Introduction
Parkinson's disease (PD) is an age-associated
neurodegenerative disorder characterized by the death of
dopaminergic (DAergic) neurons in the substantia nigra pars
compacta (1,2). It is the second most common
neurodegenerative disease after Alzheimer's disease, always
occurring in people over 60 years old (3). The principal manifestations of PD
include a resting tremor, rigidity, hypokinesia and postural
instability (4,5). It is reported that PD can be caused
by environmental risk factors and genetic factors (6–8).
Despite major advances in the understanding of PD pathophysiology,
no treatments are currently available to prevent disease
progression (9). Additionally, the
exact molecular mechanisms that initiate PD are still unclear
(10). Therefore, elucidating the
molecular mechanisms involved in the pathogenesis of PD is
imperative for the development of novel effective therapeutic
approaches for the treatment of patients with PD.
MicroRNAs (miRNAs) are small, endogenous, noncoding
RNA molecules, which have exhibited potential as biomarkers or
therapeutic targets for PD (11–13).
Bioinformatics analysis has revealed that ~200 miRNAs, including
miR-627, miR-634, miR-514, miR-563 and miR-613, are differentially
expressed between PD and healthy tissue samples (14), suggesting that miRNAs may
participate in PD pathogenesis and progression. In addition,
upregulation of miR-124 has been demonstrated to regulate autophagy
and apoptotic processes, and thus reduce the loss of DAergic
neurons in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
(MPTP)-induced model of PD (15).
Furthermore, miR-7 has been reported to possess neuroprotective
properties, exerted through the inhibition of neuronal apoptosis in
an in vitro model of PD (16). It is also reported that miR-7 can
modulate neuroinflammation in the pathogenesis of PD through
targeting Nod-like receptor protein 3 inflammasome (17). Therefore, it is hypothesized that
miRNAs serve critical roles in PD via the regulation of autophagy
and apoptotic pathways in DAergic neurons. Recently, miR-185
expression was reported to be downregulated in serum samples
isolated from patients with PD compared with in healthy individuals
(18). However, the roles of
miR-185 in PD, as well as the molecular mechanisms underlying its
involvement in the pathogenesis of the disease, have yet to be
elucidated.
The present study investigated the expression of
miR-185 in an MPTP-induced in vitro model of PD using the
human SH-SY5Y DAergic neuroblastoma cell line. In addition, the
effects of miR-185 overexpression on cellular autophagy and
apoptosis were investigated. In order to explore the molecular
mechanisms underlying the effects of miR-185 during PD
pathogenesis, the 5′-adenosine monophosphate-activated protein
kinase (AMPK)/mechanistic target of rapamycin (mTOR) signaling
pathway was examined. The aim of the present study was to
investigate whether the aberrant expression of miR-185 played a key
role in the pathogenesis of PDFindings of this study may provide a
basis for the development of new therapeutic approaches of this
disease.
Materials and methods
Cell culture
The human SH-SY5Y DAergic neuroblastoma cell line
was purchased from American Type Culture Collection (Manassas, VA,
USA). Cells were cultured in Dulbecco's modified Eagle's medium,
supplemented with 10% fetal bovine serum (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) and 100 U/ml penicillin
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), and incubated at
37°C in a humidified 5% CO2 atmosphere.
MPTP has been reported to produce irreversible and
severe Parkinsonism, and has widely been used for the establishment
of PD models (19).
1-Methyl-4-phenylpyridinium is the active metabolite of MPTP, and
is a potent neurotoxin for SH-SY5Y cells and DAergic neurons in the
substantia nigra (20). In the
present study, SH-SY5Y cells were treated with 0, 50, 100, 200 or
400 µM MPTP (Sigma-Aldrich; Merck KGaA) for 24 h to generate an
in vitro PD model. SH-SY5Y cells were then harvested for
further analysis.
Cell transfection
For SH-SY5Y cell transfection, a miR-185 mimic and
scramble miRNA were purchased from Sangon Biotech Co., Ltd.
(Shanghai, China). The miR-185 mimic sequence was
5′-UGGAGAGAAAGGCAGUUCCUGA-3′ and the scramble control sequence was
5′-UUGUACUACACAAAAGUACUG-3′. miRNAs were transfected into SH-SY5Y
cells (1×105 cells/well) using Lipofectamine®
2000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. Cells transfected with scramble
miRNA were used as the negative control. Cells were maintained in
Neurobasal medium (Invitrogen; Thermo Fisher Scientific, Inc.) with
2% B27 (Sigma-Aldrich; Merck KGaA) following transfection with
miR-185 or scramble controls for 24 h. Following 24 h of
transfection, the SH-SY5Y cells were incubated with or without AMPK
inhibitor Compound C (2 µM; Calbiochem; EMD Millipore, Billerica,
MD, USA) in the presence or absence of MPTP.
Cell apoptosis
An Annexin V-fluorescein isothiocyanate (FITC) cell
apoptosis kit (cat. no. V13241; Invitrogen; Thermo Fisher
Scientific, Inc.) was used to assess SH-SY5Y cell apoptosis by flow
cytometry. Briefly, following transfection with miR-185 and
scramble miRNAs for 48 h, cells were harvested by trypsinization
and resuspended in the staining buffer. Cells were fixed with 4%
formaldehyde for 30 min at room temperature and permeabilized with
0.2% Triton X-100 in PBS for 5 min at room temperature and were
incubated for 10 min with 5 µl Annexin-V-FITC and 5 µl propidium
iodide (PI) at room temperature (~22°C). Apoptotic cells,
identified as Annexin V-positive and PI-negative cells, were
subsequently analyzed by flow cytometry using CellQuest software
version 5.2 (BD Biosciences, San Jose, CA, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Following transfection with miR-185 or scramble
controls for 24 h, total RNA was extracted from cells
(2×106 cells) using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. Total RNA was reverse transcribed into
cDNA using the iScript™ cDNA Synthesis kit (Bio-Rad Laboratories,
Inc., Hercules, CA, USA), according to the manufacturer's protocol.
qPCR was performed using SYBR-Green Master mix (Thermo Fisher
Scientific, Inc.). Primers used for qPCR amplification are
presented in Table I, and GAPDH
was used as the internal control. Following heating to 94°C for 2
min, the experimental reaction with a volume of 50 µl was subjected
to 32 cycles of 94°C for 30 sec, 61°C for 30 sec, and 72°C for 30
sec, and the PCR products were analyzed by 1.5% gel
electrophoresis. Relative gene expression was calculated using the
2−ΔΔCq method (21).
 | Table I.Primer sequences used for reverse
transcription-quantitative polymerase chain reaction. |
Table I.
Primer sequences used for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Forward primer
(5′-3′) | Reverse primer
(5′-3′) |
|---|
| miR-185 |
GGTGGAGAGAAAGGCAGT | TGCGTGTCGTGGAGTC |
| Beclin 1 |
CAGAGCGATGGTACCTCTCTGGAGGCCTCG |
CTGACCAGGGCTGGCAACTCTAGATGGC |
| LC3I |
TCCGACCGGCCTTTCAAGCAG |
GAGAACCTGACCAGAACTCCCAG |
| LC3II |
AAACGCATTTGCCATCACAGT |
GTGAGGACTTTGGGTGTGGTTC |
| AMPK |
GCTGTGGATCGCCAAATTAT |
CACGTGCTCATCATCGAAAG |
| mTOR |
ATTTGATCAGGTGTGCCAGT |
GCTTAGGACATGGTTCATGG |
| GAPDH |
GGGAGCCAAAAGGGTCAT |
GAGTCCTTCCACGATACCAA |
Western blot analysis
Cells were washed twice with ice-cold PBS, collected
and lysed using 2 ml lysis buffer. Following transfection with
miR-185 or scramble controls for 24 h, total protein was extracted
from cells (2×106 cells) using radioimmunoprecipitation
assay lysis buffer (50 mM Tris, 150 mM NaCl, 2 mM EDTA, 1% Triton
X-100, 0.1% sodium dodecyl sulfate; pH 7.8; Sangon Biotech Co.,
Ltd.). The lysates were centrifuged at 10,000 × g for 10 min at
4°C. Protein concentration was detected using the bicinchoninic
acid Protein Assay kit (Pierce; Thermo Fisher Scientific, Inc.).
Equal quantities of extracted protein samples (~50 µg) were
separated by 10% SDS-PAGE and transferred onto polyvinylidene
difluoride membranes. Membranes were blocked with
phosphate-buffered saline containing 0.2% Tween-20 and 5% non-fat
milk at 4°C overnight, and then incubated with the following
primary antibodies overnight at 4°C: Anti-Beclin 1 (cat. no.
sc-48341; 1:1,000; Santa Cruz Biotechnology, Inc. Dallas, TX, USA),
anti-microtubule-associated protein light chain 3 (LC3)I (cat. no.
sc-16756; 1:1,000; Santa Cruz Biotechnology, Inc.), anti-LC3II,
anti-AMPK (cat. no. ab80039; 1:1,000; Abcam, Cambridge, UK),
anti-p-AMPK (cat. no. ab133448; 1:1,000; Abcam), anti-mTOR (cat.
no. sc-293089; 1:1,000; Santa Cruz Biotechnology, Inc.),
anti-p-mTOR (cat. no. sc-293133; 1:1,000; Santa Cruz Biotechnology,
Inc.), and anti-GAPDH (cat. no. sc-293335; 1:5,000; Santa Cruz
Biotechnology, Inc.). Membranes were then probed with horseradish
peroxidase-conjugated secondary antibodies (cat. nos. ab131368 and
ab191866; both 1:5,000; Abcam) for 2 h at room temperature. Protein
bands were visualized by enhanced chemiluminescence (ECL) using an
ECL Western Blotting Detection system (GE Healthcare, Chicago, IL,
USA). GAPDH was used as the internal control.
Statistical analysis
Differences among groups were assessed using one-way
analysis of variance followed by the least significant difference
test. Data are expressed as the mean ± standard deviation of at
least three independent experiments. Statistical analysis was
performed using GraphPad Prism software (version 5.0; GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
miR-185 is downregulated in
MPTP-treated SH-SY5Y cells
In the present study, RT-qPCR was used to assess the
expression of miR-185 in MPTP-treated SH-SY5Y cells. As shown in
Fig. 1, the expression levels of
miR-185 in SH-SY5Y cells exhibited a dose-dependent decrease
following treatment with increasing MPTP concentrations. Notably,
miR-185 expression was significantly downregulated following
treatment with MPTP at concentrations of >100 µM when compared
with untreated controls (100 µM, P<0.05; 200 µM, P<0.01; 400
µM, P<0.001). These results suggest that miR-185 may be
downregulated in DAergic neurons during the pathogenesis of PD.
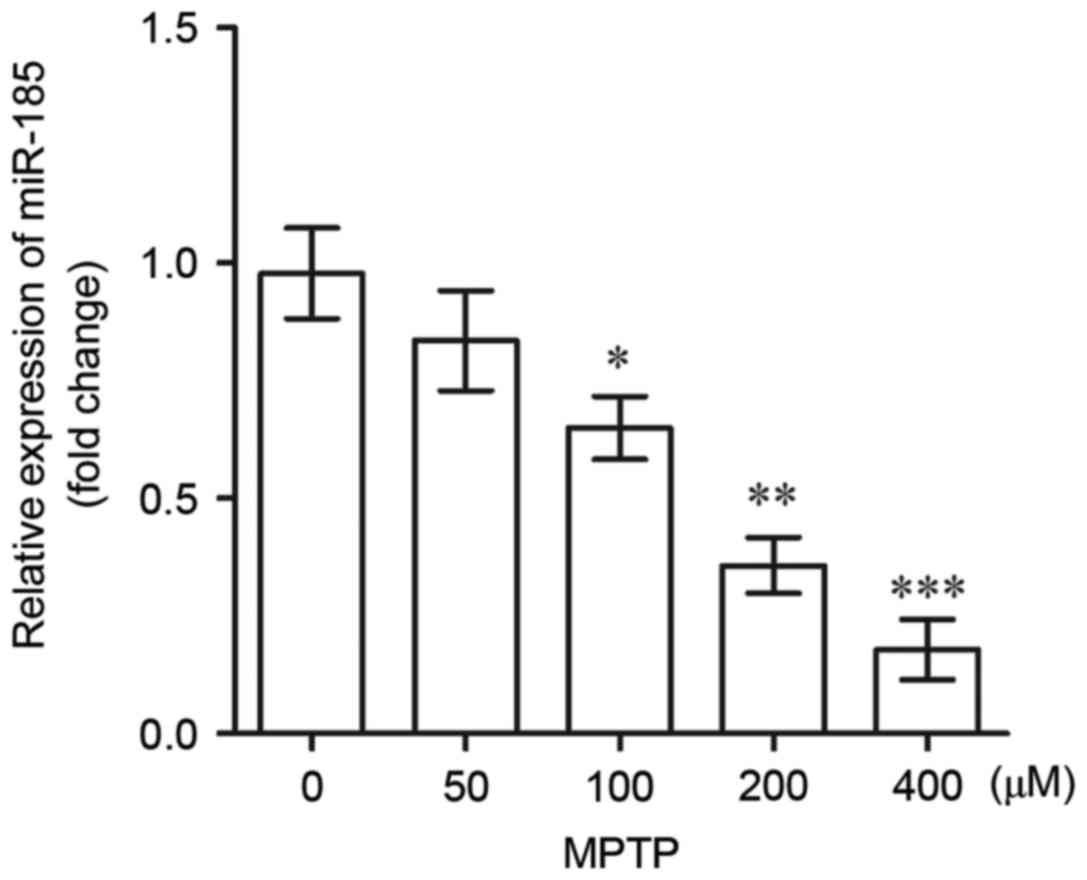 | Figure 1.miR-185 expression levels in human
SH-SY5Y dopaminergic neuroblastoma cells following treatment with
0, 50, 100, 200 and 400 µM MPTP, as determined by reverse
transcription-quantitative polymerase chain reaction. MPTP at
concentrations >100 µM significantly downregulated the
expression of miR-185. Data are expressed as the mean ± standard
deviation. *P<0.05, **P<0.01, ***P<0.001 vs. 0 µM MPTP.
miR, microRNA; MPTP,
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. |
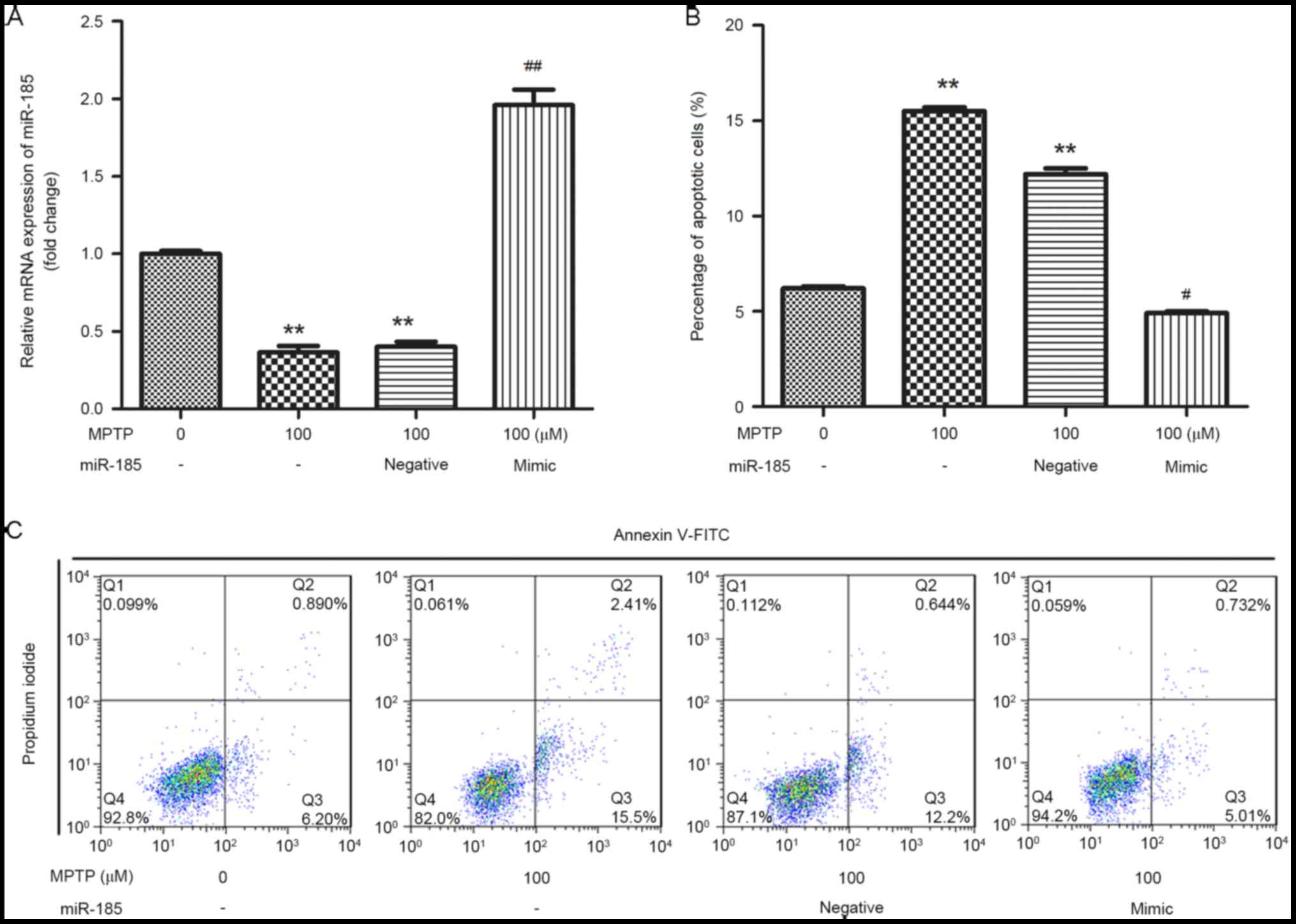
miR-185 overexpression reduces
MPTP-induced apoptosis of SH-SY5Y cells
In order to investigate the effects of miR-185 in
MPTP-induced PD, SH-SY5Y cells were transfected with a miR-185
mimic or a negative control. As shown in Fig. 2A, the expression of miR-185 in
SH-SY5Y cells was significantly increased following transfection
with the miR-185 mimic when compared with untransfected control
cells (P<0.01), indicating that miR-185 was successfully
overexpressed in MPTP-treated SH-SY5Y cells. In addition, flow
cytometric analysis revealed that treatment with MPTP was
associated with a significant increase in the percentage of
apoptotic SH-SY5Y cells, whereas miR-185 overexpression
significantly suppressed MPTP-induced cell apoptosis (P<0.05;
Fig. 2B and C).
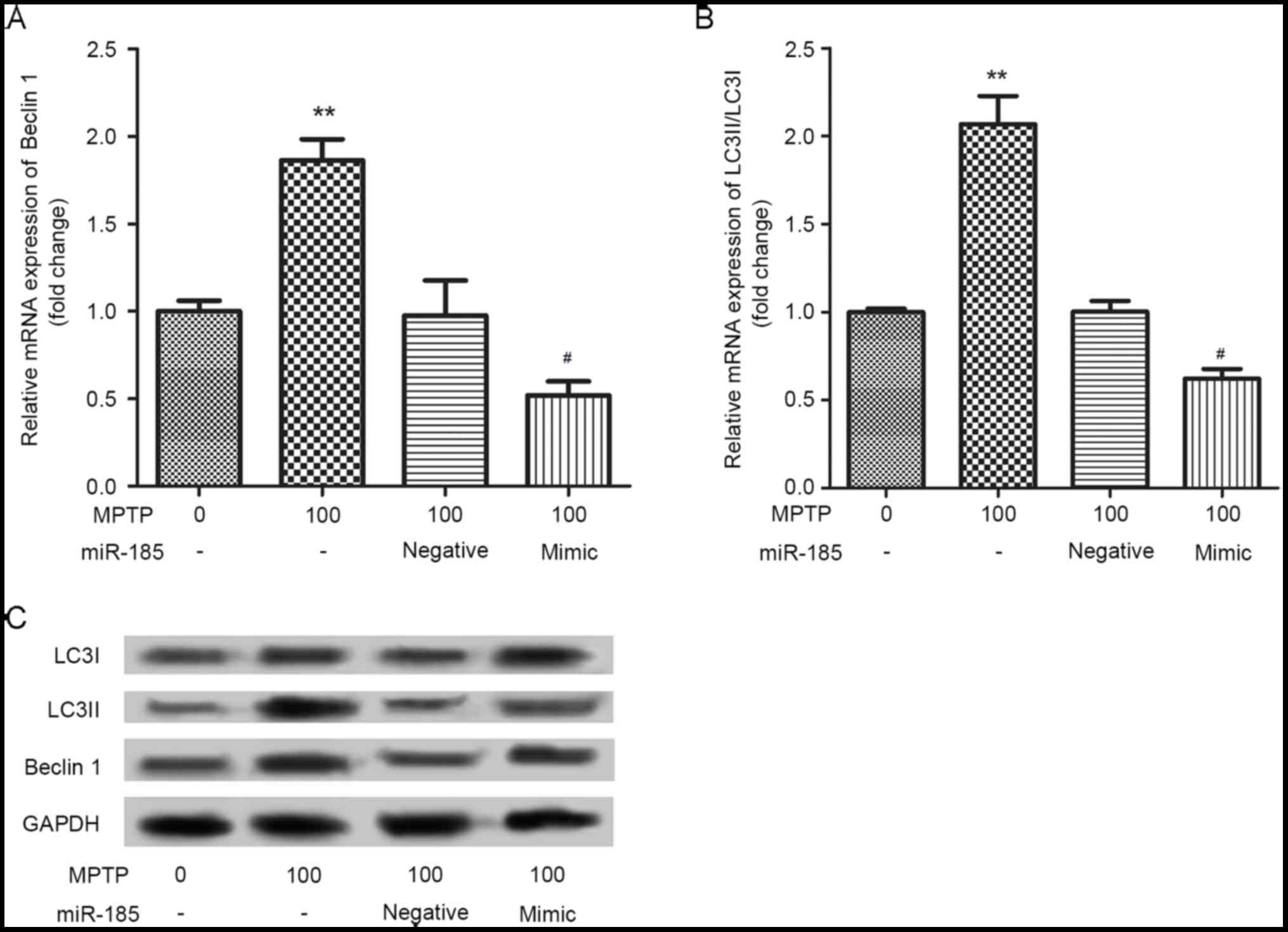
miR-185 overexpression decreases
MPTP-induced autophagy of SH-SY5Y cells
The effects of miR-185 overexpression on
MPTP-induced SH-SY5Y cell autophagy were investigated by assessing
the protein expression of the autophagy markers, Beclin 1, LC3I and
LC3II. As shown in Fig. 3, the
mRNA expression levels of Beclin 1 and the LC3II/LC3I ratio were
significantly upregulated in SH-SY5Y cells following treatment with
MPTP. Notably, miR-185 overexpression significantly suppressed the
MPTP-induced upregulation in the expression levels of these
autophagy markers (P<0.05). Similar results were identified in
the western blot analysis. These results suggested that miR-185
overexpression may decrease MPTP-induced cell autophagy.
miR-185 overexpression suppresses
activation of the AMPK/mTOR signaling pathway
To further investigate the molecular mechanisms
underlying the involvement of miR-185 in PD pathogenesis, the
AMPK/mTOR signaling pathway was examined. As shown in Fig. 4A, miR-185 overexpression was
revealed to significantly inhibit the MPTP-enhanced upregulation of
AMPK and mTOR mRNA (P<0.05), which suggests that miR-185
overexpression suppresses activation of the AMPK/mTOR signaling
pathway. As presented in Fig. 4B,
the protein expression levels of AMPK and mTOR were not altered by
MPTP or MPTP+miR-185 treatment. In addition, the MPTP-induced
upregulation of p-AMPK was reduced by miR-185 transfection, whereas
p-mTOR expression was increased following MPTP+miR-185 treatment.
These results suggested that miR-185 did not inhibit activation of
the AMPK/mTOR signaling pathway.
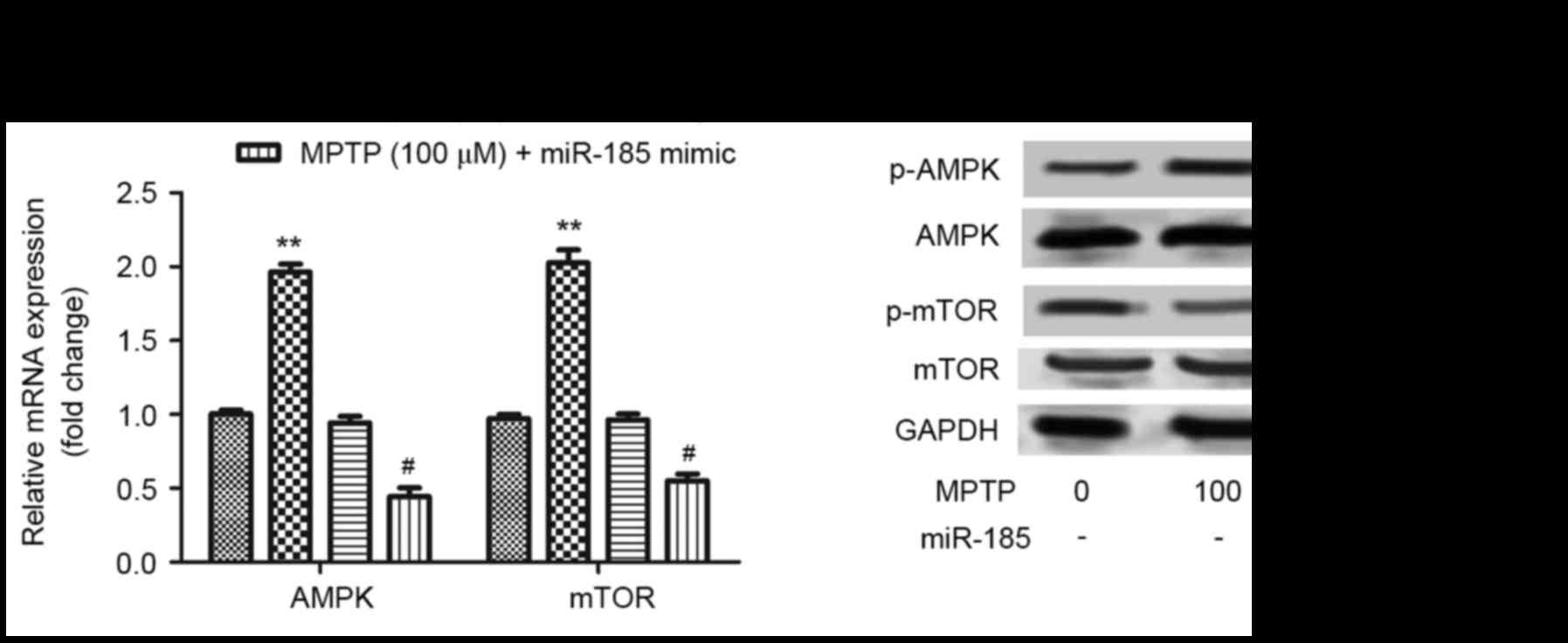 | Figure 4.miR-185 overexpression suppressed
MPTP-induced activation of the AMPK/mTOR signaling pathway in human
SH-SY5Y dopaminergic neuroblastoma cells. (A) The mRNA expression
levels of AMPK and mTOR were assessed in MPTP-treated SH-SY5Y cells
following transfection with an miR-185 mimic or negative control
miRNA using reverse transcription-quantitative polymerase chain
reaction. miR-185 overexpression significantly downregulated the
mRNA expression levels of AMPK and mTOR in MPTP-treated SH-SY5Y
cells. Data are expressed as the mean ± standard deviation.
**P<0.01 vs. untreated control; #P<0.05 vs.
MPTP-treated negative control. (B) The protein expression levels of
AMPK, p-AMPK, mTOR and p-mTOR were assessed by western blot
analysis. Control cells received no treatment with MPTP and were
not transfected. miR, microRNA; MPTP,
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; AMPK, 5′-adenosine
monophosphate-activated protein kinase; mTOR, mechanistic target of
rapamycin; p-, phosphorylated; negative, scramble miRNA. |
To further investigate the effects of miR-185 on the
AMPK/mTOR signaling pathway, cells were treated with the AMPK
inhibitor, Compound C, for 1 h, and the effects of AMPK inhibition
on SH-SY5Y cell apoptosis were assessed by flow cytometry. The
percentage of apoptotic SH-SY5Y cells was significantly increased
following treatment with MPTP and Compound C when compared with
cells treated with Compound C alone (P<0.01; Fig. 5). However, miR-185 overexpression
significantly suppressed SH-SY5Y cell apoptosis following
co-treatment with MPTP and Compound C compared with the
MPTP+Compound C+blank and MPTP+Compound C+negative groups
(P<0.05). These results suggest that miR-185 may be involved in
the regulation of MPTP-induced apoptosis via modulation of the
AMPK/mTOR signaling pathway.
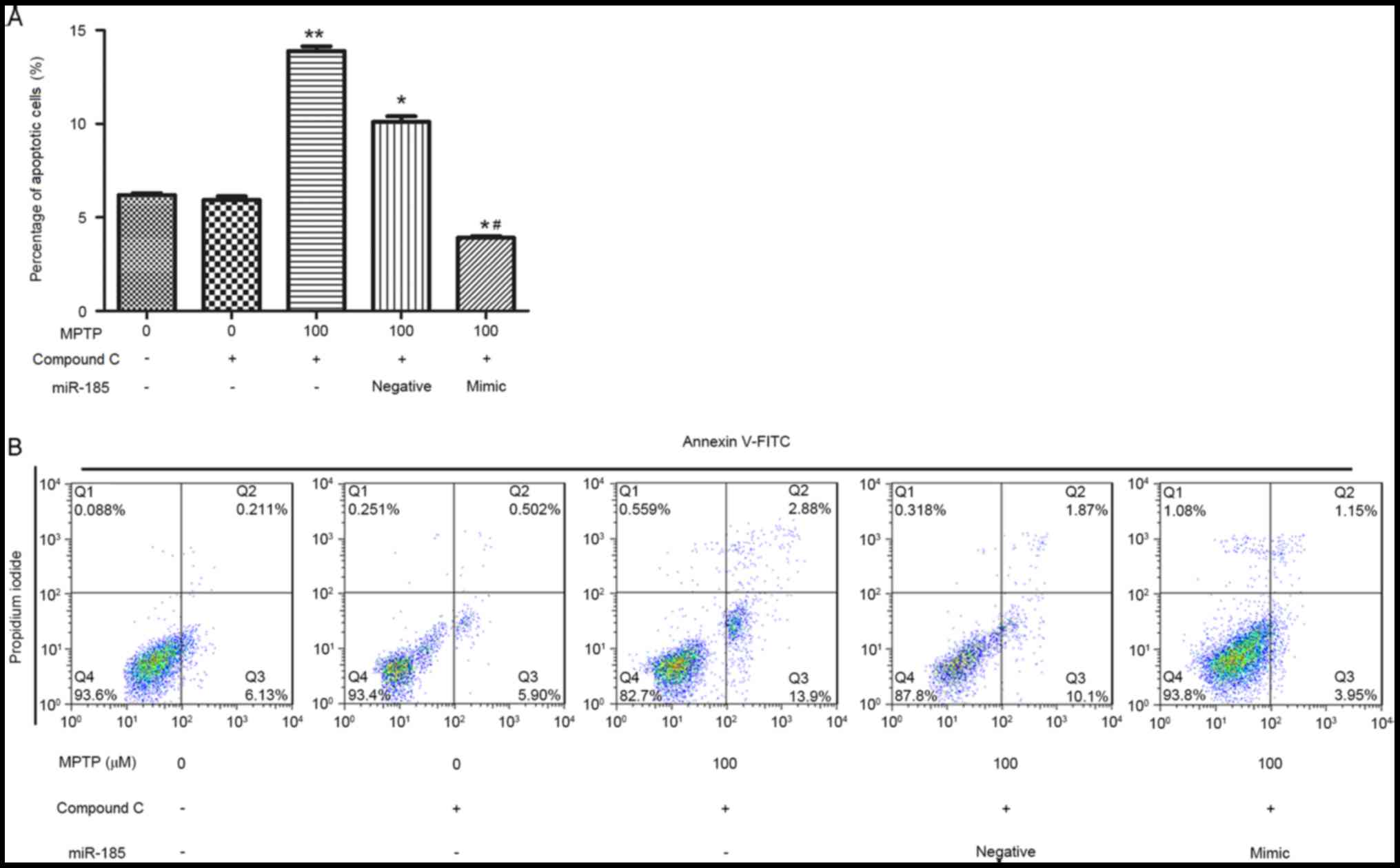 | Figure 5.miR-185 overexpression inhibited
MPTP-induced apoptosis in human SH-SY5Y dopaminergic neuroblastoma
cells following treatment with the AMPK inhibitor Compound C. (A)
The percentage of apoptotic cells was significantly decreased
following miR-185 overexpression in MPTP- and Compound C-treated
SH-SY5Y cells. (B) Flow cytometry was used to assess cellular
apoptosis following treatment with MPTP and Compound C, and
transfection with miR-185 mimic or negative control miRNA. Control
cells received no treatment with MPTP or Compound C, and were not
transfected. Data are expressed as the mean ± standard deviation.
*P<0.05 and **P<0.01 vs. untreated control;
#P<0.05 vs. MPTP-treated negative control. miR,
microRNA; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; AMPK,
5′-adenosine monophosphate-activated protein kinase; FITC,
fluorescein isothiocyanate; negative, scramble miRNA. |
Discussion
In the present study, the role of miR-185 in the
regulation of cellular autophagy and apoptosis were investigated in
an MPTP-induced in vitro model of PD, and the underlying
molecular mechanisms were explored. The results demonstrated that
miR-185 was significantly downregulated in MPTP-treated SH-SY5Y
DAergic cells. Notably, miR-185 overexpression was revealed to
significantly suppress MPTP-induced apoptosis and autophagy in
SH-SY5Y cells. In addition, miR-185 overexpression was demonstrated
to inhibit activation of the AMPK/mTOR signaling pathway in
MPTP-treated cells. These results suggest that miR-185 may be
involved in the pathogenesis and progression of PD.
Apoptosis and autophagy are basic cellular
processes, which are essential for the maintenance of neuronal
homeostasis under physiological conditions. Dysfunction of these
processes has been reported in various neurodegenerative diseases,
including PD (22–24). Apoptosis has been previously
identified as a critical factor that contributes to neuronal
degradation in PD (25). In
addition, dysregulation of autophagy processes has been reported in
animal models of PD, as well as in samples isolated from patients
with PD (26). Beclin 1
upregulation has been revealed to prevent apoptosis and enhance the
activity of autophagy signaling pathways (26). An age-dependent downregulation in
Beclin 1 expression has been demonstrated to occur in the brain,
which has been associated with a reduction in autophagy during the
progression of neurodegenerative diseases (27).
miR-185 has been identified as a crucial regulator
of cell death processes in lung epithelial cells induced by
oxidative stress (28). In
addition, miR-185-3p has been reported to modulate the growth and
apoptosis of nasopharyngeal carcinoma cells (29). In the present study, miR-185
overexpression was revealed to significantly suppress MPTP-induced
apoptosis of SH-SY5Y cells. Furthermore, miR-185 overexpression
significantly downregulated the expression of the autophagy
markers, Beclin 1 and the LC3II/LC3I ratio, in MPTP-treated cells,
thus indicating suppression of cell autophagy. The LC3II/LC3I ratio
is a well-established biochemical assay for the activation of
autophagy (30). These results are
in accordance with previous studies (22,23)
suggesting that miR-185 may be implicated in the progression of PD
via the regulation of cellular apoptosis and autophagy signaling
pathways.
To further investigate the molecular mechanisms
underlying the involvement of miR-185 in the pathogenesis of PD,
the effects of miR-185 on the AMPK/mTOR signaling pathway were
examined. mTOR has been identified as an indirect target of AMPK,
and is negatively regulated by AMPK; a process that is critical for
the regulation of cell growth and survival (31,32).
In addition, mTOR has been demonstrated to induce apoptosis and
autophagy during the development of neurocardiac complications in
diabetes mellitus (33), and
AMPK-induced mTOR inhibition reportedly occurs in autophagy
processes during neuronal cell death induced by the neurotoxin,
tributyltin (34). Hydrogen
peroxide has been revealed to inhibit mTOR signaling and lead to
neuronal apoptosis through the activation of AMPK (35). In addition, Arsikin et al
(36) demonstrated that
AMPK/mTOR-dependent autophagy contributed to the induction of
oxidative stress, thus leading to SH-SY5Y cell apoptosis. In the
present study, miR-185 overexpression downregulated the mRNA
expression of AMPK and mTOR, which had been enhanced following
treatment with MPTP. Furthermore, miR-185 overexpression
significantly suppressed apoptosis of SH-SY5Y cells treated with
MPTP and an AMPK inhibitor simultaneously. These results suggested
that miR-185 overexpression may suppress the death of DAergic
neurons in the substantia nigra, and therefore prevent the
development and progression of PD via inhibition of the AMPK/mTOR
signaling pathway. However, further studies are required to fully
elucidate the association between miR-185 and the AMPK/mTOR
signaling pathway, and explore the molecular mechanisms underlying
their roles in the pathogenesis of PD.
In conclusion, the results of the present study
suggest that miR-185 may inhibit neuronal autophagy and apoptosis
via regulation of the AMPK/mTOR intracellular signaling pathway in
PD. Therefore, the authors hypothesize that AMPK/mTOR-mediated
autophagy and apoptotic signaling pathways may be potential novel
therapeutic targets for the development of alternative strategies
for the treatment of patients with PD.
Acknowledgements
The present study was supported by the National
Nature Science Foundation of China (grant nos. 81171315 and
81227902-5).
References
|
1
|
Moore DJ, West AB, Dawson VL and Dawson
TM: Molecular pathophysiology of Parkinson's disease. Annu Rev
Neurosci. 28:57–87. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Valente EM, Abou-Sleiman PM, Caputo V,
Muqit MM, Harvey K, Gispert S, Ali Z, Del Turco D, Bentivoglio AR,
Healy DG, et al: Hereditary early-onset Parkinson's disease caused
by mutations in PINK1. Science. 304:1158–1160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dexter DT and Jenner P: Parkinson disease:
From pathology to molecular disease mechanisms. Free Radic Biol
Med. 62:132–144. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Olanow CW and Tatton WG: Etiology and
pathogenesis of Parkinson's disease. Annu Rev Neurosci. 22:123–144.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Jankovic J: Parkinson's disease: Clinical
features and diagnosis. J Neurol Neurosurg Psychiatry. 79:368–376.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bonifati V: Genetics of Parkinson's
disease-state of the art, 2013. Parkinsonism Relat Disord. 20 Suppl
1:S23–S28. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Campdelacreu J: Parkinson disease and
Alzheimer disease: Environmental risk factors. Neurologia.
29:541–549. 2014.(In English, Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Collier TJ, Kanaan NM and Kordower JH:
Ageing as a primary risk factor for Parkinson's disease: Evidence
from studies of non-human primates. Nat Rev Neurosci. 12:359–366.
2011. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hwang O: Role of oxidative stress in
Parkinson's disease. Exp Neurobiol. 22:11–17. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Noyce AJ, Bestwick JP, Silveira-Moriyama
L, Hawkes CH, Giovannoni G, Lees AJ and Schrag A: Meta-analysis of
early nonmotor features and risk factors for Parkinson disease. Ann
Neurol. 72:893–901. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mouradian MM: MicroRNAs in Parkinson's
disease. Neurobiol Dis. 46:279–284. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Petillo D, Orey S, Tan AC, Forsgren L and
Khoo SK: Parkinson's disease-related circulating microRNA
biomarkers-a validation study. AIMS Med Sci. 2:7–14. 2015.
View Article : Google Scholar
|
|
13
|
Khoo SK, Petillo D, Kang UJ, Resau JH,
Berryhill B, Linder J, Forsgren L, Neuman LA and Tan AC:
Plasma-based circulating microRNA biomarkers for Parkinson's
disease. J Parkinsons Dis. 2:321–331. 2012.PubMed/NCBI
|
|
14
|
Hao B, Chen X, Dai D, Zou C, Wu X and Chen
J: Bioinformatic analysis of microRNA expression in Parkinson's
disease. Mol Med Rep. 11:1079–1084. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang H, Ye Y, Zhu Z, Mo L, Lin C, Wang Q,
Wang H, Gong X, He X, Lu G, et al: MiR-124 regulates apoptosis and
autophagy process in MPTP model of Parkinson's disease by targeting
to bim. Brain Pathol. 26:167–176. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li S, Lv X, Zhai K, Xu R, Zhang Y, Zhao S,
Qin X, Yin L and Lou J: MicroRNA-7 inhibits neuronal apoptosis in a
cellular Parkinson's disease model by targeting Bax and Sirt2. Am J
Transl Res. 8:993–1004. 2016.PubMed/NCBI
|
|
17
|
Zhou Y, Lu M, Du RH, Qiao C, Jiang CY,
Zhang KZ, Ding JH and Hu G: MicroRNA-7 targets Nod-like receptor
protein 3 inflammasome to modulate neuroinflammation in the
pathogenesis of Parkinson's disease. Mol Neurodegener. 11:282016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ding H, Huang Z, Chen M, Wang C, Chen X,
Chen J and Zhang J: Identification of a panel of five serum miRNAs
as a biomarker for Parkinson's disease. Parkinsonism Relat Disord.
22:68–73. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Langston JW and Irwin I: MPTP: Current
concepts and controversies. Clin Neuropharmacol. 9:485–507. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Itano Y and Nomura Y:
1-Methyl-4-phenyl-pyridinium ion (MPP+) causes DNA fragmentation
and increases the Bcl-2 expression in human neuroblastoma, SH-SY5Y
cells, through different mechanisms. Brain Res. 704:240–245. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ghavami S, Shojaei S, Yeganeh B, Ande SR,
Jangamreddy JR, Mehrpour M, Christoffersson J, Chaabane W, Moghadam
AR, Kashani HH, et al: Autophagy and apoptosis dysfunction in
neurodegenerative disorders. Prog Neurobiol. 112:24–49. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiong N, Xiong J, Jia M, Liu L, Zhang X,
Chen Z, Huang J, Zhang Z, Hou L, Luo Z, et al: The role of
autophagy in Parkinson's disease: Rotenone-based modeling. Behav
Brain Funct. 9:132013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Perier C, Bové J and Vila M: Mitochondria
and programmed cell death in Parkinson's disease: Apoptosis and
beyond. Antioxid Redox Signal. 16:883–895. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tatton WG, Chalmers-Redman R, Brown D and
Tatton N: Apoptosis in Parkinson's disease: Signals for neuronal
degradation. Ann Neurol. 53 Suppl 3:S61–S72. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lynch-Day MA, Mao K, Wang K, Zhao M and
Klionsky DJ: The role of autophagy in Parkinson's disease. Cold
Spring Harb Perspect Med. 2:a0093572012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pickford F, Masliah E, Britschgi M, Lucin
K, Narasimhan R, Jaeger PA, Small S, Spencer B, Rockenstein E,
Levine B and Wyss-Coray T: The autophagy-related protein beclin 1
shows reduced expression in early Alzheimer disease and regulates
amyloid beta accumulation in mice. J Clin Invest. 118:2190–2199.
2008.PubMed/NCBI
|
|
28
|
Zhang D, Lee H, Cao Y, Dela Cruz CS and
Jin Y: MiR-185 mediates lung epithelial cell death after oxidative
stress. Am J Physiol Lung Cell Mol Physiol. 310:L700–L710. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Xu J, Ai Q, Cao H and Liu Q: MiR-185-3p
and miR-324-3p predict radiosensitivity of nasopharyngeal carcinoma
and modulate cancer cell growth and apoptosis by targeting SMAD7.
Med Sci Monit. 21:2828–2836. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mizushima N, Yamamoto A, Matsui M,
Yoshimori T and Ohsumi Y: In vivo analysis of autophagy in response
to nutrient starvation using transgenic mice expressing a
fluorescent autophagosome marker. Mol Biol Cell. 15:1101–1111.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Schmelzle T and Hall MN: TOR, a central
controller of cell growth. Cell. 103:253–262. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hay N and Sonenberg N: Upstream and
downstream of mTOR. Genes Dev. 18:1926–1945. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Maiese K: mTOR: Driving apoptosis and
autophagy for neurocardiac complications of diabetes mellitus.
World J Diabetes. 6:217–224. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Nakatsu Y, Kotake Y, Takai N and Ohta S:
Involvement of autophagy via mammalian target of rapamycin (mTOR)
inhibition in tributyltin-induced neuronal cell death. J Toxicol
Sci. 35:245–251. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Chen L, Xu B, Liu L, Luo Y, Yin J, Zhou H,
Chen W, Shen T, Han X and Huang S: Hydrogen peroxide inhibits mTOR
signaling by activation of AMPKalpha leading to apoptosis of
neuronal cells. Lab Invest. 90:762–773. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Arsikin K, Kravic-Stevovic T, Jovanovic M,
Ristic B, Tovilovic G, Zogovic N, Bumbasirevic V, Trajkovic V and
Harhaji-Trajkovic L: Autophagy-dependent and-independent
involvement of AMP-activated protein kinase in 6-hydroxydopamine
toxicity to SH-SY5Y neuroblastoma cells. Biochim Biophys Acta.
1822:1826–1836. 2012. View Article : Google Scholar : PubMed/NCBI
|