Introduction
Hair is a key component of an individual's
appearance, and in addition to protection, hair serves a role in
social communication (1).
Considering the psychological effects, hair loss has been
associated with a reduction in self-esteem. There are three phases
in the hair growth cycle including the anagen, catagen and telogen
phases, and the hair growth cycle is regulated by several growth
factors, including epithelial growth factor (EGF), insulin-like
growth factor (IGF)-1, fibroblast growth factor (FGF)-5 and −7
(2–4). Dysregulation of growth factors
results in hair loss.
At present, only two drugs, minoxidil and
finasteride, are licenced for the treatment of hair loss by the
United States Food and Drug Administration. There are several
disadvantages to the use of finasteride, including orthostatic
hypotension, chills, cold sweats and confusion. Side effects
associated with minoxidil include a burning or itching sensation of
the skin around the treated area, and hair feeling dry, straw-like
and dull subsequent to termination of the treatment (5). Therefore, it is important to seek
alternative strategies for the treatment of hair loss, with high
efficacy and fewer side effects. The use of traditional Chinese
herbal medicines has been regarded safe for use in and
complementary medicine in various diseases. BeauTop (BT) is mixture
of a variety of Chinese herbs including Ginseng radix, Astragali
radix, Radix Angelicae sinensis, Ligustri fructus, Rehmannia
glutinosa and Eclipta prostrata (Linn). A previous study
demonstrated that there was non-toxicity using BT in a Wistar rat
model (6). The aim of the current
study was to evaluate the promotion of hair growth by BT in C57BL/6
mice and the mechanism involved, for consideration of BT as a
potential therapeutic medication for alopecia.
Materials and methods
Animals
Female 8-week-old C57BL/6 mice weighing 15–20 g were
purchased from the National Laboratory Animal Center (Taiwan,
China). Mice were fed the standard laboratory diet and water ad
libitum and housed under the 12 h light/dark cycle and 21±1°C
in the Animal Technology Institute (Taiwan, China). Susbequent to
euthanasia, specimens of full-thickness skin and subcutaneous
tissue were harvested and stored at −80°C. This study was approved
by the ethics committee of Agricultural Technology Research
Institute (protocol no. 97011) (Miaoli, Taiwan).
Preparation of BT
BT is composed of Ginseng radix, Astragali radix,
Radix Angelicae sinensis, Ligustri fructus, Rehmannia glutinosa
and Eclipta prostrata (Linn) at a ratio of 7:9:7:10:7:10.
All components were purchased from a Chinese herbal shop (Sun Ten
Pharmaceutical Co. Ltd., Taiwan, China). BT at 100 mg was prepared
as a mixture of the above components and was extracted with 1 l of
distilled water at 100°C. The aqueous extracts were powdered by
heating in a rotary vacuum evaporator. BT powder was then dissolved
in distilled water.
Murine model and grouping of
wax/rosin-induced alopecia
The procedures for induction of alopecia by
wax/rosin in mice were performed as described previously (7). A total of 36 mice were divided into
the control and BT groups (18 mice/group). Mice were anesthetized
with Avertin® (2,2,2-tribromoethanol; Sigma-Aldrich;
Merck Millipore, Darmstadt, Germany) by intraperitoneal injection
at a dose of 250 mg/kg. The wax/rosin mixture was applied to the
dorsal skin of mice and each depilatory area was fixed in 4
cm2 area. The procedure induced hair follicles to enter
into telogen phase, as evidenced by the homogeneously pink skin
color on the depilatory area. When the wax/rosin mixture removed
all hair shafts, this immediately causes homogeneous anagen
development over the depilated area. A solution of BT was orally
administrated at a dose of 0.6 g/kg, while the control group was
treated with distilled water of the same volume. Each group was
treated once daily for 12 consecutive days. Mice were sacrificed by
being placed in a CO2 gas chamber.
Photo documentation
Photographs were documented on days 4, 8 and 12 to
provide a record of changes in hair growth. Images were captured
with a Canon PowerShot A470 camera on a Medalight Slim Light Panel.
Photographs were analyzed by gray scale analysis and light
absorption within specified regions was quantified using Image-Pro
Plus software version 7.0 (Media Cybernetics, Inc., Rockville, MD,
USA), with greater absorption corresponding to increased hair
growth.
Expression of growth factors via
immunohistology
Mice were sacrificed at days 4, 8 and 12, 6 mice at
each time point. The specimens of full-thickness skin and
subcutaneous tissue from the dorsal region were obtained. The
specimens were fixed in 4% paraformaldehyde at 4°C overnight and
processed for paraffin sectioning. The longitudinal slices were
processed for routine immunohistological analysis then were
preincubated with ready-to-use blocking solution (Immunotech,
Hamburg, Germany), followed by an incubation with anti-EGF (Novus
Biologicals LLC, Littleton, CO, USA), anti-IGF-1, anti-FGF-5 and
anti-FGF-7 antibodies (Abbiotec LLC San Diego, CA, USA).
Development was performed using a diaminobenzidine
tetrahydrochloride kit (Vector Laboratories, Inc., Burlingame, CA,
USA). An Olympus microscope equipped with an AxioCam ICc 3 imaging
system was used to randomly screen the field. Image-Pro Plus
software was used to calculate the percentage color area.
Statistical analysis
Data are presented as the mean ± standard deviation
unless otherwise stated. Comparison between groups was performed
using independent t-test analysis for comparison of treatment and
control groups (SPSS software, version 12.0; SPSS, Inc., Chicago,
IL, USA).
Results
Effect of BT on hair growth in mice
model
The results of the hair follicle counting
demonstrated that the BT group was significantly higher than
control group on day 8 (P<0.05; Fig. 1). These results indicated that BT
possessed superior hair growth properties compared with the
control.
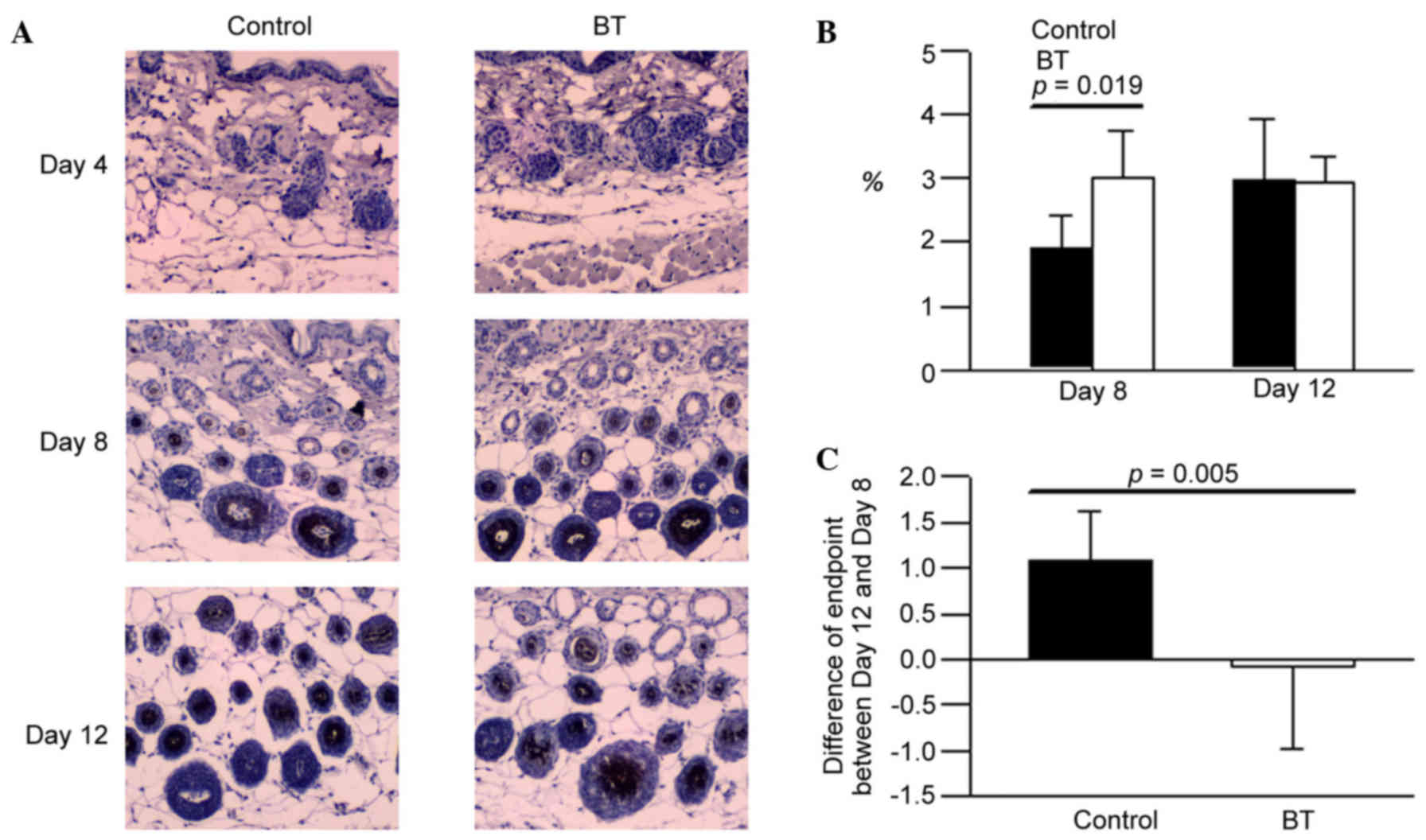
Effect of BT on EGF expression
The expression levels of EGF in the BT treatment
group on day 8 were higher compared with the control (Fig. 2A). According to the experimental
data, it was suggested that BT induced EGF production at the
depilation area of the hair follicle around day 8 (P<0.05;
Fig. 2B and C). However, BT did
not influence on EGF expression on day 12 (Fig. 2B).
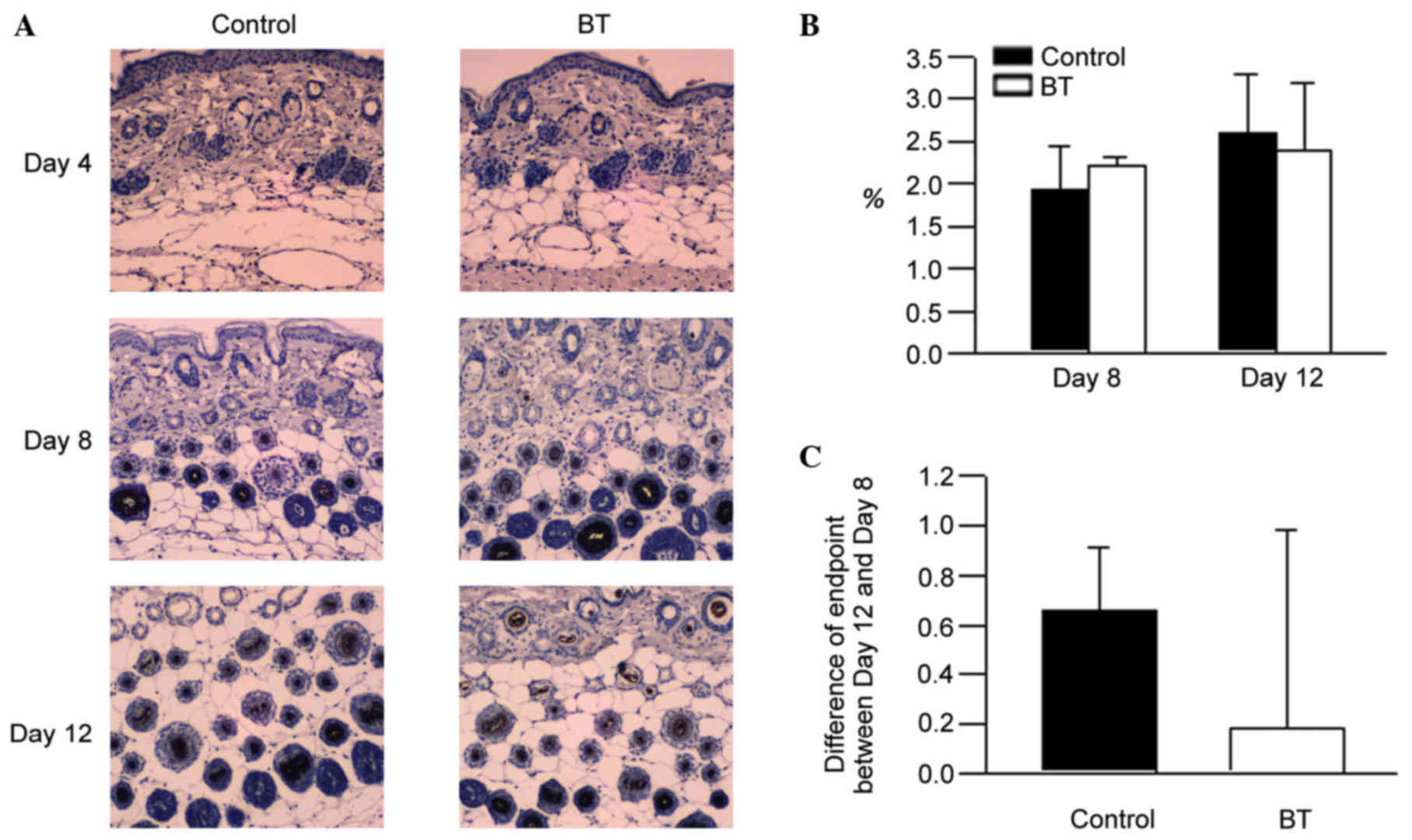
Effect of BT on IGF-1 expression
Up to day 12, BT did not result in a significant
change to the IGF-1 expression levels, as presented in Fig. 3.
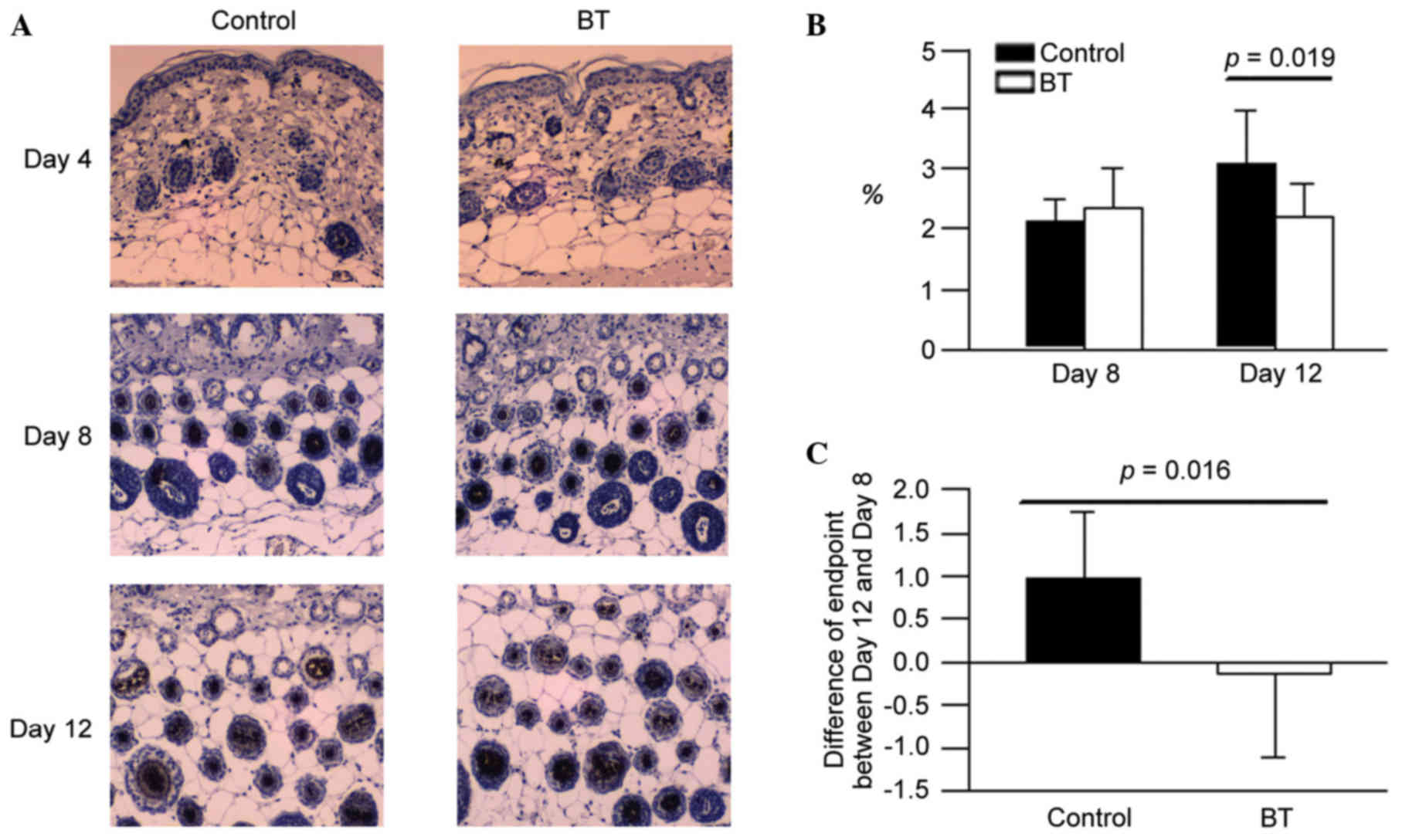
Effect of BT on FGF-5 expression
The expression levels of FGF-5 in the BT treatment
group on day 8 were observed to exhibit a lower area percentage
compared with the control (Fig.
4A). According to the experimental data, it was suggested that
BT reduced the FGF-5 production at the depilation area of the hair
follicle around day 12 (P<0.05; Fig. 4B and C). However, BT did not
influence on FGF-5 expression on day 8 (Fig. 4B).
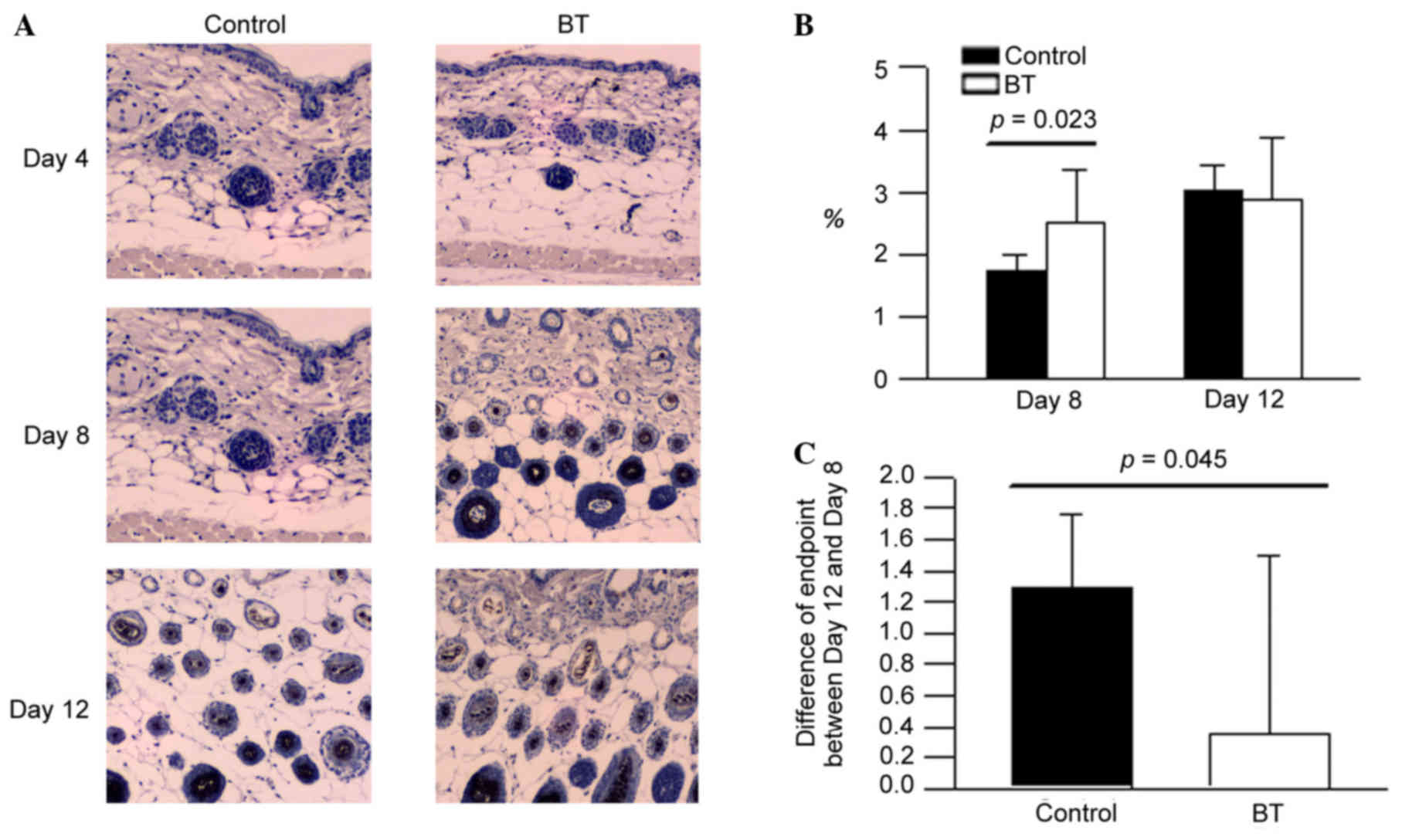
Effect of BT on FGF-7 expression
The expression levels of FGF-7 in the BT treatment
group on day 8 presented with a higher area percentage compared
with the control (Fig. 5A).
According to the experimental data, it was presumed that BT induced
the FGF-7 production at the depilation area of hair follicle around
day 8 (P<0.05; Fig. 5B and C).
However, BT did not influence FGF-7 expression at day 12 (Fig. 5B).
Discussion
Hair follicle counts demonstrated that the BT group
was significantly higher than control on day 8. From
histopathological results, EGF and FGF-7 of the BT group on day 8
presented with a higher area percentage than control, respectively.
These data suggested that BT may induce the EGF and FGF-7
production at the depilation area of hair follicles around day 8,
and additionally may reduce the FGF-5 production on day 12.
The primary ingredients of BT include Ginseng
radix, Astragali radix, Radix Angelicae sinensis, Ligustri fructus,
Rehmannia glutinosa and Eclipta prostrata (Linn).
Angiogenesis serves an important role in a wide range of
physiological processes and numerous diseases are associated with
dysregulation of angiogenesis. Ginsenoside-Re is one of the major
active components of saponin in Panax ginseng. Angiogenesis
in the ECMs was significantly enhanced by loading with basic FGF or
ginsenoside-Re (8). Promotion of
basal cell proliferation and angiogenesis by the Astragali
radix extracts was marked in the early stages of wound healing,
resulting in a significant reduction in the duration of the
wound-healing process (9).
Astragali radix contains quercetin, which has been
demonstrated to posess anti-inflammatory properties. As an
anti-inflammatory, quercetin has been observed to inhibit the
activation of NF-κB, a transcription factor that stimulates the
production of proinflammatory cytokines including tumor necrosis
factor (TNF)-α and interleukin (IL)-1, IL-2 and IL-6 (10). The natural compound quercetin has
been previously suggested to possess potential effects for alopecia
areata (AA) (11). During the
catagen stage in mice, hair follicles are also destroyed by an
inflammatory-cell infiltrate, in an apparent physiological process
of programmed organ deletion (12). From the above information, it was
hypothesized that BT may have potential for treatment of AA and
androgenetic alopecia (AGA) though the immunomodulatory effect
protecting hair follicles from immune attacks. BT was additionally
suggested to act through anti-inflammatory properties to protect
hair follicles from inflammation, which may be beneficial in AGA,
however further studies are required in order to evaluate this. The
effects of Rehmannia glutinosa oligosaccharide (RGO), a
traditional Chinese medicine, have been investigated on the
proliferation, H2O2-induced apoptosis and
secretion of vascular endothelial growth factor (VEGF) and
hepatocyte growth factor (HGF) of human adipose-derived mesenchymal
stem cells (ADMSCs) in vitro. One study demonstrated that
RGO may increase the viability and proliferative capacity and
alleviate H2O2-induced apoptosis of human
ADMSCs via the paracrine release of VEGF and HGF. These results
indicate that RGO application will enhance stem cell viability and
improve their effects in cell therapy (13). Butanol extracted from Eclipta
prostrata (Linn) has been demonstrated to produce an
anti-oxidative effect in cesarean-derived rats, and saponin is a
primary ingredient in butanol extract, which has been observed to
result in in-vitro anti-oxidative effects (14). Further study is required to
determine whether these effects are associated with the effect of
BT on improving AGA.
Normal expression of both EGF and its receptor
(EGFR) in the outer root sheath has been identified to be
downregulated with the completion of follicular growth, and
continuous expression of EGF in hair follicles of transgenic mice
arrested follicular development at the final stage of morphogenesis
(15). Binding of EGF or of
transforming growth factor α or other associated factors activates
the receptor and induces cell proliferation and differentiation
(16). The TNF-α-induced
activation of the extracellular signal-related kinase/guanine
nucleotide exchange factor H1/ras homolog family member A pathway
in tubular cells is mediated through Src- and TNF-α convertase
enzyme-dependent EGFR activation. This mechanism may couple
inflammatory and proliferative stimuli and, thus, may serve a key
role in the regulation of wound healing and fibrogenesis (17). From the results of the current
study, BT induced the EGF production significantly at the
depilation area of hair follicles around day 8, however did not
cause a significant change on day 12. It was demonstrated that the
BT has the potential effect to improve hair growth through
upregulating the expression of EGF. In the cycle of normal hair
follicles, the expression in EGF will stop increasing to avoid the
overgrowth of hair; and it was observed that the expression of EGF
did not significantly increase with BT on day 12.
The IGF-1 receptor (IGF-1R) is the only IGF receptor
to have IGF-mediated signaling functions. In circulation, this
growth factor mediates endocrine action of growth hormones on
somatic growth and is bound to specific binding proteins. The
latter control IGF transport, efflux from vascular compartments and
association with cell surface receptors. In tissues, IGF-1 is
produced by mesenchymal type cells and acts in a IGF-1 paracrine
and autocrine fashion by binding to the IGF-1R. This binding
activates the receptor tyrosine kinase that triggers the downstream
responses and finally stimulates cell division (18). The androgen metabolite
dihydrotestosterone (DHT) inhibits hair growth by negatively
modulating growth-regulatory effects of dermal papillae. DHT may
reduce IGF-1 production in dermal papillae by inhibiting sensory
neuron stimulation through interaction with the androgen receptor,
thereby inhibiting hair growth in mice (3). According to the experimental data, BT
did not induce IGF-1 production significantly on day 8 or 12. It is
suggested that BT may improve hair growth through mechanisms other
than IGF-1.
The growth and development of hair follicles is
influenced by a number of different growth factors and cytokines,
particularly members of the FGF family (19). FGF-5 is a secreted signaling
protein and inhibits hair elongation, which is suggested to be via
regulating a step in the progression (20). FGF signal transduction involves
ligand binding to a high-affinity receptor-tyrosine kinase. FGF-5
RNA was detected in the outer root sheath surrounding the transient
portion of the follicle and also in a small population of cells at
the base of the follicle in a previous study (21). It has been reported that the gene
for murine FGF-5 is expressed in the rat hair follicle and that
this expression may be associated with catagen induction (20). Mutational analysis has demonstrated
that FGF signaling mediated by FGF-5 serves a key role in
regulating the transition from anagen to catagen (21), and FGF-5 has been demonstrated to
be a particularly potent inhibitory factor (22). Other FGF family members that are
expressed during the hair growth cycle are FGF-1, FGF-2 and FGF-7
(21). Keratinocyte growth factor,
also known as FGF-7, is synthesized by skin fibroblasts (21). The 18.9 kD member of the FGF family
has been demonstrated to selectively induce keratinocyte
proliferation and differentiation in tissue culture (23). From the results of the current
study, it is suggested that BT induced the FGF-7 production at the
depilation area of hair follicles around day 8. FGF-5 is potent
inhibitor, and it was identified that BT reduced the FGF-5
production on day 12. This indicated that BT additionally has
potential to improve hair growth through upregulating the
expression of FGF-7 on day 8 and downregulating expression of FGF-5
on day 12. In the cycle of normal hair follicles, the expression of
FGF-7 will stop increasing to avoid the overgrowth of hair, and it
was observed that the expression of FGF-7 did not significantly
increase in the BT group on day 12.
These studies in mice indicated a significant
increase in the number of follicles per unit area in the BT group
compared with the controls on day 8. Thus, BT has the potential to
improve hair growth through upregulating the expression of EGF and
FGF-7 and through downregulating the expression of FGF-5. However,
no clinical trial investigating the use of BT and its effects of
inhibiting human alopecia or promoting hair growth has been
conducted, thus a trial should be undertaken in order to verify the
effects of BT in humans.
These results have demonstrated that the mechanism
of action of BT in improving AGA is potentially associated with
EGF, FGF-5 and FGF-7. Taken together, these results suggest that BT
has the potential effect of promoting hair growth.
Acknowledgements
The authors will like to thank the non-profit
organization Brion Research Institute of Taiwan (Taiwan, China) and
Chung Shan Medical University (protocol no. CSMU-G104N0002) for its
sponsorship, which has contributed to the completion of the present
study.
References
|
1
|
Lourith N and Kanlayavattanakul M: Hair
loss and herbs for treatment. J Cosmet Dermatol. 2:210–222. 2013.
View Article : Google Scholar
|
|
2
|
Alexandrescu DT, Kauffman CL and Dasanu
CA: The cutaneous epidermal growth factor network: Can it be
translated clinically to stimulate hair growth? Dermatol Online J.
15:12009.
|
|
3
|
Zhao J, Harada N and Okajima K:
Dihydrotestosterone inhibits hair growth in mice by inhibiting
insulin-like growth factor-I production in dermal papillae. Growth
Horm IGF Res. 21:260–267. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yun YR, Won JE, Jeon E, Lee S, Kang W, Jo
H, Jang JH, Shin US and Kim HW: Fibroblast growth factors: Biology,
function and application for tissue regeneration. J Tissue Eng.
2010:2181422010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rossi A, Cantisani C, Melis L, Iorio A,
Scali E and Calvieri S: Minoxidil use in dermatology, side effects
and recent patents. Recent Pat Inflamm Allergy Drug Discov.
6:130–136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee CY, Wei CC, Yu MC, Lin CC, Sheu SJ,
Yang JH, Chiang CY, Huang KH and Kuan YH: Hair growth effect of
traditional Chinese medicine BeauTop on androgenetic alopecia
patients: A randomized double-blind placebo-controlled clinical
trial. Exp Ther Med. (In press).
|
|
7
|
Katikaneni R, Ponnapakkam T, Seymour A,
Sakon J and Gensure R: Parathyroid hormone linked to a collagen
binding domain promotes hair growth in a mouse model of
chemotherapy-induced alopecia in a dose-dependent manner.
Anticancer Drugs. 25:819–825. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang YC, Chen CT, Chen SC, Lai PH, Liang
HC, Chang Y, Yu LC and Sung HW: A natural compound (ginsenoside Re)
isolated from Panax ginseng as a novel angiogenic agent for
tissue regeneration. Pharm Res. 22:636–646. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Han DO, Lee HJ and Hahm DH: Wound-healing
activity of Astragali Radix in rats. Methods Find Exp Clin
Pharmacol. 31:95–100. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Bhaskar S, Shalini V and Helen A:
Quercetin regulates oxidized LDL induced inflammatory changes in
human PBMCs by modulating the TLR-NF-κB signaling pathway.
Immunobiology. 216:367–373. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wilasrusmee C, Kittur S, Siddiqui J, Bruch
D, Wilasrusmee S and Kittur DS: In vitro immunomodulatory effects
of ten commonly used herbs on murine lymphocytes. J Altern
Complement Med. 8:467–475. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Eichmüller S, van der Veen C, Moll I,
Hermes B, Hofmann U, Müller-Röver S and Paus R: Clusters of
perifollicular macrophages in normal murine skin: Physiological
degeneration of selected hair follicles by programmed organ
deletion. J Histochem Cytochem. 46:361–370. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang Y, Wang Y, Wang L, Zhang Y, Qin Y,
Chen T, Han W and Chen G: Effects of rehmannia glutinosa
oligosaccharide on human adipose-derived mesenchymal stem cells in
vitro. Life Sci. 91:1323–1327. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim DI, Lee SH, Hong JH, Lillehoj HS, Park
HJ, Rhie SG and Lee GS: The butanol fraction of Eclipta
prostrata (Linn) increases the formation of brain acetylcholine
and decreases oxidative stress in the brain and serum of
cesarean-derived rats. Nutr Res. 30:579–584. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mak KK and Chan SY: Epidermal growth
factor as a biologic switch in hair growth cycle. J Biol Chem.
278:26120–26126. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miettinen PJ, Berger JE, Meneses J, Phung
Y, Pedersen RA, Werb Z and Derynck R: Epithelial immaturity and
multiorgan failure in mice lacking epidermal growth factor
receptor. Nature. 376:337–341. 1995. View
Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kakiashvili E, Dan Q, Vandermeer M, Zhang
Y, Waheed F, Pham M and Szászi K: The epidermal growth factor
receptor mediates tumor necrosis factor-alpha-induced activation of
the ERK/GEF-H1/RhoA pathway in tubular epithelium. J Biol Chem.
286:9268–9279. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Su HY, Hickford JG, Bickerstaffe R and
Palmer BR: Insulin-like growth factor 1 and hair growth. Dermatol
Online J. 5:11999.PubMed/NCBI
|
|
19
|
Danilenko DM, Ring BD, Yanagihara D,
Benson W, Wiemann B, Starnes CO and Pierce GF: Keratinocyte growth
factor is an important endogenous mediator of hair follicle growth,
development and differentiation. Normalization of the nu/nu
follicular differentiation defect and amelioration of
chemotherapy-induced alopecia. Am J Pathol. 147:145–154.
1995.PubMed/NCBI
|
|
20
|
Hébert JM, Rosenquist T, Götz J and Martin
GR: FGF5 as a regulator of the hair growth cycle: Evidence from
targeted and spontaneous mutations. Cell. 78:1017–1025. 1994.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rosenquist TA and Martin GR: Fibroblast
growth factor signalling in the hair growth cycle: Expression of
the fibroblast growth factor receptor and ligand genes in the
murine hair follicle. Dev Dyn. 205:379–386. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Harmon CS and Nevins TD: IL-1 alpha
inhibits human hair follicle growth and hair fiber production in
whole-organ cultures. Lymphokine Cytokine Res. 12:197–203;.
1993.PubMed/NCBI
|
|
23
|
Pierce GF, Yanagihara D, Klopchin K,
Danilenko DM, Hsu E, Kenney WC and Morris CF: Stimulation of all
epithelial elements during skin regeneration by keratinocyte growth
factor. J Exp Med. 179:831–840. 1994. View Article : Google Scholar : PubMed/NCBI
|