Introduction
Immunoglobulin A nephropathy (IgAN) is a type of
glomerular disorder, which is an important cause of kidney failure
(1). Patients usually present
macroscopic hematuria during a gastrointestinal or upper
respiratory infection (2).
However, the diagnosis of IgAN requires kidney biopsy, which may
lead to various complications (3,4).
Although IgAN is increasingly considered to be a systemic immune
complex deposition disorder, the kidney is most commonly affected,
and damage may frequently recur even subsequent to kidney
transplantation (5). Deposited
immune complexes contain a number of classes of antibody, although
IgA1 is the most frequent and is often the dominant or co-dominant
class. Certain autoreactive IgA1 antibodies that are specific for
the hinge region of IgA1 have additionally been observed in serum
from patients with IgAN (6).
Although the trigger of autoantibodies and formation of immune
complexes has remained unclear until now, certain studies have
indicated that susceptibility alleles in the antigen region of
leukocytes and adaptive immunity may serve key roles in the
pathogenesis of IgAN (7,8).
Human adaptive immunity is mediated by leukocytes,
including B and T cells, which have been demonstrated to be
selectively activated by interactions between antigens and their
receptors expressed on the surface of lymphocytes (9). Therefore, understanding the spectrum
of antigen receptors present on B and T cells, known as the immune
repertoire, may aid in identifying biomarkers and the development
of therapeutics for IgAN (10). In
the antigen-binding site of B-cell receptors (BCRs)/T-cell
receptors (TCRs), specific recognition of countless diverse
peptide-major histocompatibility complexes are determined by three
complementarity-determining regions (CDRs): CDR1, CDR2 and CDR3.
Studies have indicated that CDR1 and CDR2 are relatively conserved
and are encoded by different germline DNA sequences; however, CDR3
is encoded by genomic loci that undergo somatic recombination of
VDJ gene segments and are assembled into highly diverse junctional
residues (11,12). Therefore, the DNA sequences
encoding CDR3 in the BCR/TCR could provide structural information
about primary receptor antigenic binding sites. In the literature,
few studies have focused on the diversity of the TCR at the
nucleotide sequence level in IgAN. In a study by Wu et al
(13), 14 patients with IgAN were
divided into a stable and a progressive group, and TCR CDR3
sequences were cloned and analyzed by the dideoxy chain
determination method. The results revealed that certain conserved
amino acids in the TCR CDR3 contributed to the recognition of a
particular antigen or set of antigens. However, as the human immune
repertoire includes B and T cells, TCR CDR3 is only part of the
information. In another study, TCR diversity in IgAN was assessed
using traditional techniques, included targeted cloning and Sanger
sequencing, which are low-throughput methods and allowed for only a
descriptive assessment of 20 TCR Vβ families (14). In the present study, a
comprehensive analysis of BCR/TCR CDR3 diversity was attempted, in
order to provide novel information about IgAN by multiplex PCR,
high-throughput sequencing and bioinformatics.
Materials and methods
Study subjects
Written informed consent was obtained from each
participant. The study was approved by the Ethics Committee of
Shenzhen People's Hospital and abided by the ethical principles of
the Helsinki Declaration of 1975, as revised in 2000. Peripheral
blood samples with fresh EDTA-K2 anticoagulant were obtained from
eight South Chinese patients with IgAN and six South Chinese
healthy controls in Shenzhen People's Hospital, China. Samples were
collected at the time of the initial diagnosis of IgAN and prior to
drug treatments. The renal biopsies were performed under direct
ultrasound guidance with automated biopsy needles; the sections of
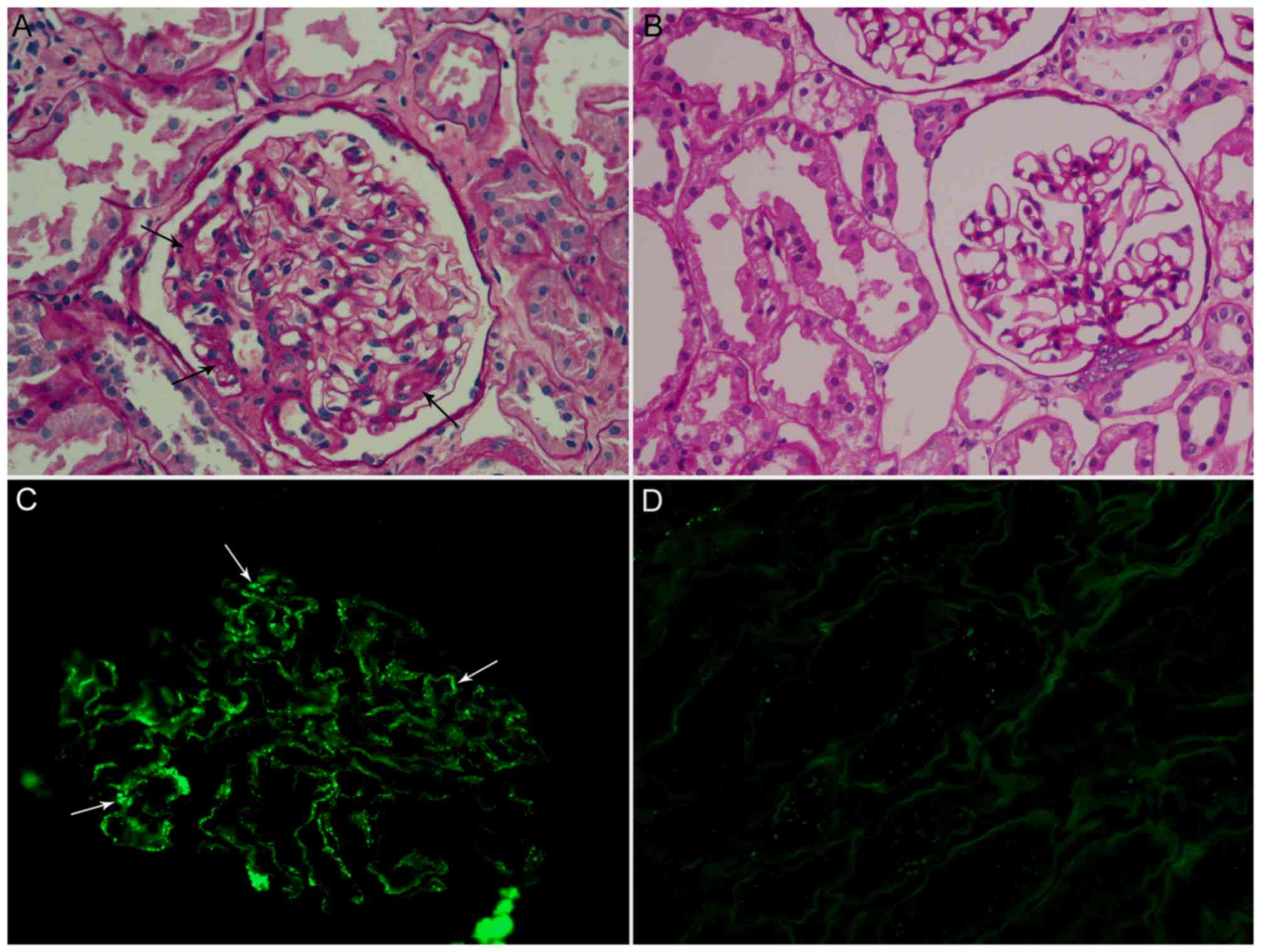
biopsy tissues were stained by periodic acid-Schiff stain at 20°C
for 10 min, and the mesangial IgA deposition was confirmed by
immunofluorescence using fluorescein isothiocyanate-labelled IgA
antibodies (Dako; Agilent Technologies, Inc., Santa Clara, CA, USA)
(15). The diagnosis of IgAN was
confirmed by clinical and biopsy findings (Fig. 1), which were according to the Lee
glomerular grading system (16).
Of the 8 patients with IgAN, 5 were female and 3 were male, and the
mean age was 34.6±6.63 years (range, 26–45 years; Table I). The six healthy controls were
healthy volunteers, 2 men and 4 women, and the mean age was
34.0±6.3 years (range, 27–45 years), and they were matched to the
patients in terms of sex, age and ethnicity.
 | Table I.Clinical data of patients with
immunoglobulin A nephropathy. |
Table I.
Clinical data of patients with
immunoglobulin A nephropathy.
| Case no. | Age, years | Lee's grading | SCR, µmol/l | BUN, mmol/l | Proteinuria, g/24
h | URBC, cells/µl |
|---|
| 1 | 27 | III | 62 | 4.10 | 0.795 | 150 |
| 2 | 44 | III | 122 | 7.48 | 1.148 | 250 |
| 3 | 36 | III | 70 | 3.50 | 0.704 | 150 |
| 4 | 36 | III | 129 | 4.35 | 1.036 | 250 |
| 5 | 26 | IV | 127 | 7.67 | 2.739 | 250 |
| 6 | 34 | IV | 102 | 4.30 | 0.840 | 150 |
| 7 | 31 | III | 69 | 3.99 | 0.730 | 250 |
| 8 | 43 | IV | 116 | 5.19 | 1.872 | 150 |
Cell sub-population isolation and DNA
extraction
Peripheral blood mononuclear cells (PBMCs) were
isolated from each peripheral blood sample using LymphoPrep (Axis
Shield Diagnostics Ltd., Dundee, UK), according to the
manufacturer's protocol. For the isolation of B cells, the non-B
cells of each PBMC sample were depleted using MicroBeads (Miltenyi
Biotec GmbH, Bergisch Gladbach, Germany) using a MACS Separator
(Miltenyi Biotec GmbH). The T cells were isolated from each PBMC
sample using CD3 MicroBeads (Miltenyi Biotec GmbH), according to
the manufacturer's protocol. DNA was extracted from the isolated B
and T cells using a QIAamp DNA Mini kit (Qiagen GmbH, Hilden,
Germany), according to the manufacturer's protocol.
Preparation of BCR/TCR libraries by
multiplex-polymerase chain reaction (PCR)
The human DNA sequences of BCR/TCR CDR3, which are
the most diverse DNA sequences, were obtained from IMGT (www.imgt.org). The relatively conserved regions in the
BCR/TCR DNA sequences were designed as the primer combining sites,
and amplification by multiplex-PCR was performed according to
previous studies, with minor modifications (17,18).
Briefly, equal amounts of DNA taken from each sample were used to
construct a library, with primers covering the majority of the V, D
and J gene families, including 16 primers to amplify BCR CDR3
(Table II), and 44 primers to
amplify TCR CDR3 (Table III).
The multiplex-PCR was performed using a QIAGEN Multiplex PCR kit.
For the amplification of BCR CDR3, the PCR mixture was amplified
under the following conditions: 95°C (15 min) followed by 25 cycles
of 94°C (15 sec), 60°C (3 min) and 72°C (10 min) For the
amplification of TCR CDR3, the PCR mixture was amplified under the
following conditions: 95°C (15 min), followed by 30 cycles of 94°C
(15 sec), 59°C (30 sec) and 72°C (11 min). The PCR products were
purified using a QIA quick PCR Purification kit (Qiagen GmbH) and
AMPure XP beads (Beckman Coulter, Inc., Brea, CA, USA). The
sequencing index was added to each PCR product sample, and the
quantity of each library was analyzed with an Agilent 2100
Bioanalyzer (Agilent Technologies, Inc.) quantitative PCR using the
KAPA Library Quantification kit, following manufacturer's protocol
(Kapa Biosystems, Inc., Wilmington, MA, USA), as previously
described (19).
| Table II.Primers for the amplification of BCR
CDR3. |
Table II.
Primers for the amplification of BCR
CDR3.
| Primer |
Forward/reverse | Sequence
(5′→3′) |
|---|
| IGHV1-18 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACCATGACCACAGAC |
| IGHV1-2/1-46 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACCAKKACCAGGGAC |
| IGHV1-24 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACCATGACCGAGGAC |
| IGHV1-3/1-45 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACCATTACYAGGGAC |
| IGHV1-69/1-f | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACGATWACCRCGGAC |
| IGHV1-8 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGAGTCACCATGACCAGGAAC |
| IGH2-70/26/5 | Forward |
CAGACGTGTGCTCTTCCGATCTAGACCAGGCTCACCATYWCCAAGG |
| IGHV3 | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGCCGATTCACCATCTCMAG |
| IGH4 | Forward |
CAGACGTGTGCTCTTCCGATCTAGCGAGTCACCATRTCMGTAGAC |
| IGHV5-51 | Forward |
CAGACGTGTGCTCTTCCGATCTAGCAGCCGACAAGTCCATCAGC |
| IGHV6-1 | Forward |
CAGACGTGTGCTCTTCCGATCTAGAGTCGAATAACCATCAACCCAG |
| IGHV7-NEW | Forward |
CAGACGTGTGCTCTTCCGATCTAGGACGGTTTGTCTTCTCCTTG |
| HIGHJ-Rev1 | Reverse |
CTACACGACGCTCTTCCGATCTCTGAGGAGACRGTGACCAGGGTG |
| HIGHJ-Rev2 | Reverse |
CTACACGACGCTCTTCCGATCTCTGAAGAGACGGTGACCATTGTC |
| HIGHJ-Rev3 | Reverse |
CTACACGACGCTCTTCCGATCTCTGAGGAGACGGTGACCAGGGT |
| HIGHJ-Rev4 | Reverse |
CTACACGACGCTCTTCCGATCTTGAGGAGACGGTGACCGTGGTC |
| Table III.Primers for the amplification of TCR
CDR3. |
Table III.
Primers for the amplification of TCR
CDR3.
| Primer |
Forward/reverse | Sequence
(5′→3′) |
|---|
| TRBV2F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGATTTCACTCTGAAGATCCGGTCCAC |
| TRBV9F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCCTGACTTGCACTCTGAACTAAACCT |
| TRBV14F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGAGGGACGTATTCTACTCTGAAGG |
| TRBV15F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTTCTTGACATCCGCTCACCAGG |
| TRBV19F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTCCTTTCCTCTCACTGTGACATCGG |
| TRBV3-1-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGAAACAGTTCCAAATCGMTTCTCAC |
|
TRBV4-1/2/3-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCAAGTCGCTTCTCACCTGAATG |
| TRBV5-1-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGCCAGTTCTCTAACTCTCGCTCT |
|
TRBV5-4/5/6/8-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTCAGGTCGCCAGTTCCCTAAYTAT |
|
TRBV6-1/2/3/5/8-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCAATGGCTACAATGTCTCYAGAT |
| TRBV6-4-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTGATGGTTATAGTGTCTCCAGAG |
| TRBV6-9-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCGATGGCTACAATGTATCCAGAT |
| TRBV6-6-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGAATGGCTACAACGTCTCCAGAT |
|
TRBV7-2/4/6/7/8-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGGATCCGTCTCCACTCTGAMGAT |
| TRBV7-3-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGGATCCGTCTCTACTCTGAAGAT |
| TRBV7-9-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGGATCTTTCTCCACCTTGGAGAT |
|
TRBV10-1-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCCTCACTCTGGAGTCTGCTGCC |
|
TRBV10-2/3-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCCTCACTCTGGAGTCMGCTACC |
|
TRBV11-1/2/3-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGCAGAGAGGCTCAAAGGAGTAGACT |
|
TRBV12-3/4-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGATCGATTCTCAGCTAAGATGCCT |
|
TRBV12-5-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGATCGATTCTCAGCAGAGATGCCT |
| TRBV13-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTCGATTCTCAGCTCAACAGTTC |
| TRBV18-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGTAGATGAGTCAGGAATGCCAAAG |
|
TRBV20-1-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGAACCATGCAAGCCTGACCTT |
|
TRBV24-1-F2-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCTCCCTGTCCCTAGAGTCTGCCAT |
| TRBV25-1F-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGCCCTCACATACCTCTCAGTACCTC |
|
TRBV27/28-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGGGAGATGTTCCTGARGGGTACA |
|
TRBV29-1-F4-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGAACTCTGACTGTGAGCAACATGAG |
| TRBV16-F2-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCTGTAGCCTTGAGATCCAGGCTACGA |
| TRBV30-F5-IND | Forward |
CAGACGTGTGCTCTTCCGATCTAGCAGATCAGCTCTGAGGTGCCCCA |
| TRBJ1.1-R2-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCTACAACTGTGAGTCTGGTG |
| TRBJ1.2R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCTACAACGGTTAACCTGGTC |
| TRBJ1.3R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCTACAACAGTGAGCCAACTT |
| TRBJ1.4R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCATACCCAAGACAGAGAGCTGGGTTC |
| TRBJ1.5R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCTAGGATGGAGAGTCGAGTC |
| TRBJ1.6R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCATACCTGTCACAGTGAGCCTG |
| TRBJ2.1R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCCTTCTTACCTAGCACGGTGA |
| TRBJ2.2R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCCAGTACGGTCAGCCT |
| TRBJ2.3R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCCGCTTACCGAGCACTGTCAG |
| TRBJ2.4R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCCAGCTTACCCAGCACTGAGA |
| TRBJ2.5-R2-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCGAGCACCAGGAGCCGCGT |
| TRBJ2.6R-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTCGCCCAGCACGGTCAGCCT |
| TRBJ2.7-R2-P1 | Reverse |
CTACACGACGCTCTTCCGATCTCTTACCTGTGACCGTGAGCCTG |
| P1 | Reverse |
AATGATACGGCGACCACCGAGATCTACACTCTTTCCCTACACGACGCTCTTCCGATCT |
Deep sequencing and data analysis
The libraries were sequenced using Hiseq 2000
following the manufacturer's protocol (Illumina, Inc., San Diego,
CA, USA). In order to control the sequencing quality, internal
controls consisting of defined DNA fragments were used in the
sequence detection. The defined DNA fragments ranged between 170
and 800 bp, and enabled the estimation of error rates (E). The
quality scores of sequencing (sQ), which were one important
criterion for read filtering, ranged from 0 to 40, and the
relationship between sQ and E was calculated by the following
formula:
sQ=-10x(logE/1-E)/log10E=Y/1+YY=sQ/(e-10xlog10)
In order to obtain clean immune sequences, the
adapters and the reads with sQ <15 or N bases >5% in the raw
data generated by Hiseq 2,000, were filtered, as previously
described (17). The clean data
were expected to have sequence lengths of >60 nucleotides; and
the pair-end read pairs, which had identity-aligning tail regions
or sequences with 90% matched bases (≥10 bp overlapping), were
merged into a single-contig sequence using COPE v1.1.3 and FqMerger
(both BGI, Shenzhen, China). Following low-quality read filtering,
the clean data were used in further alignments with the online tool
IMGT/HighV-QUEST (20). The web
portal of IMGT® and the international ImMunoGeneTics
information system® (www.imgt.org)
was used to analyze the nucleotide sequence rearrangement of
BCRs/TCRs obtained from high-throughput sequencing. The output data
included each distinct DNA sequence of BCR/TCR, IMGT clonotypes
(AA), VDJ gene assignment and distribution of CDR3 length.
Statistical analysis was performed using SPSS Statistics 17.0
(SPSS, Inc., Chicago, IL, USA), and the comparisons of frequency
estimates between groups were performed by χ2 testes.
The residue alignment was performed by MEGA7.0 software (www.megasoftware.net) (21).
Results
Summary of BCR/TCR sequencing
For the BCR libraries, 466,667 and 408,506 reads
from the IgAN and the control group, respectively, were obtained.
Following filtering out of unknown sequences, 462,372 and 402,373
clean immune sequences were identified, of which 349,990 and
321,803 were CDR3 sequences. 34,078 unique CDR3 nucleotide
sequences and 24,049 unique CDR3 amino acid clonotypes were
detected in the IgAN group, and 38,191 unique CDR3 nucleotide
sequences and 26,919 unique CDR3 amino acid clonotypes were
detected in the control group. For the TCR libraries, 467,542 and
491,499 reads from the IgAN and the control group, respectively,
were obtained. There were 464,053 and 487,974 clean immune
sequences, of which 344,679 and 362,658 were CDR3 sequences; and
46,735 unique CDR3 nucleotide sequences and 41,573 unique CDR3
amino acid clonotypes were identified in the IgAN group, and 27,272
unique CDR3 nucleotide sequences and 22,918 unique CDR3 amino acid
clonotypes were identified in the control group. The numbers of
unique CDR3 nucleotide sequences and unique CDR3 amino acid
clonotypes indicated that 29.43 and 11.05% nucleotide sequences
would encode identical amino acid clonotypes in the BCR and TCR,
respectively.
Characteristics of the BCR/TCR CDR3
length distribution
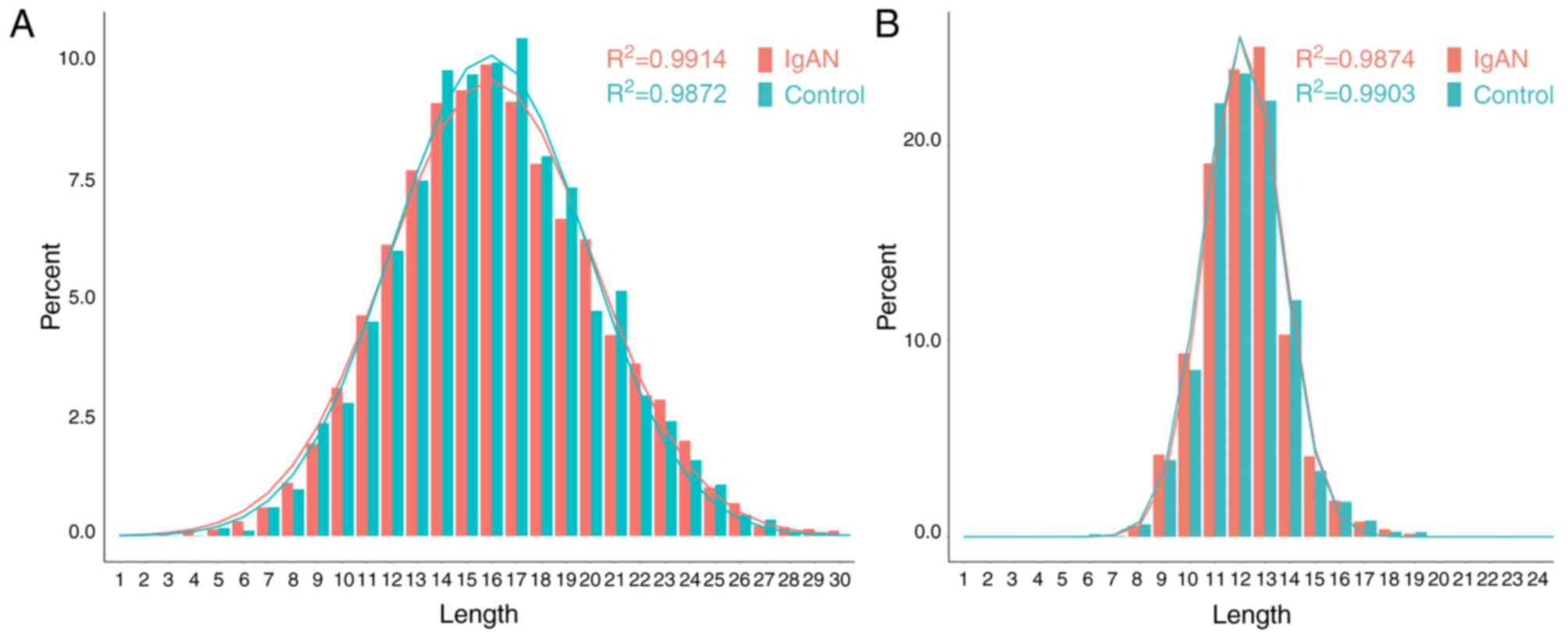
The length distribution of the BCR/TCR CDR3 loop is
an important index for the determination of B/T cell repertoire
diversity. In the present study, the average BCR CDR3 lengths did
not differ significantly between the IgAN (17.01±4.27) and the
control group (17.25±4.15; P>0.05). The average TCR CDR3 lengths
did not differ significantly between the IgAN (12.56±2.56) and the
control group (12.42±2.20; P>0.05). The length distribution of
the BCR/TCR CDR3 loops in the IgAN group and the control group were
observed to fit well to a Gaussian distribution curve
(R2≈1.0; Fig. 2). In
Gaussian distribution curves, the R2 index ranges
between 0 and 1, corresponding to worst and best fit.
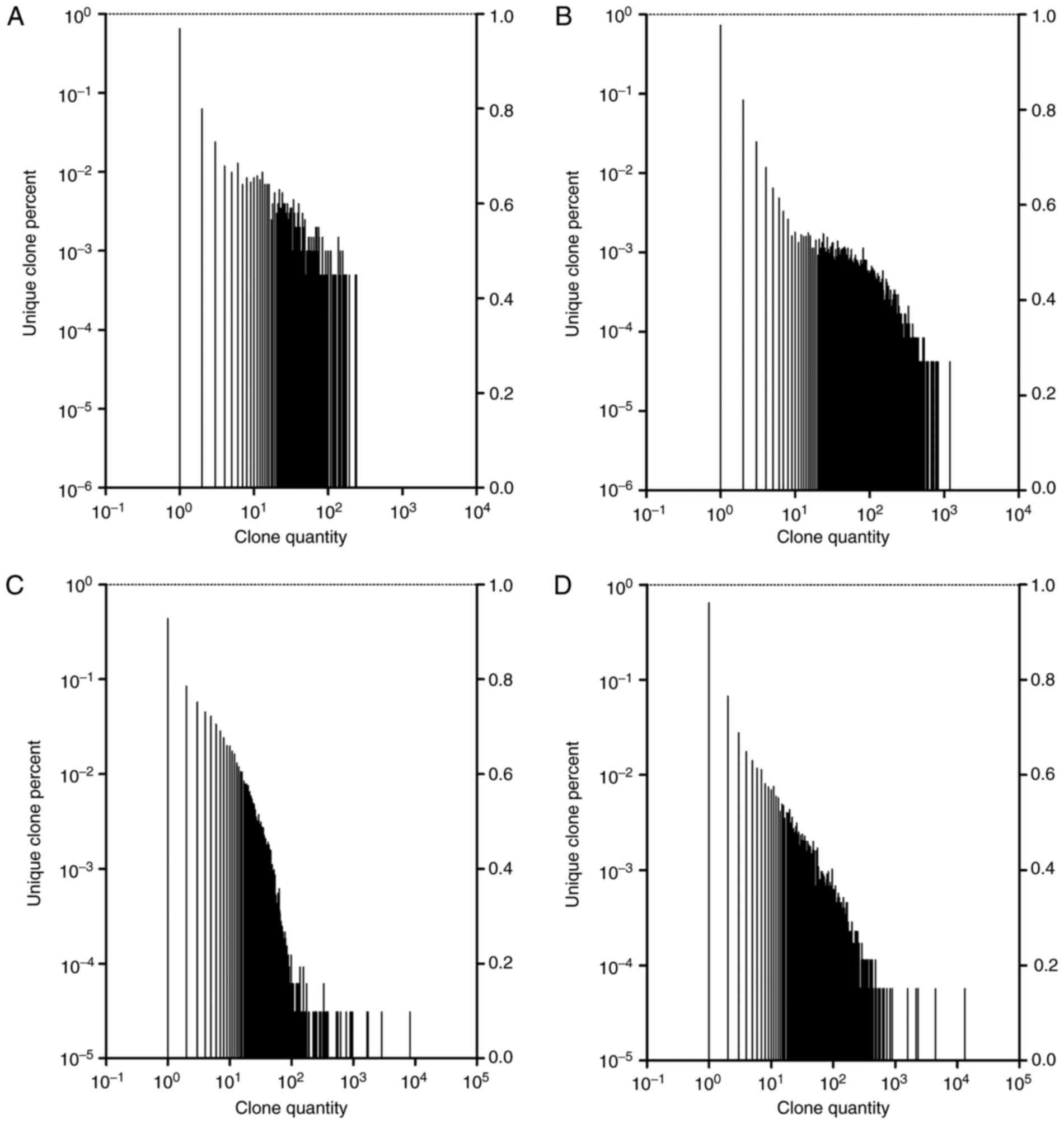
BCR/TCR CDR3 with highly expanded
clones
The expression level of each unique BCR/TCR CDR3
amino acid sequence was evaluated by the unique BCR/TCR CDR3 DNA
sequence frequency following alignment. Highly expanded clones
(HECs) were defined as those with a frequency >0.01% (22). For the BCR CDR3, a total of 3,133
clones were identified as HECs, with a median HEC ratio of
0.0192664% in the IgAN group; whereas, in the control group, 2,338
clones were HECs, with a median HEC ratio of 0.0277151%. A
χ2 test indicated that there were significantly more BCR
CDR3 HECs in the IgAN compared with the control group (P<0.05;
data not shown). The BCR CDR3 HECs in each group were highly
diverse, and all of the HECs in the IgAN group were unique clones.
For the TCR CDR3, a total of 1,804 clones were identified as HECs,
with a median HEC ratio of 0.0133458% in the IgAN group; and in the
control group, 2277 clones were identified as HECs, with a median
HEC ratio of 0.0228866%. The HECs of the TCR CDR3 in each group
were highly diverse, with 98.7804878% of TCR HECs in the IgAN group
and 98.3311375% of TCR HECs in the control group observed to be
unique clones. Furthermore, when the frequency distribution of the
BCR/TCR repertoire was plotted, a great variation between the two
groups of samples was identified, and the frequency distribution
was markedly more skewed in the IgAN group (Fig. 3).
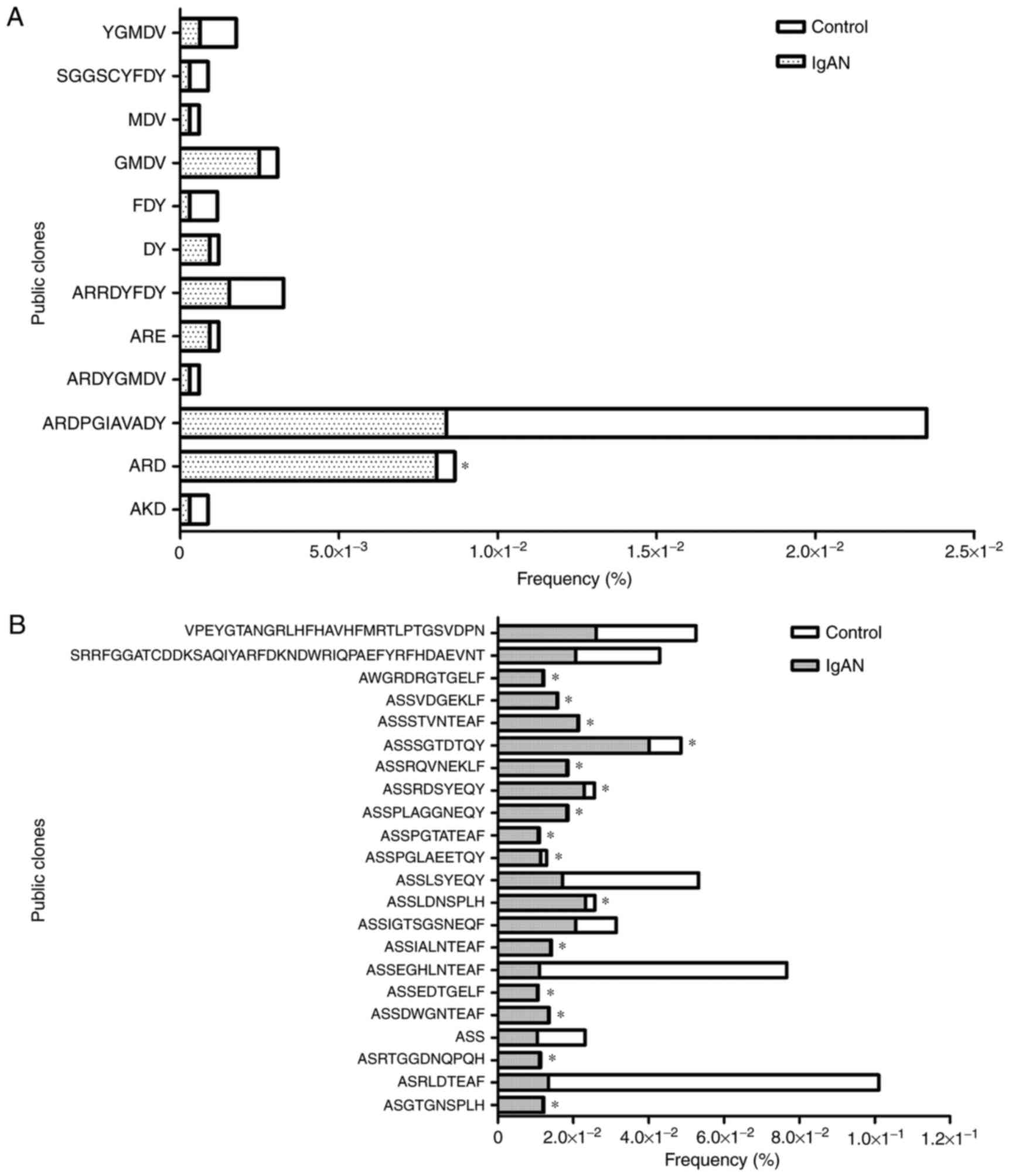
Conserved BCR/TCR CDR3 in the IgAN and
control groups
The public clones in the IgAN and control groups
were considered to be an important aspect of the conserved clones;
a total of 12 BCR CDR3 clones and 228 TCR CDR3 clones were
identified as public ones, of which 22 in the IgAN group were HECs.
The χ2 test indicated that a total of 16 public clones,
including one clone of BCR CDR3 and 15 clones of TCR CDR3, were
expressed with a significantly higher frequency in IgAN patients
compared with the control group (P<0.001; Fig. 4). In order to identify whether
there were any conserved BCR/TCR CDR3 residues among the unique
clones, the top 30 highly expressed BCR/TCR CDR3 clones were
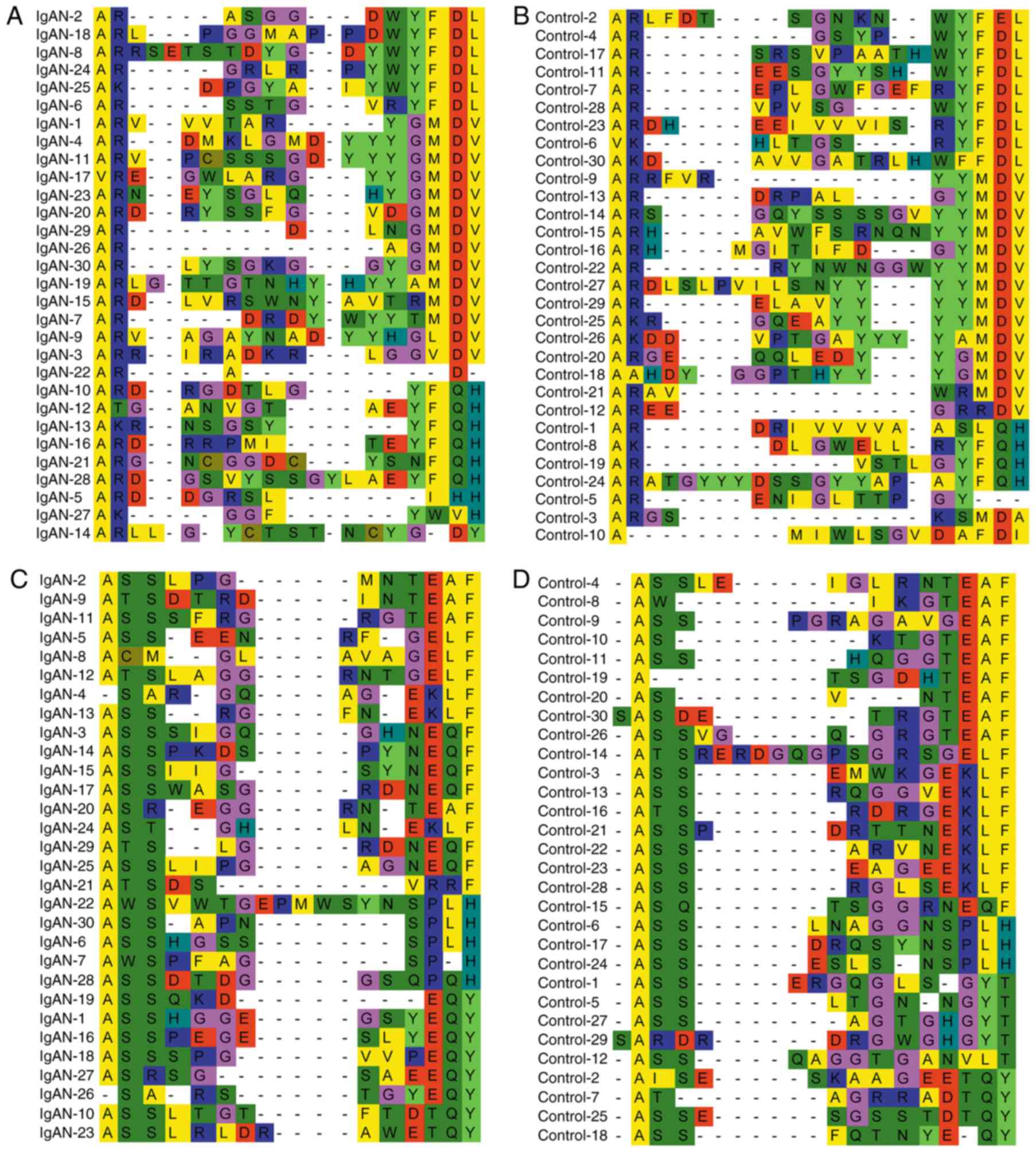
aligned, and it was demonstrated that the amino acid residues of
BCR/TCR CDR3 may not be encoded randomly; there were several
conserved BCR/TCR CDR3 residues, including GMDV, EQY and EQF, that
were recurrent >3 times only in the IgAN group (Fig. 5).
VDJ gene recombination of the BCR/TCR
CDR3
Diversity of the BCR/TCR CDR3 is generated by the
process of VDJ genetic recombination, in which certain genes in the
V, D and J gene family are selected. In order to avoid distortion
by dominant clones, which may be expanded as a consequence of an
immune response, the composition of unique CDR3 sequences was
analyzed and their copies disregarded. The composition of the BCR
was additionally analyzed by the gene sub-segment usage of IGHV,
IGHD, IGHJ; and TCR by the gene sub-segment usage of TRBV, TRBD and
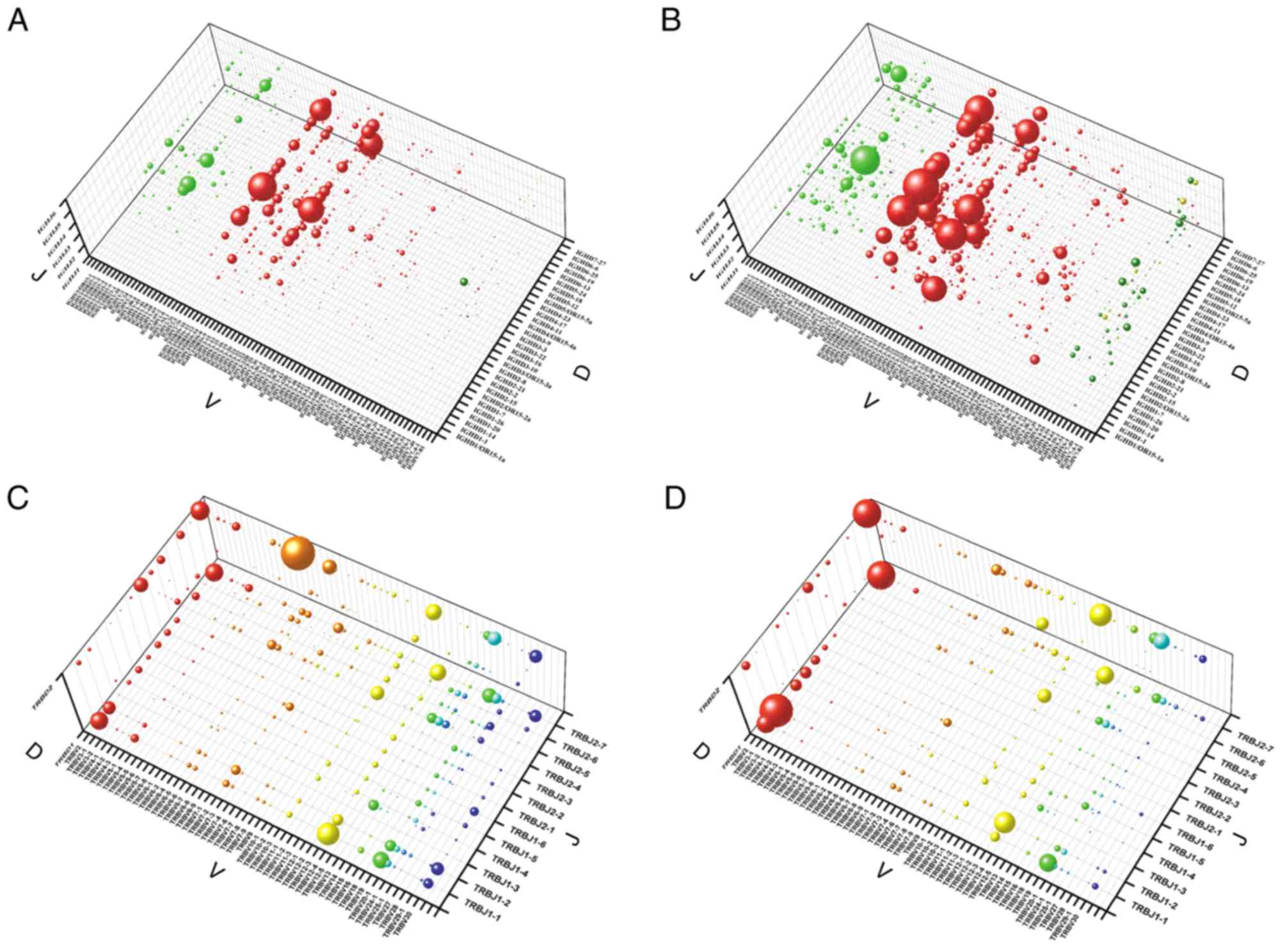
TRBJ. The recombination of genes from the V, D and J gene family in
the BCR/TCR CDR3 appeared to be conserved, and it was demonstrated
that 73 genes in the IGHV subfamily, 30 genes in the IGHD
subfamily, six genes in the IGHJ subfamily, 53 genes in the TRBV
subfamily, two genes in the TRBD subfamily and 12 genes in the TRBJ
subfamily were commonly used in both groups (Fig. 6). However, the VDJ recombination of
BCR/TCR CDR3 exhibited great variation between the two groups of
samples, with the χ2 test indicating that four of the
seven high frequency VDJ gene recombinations (frequency >1.0%),
including IGHV3-11/IGHD3-10/IGHJ6, IGHV3-30/IGHD3-10/IGHJ6,
TRBV6-6/TRBJ2-7/TRBD2 and TRBV12-3/TRBJ1-1/TRBD1, were present with
a significantly higher frequency in the IgAN compared with the
control group (P<0.001; data not shown).
Discussion
B and T cells serve important roles in the human
immune system, and understanding their receptor characteristics in
human autoimmune diseases may help the development of specific
immunotherapeutic interventions for diseases in the clinic
(23). IgAN is widely defined as a
human autoimmune disease, and certain studies have indicated that a
cytokine-mediated switch of naive B cells to antibody-secreting
cells or Toll-like receptor ligation pathways in T-cells may be
important in the pathogenesis of IgAN (6,24).
In the clinical setting, certain classical immunosuppressive drugs
that exhibit pleiotropic effects, including cyclophosphamide and
azathioprine, have been used for the treatment of IgAN (25,26).
With the development of next-generation sequencing, it is possible
to analyze the human immune repertoires in different physiological
and pathological conditions with a high depth, which was not
achievable previously (27,28).
In the present study, a large number of BCR/TCR CDR3
sequences at the DNA level were resolved, providing a large
quantity of novel information regarding immune repertoires in IgAN.
With regard to the common index for the determination of B/T cell
repertoire diversity, the length distribution of the BCR/TCR CDR3
loop appeared to fit well to a Gaussian distribution curve in the
IgAN and control groups. This indicated that there may be no
notable alterations in length distribution compared with certain
typical autoimmune disorders (18). As another common index, the HECs of
BCRs and TCRs exhibited certain differences; for example, the
number of BCR HECs was increased in the IgAN compared with the
control group, whereas the TCR HECs did not exhibit the same trend.
This result indicated that there were more BCR CDR3 sequences with
higher frequencies in the disease condition of IgAN. As IgAN is a
disease characterized by mesangial IgA deposits, a higher frequency
of BCR HECs may contribute to a higher frequency of class-switching
of prime naive B cells to IgA+ antibody-secreting cells
following mucosal infection (6).
BCR/TCR CDR3 peptides are believed to be among the
most complex and diverse amino acid sequences in humans (29). Studies have indicated that BCR/TCR
CDR3 is encoded by somatic rearrangement between IGHV/IGHD/IGHJ
(TRBV/TRBD/TRBJ) gene segments, and that receptor diversity may be
augmented by non-template nucleotide trimming or addition at
different gene segment junction sites (30,31).
Therefore, the original hypervariable sequences of CDR3 may be
encoded randomly and are able to recognize millions of antigens
(32). Notably, public clones of
BCR/TCR CDR3 sequences in the two groups of samples were
identified, including 12 BCR and 228 TCR CDR3 clones, certain of
which were HECs. Furthermore, certain of the BCR/TCR CDR3 residues
appeared to be conserved in different clones. These findings
indicated that the BCR/TCR CDR3 may not be produced randomly.
Public (conserved) B/T cells are known to be key factors affecting
human health (33). Notably, there
were public sequences with significantly different frequencies in
the IgAN group. It is possible that these public clones or residues
with significantly higher frequency may be the initiators of IgAN,
and may represent potential targets for immunotherapeutic
intervention.
Regarding the use of IGHV/IGHD/IGHJ in encoding the
BCR CDR3, there were certain common characteristics in the two
groups of samples, including the most highly used gene segment,
IGHV3-11/IGHD3-10/IGHJ6; this indicated that gene segment usage of
BCR CDR3 may be conserved. Regarding the use of TRBV/TRBD/TRBJ in
the TCR CDR3, there were certain common gene combinations in the
two groups, while the most highly used gene segment in the IgAN
group, TRBV6-6/TRBJ2-7/TRBD2, had a lower frequency in the control
group. Although it may be difficult to fully understand the
different degrees of BCR/TCR (V/D/J) repertoire usage, fully
describing the gene segment usages of BCR/TCR CDR3 may be valuable
for understanding the background of IgAN.
In conclusion, the present study attempted to
profile the entire B- and T-cell repertoires in patients with IgAN
by high-throughput sequencing for the first time, to the best of
our knowledge. The global repertoires contain a wealth of conserved
characteristics, especially the conserved BCR/TCR CDR3 sequences
and the usage of gene segments, which may provide novel information
for the treatment of IgAN.
Acknowledgements
Not applicable.
Funding
Funding was received from Science and Technology
Plan of Shenzhen (grant nos. JCYJ20160422164313440 and
JCYJ20150403093323885).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Author's contributions
YD and JQ carried out the study design, MO
participated in experiment and data analysis, and drafted the
manuscript. FZ and XZ participated in sample collection and
clinical data analysis. SL, DT and PZ participated in experiment
and analysis. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Ethics Committee of
Shenzhen People's Hospital. Written informed consent was obtained
from all patricpants.
Consent for publication
Written informed consent for the publication of any
associated data and accompanying images was obtained from each
participant.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Berthelot L, Papista C, Maciel TT,
Biarnes-Pelicot M, Tissandie E, Wang PH, Tamouza H, Jamin A,
Bex-Coudrat J, Gestin A, et al: Transglutaminase is essential for
IgA nephropathy development acting through IgA receptors. J Exp
Med. 209:793–806. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Riispere Ž, Kuudeberg A, Seppet E, Sepp K,
Ilmoja M, Luman M, Kõlvald K, Auerbach A and Ots-Rosenberg M:
Significance of clinical and morphological prognostic risk factors
in IgA nephropathy: Follow-up study of comparison patient groups
with and without renoprotection. BMC Nephrol. 18:892017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fiorentino M, Bolignano D, Tesar V, Pisano
A, Van Biesen W, D'Arrigo G, Tripepi G and Gesualdo L: ERA-EDTA
Immunonephrology Working Group: Renal biopsy in 2015-from
epidemiology to evidence-based indications. Am J Nephrol. 43:1–19.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Le WB and Liu ZH: Prognostic indicators
and treatment of IgA Nephropathy in China. Pathogen Treat IgA
Nephropat. 127–139. 2016. View Article : Google Scholar
|
|
5
|
Wyatt RJ and Julian BA: IgA Nephropathy. N
Engl J Med. 368:2402–2414. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Boyd JK, Cheung CK, Molyneux K, Feehally J
and Barratt J: An update on the pathogenesis and treatment of IgA
nephropathy. Kidney Int. 81:833–843. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gharavi AG, Kiryluk K, Choi M, Li Y, Hou
P, Xie J, Sanna-Cherchi S, Men CJ, Julian BA, Wyatt RJ, et al:
Genome-wide association study identifies susceptibility loci for
IgA nephropathy. Nat Genet. 43:321–327. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suzuki H, Kiryluk K, Novak J, Moldoveanu
Z, Herr AB, Renfrow MB, Wyatt RJ, Scolari F, Mestecky J, Gharavi AG
and Julian BA: The pathophysiology of IgA nephropathy. J Am Soc
Nephrol. 22:1795–1803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mekori YA, Hershko AY, Frossi B, Mion F
and Pucillo CE: Integrating innate and adaptive immune cells: Mast
cells as crossroads between regulatory and effector B and T cells.
Eur J Pharmacol. 778:84–89. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han Y, Li H, Guan Y and Huang J: Immune
repertoire: A potential biomarker and therapeutic for
hepatocellular carcinoma. Cancer Lett. 379:206–212. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hou X, Lu C, Chen S, Xie Q, Cui G, Chen J,
Chen Z, Wu Z, Ding Y, Ye P, et al: High throughput sequencing of T
cell antigen receptors reveals a conserved TCR repertoire. Medicine
(Baltimore). 95:e28392016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Madi A, Shifrut E, Reich-Zeliger S, Gal H,
Best K, Ndifon W, Chain B, Cohen IR and Friedman N: T-cell receptor
repertoires share a restricted set of public and abundant CDR3
sequences that are associated with self-related immunity. Genome
Res. 24:1603–1612. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu H, Zhang GY, Clarkson AR and Knight JF:
Conserved T-cell receptor beta chain CDR3 sequences in IgA
nephropathy biopsies. Kidney Int. 55:109–119. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
O'Connell AE, Volpi S, Dobbs K, Fiorini C,
Tsitsikov E, de Boer H, Barlan IB, Despotovic JM, Espinosa-Rosales
FJ, Hanson IC, et al: Next generation sequencing reveals skewing of
the T and B cell receptor repertoires in patients with
wiskott-Aldrich syndrome. Front Immunol. 5:3402014.PubMed/NCBI
|
|
15
|
Suzuki K, Honda K, Tanabe K, Toma H, Nihei
H and Yamaguchi Y: Incidence of latent mesangial IgA deposition in
renal allograft donors in Japan. Kidney Int. 63:2286–2294. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee HS, Lee MS, Lee SM, Lee SY, Lee ES,
Lee EY, Park SY, Han JS, Kim S and Lee JS: Histological grading of
IgA nephropathy predicting renal outcome: Revisiting H. S. Lee's
glomerular grading system. Nephrol Dial Transplant. 20:342–348.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hou X, Chen J, Lu C, Wang L and Wu W: The
conserved T cell receptor repertoire observed in patients with
systemic lupus erythematosus. Int J Clin Exp Med. 10:2053–2065.
2017.
|
|
18
|
Liu S, Hou XL, Sui WG, Lu QJ, Hu YL and
Dai Y: Direct measurement of B-cell receptor repertoire's
composition and variation in systemic lupus erythematosus. Genes
Immun. 18:22–27. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guda K, Veigl ML, Varadan V, Nosrati A,
Ravi L, Lutterbaugh J, Beard L, Willson JK, Sedwick WD, Wang ZJ, et
al: Novel recurrently mutated genes in African American colon
cancers. Proc Natl Acad Sci USA. 112:pp. 1149–1154. 2015;
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Meyer EH, Hsu AR, Liliental J, Löhr A,
Florek M, Zehnder JL, Strober S, Lavori P, Miklos DB, Johnson DS
and Negrin RS: A distinct evolution of the T-cell repertoire
categorizes treatment refractory gastrointestinal acute
graft-versus-host disease. Blood. 121:4955–4962. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kumar S, Stecher G and Tamura K: MEGA7:
Molecular evolutionary genetics analysis version 7.0 for bigger
datasets. Mol Biol Evol. 33:1870–1874. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xu H, Wang C, Liu X, Zhao Y and Zhang W:
Characterization of the T cell receptor repertoires during
pregnancy. Biomed Res. 28:1161–1166. 2017.
|
|
23
|
Borroto A, Reyes-Garau D, Jiménez MA,
Carrasco E, Moreno B, Martínez-Pasamar S, Cortés JR, Perona A, Abia
D, Blanco S, et al: First-in-class inhibitor of the T cell receptor
for the treatment of autoimmune diseases. Sci Transl Med.
8:370ra1842016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Renfrow MB and Novak J: What insights can
proteomics give us into IgA nephropathy (Berger's disease)? Expert
Rev Proteomics. 14:645–647. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Rasche FM, Keller F, Rasche WG, Schiekofer
S, Boldt A, Sack U and Fahnert J: Why, when and how should
immunosuppressive therapy considered in patients with
immunoglobulin A nephropathy. Clin Exp Immunol. 186:115–133. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim MJ, Schaub S, Molyneux K, Koller MT,
Stampf S and Barratt J: Effect of immunosuppressive drugs on the
changes of serum galactose-deficient IgA1 in patients with IgA
nephropathy. PLoS One. 11:e01668302016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Callan CG Jr, Mora T and Walczak AM:
Repertoire sequencing and the statistical ensemble approach to
adaptive immunity. Curr Opin Syst Biol. 1:44–47. 2017. View Article : Google Scholar
|
|
28
|
Hou XL, Wang L, Ding YL, Xie Q and Diao
HY: Current status and recent advances of next generation
sequencing techniques in immunological repertoire. Genes Immun.
17:153–164. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Berry R and Call ME: Modular activating
receptors in innate and adaptive immunity. Biochemistry.
56:1383–1402. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Krell PF, Reuther S, Fischer U, Keller T,
Weber S, Gombert M, Schuster FR, Asang C, Stepensky P, Strahm B, et
al: Next-generation-sequencing-spectratyping reveals public T-cell
receptor repertoires in pediatric very severe aplastic anemia and
identifies a β chain CDR3 sequence associated with
hepatitis-induced pathogenesis. Haematologica. 98:1388–1396. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Aouinti S, Malouche D, Giudicelli V,
Kossida S and Lefranc MP: IMGT/HighV-QUEST statistical significance
of IMGT clonotype (AA) diversity per gene for standardized
comparisons of next generation sequencing immunoprofiles of
immunoglobulins and T cell receptors. PLoS One. 10:e01423532015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Saada R, Weinberger M, Shahaf G and Mehr
R: Models for antigen receptor gene rearrangement: CDR3 length.
Immunol Cell Biol. 85:323–332. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kitaura K, Shini T, Matsutani T and Suzuki
R: A new high-throughput sequencing method for determining
diversity and similarity of T cell receptor (TCR) α and β
repertoires and identifying potential new invariant TCR α chains.
BMC Immunol. 17:382016. View Article : Google Scholar : PubMed/NCBI
|