Introduction
Foeniculum vulgare is from the Umbelliferae
family of plants and is used as an antiaging agent to promote
longevity. It is more commonly called fennel and widely used as a
medicine especially in Asia, but also in other places worldwide
(1,2). Traditional practices and the
scientific assessment of fennel indicate that it is a widely
applied herbal plant in human health care in many developing and
developed countries (3). In fact,
it has been attested to have a number of therapeutic properties for
the treatment of internal organs, especially in the endocrine,
gastrointestinal tract and respiratory systems (4–6). In
addition, fennel is widely used for the treatment of inflammatory
diseases in conventional medicine (7).
The volatile chemical composition of wild fennel
samples harvested in different geographical areas has been widely
reported (8). Trans-anethole
[1-Methoxy-4-(E)-propenyl-benzene] is an organic compound isolated
from fennel (9). The uses of
fennel have been evaluated in a number of biopharmacological
studies that suggest that the various constituents of fennel
(seeds, leaves and fruits) are rich in molecules with antioxidant
activity (10–12). Furthermore, the extracts and
components of fennel have been reported to have anti-inflammatory,
anti-bacterial and antiviral effects (13–15).
In 1967, acute lung injury (ALI) was defined as mild
acute respiratory distress syndrome (ARDS), with a P/F ratio of
<300 mmHg (40 kPa) and positive end-expiratory
pressure/continuous positive airway pressure levels of >5 cm
H2O, according to the Berlin Definition (16). Classical signs of ALI include
accumulation of neutrophils, increased alveolar fluid and
pro-inflammatory cytokines in the lungs (17). Worldwide, 5% of the population
succumb to ALI per year. This syndrome involves acute inflammation
of the bronchi in the lungs. The clinical manifestations of ALI are
a productive cough, wheezing, shortness of breath, fever and chest
distention. Regeneration of the alveolar epithelium after ALI is
important for maintaining homeostasis, and the latest studies
demonstrate that the immune system may influence the protection of
epithelial surfaces by increasing barrier function and supporting
healing (18,19).
A recent study demonstrated that CD4+ T
lymphocytes, particularly T helper 17 (Th17) cells and regulatory T
cells (Tregs) are an essential subject for research into the
etiology or pathogenesis of ARDS. Th17 cells, a recently identified
active subdivision of CD4+ T cells and the main origin
of interleukin (IL)-17 (IL-17A), play a major role in protecting
the body from microorganisms, such as Staphylococcus aureus
(20,21). Patients who suffer from ALI/ARDS
have increased levels of T-cell proliferation and activation,
especially Th17 cells (22). Tregs
act in an anti-inflammatory role primarily by contact-dependent
destruction of immunocytes or release of cytokines like IL-10
(23). A reduction in the
proportion of Treg cells or a decrease in Treg cell function is
associated with a number of autoimmune disorders (24). Tregs alter innate immune reactions
during lung injury and may play a role in treating ALI as indicated
by Guo et al (25). Mature
Tregs can be classified as inducible Tregs (iTregs) or natural
Tregs (nTregs). iTregs are able to perform their inhibitory
function by producing immunosuppressive cytokines like IL-10
(26). Tregs differ from Th17
cells and have a powerful pro-inflammatory action by generating
IL-17. In pre-clinical and clinical trials on pulmonary and
autoimmune diseases, Th17 cells have been reported to play a role
(27–30). The IL-17 from Th17 cells inhibits
Treg differentiation (31,32). Additionally, IL-17A could be a
pro-inflammatory cytokine and may play a key role in ALI induced by
lipopolysaccharide (LPS), as shown by previous studies (33,34).
In addition, lung injury by Bordetella pertussis is promoted
by increased levels of Th17 cells (35). However, the main mechanism of
CD4+ T cells in ALI requires further investigation.
These studies have revealed that mice with LPS-induced lung injury
may be treated by both Cozaar™ and alanylglutamine,
which act by decreasing the Th17 cell immune responses and
affecting the Th17/Treg balance in favor of Treg cells (36). The pathological mechanism of
autoimmune and inflammatory disease involves a balance between Th17
and Tregs. A Treg/Th17 imbalance is associated with the occurrence
of autoimmune diseases. Further investigation into these cell
populations in airway inflammation may improve the understanding of
the immune mechanism of lung disease.
In the present study, we first established a model
of LPS-induced ALI and then treated the mice with fennel volatile
oil. The volatile oil reduced exudation in the lungs and also
significantly reduced the quantity of leukocytes infiltrating into
bronchoalveolar lavage fluid (BALF). The above results showed that
the volatile oil of fennel could inhibit ALI in mice.
As a result of the comprehensive analysis of the
function of the fennel active constituent trans-anethole in the
literature, we know that it has anti-inflammatory, insecticidal
activity and estrogen activity (37). In our study we investigated the
effect of trans-anethole on lung inflammation in ALI, as well as
the related modalities for the function of Th17 and Tregs.
Materials and methods
Animals and cell culture
According to the guidelines of the Tianjin
University of Traditional Chinese Medicine (Tianjin, China) and the
Animal Ethics Committee of Laboratory Animal Care, all animal
studies received ethical approval. Sixty healthy 5-week-old male
C57BL/6 mice (18–22 g) were ordered from the Animal Research Center
of the Chinese Academy of Medical Sciences (Beijing, China) and
were housed under suitable conditions. Mice were raised on standard
water and food.
Drugs and reagents
LPS, dexamethasone and trans-anethole were purchased
from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Oligo DNA was
purchased from Shanghai Bio-engineering Co. (Shanghai, China).
TransScript First-Strand cDNA Synthesis SuperMix was obtained from
TransGen Biotech (Beijing, China). SuperReal PreMix Plus was
obtained from Tiangen (Beijing, China). PE-labeled anti-mouse IL-17
and IL-10, fluorescein isothiocyanate (FITC)-labeled anti-mouse
CD4, Alexa Fluor® 647-labeled anti-mouse Foxp3,
anti-mouse M1/70+ and anti-mouse F4\80+ were purchased from Santa
Cruz Biotechnology, Inc. (Dallas, TX, USA). The PE anti-mouse Foxp3
Staining Set was from eBioscience (Thermo Fisher Scientific, Inc.,
Waltham, MA, USA).
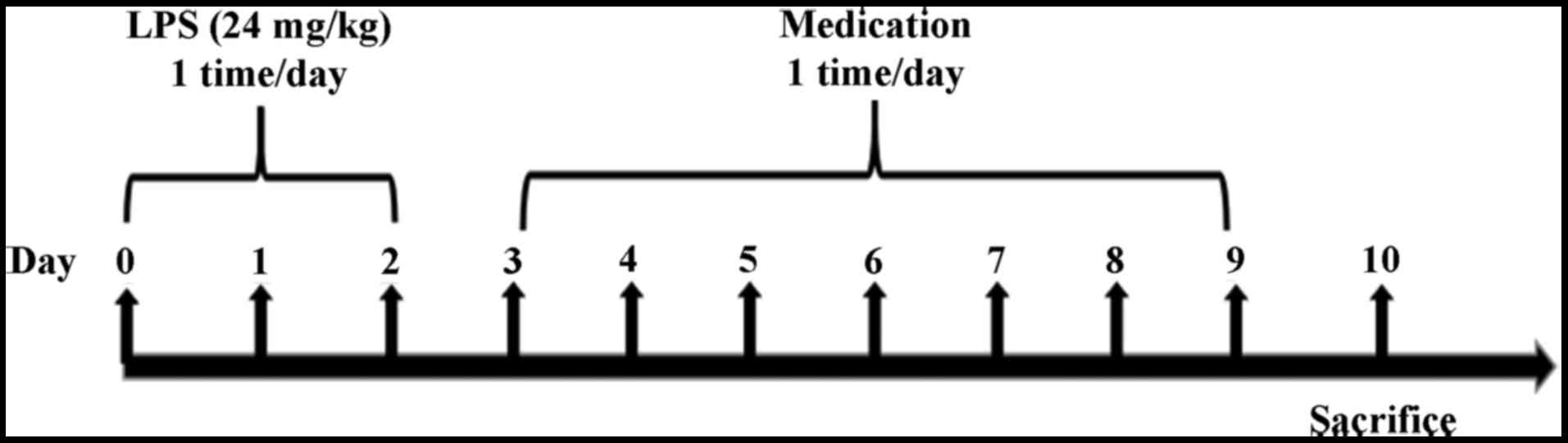
Animal model and experimental
procedures
These sixty mice were allowed to adapt to normal
laboratory conditions during the experiment. The experimental
groups (10 mice/group) were as follows: normal control, LPS-induced
model, trans-anethole treatment (low: 36.4 mg/kg; medium: 72.8
mg/kg; and high: 145.6 mg/kg) and dexamethasone treatment (5
mg/kg). As presented in Fig. 1,
the pulmonary delivery model was established in normal mice. The
C57BL/6 mice bronchitis model was induced by intratracheal
instillation of LPS suspension in normal saline (24 mg/kg) for 3
days. On the 4th day, the mice were administered trans-anethole
(36.4, 72.8 or 145.6 mg/kg) and dexamethasone (5 mg kg/kg) once a
day for 7 consecutive days. The mice were sacrificed 24 h after the
end of treatment.
Histopathology analysis
The lung tissue was stained with hematoxylin and
eosin and the inflammatory changes and morphological lesions were
analyzed using a light microscope (magnification, ×400).
Inflammatory cell count in BALF
A total of 50 µl/g sterile saline was lavaged twice,
then the BALF was collected and centrifuged at 3,000 × g for 15
min. The white blood cells in the BALF were divided and counted
into two types, containing monocytes and eosinophils.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis in spleen and lung
tissue
Total RNA from the isolated spleen and lung tissues
was reverse transcribed into first strand cDNA, according to the
manufacturer's instructions. The relative mRNA levels of samples
were measured using a NanoDrop 2000. qPCR was performed with
sequence-specific primer pairs to execute a 40-cycle two-step PCR
procedure. The qPCR thermocycling conditions were as follows: 95°C
for 3 min, 40 cycles of 95°C for 15 sec and 60°C for 30 sec, then
95°C for 30 sec. Primers were designed by Primer Premier 3.0
(Premier, Vancouver, BC, Canada) and were as follows: IL17,
forward, 5′-CCTCAAAGCTCAGCGTGTCC-3′ and reverse,
5′-GAGCTCACTTTTGCGCCAAG-3′; IL-10, forward, 5′-AAGTGATGCCCCAGGCA-3′
and reverse, 5′-TCTCACCCAGGGAATTCAAA-3′; GAPDH, forward,
5′-AACGGATTTGGCCGTATTGG-3′ and reverse, 5′-CCTTGACTGTGCCGTTGAAT-3′.
Gene expression of IL-17 and IL-10 was measured with qPCR using
SuperReal PreMix Plus (Tiangen). The fold-change of every gene was
quantified by the 2−∆∆Cq method and the internal control
was GAPDH gene. The data were normalized to the internal reference
gene.
Th17/Treg examination in the splenic
tissue
Flow cytometry instrument (BD Biosciences, Franklin
Lakes, NJ, USA) was used to measure Th17 and Tregs in the splenic
tissue. As described in the manufacturer's protocol, the spleen
cells were stained with the FITC anti-mouse CD4 antibody, then
fixed and permeabilized with PE anti-mouse Foxp3 Staining Set.
Then, the cells were stained with anti-mouse IL-10 and anti-mouse
IL-17 (eBioscience; Thermo Fisher Scientific, Inc.).
IL-17 and IL-10 cytokine expression in
CD4+ T cells isolated from the spleen of WT mice
The spleen cells were stained using anti-CD4
antibodies, incubated and fixed with IL-10 and IL-17 staining sets.
According to the manufacturer's instructions, the cells were
stained with PE-labeled anti-mouse IL-17 and IL-10 and then tested
by flow cytometry (BD Biosciences).
Statistical analysis
All data are presented as the mean ± standard
deviation of the mean and compared by one-way analysis of variance
followed by Tukey's post hoc test. All data were analyzed by SPSS
19 (SPSS, Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
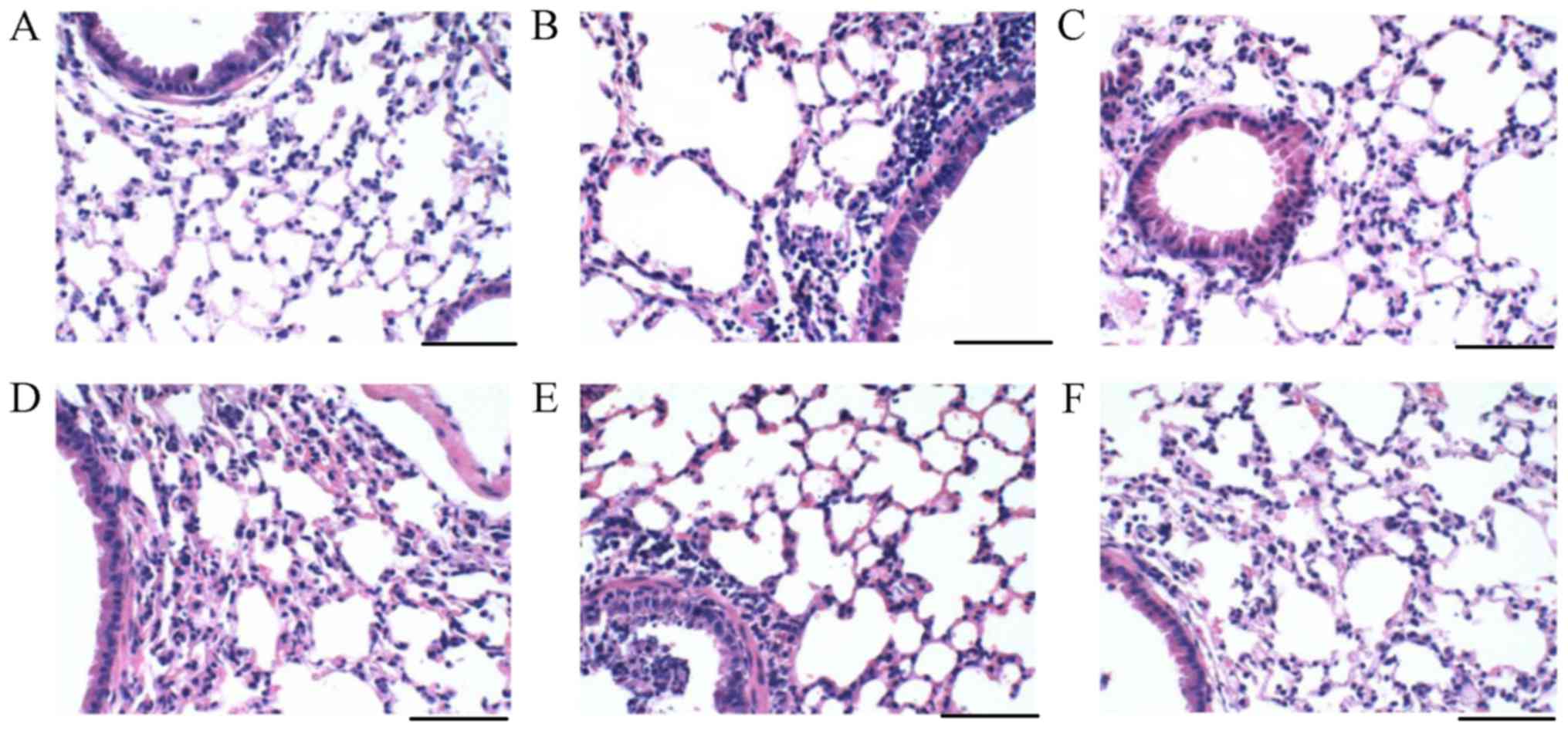
Trans-anethole attenuates airway
inflammation in ALI mice
Lung tissues were collected in each group. From
analyzing the histological results, the lung sections of the normal
group exhibited normal architecture and there was no obvious
neutrophils infiltration, erythrocytes or protein exudate (Fig. 2A). However, severe protein
exudation and lung cell infiltration were demonstrated in the ALI
mouse model (Fig. 2B). These were
relieved after trans-anethole and dexamethasone treatments.
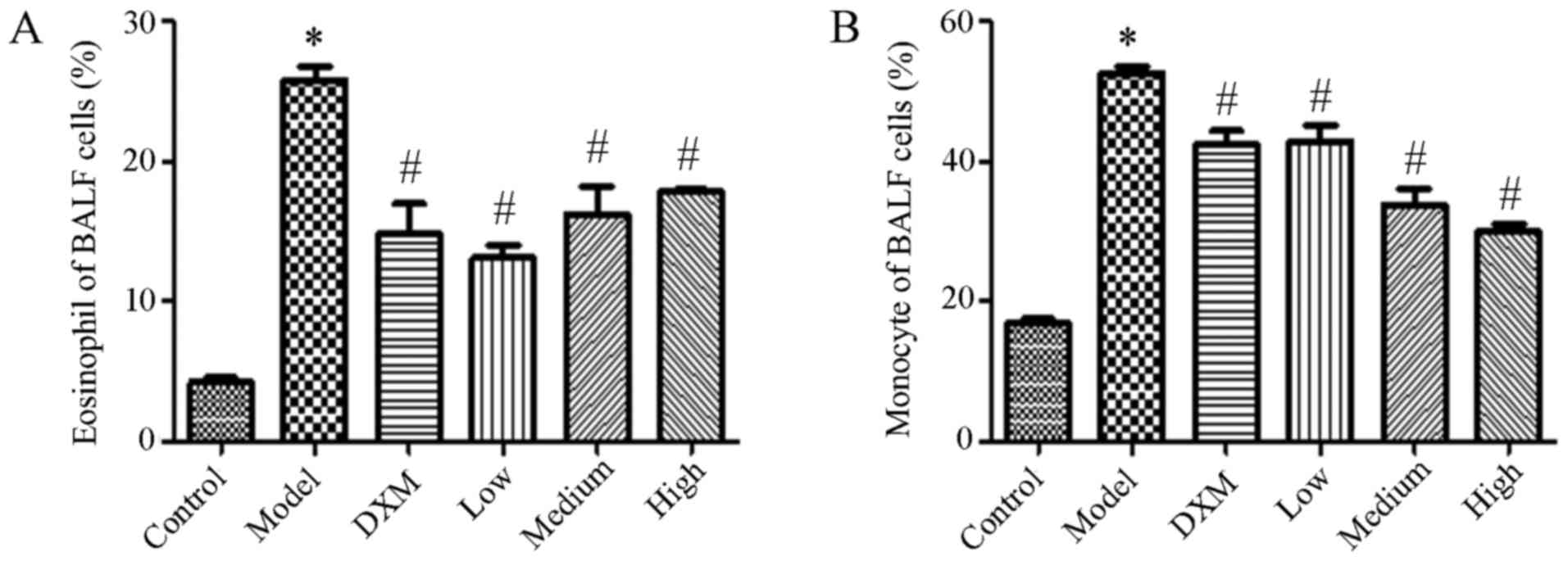
Trans-anethole decreases the number of
inflammatory cells in the BALF of ALI mice
The number of inflammatory cells was measured in the
BALF. An increase in the level of leukocytes was caused by LPS with
both eosinophilic granulocytes and mononuclear cells being enhanced
compared with the normal group. Compared with model group,
Trans-anethole treatment significantly decreased the level of
inflammation cells in the BALF. Compared with dexamethasone, the
numbers of eosinophils and monocytes were not significantly
different in the BALF compared with the trans-anethole group. The
number of monocytes was reduced in the high group compared with the
other groups. As expected, the level of inflammatory cells was
markedly decreased in BALF following trans-anethole treatment
(Fig. 3).
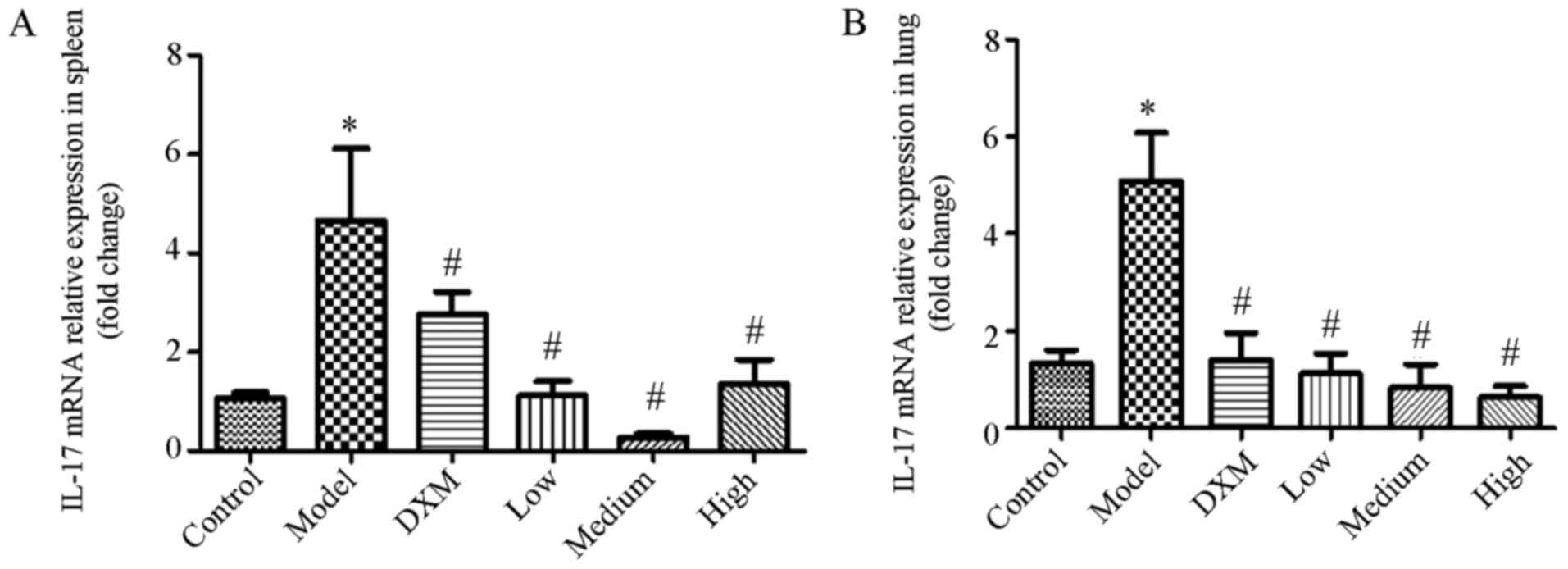
Trans-anethole regulates cytokine mRNA
expression in isolated spleen and lung tissues of ALI mice
Both anti-inflammatory cytokine IL-10 and
pro-inflammatory cytokine IL-17 are critical for inflammation.
Tregs can decrease inflammation via the production of
immunosuppressive agents such as IL-10. To identify the role of
trans-anethole on IL-17 and IL-10 expression, we measured the
expression of both cytokines in the lung and splenic tissue of mice
in each group.
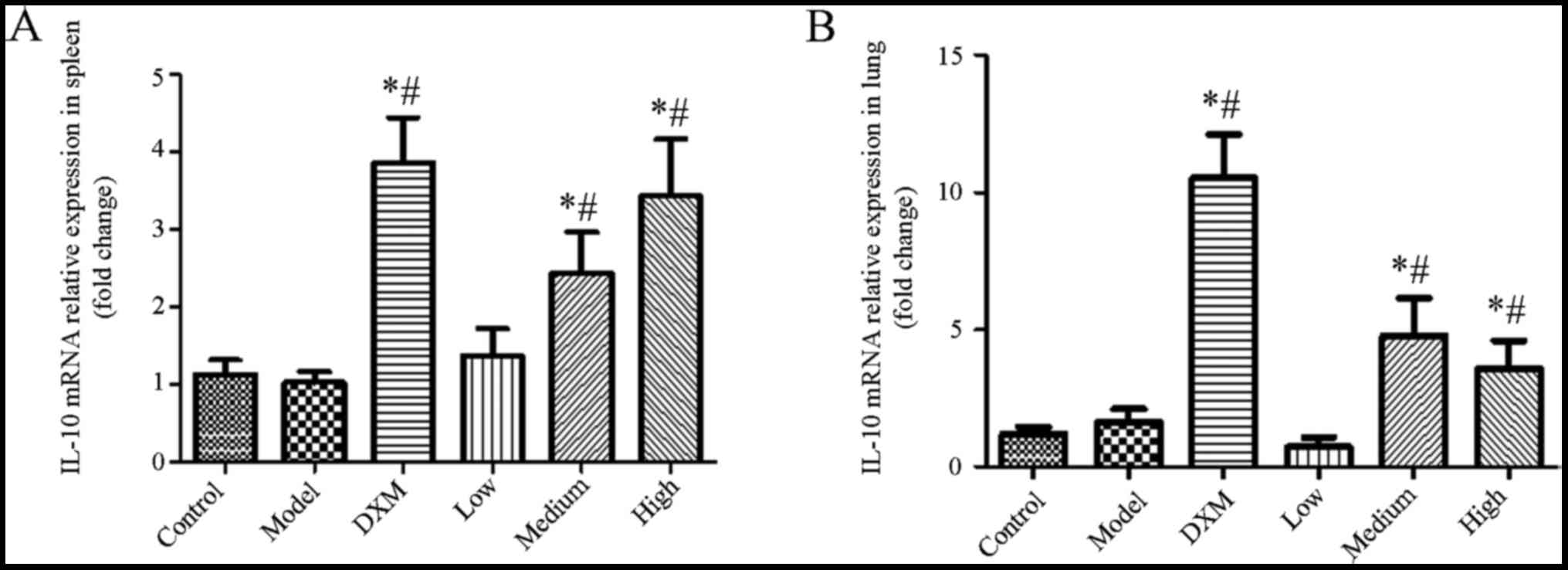
As shown in Fig. 4,
IL-17 was markedly elevated in the model group compared with the
normal group in the spleen and lung tissue. The level of IL-17 was
significantly reduced in the trans-anethole group compared with the
model groups. Compared with dexamethasone, the level of IL-17 was
decreased in the trans-anethole group. The level of IL-17 in spleen
and lung tissue was significantly reduced in the trans-anethole
medium group compared with the other groups.
As shown in Fig. 5,
the level of IL-10 was not changed in the model group compared with
the normal group in the spleen and lung tissue. Although compared
with the dexamethasone group, the level of IL-10 was slightly
decreased in the spleen and lung tissue in the trans-anethole
group, trans-anethole treatment dramatically increased IL-10
expression compared with the model group in the spleen and lung
tissue. These results showed that trans-anethole can decrease IL-17
and increase IL-10 in ALI mice, which demonstrates the
effectiveness of the treatment on lung inflammation.
Trans-anethole controls the
proportions of Th17 and Treg in the spleen of ALI mice
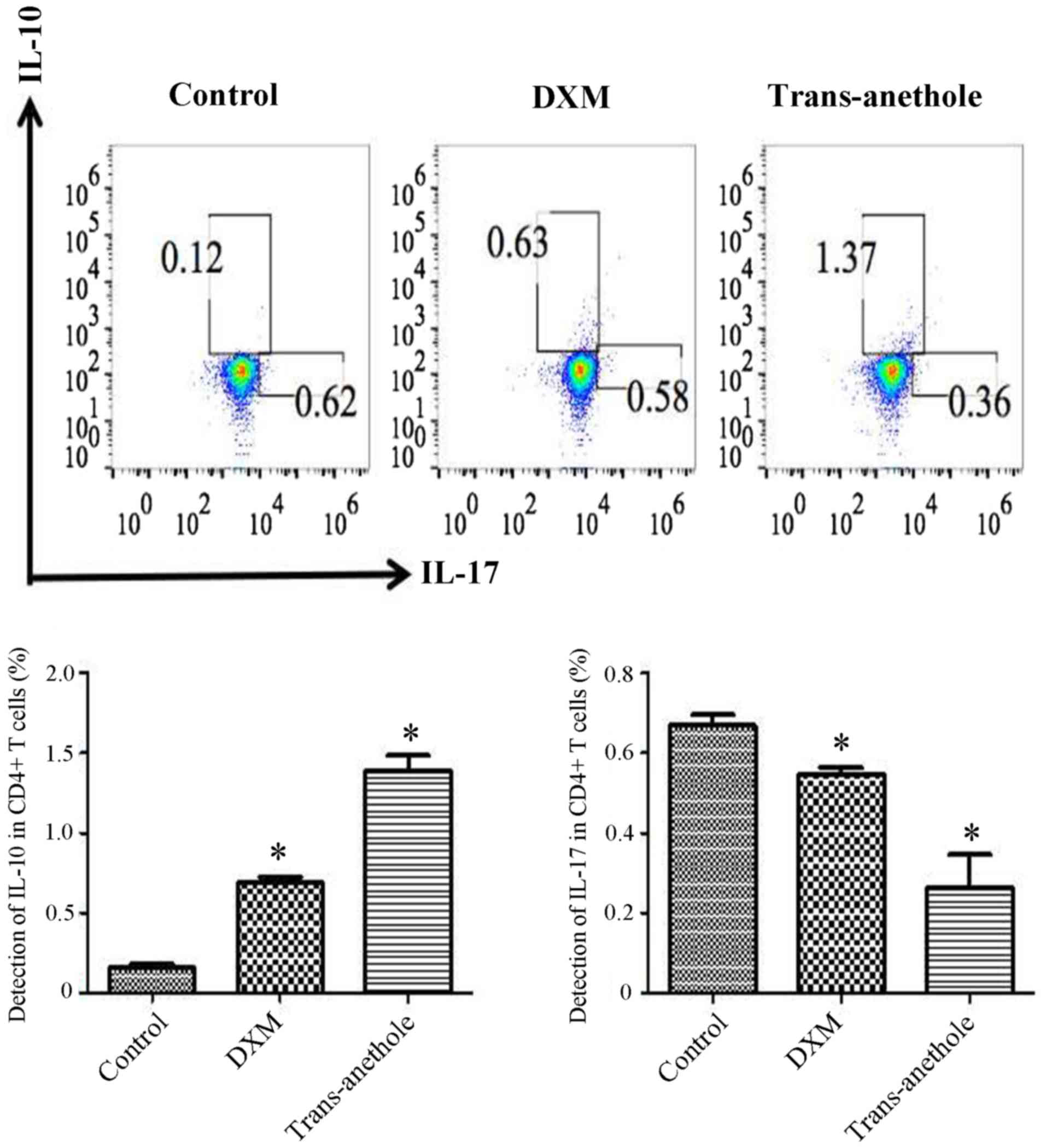
Given the important role of IL-10 and IL-17 for Th17
differentiation, we aimed to analyze the effect of trans-anethole
on these two cytokines. CD4+ T cells were collected from
the spleen of mice in the absence or presence of trans-anethole. We
observed that the compound selectively inhibited IL-17 production
without affecting the secretion of IL-10 (Fig. 6).
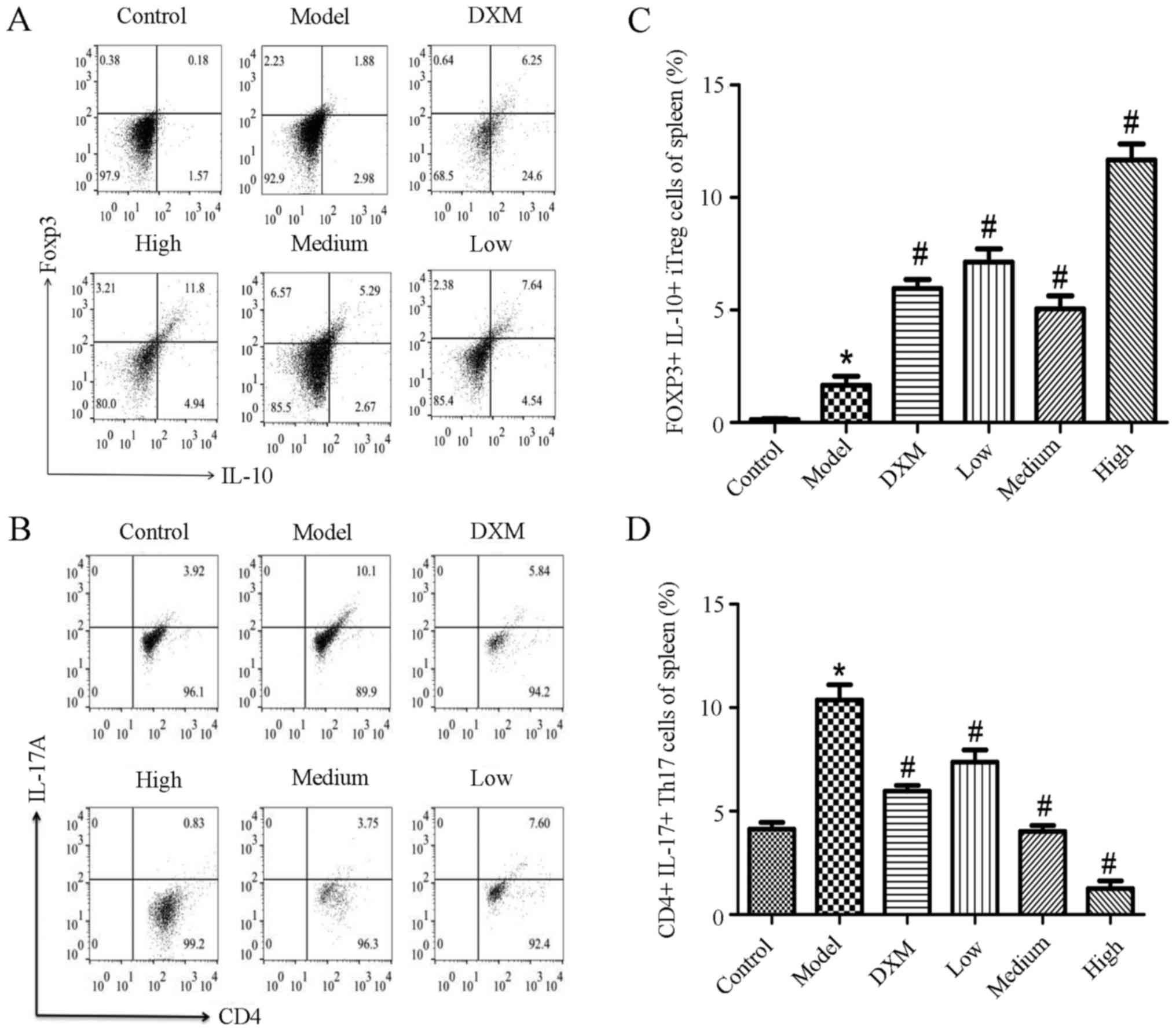
To assess the proportion of Tregs and Th17 cells
that were involved in lung inflammation, changes in the ratio of
the two groups of cells in the mice spleens was measured using flow
cytometry. As shown in Fig. 7, an
obvious increase in Th17 cells was demonstrated in the model group
compared with the normal group in the spleen. Trans-anethole
treatment caused a significant decrease of the proportion of Th17
cells and a significant increase in the percentage of iTreg cells
compared with the LPS-induced ALI mice. Compared with the
dexamethasone-treatment group, the changes to the ratio of Th17 and
Treg cells were more obvious in the spleen in the trans-anethole
group.
Discussion
LPS is a stimulator of pro-inflammatory cells,
therefore intratracheal administration of LPS was used to establish
the ALI mouse model. Histological examination of the lung tissue
showed thinning alveolar walls, diffuse hemorrhage, interstitial
tissue inflammation and decrease of the size of the alveolar
cavity. Our results also showed that inflammatory cells
(eosinophils and monocytes) were reduced in the BALF after
treatment.
ALI is a common disease. The early stages of
inflammatory diseases start with increasing levels of inflammatory
cells and mediators The CD4+Foxp3+ T cell
Treg subdivision is important for influencing two parts of the
immune process (1): To subdue an
extensive range of effector immune cells, such as B cells and
macrophages, and (2) to produce
relevant cytokines, such as IL-10 (38,39).
In our study, we reported a decrease of Th17 cells and an increase
of Tregs in the spleen tissue of the bronchitis model induced by
LPS. The above results demonstrate that the change in ratio of
Th17/Tregs may influence the pathogenesis of ALI caused by LPS.
Tregs are a subdivision of CD4+ T cells that are
important in maintaining self-tolerance to prevent autoimmunity,
control chronic inflammatory diseases and to regulate lymphocytes
(40). During the investigation of
lymphocyte function in ALI caused by LPS, the study demonstrated
that decreasing numbers of Tregs in mice with ALI decreased their
capacity for recovery. This can be explained by a deficiency in the
number of Tregs changing into lymphocytes that caused elevation of
pro-inflammatory cytokines in the alveolar cavity and enhanced
neutrophil apoptosis. This was confirmed in the present study as
after treatment, Treg cells were obviously increased in the spleen
tissue. Furthermore our study results are consistent with a recent
study, which demonstrated that decreasing levels of Tregs and IL-10
are widely observed in mice with ALI (41).
Our results demonstrated that gene expression of
IL-10 and IL-17 was changed in spleen and lung tissue after drug
treatment, which was measured using RT-qPCR. Fennel can resulte in
a notable reduction in the expression of IL-17 mRNA and a evident
increasement in the expression of IL-10 mRNA. Then, we examined the
change in Treg cell proportion in mice spleens, which was analyzed
by flow cytometry. The trans-anethole group showed a significantly
increased level of Tregs compared with the dexamethasone group. Our
preliminary results suggest that trans-anethole could increase the
percentage of Treg cells. We also found that Th17 cells were
decreased in the spleen tissue of mice with ALI. The present
results show that trans-anethole has anti-inflammatory activity,
which is associated with the regulation of Th17/Treg responses.
Fennel is one of the commonly used herbs in Chinese
traditional medicine, and has been demonstrated by studies in
modern pharmacology to have potential as a treatment of
inflammatory diseases. In the current study, we demonstrated that
trans-anethole, the main component of fennel, effectively
alleviated airway inflammatory in ALI mice. The airway inflammation
was alleviated via regulatory responses by Th17 and Tregs. The
anti-inflammatory function of fennel has not been fully explained
by molecular biology but the anti-inflammatory function of
trans-anethole may be related to the reaction of Th17 and Tregs.
Therefore, fennel may have potential as an effective treatment
against inflammatory conditions.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Projects from
Administration Bureau of Traditional Chinese Medicine and Ethnic
Group Medicine of Xinjiang Uyghur Autonomous Region (grant nos.
Q2015-01-10 and 2017-01-10), Key Research and Development Program
of Shandong Province (grant no. 2016ZDJS07A21) and the Program for
Changjiang Scholars and Innovative Research Team in University
(grant no. IRT_14R41).
Availability of data and materials
The datasets used or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SZ implemented the hematoxylin and eosin staining,
analyzed the data and wrote the manuscript. CH and LA executed the
Th17/Treg examination in spleen tissue. QY and ID conducted reverse
transcription-quantitative polymerase chain reaction. XC and SD
counted the number of inflammatory cells. XH conceived the
experiments and obtained the funding for this project. All authors
read and agreed to the final manuscript.
Ethics approval and consent to
participate
The present study was approved by the Animal Ethics
Committee Of Laboratory Animal Care of Tianjin University of
Traditional Chinese Medicine and was performed according to the
guidelines of the Tianjin University of Traditional Chinese
Medicine.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Badgujar SB, Patel VV and Bandivdekar AH:
Foeniculum vulgare Mill: A review of its botany, phytochemistry,
pharmacology, contemporary application, and toxicology. Biomed Res
Int. 2014:8426742014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garg C, Khan SA, Ansari SH, Suman A and
Garg M: Chemical composition, therapeutic potential and
perspectives of Foeniculum vulgare. Pharmacogn Rev. 3:346–352.
2009.
|
|
3
|
Muckensturm B, Foechterlen D, Reduron JP,
Danton P and Hildenbrand M: Phytochemical and chemotaxonomic
studies of Foeniculum vulgare. Biochem Syst Ecol. 25:353–358. 1997.
View Article : Google Scholar
|
|
4
|
Ostad SN, Soodi M, Shariffzadeh M,
Khorshidi N and Marzban H: The effect of fennel essential oil on
uterine contraction as a model for dysmenorrhea, pharmacology and
toxicology study. J Ethnopharmacol. 76:299–304. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Malini T, Vanithakumari G, Megala N,
Anusya S, Devi K and Elango V: Effect of Foeniculum vulgare Mill.
seed extract on the genital organs of male and female rats. Indian
J Physiol Pharmacol. 29:21–26. 1985.PubMed/NCBI
|
|
6
|
Ghodsi Z and Asltoghiri M: The effect of
fennel on pain quality, symptoms and menstrual duration in primary
dysmenorrhea. J Pediatr Adol Gynec. 27:283–286. 2014. View Article : Google Scholar
|
|
7
|
Piras A, Falconieri D, Porcedda S,
Marongiu B, Gonçalves MJ, Cavaleiro C and Salgueiro L:
Supercritical CO2 extraction of volatile oils from Sardinian
Foeniculum vulgare ssp. vulgare (Apiaceae): Chemical composition
and biological activity. Nat Prod Res. 28:1819–1825. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Damjanović B, Lepojević Ž, Živković V and
Tolić A: Extraction of fennel (Foeniculum vulgare Mill.) seeds with
supercritical CO2: Comparison with hydrodistillation.
Food Chem. 92:143–149. 2005. View Article : Google Scholar
|
|
9
|
Fang L, Qi M, Li T, Shao Q and Fu R:
Headspace solvent microextraction-gas chromatography-mass
spectrometry for the analysis of volatile compounds from Foeniculum
vulgare Mill. J Pharm Biomed Anal. 41:791–797. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pacifico S, Galasso S, Piccolella S,
Kretschmer N, Pan SP, Nocera P, Lettieri A, Bauer R and Monaco P:
Winter wild fennel leaves as a source of anti-inflammatory and
antioxidant polyphenols. Arab J Chem. (In press).
|
|
11
|
Kaur GJ and Arora DS: Antibacterial and
phytochemical screening of Anethum graveolens, Foeniculum vulgare
and Trachyspermum ammi. BMC Complement Altern Med. 9:302009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kang P, Kim KY, Lee HS, Min SS and Seol
GH: Anti-inflammatory effects of anethole in
lipopolysaccharide-induced acute lung injury in mice. Life Sci.
93:955–961. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jahromi Namavar B, Tartifizadeh A and
Khabnadideh S: Comparison of fennel and mefenamic acid for the
treatment of primary dysmenorrhea. Int J Gynaecol Obstet.
80:153–157. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Elsoud N, Ellaithy N, Elsaeed G, Wahby M,
Khalil M, Morsy F and Shaffie N: Antidiabetic activities of
Foeniculum vulgare Mill. essential oil in streptozotocin-induced
diabetic rats. Maced J Med Sci. 4:139–146. 2011.
|
|
15
|
Koppula S and Kumar H: Foeniculum vulgare
Mill (Umbelliferae) attenuates stress and improves memory in wister
rats. Trop J Pharm Res. 12:553–558. 2013.
|
|
16
|
Ferguson ND, Fan E, Camporota L, Antonelli
M, Anzueto A, Beale R, Brochard L, Brower R, Esteban A, Gattinoni
L, et al: The Berlin definition of ARDS: An expanded rationale,
justification, and supplementary material. Intensive Care Med.
38:1573–1582. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Matthay MA, Ware LB and Zimmerman GA: The
acute respiratory distress syndrome. J Clin Invest. 122:2731–2740.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mizgerd JP: Respiratory infection and the
impact of pulmonary immunity on lung health and disease. Am J
Respir Crit Care Med. 186:824–829. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bhattacharya J and Matthay MA: Regulation
and repair of the alveolar-capillary barrier in acute lung injury.
Ann Rev Physiol. 75:593–615. 2013. View Article : Google Scholar
|
|
20
|
Kim MR, Hong SW, Choi EB, Lee WH, Kim YS,
Jeon SG, Jang MH, Gho YS and Kim YK: Staphylococcus aureus-derived
extracellular vesicles induce neutrophilic pulmonary inflammation
via both Th1 and Th17 cell responses. Allergy. 67:1271–1281. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Iwakura Y, Ishigame H, Saijo S and Nakae
S: Functional specialization of interleukin-17 family members.
Immunity. 34:149–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Risso K, Kumar G, Ticchioni M, Sanfiorenzo
C, Dellamonica J, Guillouet-de Salvador F, Bernardin G, Marquette
CH and Roger PM: Early infectious acute respiratory distress
syndrome is characterized by activation and proliferation of
alveolar T-cells. Eur J Clin Microbiol Infect Dis. 34:1111–1118.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Noack M and Miossec P: Th17 and regulatory
T cell balance in autoimmune and inflammatory diseases. Autoimmun
Rev. 13:668–677. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Costantino CM, Baecher-Allan CM and Hafler
DA: Human regulatory T cells and autoimmunity. Eur J Immunol.
38:921–924. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Guo Z, Wen Z, Qin A, Zhou Y, Liao Z, Liu
Z, Liang Y, Ren T and Xu L: Antisense oligonucleotide treatment
enhances the recovery of acute lung injury through IL-10-secreting
M2-like macrophage-induced expansion of CD4+ regulatory T cells. J
Immunol. 190:4337–4348. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mills KH: Regulatory T cells: Friend or
foe in immunity to infection? Nat Rev Immunol. 4:841–855. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Miossec P, Korn T and Kuchroo VK:
Interleukin-17 and type 17 helper T cells. New Engl J Med.
361:888–898. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nistala K, Moncrieffe H, Newton KR,
Varsani H, Hunter P and Wedderburn LR: Interleukin-17-producing T
cells are enriched in the joints of children with arthritis, but
have a reciprocal relationship to regulatory T cell numbers.
Arthritis Rheum. 58:875–887. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tzartos JS, Friese MA, Craner MJ, Palace
J, Newcombe J, Esiri MM and Fugger L: Interleukin-17 production in
central nervous system-infiltrating T cells and glial cells is
associated with active disease in multiple sclerosis. Am J Pathol.
172:146–155. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hirota K, Yoshitomi H, Hashimoto M, Maeda
S, Teradaira S, Sugimoto N, Yamaguchi T, Nomura T, Ito H, Nakamura
T, et al: Preferential recruitment of CCR6-expressing Th17 cells to
inflamed joints via CCL20 in rheumatoid arthritis and its animal
model. J Exp Med. 204:2803–2812. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Halwani R, Al-Muhsen S and Hamid Q: T
helper 17 cells in airway diseases: From laboratory bench to
bedside. Chest. 143:494–501. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Hartigan-O'Connor DJ, Hirao LA, McCune JM
and Dandekar S: Th17 cells and regulatory T cells in elite control
over HIV and SIV. Curr Opin HIV AIDS. 6:221–227. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
You QH, Zhang D, Niu CC, Zhu ZM, Wang N,
Yue Y and Sun GY: Expression of IL-17A and IL-17F in
lipopolysaccharide-induced acute lung injury and the counteraction
of anisodamine or methylprednisolone. Cytokine. 66:78–86. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li C, Yang P, Sun Y, Li T, Wang C, Wang Z,
Zou Z, Yan Y, Wang W, Wang C, et al: IL-17 response mediates acute
lung injury induced by the 2009 pandemic influenza A (H1N1) virus.
Cell Res. 22:528–538. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fedele G, Spensieri F, Palazzo R, Nasso M,
Cheung GY, Coote JG and Ausiello CM: Bordetella pertussis commits
human dendritic cells to promote a Th1/Th17 response through the
activity of adenylate cyclase toxin and MAPK-pathways. PLoS One.
5:e87342010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu J, Zhang PS, Yu Q, Liu L, Yang Y, Guo
FM and Qiu HB: Losartan inhibits conventional dendritic cell
maturation and Th1 and Th17 polarization responses: Νovel
mechanisms of preventive effects on lipopolysaccharide-induced
acute lung injury. Int J Mol Med. 29:269–276. 2012.PubMed/NCBI
|
|
37
|
Choi EM and Hwang JK: Antiinflammatory,
analgesic and antioxidant activities of the fruit of Foeniculum
vulgare. Fitoterapia. 75:557–565. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Taylor A, Verhagen J, Blaser K, Akdis M
and Akdis CA: Mechanisms of immune suppression by interleukin-10
and transforming growth factor-beta: The role of T regulatory
cells. Immunology. 117:433–442. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
De M, De AK, Sen P and Banerjee AB:
Antimicrobial properties of star anise (Illicium verum Hook f).
Phytother Res. 16:94–95. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang H, Peng W, Weng Y, Ying H, Li H, Xia
D and Yu W: Imbalance of Th17/Treg cells in mice with chronic
cigarette smoke exposure. Int Immunopharmacol. 14:504–512. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Scheffold A, Murphy KM and Höfer T:
Competition for cytokines: T(reg) cells take all. Nat Immunol.
8:1285–1287. 2007. View Article : Google Scholar : PubMed/NCBI
|