µg/ml) of Am, Ag, Tk or SH003
for 72 h, followed by MTT assay analysis. Cells were treated with
MTT solution for 2 h, followed by 100 µl of DMSO. Cell viability
was analyzed by an ELISA microplate reader. Experiments were
performed in triplicate, and results are presented as the mean ±
standard deviation. *P<0.05, **P<0.01 and ***P<0.001 vs.
control (0 µg/ml). Ag, Angelica gigas; Am, Astragalus
membranaceus; Conc., concentration; MTT, methyl thiazolyl
tetrazolium; RIE, rat intestinal epithelium; Tk, Trichosanthes
kirilowii Maximowicz. |
SH003 induces apoptosis in HeLa
cells
SH003-induced apoptosis was examined in HeLa cells.
Cells were stained with annexin V-FITC and 7-AAD and analyzed by
flow cytometry. SH003 treatments at 200 and 400 µg/ml were revealed
to significantly increase the percentage of cells in the early and
late apoptosis regions (Fig. 2A).
In accordance with these data, SH003 treatment appeared to increase
the protein expression levels of C-casp3, C-casp8 and PARP1, but
not C-casp9 (Fig. 2B), which
indicated that SH003 treatment may induce extrinsic apoptotic cell
death in HeLa cells. However, these results were not statistically
significant.
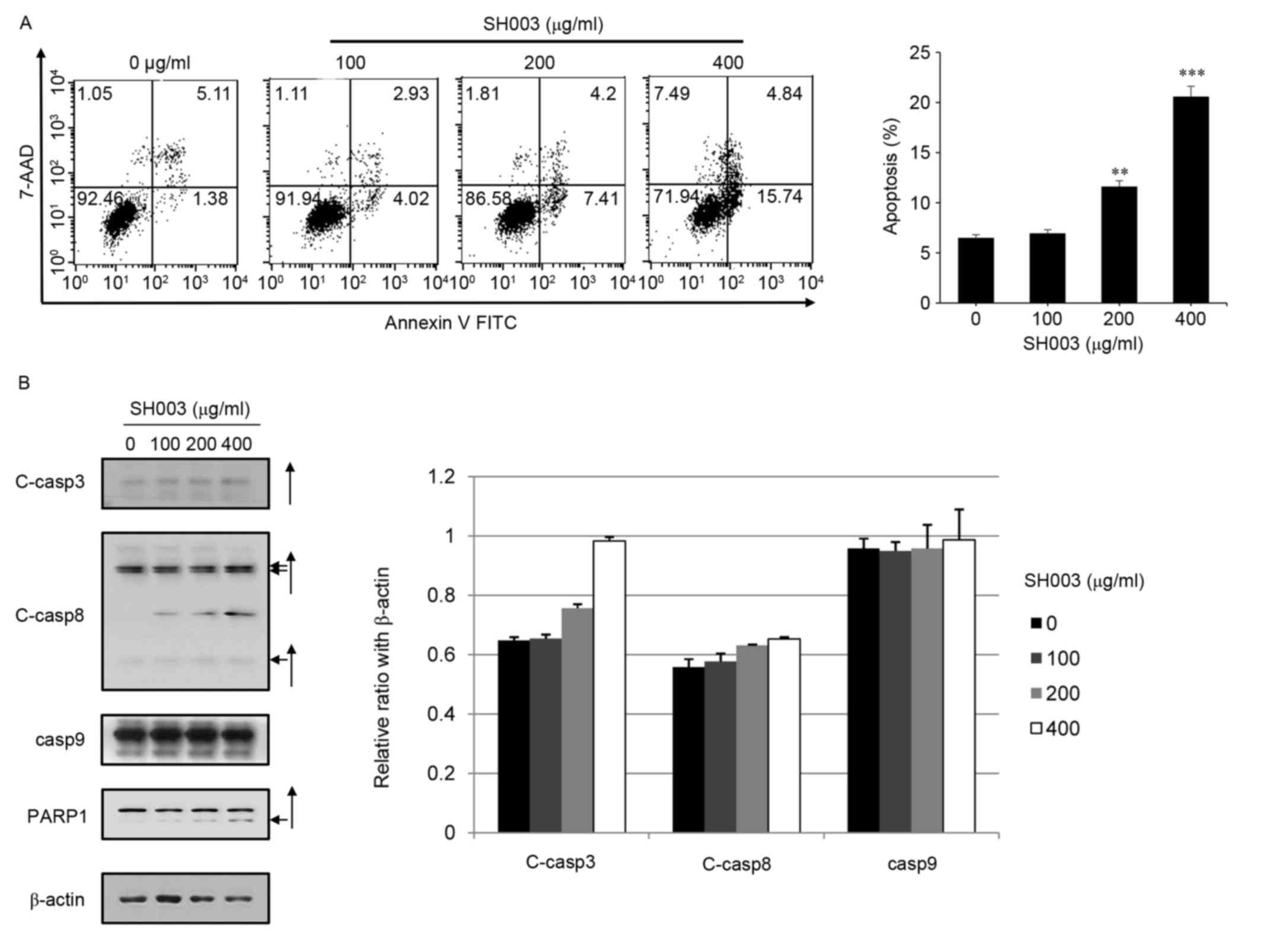 | Figure 2.SH003 treatment induces apoptosis in
HeLa cells. (A) HeLa cells (1×105 cells/dish) were
seeded on 60 mm dishes and treated with 0, 100, 200 or 400 µg/ml
SH003 for 48 h. Cells were harvested and stained with 7-AAD and
annexin V-FITC in 1X binding buffer. Apoptotic cell death was
analyzed by FACSCalibur flow cytometer FL-1 and FL-3 channels.
Experiments were performed in triplicate, and results are presented
as the mean ± standard deviation; **P<0.01 and ***P<0.001 vs.
control (0 µg/ml). (B) Total cell lysates were prepared and the
equal amounts of protein (15 µg) were separated by SDS-PAGE, and
the membranes were probed with C-casp3, C-casp8, C-casp9 and PARP1
antibodies. β-Actin was used as a loading control. Arrows indicate
the cleaved forms of the proteins. 7-AAD, 7-aminoactinomycin D;
C-casp, cleaved caspase; FITC, fluorescein isothiocyanate; PARP1,
poly(ADP-ribose) polymerase 1. |
SH003 does not affect the intrinsic
mitochondria-mediated apoptosis pathway in HeLa cells
As SH003 treatment did not appear to affect the
expression level of C-casp9 (Fig.
2B), the effects of SH003 on the intrinsic
mitochondria-mediated apoptosis pathway were examined by
measurement of the Bax/Bcl-2 ratio, which is important in the
intrinsic apoptosis pathway. No significant differences in
Bax/Bcl-2 ratios were indicted for HeLa cells exposed to SH003,
which suggested that SH003 may not induce intrinsic apoptosis
(Fig. 3A). In addition, changes in
mitochondrial transmembrane potential within the cells were
analyzed by staining HeLa cells with JC-1 and measuring the
monomeric form (via channel FL-1) and J-aggregates (via FL-2).
SH003 did not induce a low mitochondrial transmembrane potential,
suggesting that SH003 does may not induce apoptosis through the
mitochondria-mediated pathway (Fig.
3B), but through the extrinsic pathway.
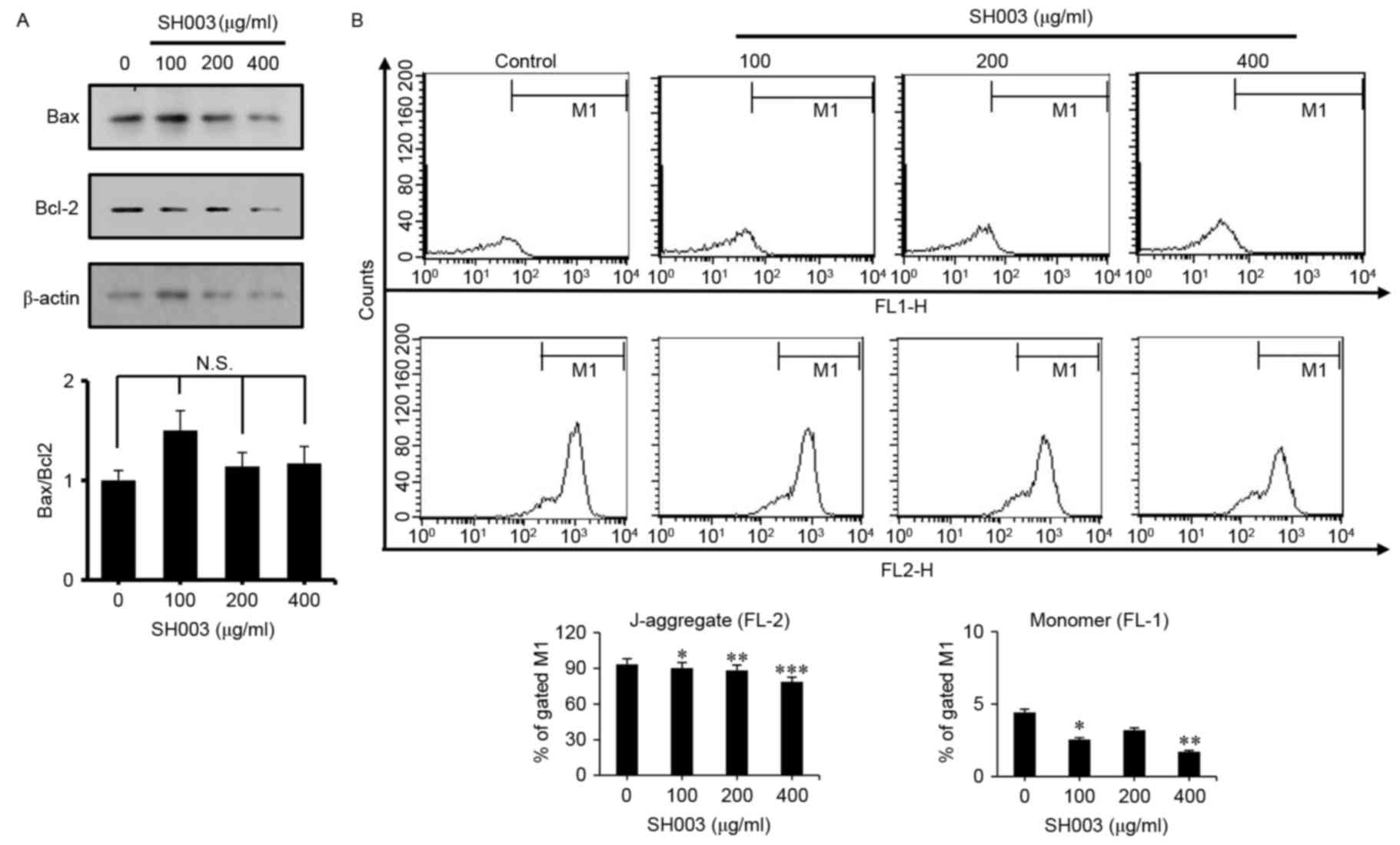 | Figure 3.SH003 treatment does not affect
intrinsic mitochondria-mediated apoptosis. HeLa cells were treated
with 100, 200 and 400 µg/ml of SH003 for 24 h prior to testing. (A)
Western blotting was performed by using anti-Bax and anti-Bcl-2
antibodies; β-actin was used as a loading control. Bax/Bcl-2 ratio
was calculated by ImageJ software, following densitometric
analysis. (B) Following SH003 treatment, cells were harvested and
stained with JC-1, and mitochondrial membrane potentials were
detected by FACSCalibur flow cytometry using FL-1 and FL-2 channels
to detect JC-1 monomers and J-aggregates, respectively. Experiments
were performed three times, and results are presented as the mean ±
standard deviation; *P<0.05, **P<0.01 and ***P<0.001 vs.
untreated control. Bcl, B-cell lymphoma; Bax, Bcl-2-like protein;
JC-1,
5,5′,6,6′-tetrachloro-1,1′,3,3′tetraethylbenzimidazolylcarbocyanine;
N.S., not significant. |
SH003 increases ROS generation in HeLa
cells
Elevated ROS levels have previously been implicated
in apoptosis (39–41). To investigate whether SH003
treatment influenced ROS generation, HeLa cells were stained with
DCFH-DA and analyzed by flow cytometry. The varying concentrations
of SH003 treatment significantly increased ROS generation in a
dose-dependent manner (Fig. 4A).
However, SH003-induced apoptosis was not related with an increase
in ROS levels, as co-treatment with SH003 and NAC resulted in
increased apoptosis (Fig. 4B) and
reduced cell viability compared with SH003 alone (Fig. 4C). These results indicated that
apoptosis induced by SH003 is not dependent on the ROS-mediated
pathway.
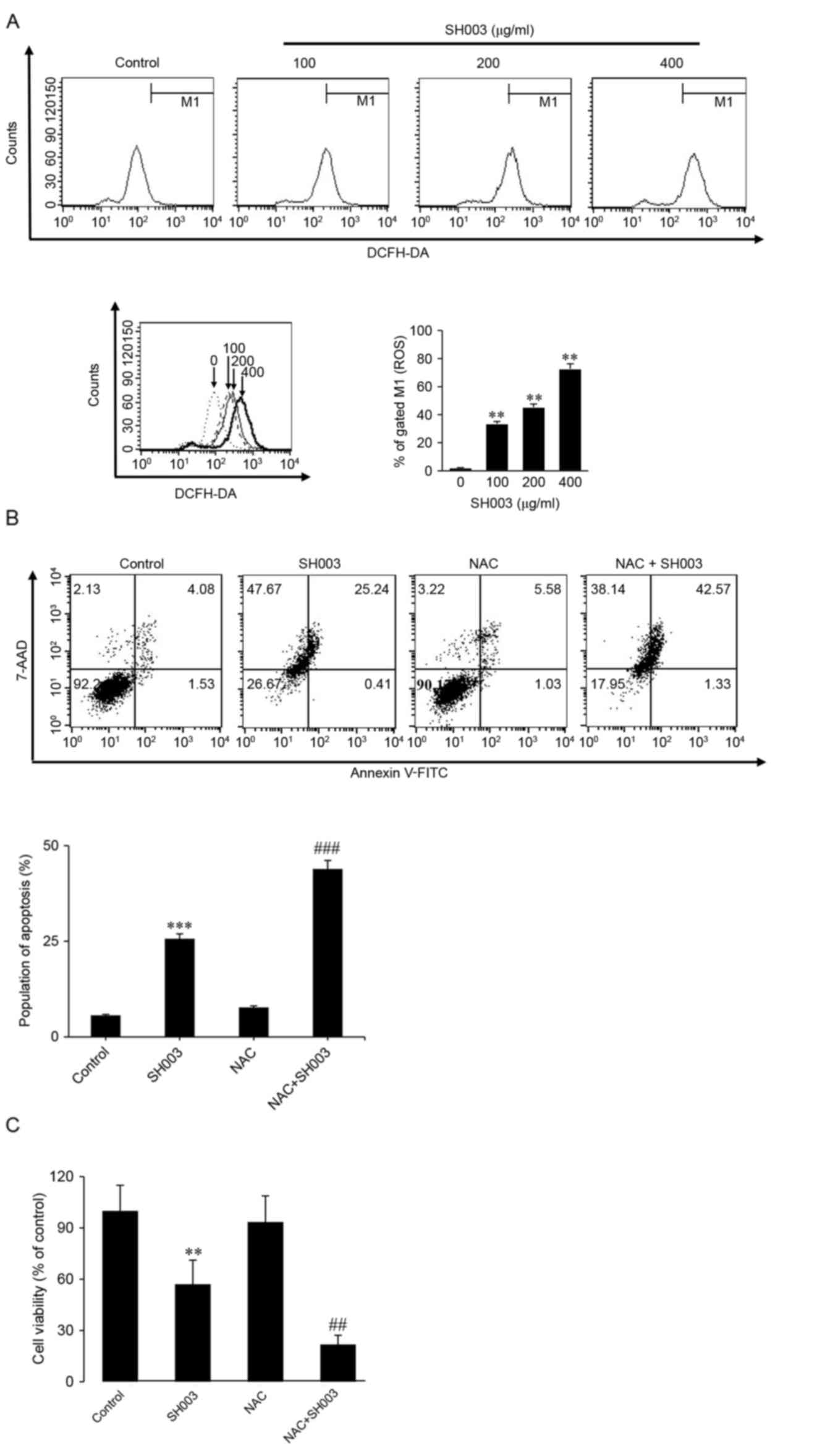 | Figure 4.SH003 treatment increases ROS
generation in HeLa cells. (A) HeLa cells were treated with various
concentrations (0, 100, 200 and 400 µg/ml) of SH003 and incubated
with DCFH-DA for 1 h. ROS generation of HeLa cells was measured by
FACSCalibur flow cytometry. **P<0.01 vs. untreated control. (B)
Cells were pretreated with 2 mM NAC for 30 min, followed by
treatment with concentrations of SH003 for 48 h. Cells were stained
with DCFH-DA for 1 h, then stained with 7-AAD and annexin V-FITC in
1X binding buffer. Apoptotic cell death was analyzed by FACSCalibur
flow cytometer. ***P<0.001 vs. untreated control;
###P<0.001 vs. SH003 treatment only. (C) HeLa cells
were pretreated with 2 mM NAC for 30 min, followed by treatment
with SH003 for 48 h. MTT solution was added to the cells and
incubated for 2 h, followed by the addition of 100 µl of DMSO to
each well. Cell viability was analyzed by ELISA microplate reader.
**P<0.01 vs. untreated control; ##P<0.01 vs. SH003
treatment only. All data are presented as the mean ± and standard
deviation of 3 independent experiments. DCFH-DA,
2′,7′-dichlorofluorescin diacetate; FITC, fluorescein
isothiocyanate; MTT, methyl thiazolyl tetrazolium; NAC, N-acetyl
L-cysteine; ROS, reactive oxygen species. |
SH003 induces G1 cell cycle
arrest in HeLa cells
Chemotherapy commonly inhibits cancer growth by
inducing cell cycle arrests (42–44)
and the present findings indicated that SH003 may have potential to
induce cell cycle arrest in cancer cells. Western blot analysis was
conducted to investigate whether SH003 induced cell cycle arrest in
HeLa cells. SH003 treatment was demonstrated to decrease the
protein expression levels of CDK2, CDK4, CDK6 and cyclin D. In
addition, SH003 treatment decreased p-p53 and pRb levels (Fig. 5A). In addition, cell cycle analysis
was performed with a Muse Cell Analyzer. The number of cells at the
G1 phase increased in HeLa cultures treated with varying
concentrations of SH003 (Fig. 5B).
These results indicated that SH003 may reduce G1 phase
cell cycle associated protein expression and arrests the cell cycle
at the G1 phase.
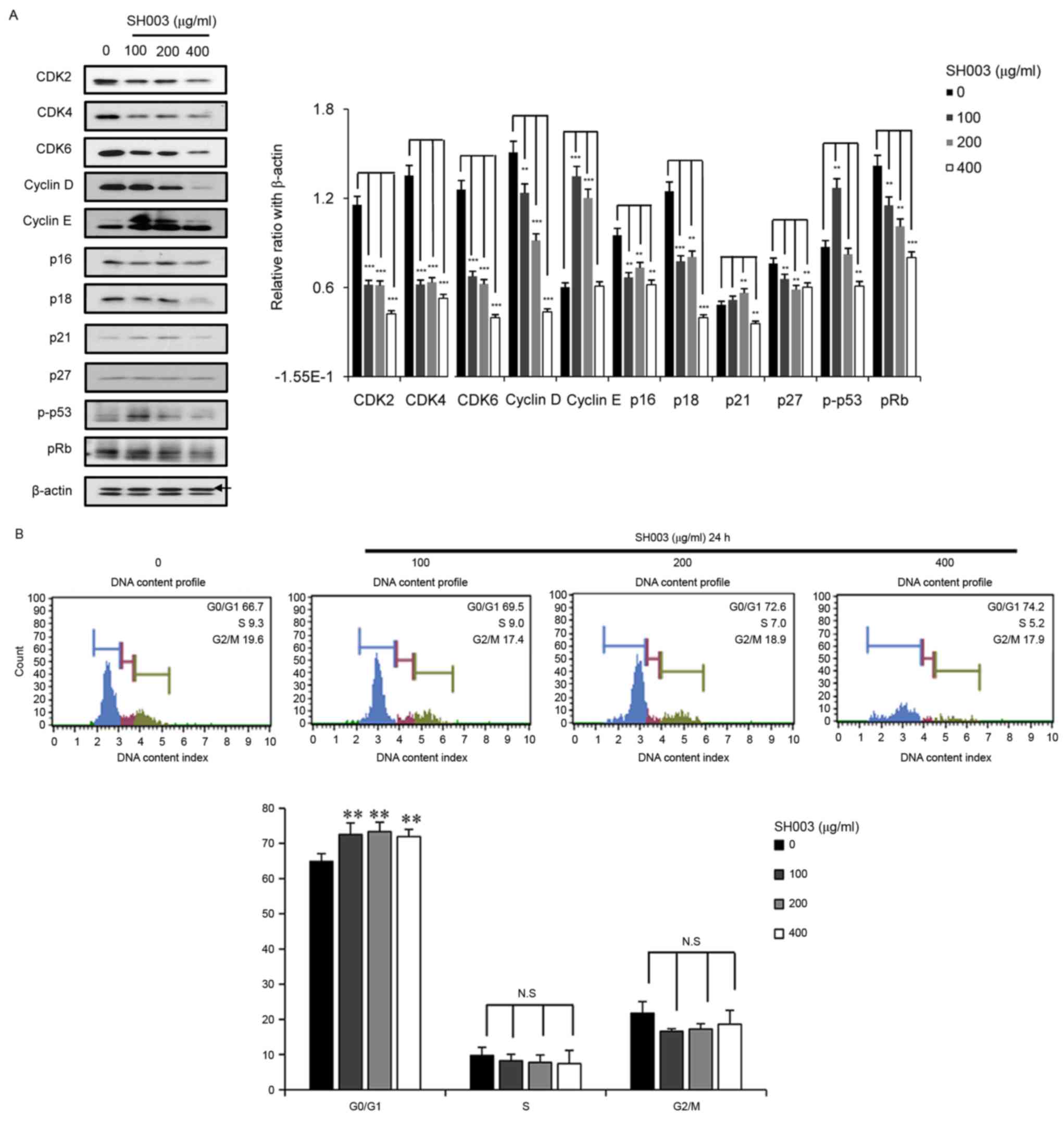 | Figure 5.SH003 induces G1 cell
cycle arrest in HeLa cells. (A) Cells were treated with varying
concentrations (0, 100, 200 and 400 µg/ml) of SH003 for 24 h,
harvested and used for western blot analysis. Each sample was
separated by SDS-PAGE, and then transferred to a membrane. The
membrane was probed with antibodies against CDK2, CDK4, CDK6,
cyclin D, cyclin E, p-p53 and pRb. β-Actin (upper band) was used as
a loading control. Cell cycle associated protein levels were
calculated by densitometric analysis using ImageJ software. (B)
Cells were plated 7×105 and treated with varying
concentrations of SH003 for 24 h. Cells were harvested, fixed in
95% ethanol and stained with 50 µl of Muse Cell Cycle Assay
reagent; cell cycle profiles were analyzed using Muse Cell
Analyzer. Experiments were performed in triplicate, and results are
presented as the mean ± standard deviation. **P<0.01 and
***P<0.001 vs. untreated control. CDK, cyclin dependent kinase;
N.S., not significant; p, phosphorylated; pRB,
retinoblastoma-associated protein. |
Discussion
As therapeutic agents without toxic side effects,
traditional herbal medicines have been the focus of several studies
on cancer (45,46). One such traditional herbal
medicine, SH003, has been reported previously as a potential novel
anticancer agent for breast and prostate cancer (12). In the present study, SH003
treatment was demonstrated to induce extrinsic apoptosis in HeLa
cervical cancer cells.
Conventional chemotherapeutic agents for cancer
treatment commonly induce apoptotic death (9–11),
and traditional herbal medicines have exhibited anticancer effects
against cervical cancer cells by promoting apoptosis (9–11).
Previous studies have demonstrated that SH003 treatment suppressed
the growth of breast and prostate cancer cells by inducing
apoptosis (12,47,48).
It has been also reported that SH003 co-treatment with doxorubicin
may have synergistic effects on the promotion of apoptotic death in
MDA-MB-231 human breast cancer cells (49). Consistent with these previous
results, the present study demonstrated that SH003-induced growth
inhibition of HeLa cells may be mediated by the induction of
apoptosis.
SH003 comprises Am, Ag and Tk, which have previously
been indicated as effective treatments for a variety of diseases,
such as hematologic diseases, endocrine disorders, leukemia,
hepatocellular carcinoma, colon cancer, non-small cell lung cancer
and gastric cancer (14–25). Although the mechanism of action
remains unclear, the mixture of Ag, Am and Tk may complement the
individual properties of each component. Previous studies have
demonstrated that SH003 may be more effective to inhibit the growth
of breast cancer cells compared with the effects of the separate
components (12,48,49).
Consistent with these previous results, the present data indicated
that SH003 treatment exhibited better effects on the inhibition of
HeLa cancer cell growth than its constituents' effect. In addition,
SH003 appeared to be less toxic to normal RIE cells compared with
cells treated with Am, Ag or Tk alone, which suggested that SH003
may be a possible treatment for cervical cancer without serious
adverse effects. However, further studies, investigating the
effects of SH003 and its components on apoptosis, ROS levels and
cell cycle arrest on normal cervical cells, are required in order
to establish the safety profile of SH003.
Cell cycle entry is critical for homeostatic control
in the cell (31); however, when
cells suffer from cellular damage, cell cycle arrest is induced by
various mechanisms such as the inhibition of cyclin and CDK
expression. These regulations are commonly referred to as cell
cycle checkpoint (50). In the
present study, SH003 treatment reduced the levels of expression for
G1 phase associated CDKs (CDK2, CDK4 and CDK6) and
cyclin D. Although the present data indicated that inhibition of
cell growth by SH003 is mediated through cell cycle arrest at
G1 phase, further study should be performed to determine
the mechanisms of CDK inhibition.
Increased ROS production has been associated with
promoting apoptosis (40,51,52).
In the present study, SH003 treatment increased ROS production in a
dose-dependent manner, whereas treatment with NAC did not block ROS
generation (data not shown) and apoptosis in SH003-treated HeLa
cells. The present data indicated that SH003-mediated apoptosis was
not correlated with increased ROS production.
In conclusion, the present study is the first to
suggest, to the best of our knowledge, that SH003 induces extrinsic
apoptosis in HeLa cells. Although a mechanism of SH003 action on
treating cervical cancer cells is required to clarify which of the
active compounds of SH003 show anticancer effects against cervical
cancer cells, the present study suggests that SH003 may be
effective for the treatment of cervical cancer.
Acknowledgements
This study was supported by The Korean Medicine
R&D Project of the Ministry of Health and Welfare (grant no.
B110043).
References
|
1
|
Popat K, McQueen K and Feeley TW: The
global burden of cancer. Best Pract Res Clin Anaesthesiol.
27:399–408. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Vaccarella S, Bruni L and Seoud M: Burden
of human papillomavirus infections and related diseases in the
extended Middle East and North Africa region. Vaccine. 31 Suppl
6:G32–G44. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pecorelli S, Zigliani L and Odicino F:
Revised FIGO staging for carcinoma of the cervix. Int J Gynaecol
Obstet. 105:107–108. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dueñas-González A, Cetina L, Coronel J and
González-Fierro A: The safety of drug treatments for cervical
cancer. Expert Opin Drug Saf. 15:169–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Choi EJ and Kim GH: Antioxidant and
anticancer activity of Artemisia princeps var. Orientalis extract
in HepG2 and Hep3B hepatocellular carcinoma cells. Chin J Cancer
Res. 25:536–543. 2013.PubMed/NCBI
|
|
7
|
Fang L, Wang Z, Kong WY, Feng JG, Ma SL
and Lin NM: Anti-tumor and apoptotic effects in vitro and in vivo
of a traditional Chinese medicine prescription. Chin Med J (Engl).
124:3583–3587. 2011.PubMed/NCBI
|
|
8
|
Cheng YL, Chang WL, Lee SC, Liu YG, Chen
CJ, Lin SZ, Tsai NM, Yu DS, Yen CY and Harn HJ: Acetone extract of
Angelica sinensis inhibits proliferation of human cancer cells via
inducing cell cycle arrest and apoptosis. Life Sci. 75:1579–1594.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sato T, Kita K, Sato C and Kaneda A:
Hochu-ekki-to (Bu-zhong-yi-qi-tang), a herbal medicine, enhances
cisplatin-induced apoptosis in HeLa cells. Mol Med Rep.
12:6215–6220. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Duan D, Zhang J, Yao J, Liu Y and Fang J:
Targeting thioredoxin reductase by parthenolide contributes to
inducing apoptosis of HeLa cells. J Biol Chem. 291:10021–10031.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Qian S and Li M: Chamaejasmine induces
apoptosis in HeLa cells through the PI3K/Akt signaling pathway.
Anticancer Drugs. 28:40–50. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Choi YK, Cho SG, Woo SM, Yun YJ, Park S,
Shin YC and Ko SG: Herbal extract SH003 suppresses tumor growth and
metastasis of MDA-MB-231 breast cancer cells by inhibiting
STAT3-IL-6 signaling. Mediators Inflamm. 2014:4921732014.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Choi HS, Kim MK, Lee K, Lee KM, Choi YK,
Shin YC, Cho SG and Ko SG: SH003 represses tumor angiogenesis by
blocking VEGF binding to VEGFR2. Oncotarget. 7:32969–32979. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Li L, Jiang C, Xing C, Kim SH and
Lü J: Anti-cancer and other bioactivities of Korean Angelica gigas
Nakai (AGN) and its major pyranocoumarin compounds. Anticancer
Agents Med Chem. 12:1239–1254. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yang B, Xiao B and Sun T: Antitumor and
immunomodulatory activity of Astragalus membranaceus
polysaccharides in H22 tumor-bearing mice. Int J Biol Macromol.
62:287–290. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhang WL, Zheng KY, Zhu KY, Zhan JY, Bi
CW, Chen JP, Du CY, Zhao KJ, Lau DT, Dong TT and Tsim KW: Chemical
and biological assessment of Angelica herbal decoction: Comparison
of different preparations during historical applications.
Phytomedicine. 19:1042–1048. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kim BS, Seo H, Kim HJ, Bae SM, Son HN, Lee
YJ, Ryu S, Park RW and Nam JO: Decursin from Angelica gigas Nakai
inhibits B16F10 melanoma growth through induction of apoptosis. J
Med Food. 18:1121–1127. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui R, He J, Wang B, Zhang F, Chen G, Yin
S and Shen H: Suppressive effect of Astragalus membranaceus Bunge
on chemical hepatocarcinogenesis in rats. Cancer Chemother
Pharmacol. 51:75–80. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lv J, Zhao Z, Chen Y, Wang Q, Tao Y, Yang
L, Fan TP and Liu C: The chinese herbal decoction danggui buxue
tang inhibits angiogenesis in a rat model of liver fibrosis. Evid
Based Complement Alternat Med. 2012:2849632012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shin JW, Son JY, Kang JK, Han SH, Cho CK
and Son CG: Trichosanthes kirilowii tuber extract induces G2/M
phase arrest via inhibition of tubulin polymerization in HepG2
cells. J Ethnopharmacol. 115:209–216. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kongtun S, Jiratchariyakul W, Kummalue T,
Tan-ariya P, Kunnachak S and Frahm AW: Cytotoxic properties of root
extract and fruit juice of Trichosanthes cucumerina. Planta Med.
75:839–842. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Heo BG, Chon SU, Park YJ, Bae JH, Park SM,
Park YS, Jang HG and Gorinstein S: Antiproliferative activity of
Korean wild vegetables on different human tumor cell lines. Plant
Foods Hum Nutr. 64:257–263. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li LK, Kuang WJ, Huang YF, Xie HH, Chen G,
Zhou QC, Wang BR and Wan LH: Anti-tumor effects of Astragalus on
hepatocellular carcinoma in vivo. Indian J Pharmacol. 44:78–81.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cho WC and Leung KN: In vitro and in vivo
anti-tumor effects of Astragalus membranaceus. Cancer Lett.
252:43–54. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng XD, Hou CH, Zhang XJ, Xie HY, Zhou
WY, Yang L, Zhang SB and Qian RL: Effects of Huangqi (Hex) on
inducing cell differentiation and cell death in K562 and HEL cells.
Acta Biochim Biophys Sin (Shanghai). 36:211–217. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yin G, Tang D, Dai J, Liu M, Wu M, Sun YU,
Yang Z, Hoffman RM, Li L, Zhang S and Guo X: Combination efficacy
of astragalus membranaceus and curcuma wenyujin at different stages
of tumor progression in an imageable orthotopic nude mouse model of
metastatic human ovarian cancer expressing red fluorescent protein.
Anticancer Res. 35:3193–3207. 2015.PubMed/NCBI
|
|
27
|
Lee HJ, Lee HJ, Lee EO, Lee JH, Lee KS,
Kim KH, Kim SH and Lü J: In vivo anti-cancer activity of Korean
Angelica gigas and its major pyranocoumarin decursin. Am J Chin
Med. 37:127–142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fang EF, Zhang CZ, Zhang L, Wong JH, Chan
YS, Pan WL, Dan XL, Yin CM, Cho CH and Ng TB: Trichosanthin
inhibits breast cancer cell proliferation in both cell lines and
nude mice by promotion of apoptosis. PLoS One. 7:e415922012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li M, Li X and Li JC: Possible mechanisms
of trichosanthin-induced apoptosis of tumor cells. Anat Rec
(Hoboken). 293:986–992. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Woo SM, Choi YK, Cho SG, Park S and Ko SG:
A New Herbal Formula, KSG-002, suppresses breast cancer growth and
metastasis by Targeting NF-κB-dependent TNF α production in
macrophages. Evid Based Complement Alternat Med. 2013:7282582013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Degterev A and Yuan J: Expansion and
evolution of cell death programmes. Nat Rev Mol Cell Biol.
9:378–390. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kerr JF, Wyllie AH and Currie AR:
Apoptosis: A basic biological phenomenon with wide-ranging
implications in tissue kinetics. Br J Cancer. 26:239–257. 1972.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Youle RJ and Strasser A: The BCL-2 protein
family: Opposing activities that mediate cell death. Nat Rev Mol
Cell Biol. 9:47–59. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ola MS, Nawaz M and Ahsan H: Role of Bcl-2
family proteins and caspases in the regulation of apoptosis. Mol
Cell Biochem. 351:41–58. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
McIlwain DR, Berger T and Mak TW: Caspase
functions in cell death and disease. Cold Spring Harb Perspect
Biol. 5:a0086562013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lavrik IN and Krammer PH: Regulation of
CD95/Fas signaling at the DISC. Cell Death Differ. 19:36–41. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vermeulen K, Van Bockstaele DR and
Berneman ZN: The cell cycle: A review of regulation, deregulation
and therapeutic targets in cancer. Cell Prolif. 36:131–149. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Song J, Li J, Qiao J, Jain S, Evers B Mark
and Chung DH: PKD prevents H2O2-induced apoptosis via NF-kappaB and
p38 MAPK in RIE-1 cells. Biochem Biophys Res Commun. 378:610–614.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Song Y, Li X, Li Y, Li N, Shi X, Ding H,
Zhang Y, Li X, Liu G and Wang Z: Non-esterified fatty acids
activate the ROS-p38-p53/Nrf2 signaling pathway to induce bovine
hepatocyte apoptosis in vitro. Apoptosis. 19:984–997. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Circu ML and Aw TY: Reactive oxygen
species, cellular redox systems, and apoptosis. Free Radic Biol
Med. 48:749–762. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Trachootham D, Alexandre J and Huang P:
Targeting cancer cells by ROS-mediated mechanisms: A radical
therapeutic approach? Nat Rev Drug Discov. 8:579–591. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chaithongyot S, Asgar A, Senawong G,
Yowapuy A, Lattmann E, Sattayasai N and Senawong T: Anticancer
effects of curcuma C20-dialdehyde against colon and cervical cancer
cell lines. Asian Pac J Cancer Prev. 16:6513–6519. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Luo H, Wang F, Bai Y, Chen T and Zheng W:
Selenium nanoparticles inhibit the growth of HeLa and MDA-MB-231
cells through induction of S phase arrest. Colloids Surf B
Biointerfaces. 94:304–308. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Song JM, Anandharaj A, Upadhyaya P,
Kirtane AR, Kim JH, Hong KH, Panyam J and Kassie F: Honokiol
suppresses lung tumorigenesis by targeting EGFR and its downstream
effectors. Oncotarget. 7:57752–57769. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Wang S, Wu X, Tan M, Gong J, Tan W, Bian
B, Chen M and Wang Y: Fighting fire with fire: Poisonous Chinese
herbal medicine for cancer therapy. J Ethnopharmacol. 140:33–45.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cheng HM, Li CC, Chen CY, Lo HY, Cheng WY,
Lee CH, Yang SZ, Wu SL, Hsiang CY and Ho TY: Application of
bioactivity database of Chinese herbal medicine on the therapeutic
prediction, drug development, and safety evaluation. J
Ethnopharmacol. 132:429–437. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Choi YJ, Choi YK, Lee KM, Cho SG, Kang SY
and Ko SG: SH003 induces apoptosis of DU145 prostate cancer cells
by inhibiting ERK-involved pathway. BMC Complement Altern Med.
16:5072016. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Choi EK, Kim SM, Hong SW, Moon JH, Shin
JS, Kim JH, Hwang IY, Jung SA, Lee DH, Lee EY, et al: SH003
selectively induces p73-dependent apoptosis in triple-negative
breast cancer cells. Mol Med Rep. 14:3955–3960. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Woo SM, Kim AJ, Choi YK, Shin YC, Cho SG
and Ko SG: Synergistic effect of SH003 and doxorubicin in
triple-negative breast cancer. Phytother Res. 30:1817–1823. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hartwell LH and Weinert TA: Checkpoints:
Controls that ensure the order of cell cycle events. Science.
246:629–634. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Liou GY and Storz P: Reactive oxygen
species in cancer. Free Radic Res. 44:479–496. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Storz P: Reactive oxygen species in tumor
progression. Front Biosci. 10:1881–1896. 2005. View Article : Google Scholar : PubMed/NCBI
|














