Introduction
Depression has become one of the most disabling
psychiatric conditions (1,2). It is a multifactorial disorder that
may involve changes in gene expression and regulation. Previous
studies have focused on the role of genetic factors in the
development of depression, and several genetically altered loci
have been identified (3,4). However, the pathogenesis of
depression remains unclear.
Salmena et al (5) proposed a novel RNA regulation
mechanism involving competitive endogenous RNA (ceRNA) regulation,
whereby RNA transcripts crosstalk with one another via decreasing
targeting concentrations of microRNAs (miRNAs), which suppresses
other RNAs that share common miRNA response elements (5). Currently, several studies have
demonstrated that ceRNA regulation serves critical roles in human
disease, including cancer and rheumatoid arthritis (6–8). For
example, phosphatase and tensin homolog (PTEN), which is a tumor
suppressor gene, may be regulated by ceRNA activity in multiple
tumors (9,10). RNA-Sequencing (Seq) technology has
allowed for the identification of long non-coding RNAs (lncRNAs);
lncRNAs may act as ceRNAs in human diseases. For example, high
mobility group AT-hook 1 (HMGA1) pseudogenes may act as ceRNA
decoys. H19 affects HMGA1 expression by attenuating the suppression
of let-7, thereby promoting pancreatic cancer metastasis (11). Small nucleolar RNA host gene 6-003
may act as a ceRNA to promote the progression of hepatocellular
carcinoma (12). Zhou et al
(13) constructed a breast
cancer-specific ceRNA network based on the mRNA expression profile.
Furthermore, the integrative analyses of Du et al (14) uncovered a lncRNA-mediated sponge
regulatory network in prostate cancer. Previous studies have
focused on investigating the ceRNA regulation in cancer. Jiang
et al (15) revealed
functional lncRNAs in rheumatoid arthritis based on the ceRNA
theory. Lai et al (16)
demonstrated that HOX transcript antisense RNA acts as a ceRNA and
may regulate PTEN expression by inhibiting miR-19 in cardiac
hypertrophy. Recently, a previous study revealed that have revealed
that noncoding RNAs serve important roles in depression (17). However, a systematical dissection
of the ceRNA network in depression has not been performed.
In the present study, a ceRNA network was
constructed using samples obtained from non-psychiatric individuals
and patients with major depression. Further analysis identified
critical ceRNA modules in depression. Such systematic construction
and dissection of the ceRNA regulatory network in depression may
aid to elucidate the underlying pathogenic mechanisms involved in
depression.
Materials and methods
RNA-seq expression data
RNA-Seq data were downloaded from the Gene
Expression Omnibus (GEO) database (18). The accession number was GSE42546
(2), which refers to a study on
the transcriptome profiling of human hippocampus dentate gyrus
granule cells in mental illness. The expression profile contains 79
samples of patients with mental illness including 17 patients with
schizophrenia, 16 patients with bipolar disorder, 17 patients with
major depression and 29 non-psychiatric controls. The focus of the
present study was on depression, therefore the data on 17 patients
with major depression and 29 non-psychiatric control samples were
used for further analysis. RNAs (mRNAs and lncRNAs) involved the
expression profile were unified as genes.
Construction of the ceRNA regulation
network
The ceRNA network was constructed based on the
gene-miRNA interactions and the RNA expression data. Firstly, the
lncRNA-miRNA and gene-miRNA interaction data were downloaded from
starBase V2.0 database (starbase.sysu.edu.cn/) (19). A total of 10,212 interactions from
1,065 lncRNA genes and 277 miRNAs, and 606,408 interactions from
13,801 mRNA genes and 386 miRNAs were obtained. These lncRNA-miRNA
and gene-miRNA interactions were combined and 616,620 interactions
between 386 miRNAs and 14,816 genes were obtained. Candidate ceRNA
pairs were constructed by screening all possible RNA-RNA pairs. RNA
pairs that had at least 3 common miRNAs and a Jacquard coefficient
of shared miRNAs between two RNAs of >0.2 were identified as
candidate ceRNA pairs. In addition, the ceRNA regulatory pair was
identified based on the expression data obtained from 29
non-psychiatric controls and 17 patients with major depression. To
ensure the volatility of expression data and to obtain reliable
associations, genes with zero expression values were removed in
>50% samples. Finally, a total of 14,443 genes were retained in
the expression profile. At least 9 samples could lead to
statistically significant results. A total of 2 ceRNAs were
required to exhibit positively correlated expression using a
Pearson correlation coefficient of >0.4 and an adjusted
significance P-value of <0.05. Correlation analysis was
evaluated using the Fisher's method in the Weighted Correlation
Network Analysis package (horvath.genetics.ucla.edu/html/CoexpressionNetwork/Rpackages/WGCNA/)
(20). The identified ceRNA pairs
were combined and the ceRNA regulatory networks were then
constructed.
Differential expression analysis
To ensure the accuracy of differential gene
recognition, a t-test was conducted and fold-change values were
used (t-test, P-value <0.05; fold-change values >1.5 or
<0.667). The t-test was conducted and the fold-change value for
each gene was calculated using R 2.15.3 (cran.r-project.org). P<0.05 was considered to
indicate a statistically significant difference.
Functional enrichment analysis
The enrichment analysis was performed to investigate
the function of genes specific for the ceRNA networks in the normal
and depression states. In this process, the Database for
Annotation, Visualization and Integrated Discovery (DAVID v6.8)
tool (david.ncifcrf.gov) (21) was used; briefly, genes specific for
the ceRNA networks in the normal and depression states were
inputted as gene lists. From the results page, the Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathways (www.genome.jp/kegg/) (22) chart was downloaded for further
analysis.
Identifying ceRNA modules
The MCODE v1.4.2 plugin in Cytoscape v3.4.0
(cytoscape.org) (23) was used to examine the closely
associated ceRNA modules for normal- and depression-associated
networks. The default selection for MCODE was used.
Identifying key modules
Modules that were significantly enriched with
differentially expressed genes were identified as key modules using
the Gene Set Enrichment Analysis (GSEA) method (24) and hypergeometric test. The GSEA
method was performed using the GSEA 1.0 R script (24).
Screening of drug-like small
molecules
The CMAP (build 02) database (www.cmap-software.com) (25) collects genome-wide transcriptional
expression data from cultured human cells treated with bioactive
small molecules. A total of >6,000 gene expression profiles
representing 1,309 compounds were downloaded. For each compound,
the treatment group was compared with the control group in order to
identify genes associated with the compound intervention under
different thresholds (fold-change values >1.5 or fold-change
values <0.667 fold-change values >2 or fold-change values
<0.5). Genes with fold-change values of >1.5 or <0.667
were considered to be genes associated with the compound
intervention according to the fold-change method. In addition, a
more stringent threshold (fold-change values of >2 or <0.5)
was also used to explore the reproducibility of the detection
results.
Results and Discussion
Properties of the ceRNA network in
depression
The ceRNA regulatory network was constructed using
data obtained from the GEO database from 17 patients with major
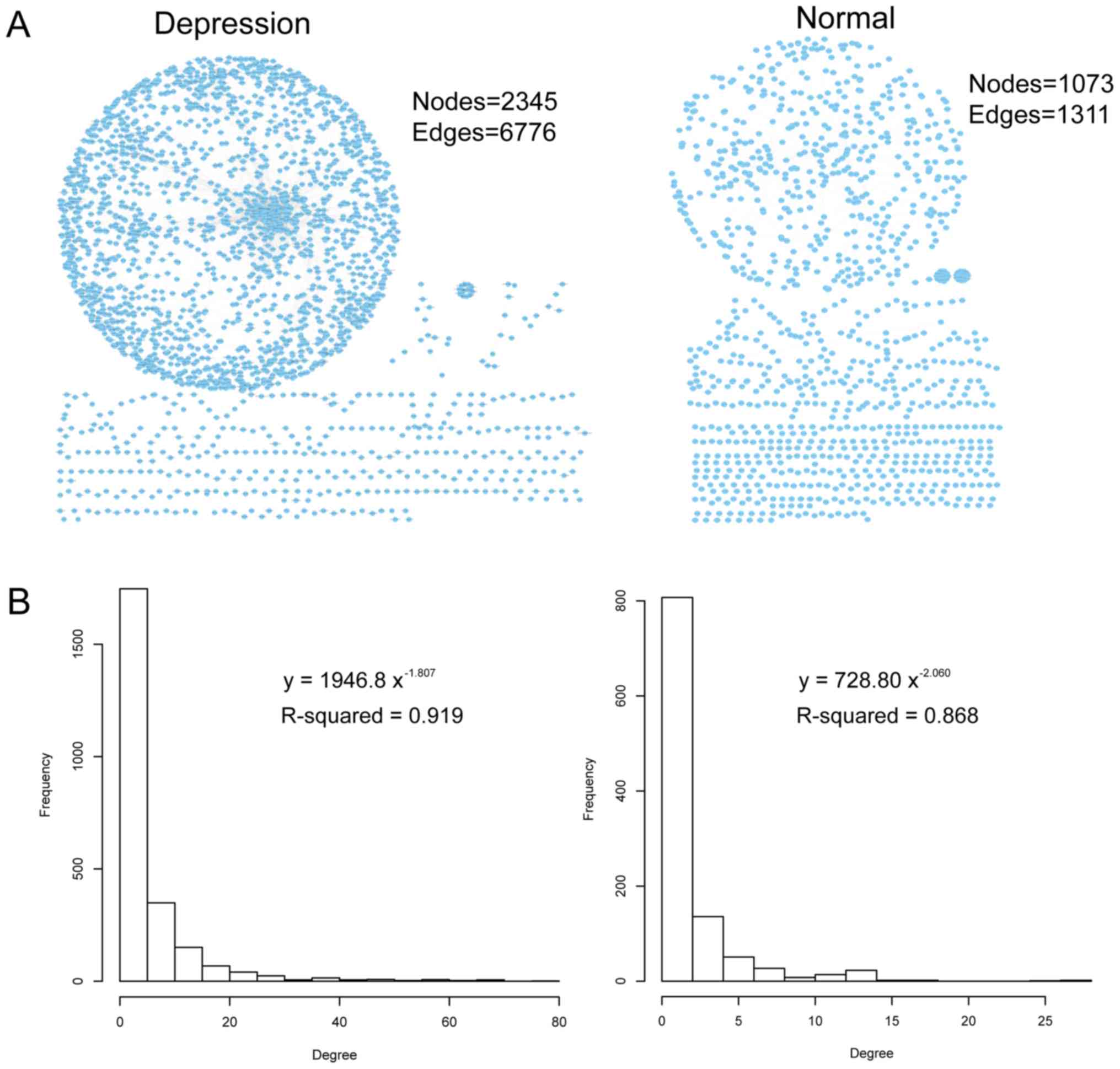
depression and 29 non-psychiatric control samples. A total of 1,311
ceRNA associations between 1,073 genes, and 6,776 ceRNA
associations between 2,345 genes were identified under normal
control and disease states, respectively (Fig. 1A). Then, the degree distribution of
the ceRNA networks was examined (with degree defined as the number
of edges connected to a certain node; Fig. 1B). The majority of ceRNAs exhibited
a low connectivity, whereas a few ceRNAs exhibited a relatively
high connectivity. Further examination of the degree distribution
of the ceRNA networks demonstrated that the degree of these two
ceRNA networks was subjected to a power-law distribution with a
slope of −2.06 and R2=0.868 for the ceRNA control
network, and a slope of −1.807 and R2=0.919 for the
ceRNA network of depression (Fig.
1B). This suggests that ceRNA networks in depression are
similar to the majority of biological networks. However there are a
number of core nodes that maintain the ceRNA network organizational
structure and distinguish it from random networks.
Next, the core nodes of the ceRNA networks were
examined. Firstly, the hub nodes (defined as the nodes with a
degree of >5 and as the top 5%) for each ceRNA network were
identified. A total of 53 hub nodes in the ceRNA control network
and 121 hub nodes in the disease-associated ceRNA network were
identified. Table I presents the
top 30 hub nodes with regard to degree number in the normal and
disease networks. Further exploration of these hub nodes
demonstrated that a number of these have been reported to be
associated with depression. For example, the amount of
mitochondrial DNA (mtDNA) has been implicated in stress-associated
diseases, including major depressive disorder (26). Cyclin-dependent kinase (CDK)-6, a
hub gene in the normal ceRNA network, has been identified as a
novel molecule involved in the control of mtDNA (26). High mobility group nucleosomal
binding domain 3 (HMGN3), peroxisome proliferator-activated
receptor-γ coactivator 1β (PPARGC1B) and leukemia inhibitory factor
receptor-α (LIFR) were revealed to be hub ceRNAs in the
depression-associated ceRNA network in this study. The Hmgn3 gene
exhibits differential expression between anxiety-associated
behavior and control mouse models (27). Kripke et al (28) suggested that PPARGC1B polymorphisms
may be associated with bipolar disorder. Furthermore, Kripke et
al also weakly replicated the association between depression
and PPARGC1B rs7732671 in another study (29). Additionally, previous research has
indicated that plasma tryptophan levels are decreased during
pregnancy and puerperium, as well as in patients with major
depression and inflammation (30).
According to a study by Maes et al (30) LIFR is significantly associated with
the changes in the tryptophan/competing amino acid ratio.
Therefore, these hub ceRNAs, including HMGN3, PPARGC1B and LIFR,
are likely associated with depression, and their regulation may
serve important roles in the initiation and progression of
depression.
 | Table I.Top 30 hub nodes in the normal and
depression competitive endogenous RNA networks. |
Table I.
Top 30 hub nodes in the normal and
depression competitive endogenous RNA networks.
| A, Normal
network |
|---|
|
|---|
| Node name | Degree |
|---|
| ATMIN | 27 |
| RBM12B | 27 |
| RAB11FIP2 | 25 |
| FOXJ2 | 17 |
| LOC374443 | 17 |
| KLF3 | 16 |
| ARMC8 | 15 |
| C2orf69 | 14 |
| PCDHGB7 | 13 |
| PCDHGA5 | 13 |
| PCDHGA12 | 13 |
| PCDHGB4 | 13 |
| PCDHGB6 | 13 |
| PCDHGA10 | 13 |
| PCDHGA9 | 13 |
| PCDHGB5 | 13 |
| PCDHGA8 | 13 |
| PCDHGB2 | 13 |
| PCDHGA6 | 13 |
| PCDHGB3 | 13 |
| PCDHGA7 | 13 |
| PCDHGA11 | 13 |
| PCDHA7 | 13 |
| PCDHA6 | 13 |
| PCDHA5 | 13 |
| PCDHA8 | 13 |
| TTC32 | 13 |
| C3orf58 | 13 |
| PNRC1 | 13 |
| SATB1 | 13 |
|
| B, Depression
network |
|
| Node
name | Degree |
|
| GOLGA7 | 78 |
| MED8 | 70 |
| C3orf23 | 70 |
| 8-Sep | 69 |
| MTF2 | 68 |
| CPOX | 66 |
| HMGN3 | 66 |
| FAM199X | 65 |
| CTPS | 64 |
| GPR180 | 62 |
| ADSS | 59 |
| TNKS2 | 57 |
| MSL1 | 57 |
| ZMYM1 | 57 |
| KLF3 | 57 |
| C18orf25 | 56 |
| TXNRD1 | 56 |
| C14orf28 | 55 |
| CAND1 | 55 |
| TNFAIP1 | 51 |
| COG3 | 50 |
| IL15 | 50 |
| CHD7 | 50 |
| ZNF236 | 49 |
| RAB14 | 49 |
| ZBTB41 | 48 |
| RBM24 | 47 |
| CLDND1 | 46 |
| TAOK3 | 45 |
| ZDHHC9 | 44 |
Comparison of the ceRNA regulation
network under normal physiological and disease states
In order to further understand ceRNA regulation in
depression, the ceRNA networks under normal physiological and
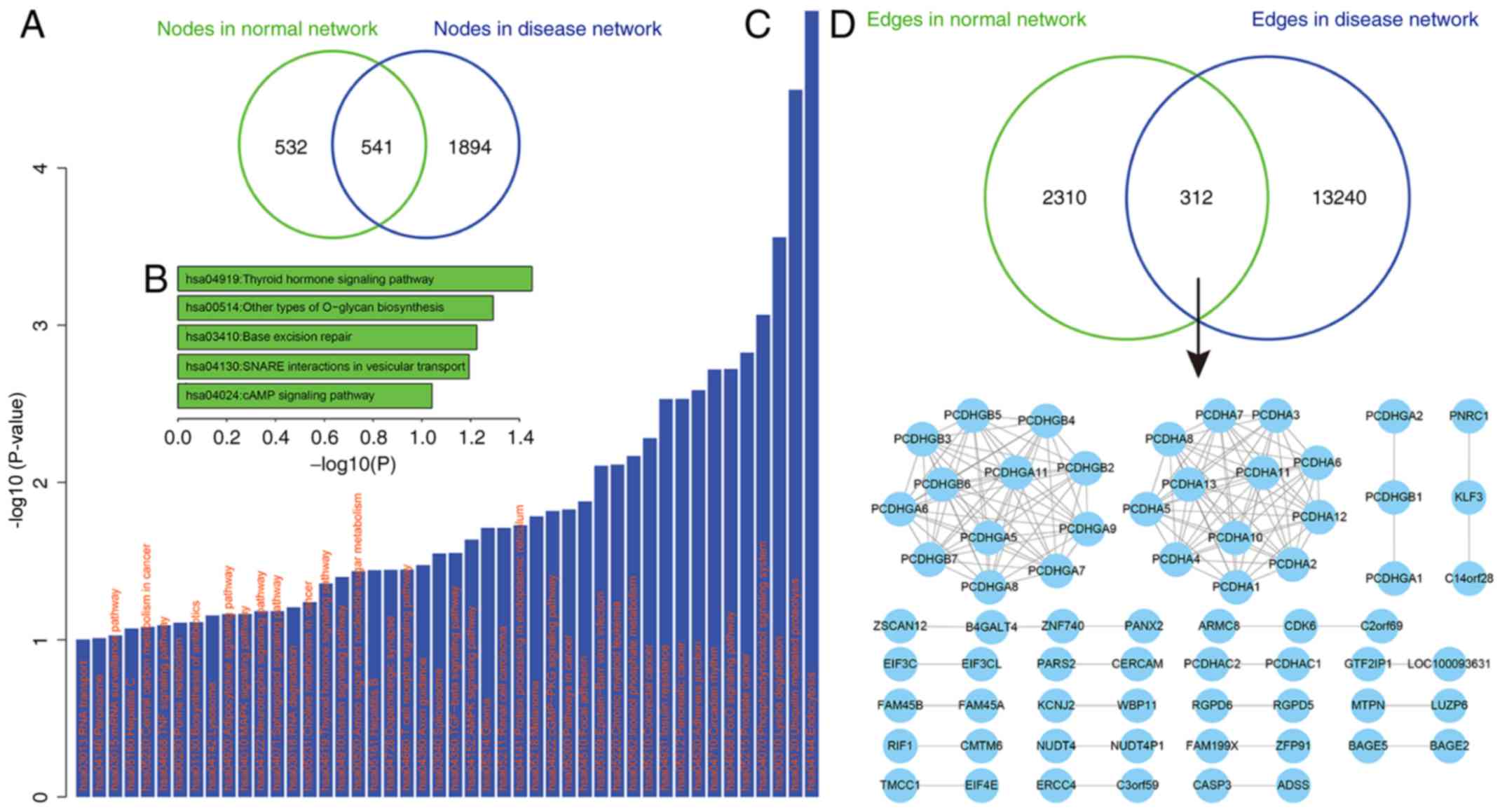
disease states were compared. Firstly, the nodes between the ceRNA
networks were compared, and it was revealed that half of the ceRNAs
(50.4%) in the normal ceRNA network were also involved in the
disease-associated ceRNA network, whereas 532 ceRNA nodes (49.6%)
and up to 1,894 ceRNAs (77.7%) were specifically involved in the
normal and disease networks, respectively (Fig. 2A). This suggests that the
initiation of depression may be associated with these ceRNA
activities. Next, the function of these specific ceRNAs in the
normal and depression states was investigated. Functional
enrichment analysis for these ceRNA sets (P<0.05) was performed
to identify significantly enriched KEGG pathways (22) using the DAVID tool (21). A total of 532 ceRNA nodes that were
specifically involved in the normal ceRNA network were
significantly enriched for the ‘thyroid hormone signaling pathway’
(P<0.05; Fig. 2B). Furthermore,
four pathways were enriched with a significant (P<0.01),
including ‘other types of O-glycan biosynthesis’ (P=0.051), ‘base
excision repair’ (P=0.059), ‘soluble N-ethylmaleimide-sensitive
fusion protein-attachment protein receptors interactions in
vesicular transport’ (P=0.064) and ‘cyclic adenosine monophosphate
signaling pathway’ (P=0.091). These 1,894 ceRNAs, which were
specific for the depression network, were enriched in up to 31
pathways (P<0.05; Fig. 2C).
These results indicate that these ceRNAs may regulate a wide
variety of biological functions. In addition, a number of these
pathways may be associated with depression, including ‘circadian
rhythm’ (31,32) and ‘insulin resistance’ (33).
According to the results of the present study, only
a small fraction of ceRNA pairs (11.9%) were shared between the
normal and disease networks. This indicates that the differential
regulation of ceRNAs may be implicated in depression. Firstly,
common ceRNA pairs were examined and it was revealed that these 312
shared ceRNA pairs of the ceRNA networks may form a core subnetwork
(Fig. 2D), which mainly contains
protocadherin (PCDH) family members. Chen et al (34) revealed that conditional deletion of
the mouse Pcdh-α (PCDHA) gene cluster was associated with
depression-like behaviors, indicating that these ceRNA regulations
may serve critical roles in depression. In the present study, ceRNA
regulation pairs that were specific for depression were assessed.
It was demonstrated that maternally expressed 8 (MEG8; a lncRNA)
competitively regulated four genes including integrin subunit
β-like 1 (ITGBL1), SSU72 homolog RNA polymerase II C-terminal
domain phosphatase, potassium voltage-gated channel modifier
subfamily G member 3 (KCNG3) and Acyl-coenzyme A synthetase short
chain family member 2 (ACSS2) in depression; MEG8 was not present
in the normal ceRNA network. Additionally, it was revealed that
miR-16 participates in three of these ceRNA regulations, namely
MEG8-ITGBL1, MEG8-KCNG3 and MEG8-ACSS2. miR-16 expression is
associated with major depression (35–37)
and has therefore been used as a serum biomarker for depression
(38). This indicates that the
lncRNA MEG8 may be a novel molecule implicated in the pathogenesis
of depression. However, further investigation is required.
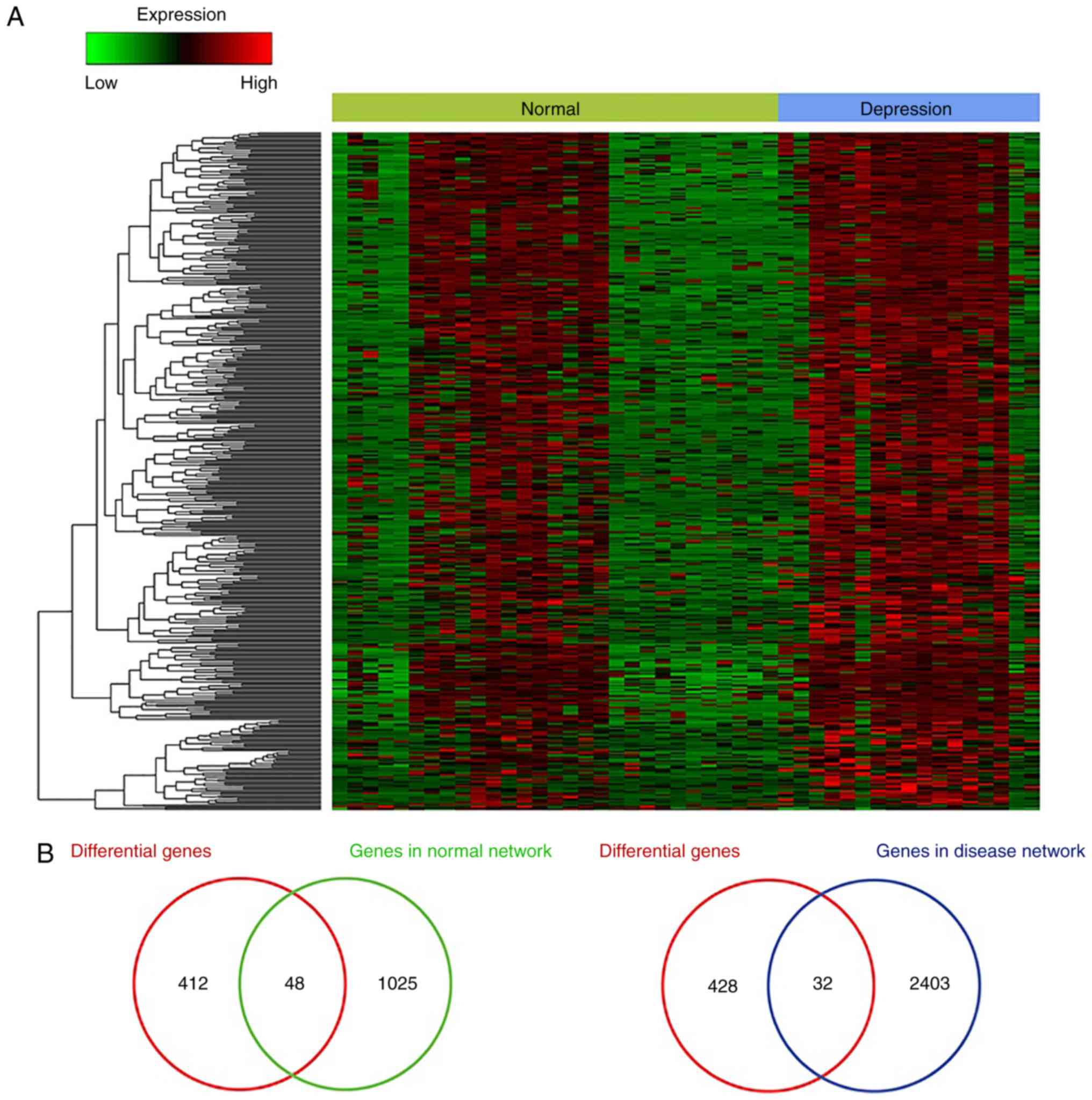
Expression analysis of ceRNAs
The expression of ceRNAs in the normal and
depression-associated networks was examined. Comparing samples from
non-psychiatric individuals and patients with major depression
identified a total of 460 genes (data not shown). Cluster analysis
of these genes was also performed (Fig. 3A). There were 48 and 32
differentially expressed ceRNAs in the normal and
depression-associated networks, respectively (Fig. 3B). The fold-change value was
>1.5 or <0.667 and the P-value was <0.05. A hypergeometric
test was conducted to identify overlapping ceRNAs and it was
revealed that the overlap between the differential genes and ceRNAs
in the normal networks was significant (P<0.05) (data not
shown). This suggests that these differentially expressed ceRNAs
may lead to ceRNA dysregulation and thus be involved in the
occurrence of depression.
Dissecting ceRNA modules in
depression
Compared with the global ceRNA regulatory networks,
subnetwork modules may provide key insights into ceRNA regulation.
In the present study, ceRNA regulation modules were examined. The
MCODE v1.4.2 plugin (23) in
Cytoscape v3.4.0 was used to identify ceRNA modules for normal and
depression-associated networks. The default selection for MCODE was
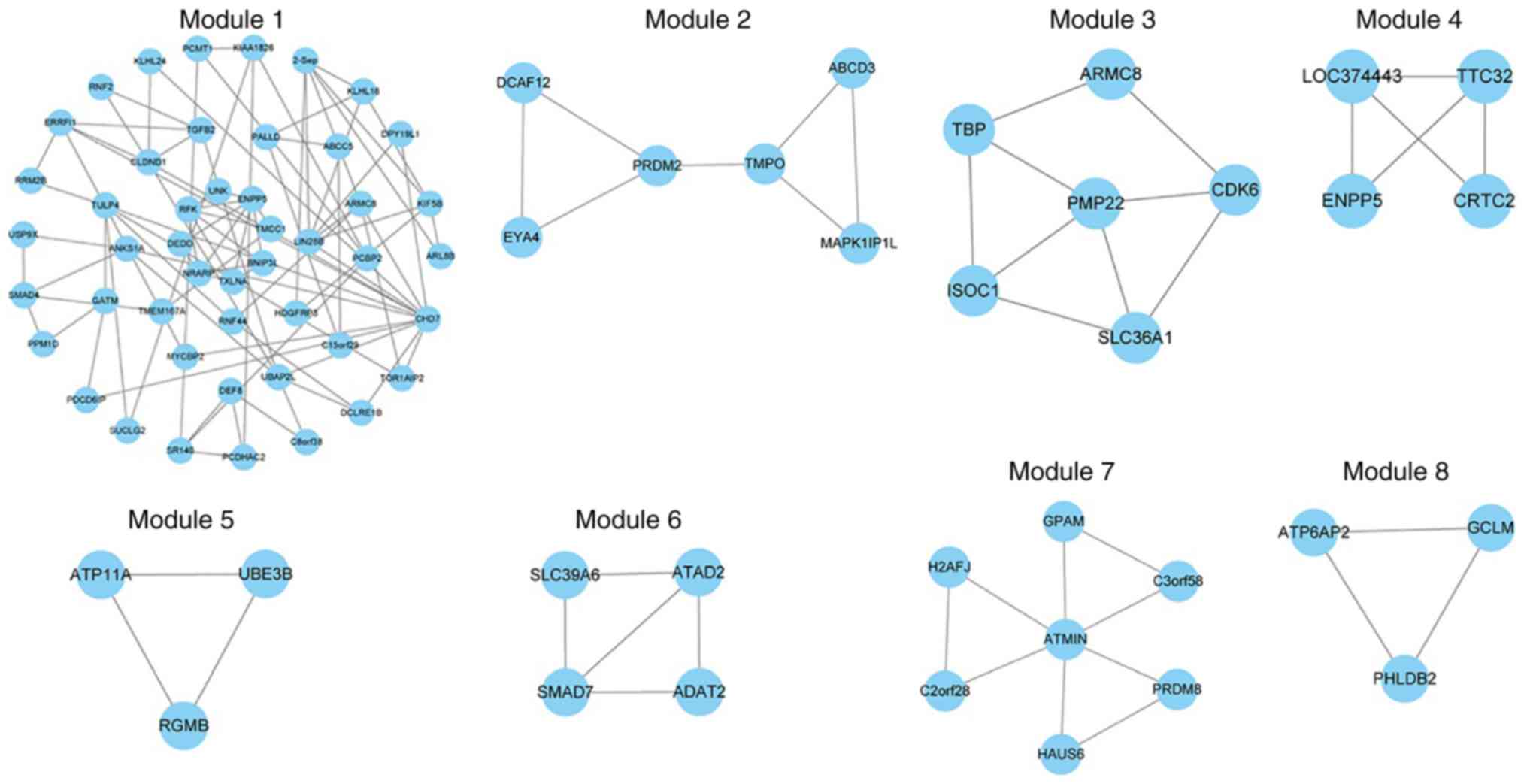
used. A total of 22 and 62 ceRNA modules were identified for the
normal and depression-associated networks, respectively. At least 3
ceRNAs (that were closely associated with each other in the
network) were involved in each module.
Next, key modules, defined as modules that were
significantly enriched with differentially expressed genes, were
examined. GSEA (24) and a
hypergeometric test were used to assess dysfunctional modules. A
total of 4 differential modules were identified using GSEA and 4
were identified via the hypergeometric test (P<0.05; Fig. 4). These modules were dysfunctional
in the disease state and several of these ceRNAs, including PCDHA
subfamily C2 (PCDHAC2; module 1) and CDK6 (module 3), may be
implicated in the occurrence and development of depression. For
example, mtDNA is implicated in stress-associated diseases,
including major depressive disorder (22). Cai et al (26) reported that CDK6 may regulate
mtDNA. Among these 8 key modules, module 1 contained the largest
number of ceRNAs and notable complex interactions. Functional
analysis of these modules revealed an association with the
following pathways: ‘Role of Tob in T-cell activation’,
‘transforming growth factor (TGF)-β signaling pathway’, ‘First
Multivalent Nuclear Factor’, ‘Cell Cycle (G1/S Check Point)’ and
‘anaplastic lymphoma kinase’ in cardiac myocytes. Some of these
functions have been reported to be associated with depression, such
as Mihailova et al who observed low TGF-β1 expression in
patients with depression (39).
This indicates that other mechanisms may be involved in the
pathophysiology of depression.
Potential relevance drug screening for
key ceRNA modules
Small molecules may affect the expression of genes
and improve disease prognosis. In the present study, small
molecules that may regulate dysfunctional key modules in depression
were screened. A total of 1,309 small molecules were analyzed and
screened using a hypergeometric test. The following thresholds were
used for the analysis: Fold-change values >1.5 or <0.667;
fold-change values >2 or <0.5. Significantly associated small
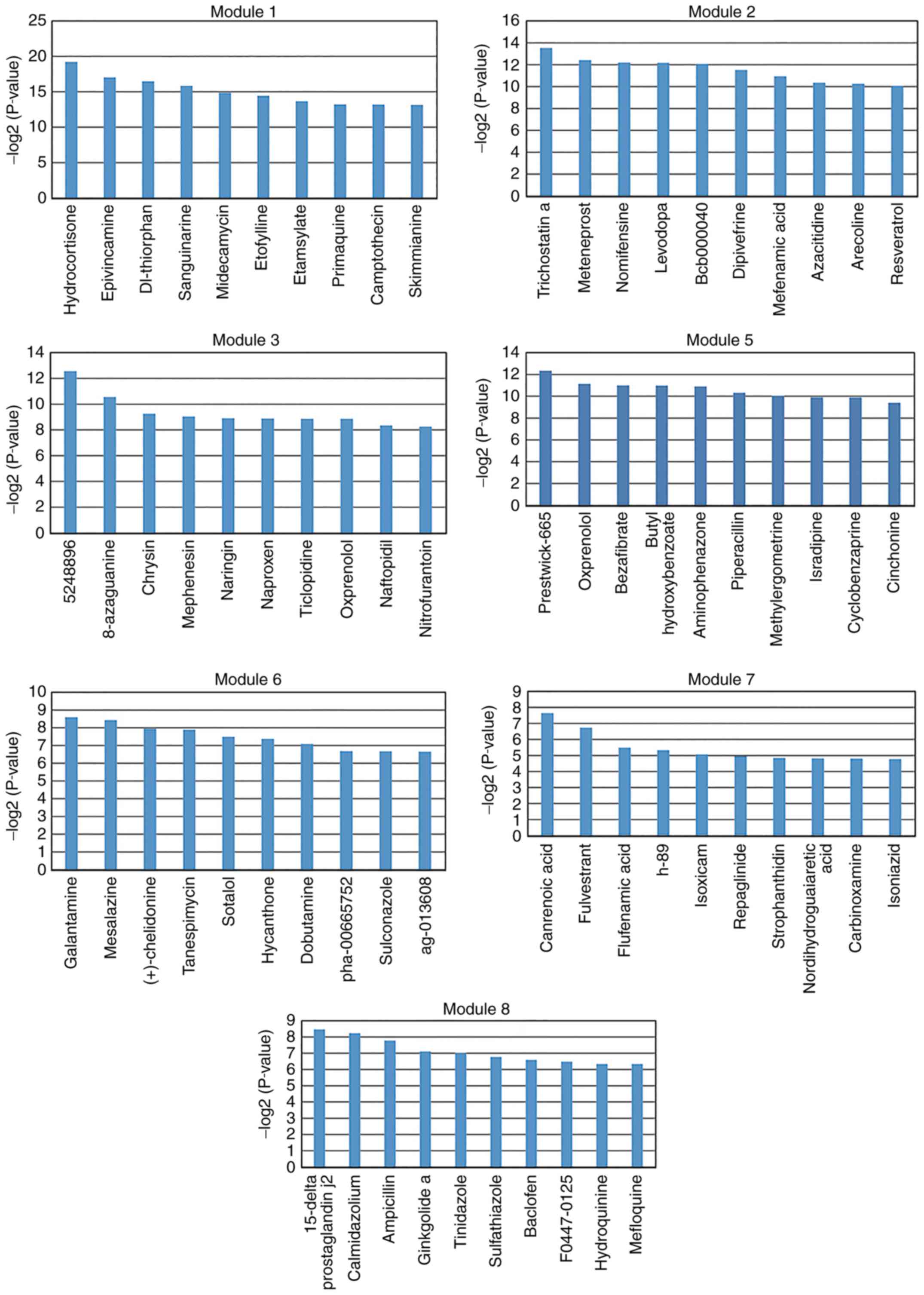
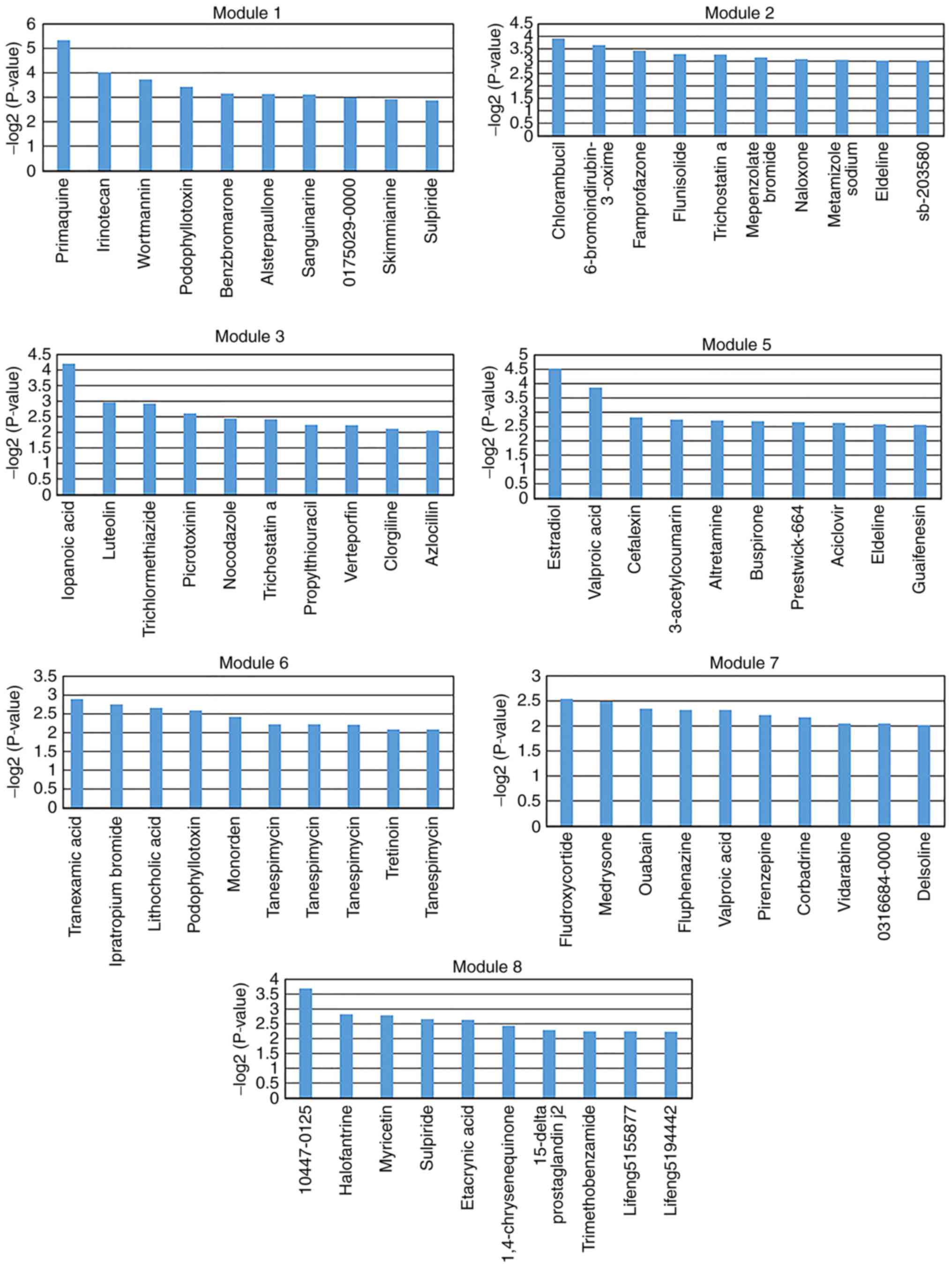
molecules (P<0.05) were identified for each key module. The top
10 significantly associated small molecules were selected for each
key module (except module 4; Figs.
5 and 6). There were no small
molecules significantly associated with module 4. These selected
small molecules may be associated with improved disease outcomes or
lead to side effects. For example, isradipine was screened for
module 5 (Fig. 5). A preliminary
study by Ostacher et al (40) suggested that isradipine may be
safely administrated to patients with bipolar depression. However,
further investigation is required.
In conclusion, in the present study a
depression-associated ceRNA network was constructed and a
systematical analysis of ceRNA regulation in depression was
performed. The results of the present study may provide novel
perspectives on understanding the pathogenesis and treatment
mechanisms of depression.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JZ and ZY conducted the analyses and interpreted the
data. YL was a major contributor in designing the study and writing
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sullivan PF, Neale MC and Kendler KS:
Genetic epidemiology of major depression: Review and meta-analysis.
Am J Psychiatry. 157:1552–1562. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kohen R, Dobra A, Tracy JH and Haugen E:
Transcriptome profiling of human hippocampus dentate gyrus granule
cells in mental illness. Transl Psychiatry. 4:e3662014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lacerda-Pinheiro SF, Pinheiro Junior RF,
Pereira de Lima MA, Lima da Silva CG, Vieira dos Santos Mdo S,
Teixeira Júnior AG, Lima de Oliveira PN, Ribeiro KD, Rolim-Neto ML
and Bianco BA: Are there depression and anxiety genetic markers and
mutations? A systematic review. J Affect Disord. 168:387–398. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Heiman GA, Kamberakis K, Gill R,
Kalachikov S, Pedley TA, Hauser WA and Ottman R: Evaluation of
depression risk in LGI1 mutation carriers. Epilepsia. 51:1685–1690.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Salmena L, Poliseno L, Tay Y, Kats L and
Pandolfi PP: A ceRNA hypothesis: The Rosetta Stone of a hidden RNA
language? Cell. 146:353–358. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ergun S and Oztuzcu S: Oncocers:
ceRNA-mediated cross-talk by sponging miRNAs in oncogenic pathways.
Tumour Biol. 36:3129–3136. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xia T, Liao Q, Jiang X, Shao Y, Xiao B, Xi
Y and Guo J: Long noncoding RNA associated-competing endogenous
RNAs in gastric cancer. Sci Rep. 4:60882014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sumazin P, Yang X, Chiu HS, Chung WJ, Iyer
A, Llobet-Navas D, Rajbhandari P, Bansal M, Guarnieri P, Silva J
and Califano A: An extensive microRNA-mediated network of RNA-RNA
interactions regulates established oncogenic pathways in
glioblastoma. Cell. 147:370–381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Karreth FA, Tay Y, Perna D, Ala U, Tan SM,
Rust AG, DeNicola G, Webster KA, Weiss D, Perez-Mancera PA, et al:
In vivo identification of tumor-suppressive PTEN ceRNAs in an
oncogenic BRAF-induced mouse model of melanoma. Cell. 147:382–395.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tay Y, Kats L, Salmena L, Weiss D, Tan SM,
Ala U, Karreth F, Poliseno L, Provero P, Di Cunto F, et al:
Coding-independent regulation of the tumor suppressor PTEN by
competing endogenous mRNAs. Cell. 147:344–357. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ma C, Nong K, Zhu H, Wang W, Huang X, Yuan
Z and Ai K: H19 promotes pancreatic cancer metastasis by
derepressing let-7's suppression on its target HMGA2-mediated EMT.
Tumour Biol. 35:9163–9169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cao C, Zhang T, Zhang D, Xie L, Zou X, Lei
L, Wu D and Liu L: The long non-coding RNA, SNHG6-003, functions as
a competing endogenous RNA to promote the progression of
hepatocellular carcinoma. Oncogene. 36:1112–1122. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhou X, Liu J and Wang W: Construction and
investigation of breast-cancer-specific ceRNA network based on the
mRNA and miRNA expression data. IET Syst Biol. 8:96–103. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Du Z, Sun T, Hacisuleyman E, Fei T, Wang
X, Brown M, Rinn JL, Lee MG, Chen Y, Kantoff PW and Liu XS:
Integrative analyses reveal a long noncoding RNA-mediated sponge
regulatory network in prostate cancer. Nat Commun. 7:109822016.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jiang H, Ma R, Zou S, Wang Y, Li Z and Li
W: Reconstruction and analysis of the lncRNA-miRNA-mRNA network
based on competitive endogenous RNA reveal functional lncRNAs in
rheumatoid arthritis. Mol Biosyst. 13:1182–1192. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lai Y, He S, Ma L, Lin H, Ren B, Ma J, Zhu
X and Zhuang S: HOTAIR functions as a competing endogenous RNA to
regulate PTEN expression by inhibiting miR-19 in cardiac
hypertrophy. Mol Cell Biochem. 432:179–187. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lin R and Turecki G: Noncoding RNAs in
depression. Adv Exp Med Biol. 978:197–210. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Barrett T, Wilhite SE, Ledoux P,
Evangelista C, Kim IF, Tomashevsky M, Marshall KA, Phillippy KH,
Sherman PM, Holko M, et al: NCBI GEO: Archive for functional
genomics data sets-update. Nucleic Acids Res. 41:D991–D995. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li JH, Liu S, Zhou H, Qu LH and Yang JH:
starBase v2.0: Decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA
interaction networks from large-scale CLIP-Seq data. Nucleic Acids
Res. 42:D92–D97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Langfelder P and Horvath S: WGCNA: An R
package for weighted correlation network analysis. BMC
Bioinformatics. 9:5592008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: New perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bader GD and Hogue CW: An automated method
for finding molecular complexes in large protein interaction
networks. BMC Bioinformatics. 4:22003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Subramanian A, Tamayo P, Mootha VK,
Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub
TR, Lander ES and Mesirov JP: Gene set enrichment analysis: A
knowledge-based approach for interpreting genome-wide expression
profiles. Proc Natl Acad Sci USA. 102:15545–15550. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lamb J, Crawford ED, Peck D, Modell JW,
Blat IC, Wrobel MJ, Lerner J, Brunet JP, Subramanian A, Ross KN, et
al: The connectivity map: Using gene-expression signatures to
connect small molecules, genes, and disease. Science.
313:1929–1935. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cai N, Li Y, Chang S, Liang J, Lin C,
Zhang X, Liang L, Hu J, Chan W, Kendler KS, et al: Genetic control
over mtDNA and Its relationship to major depressive disorder. Curr
Biol. 25:3170–3177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Czibere L, Baur LA, Wittmann A, Gemmeke K,
Steiner A, Weber P, Pütz B, Ahmad N, Bunck M, Graf C, et al:
Profiling trait anxiety: Transcriptome analysis reveals cathepsin B
(Ctsb) as a novel candidate gene for emotionality in mice. PLoS
One. 6:e236042011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kripke DF, Nievergelt CM, Joo E, Shekhtman
T and Kelsoe JR: Circadian polymorphisms associated with affective
disorders. J Circadian Rhythms. 7:22009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kripke DF, Nievergelt CM, Tranah GJ,
Murray SS, Rex KM, Grizas AP, Hahn EK, Lee HJ, Kelsoe JR and Kline
LE: FMR1, circadian genes and depression: Suggestive associations
or false discovery? J Circadian Rhythms. 11:32013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Maes M, Ombelet W, Verkerk R, Bosmans E
and Scharpé S: Effects of pregnancy and delivery on the
availability of plasma tryptophan to the brain: Relationships to
delivery-induced immune activation and early post-partum anxiety
and depression. Psychol Med. 31:847–858. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Murray JM, Sletten TL, Magee M, Gordon C,
Lovato N, Bartlett DJ, Kennaway DJ, Lack LC, Grunstein RR, Lockley
SW, et al: Prevalence of circadian misalignment and its association
with depressive symptoms in delayed sleep phase disorder. Sleep.
402017.doi: 10.1093/sleep/zsw002.
|
|
32
|
Takaesu Y, Inoue Y, Ono K, Murakoshi A,
Futenma K, Komada Y and Inoue T: Circadian rhythm sleep-wake
disorders as predictors for bipolar disorder in patients with
remitted mood disorders. J Affect Disord. 220:57–61. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee JH, Park SK, Ryoo JH, Oh CM, Mansur
RB, Alfonsi JE, Cha DS, Lee Y, McIntyre RS and Jung JY: The
association between insulin resistance and depression in the Korean
general population. J Affect Disord. 208:553–559. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chen WV, Nwakeze CL, Denny CA, O'Keeffe S,
Rieger MA, Mountoufaris G, Kirner A, Dougherty JD, Hen R, Wu Q and
Maniatis T: Pcdhalphac2 is required for axonal tiling and assembly
of serotonergic circuitries in mice. Science. 356:406–411. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bai M, Zhu X, Zhang Y, Zhang S, Zhang L,
Xue L, Yi J, Yao S and Zhang X: Abnormal hippocampal BDNF and
miR-16 expression is associated with depression-like behaviors
induced by stress during early life. PLoS One. 7:e469212012.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Baudry A, Mouillet-Richard S, Schneider B,
Launay JM and Kellermann O: miR-16 targets the serotonin
transporter: A new facet for adaptive responses to antidepressants.
Science. 329:1537–1541. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Shao QY, You F, Zhang YH, Hu LL, Liu WJ,
Liu Y, Li J, Wang SD and Song MF: CSF miR-16 expression and its
association with miR-16 and serotonin transporter in the raphe of a
rat model of depression. J Affect Disord. 238:609–614. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Gheysarzadeh A, Sadeghifard N, Afraidooni
L, Pooyan F, Mofid MR, Valadbeigi H, Bakhtiari H and Keikhavani S:
Serum-based microRNA biomarkers for major depression: MiR-16,
miR-135a, and miR-1202. J Res Med Sci. 23:692018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mihailova S, Ivanova-Genova E, Lukanov T,
Stoyanova V, Milanova V and Naumova E: A study of TNF-α, TGF-β,
IL-10, IL-6, and IFN-γ gene polymorphisms in patients with
depression. J Neuroimmunol. 293:123–128. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ostacher MJ, Iosifescu DV, Hay A,
Blumenthal SR, Sklar P and Perlis RH: Pilot investigation of
isradipine in the treatment of bipolar depression motivated by
genome-wide association. Bipolar Disord. 16:199–203. 2014.
View Article : Google Scholar : PubMed/NCBI
|