Introduction
Spinal cord injury (SCI) can lead to permanent
neurological deficits, including motor and sensory impairments,
with high physical disability and mortality rates. It can lead to
serious damage to patients' physical and mental health, and cause
severe social problems and economic burden (1). Although there are currently no
effective therapies for the treatment of SCI, various types of stem
cells, including embryonic, fetal and adult stem cells, have been
widely used in cellular transplantation therapies for SCI,
attracting increasing attention due to their satisfactory
pre-clinical therapeutic effects (2). Among them, bone marrow mesenchymal
stem cells (BMSCs) have emerged as one of the most promising types
of stem cells due to their favorable ethical and safety profiles
(3). Various studies have
suggested that BMSCs transplanted directly into the lesion can
promote axonal regeneration and contribute to motor and sensory
improvement following SCI (4). The
underlying mechanisms may include the differentiation of BMSCs
toward neuronal cells for improving structural and functional
repair of SCI and the secretion of various neurotrophic cytokines
stimulating nerve growth, including brain-derived neurotrophic
factor, nerve growth factor, vascular endothelial growth factor,
fibroblast growth factor, hepatocyte growth factor, insulin-like
growth factor and stem cell-derived factor (5,6).
Neuregulins (NRGs) comprise a large family of widely
expressed epidermal growth factor (EGF)-like proteins which are
essential in neural development and brain activity homeostasis
(7). Four NRG genes
(NRG1-4) and 30 NRG isoforms have been reported previously
(8). The NRG1 gene isoforms
are composed of three types and the NRG1 type I isoform, also known
as heregulin (HRG, 44 kDa) containing an Ig-like region and a
glycosylation-rich segment, is predominantly expressed in the
nervous system due to its acetylcholine receptor-inducing activity
(9). HRG can promote the survival
of neurons, glial cells (10,11),
epithelial cells (12),
cardiomyocytes (13,14) and other cell types (15) binding with ErbB tyrosine kinase
transmembrane receptors, including ErbB1/EGFR (16), ErbB2/neu (17), ErbB3 (18) and ErbB4 (19). The NRG/ErbB3/4 signaling pathway is
important in the regulation of numerous neurodevelopmental and
activity-dependent processes, including cellular growth,
proliferation, differentiation, migration and apoptosis (20).
Originally, the epithelial-mesenchymal transition
(EMT) was identified as a morphological conversion occurring at
specific stage of embryonic development and in the transitional
stage between the early stage and progressive stage of tumors
(21). The Snail signaling pathway
may be involved in the migration of stem cells due to its
well-established role in the EMT process (22). Although the role of Snail in
tumor-related invasion and metastasis has been elucidated, there
remains no clear understanding on its effects on the function of
BMSCs.
Therefore, the present study aimed to clarify the
underlying mechanism of NRG1 in the migration of BMSCs contributing
to the functional recovery of SCI. Firstly, the association between
NRG1 and Snail was investigated. Subsequently, an animal model of
SCI was established using Allen's method and BMSCs modified with a
Snail-overexpression plasmid vector were transplanted into the
lesion of the spinal cord. The expression levels of NRG1, Snail and
matrix metalloproteinase-2 (MMP-2) were detected by western blot
and reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) analyses. Furthermore, the functional recovery of the
spinal cord was evaluated using the Basso, Beattie, and Bresnahan
(BBB) rating scales.
Materials and methods
Cell cultures
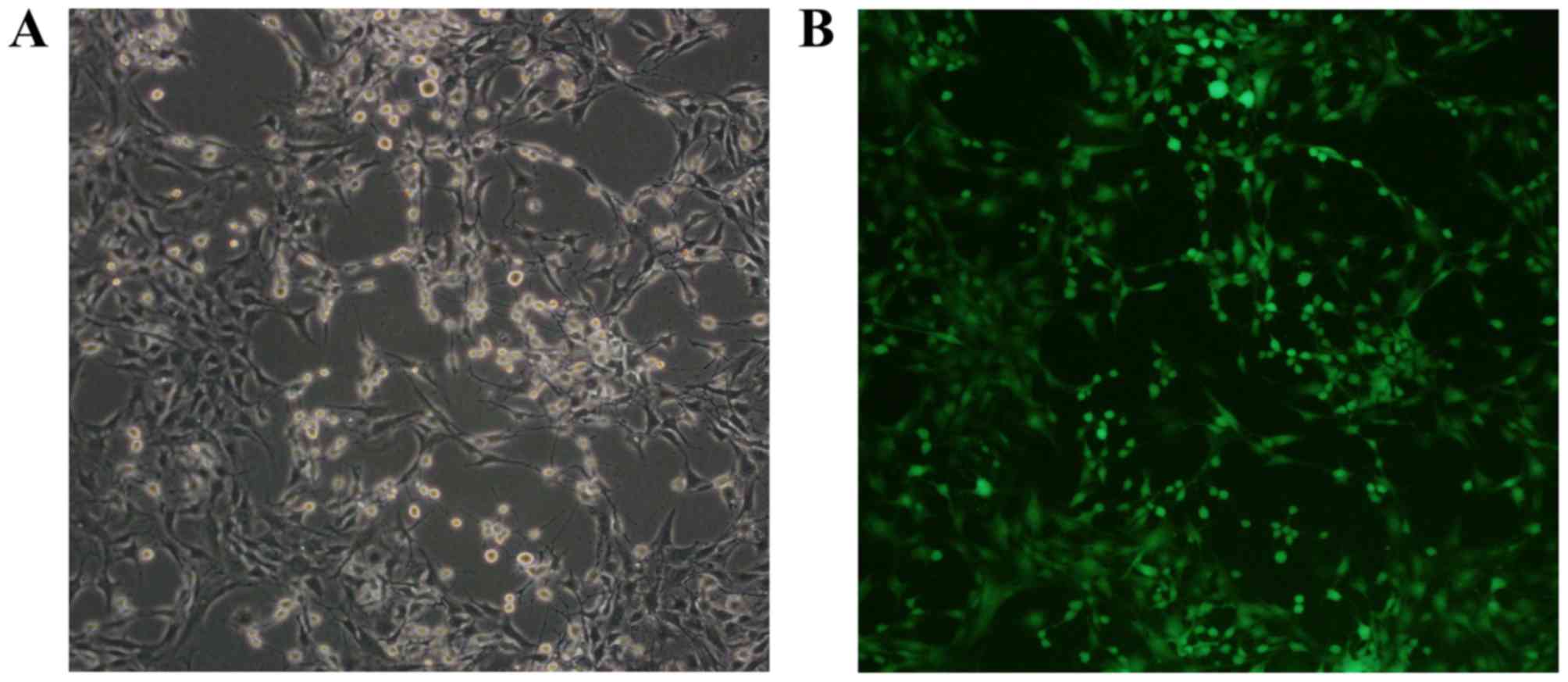
The green fluorescent protein (GFP)-labeled rat
BMSCs were purchased from Cyagen Biosciences, Inc. (Guangzhou,
China) and >95% BMSCs expressed GFP (Fig. 1). The surface markers of
CD44+, CD90+, CD11b−,
CD34− and CD45− were confirmed by the
manufacturer. The BMSCs were cultivated in Dulbecco's modified
Eagle's medium (DMEM)/F12 with 10% fetal bovine serum (FBS), 100 U
penicillin and 100 mg/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.). The cells were maintained at 37°C in a
humidified atmosphere containing 5% CO2 and passaged
routinely for experiments.
RT-qPCR analysis
Total RNA from the BMSCs was extracted using TRIzol
(Takara Bio, Inc., Otsu, Japan) according to the manufacturer's
protocol. The concentration was detected by measuring the
UV-absorption at 260 nm. The reaction mixture obtained following
the manufacturer instruction, was incubated at 42°C for 45 min and
85°C for 10 min. The primers were designed as follows: Snail,
forward 5′-ATGCACATCCGAAGCCACACGC-3′ and reverse
5′-GTAGGTTGGAGCGGTCGGCAAA-3′; MMP-2, forward
5′-TTTCCATTCCGCTTCCAGGGCACAT-3′ and reverse
5′-TCGCACACCACATCTTTCCGTCACT-3′; GAPDH, forward
5′-GTCTTCACCACCATGGAGAAGGCTG-3′ and reverse
5′-TGAGGTCCACCACCCTGTTGCTGTA-3′. qPCR analysis was performed using
a SYBR Premix Ex Taq II kit (Takara Bio, Inc., Otsu, Japan) with 20
µl solution containing 10 µl reaction mixture, 2 µl of cDNA, and
300 nM of gene-specific primers. The PCR samples were denatured at
95°C for 3 min and 40 cycles were performed at 95°C for 12 sec, and
at 62°C for 40 sec. The efficiency of the reaction was measured
with primers using serial dilutions of the cDNA (1:1, 1:5, 1:25,
1:125, 1:625 and 1:3,125). Each sample was tested in triplicate.
The relative gene expression levels were analyzed using the Pfaffl
method. The data are expressed as fold change relative to untreated
controls after normalizing to β-actin (23).
Overexpression plasmid and
transfection
The Snail overexpression plasmid (pBabe-puro-Snail),
the mammalian expression vector with full length human Snail cDNA,
and the negative comparing plasmid (pBabe-puro) were provided by Dr
Bob Weinberg (Five Cambridge Centre, Cambridge, MA, USA) from
Addgene, Inc. (Cambridge, MA, USA). The BMSCs were transfected with
the Snail overexpression plasmid using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA)
based on the manufacturer's protocol. Ampicillin-resistant clones
were isolated as single colonies.
Methyl thiazolyl tetrazolium (MTT)
assay
Each group (5×104 cells per well) of
cells were incubated in 96-well plates containing 100 µl DMEM
(Gibco; Thermo Fisher Scientific, Inc.) in each well. The rates of
cellular proliferation were measured following 24, 48, 72, and 96 h
of incubation. Subsequently, 20 µl of MTT (5 mg/ml) was added into
each well and incubated for 4 h at 37°C. Following removal of the
culture medium from each well, 150 µl of DMSO was added to each
well and the well-plate was detected using a spectrometer at 490
nm.
Western blot analysis
RIPA lysis buffer was added into the cell suspension
and reacted for 30 min at 4°C. Subsequently, the samples were
boiled for 10 min and centrifuged at 12,000 × g for 5 min at room
temperature. The supernatants were collected and the quantity of
protein loaded per well was adjusted. The protein sample (20 mg)
from each lysate was separated by using 10–12% sodium dodecyl
sulfate-polyacrylamide (SDS-PAGE) gel electrophoresis, transferred
onto nitrocellulose membranes. The membranes were blocked using 5%
non-fat milk in PBS (pH 7.2) for 2 h at room temperature and
incubated with primary antibodies overnight at 4°C, including
anti-Snail (1:500; ab53519; Abcam, Cambridge, MA, USA), anti-MMP-2
(1:100; ab135562; Abcam) and GAPDH (1:400; ab8245; Abcam). The
membranes were then washed with 1% TBS-Tween and incubated with
secondary antibodies for 2 h at room temperature. An ECL
chemiluminescent substrate kit was used to detect the reaction and
images were analyzed using AlphaEase FC software version 3.0
(ProteinSimple, San Jose CA, USA). The western blot assays were
conducted in at least three independent experiments.
Cell migration assay
The migration of stem cells was measured based on
counting the number of cells that passed through a gelation-coated
polycarbonate membrane with an 8-µm pore size. The cells were
suspended and seeded onto the Transwell membrane with the density
of 1×103 cells/well. For chemotaxis experiments, 15% FBS
(Gibco; Thermo Fisher Scientific, Inc.) were placed into the wells
of the lower chamber compartment. The cells were fixed with
methanol at −20°C for 30 min and stained with Giemsa at room
temperature for 30 min. The number of migratory cells was counted
with an inverted microscope at ×200 magnification in five
fields.
SCI model
A total of 36 six-week-old female Sprague-Dawley
(SD) rats were purchased from the Laboratory of Animal Technology
of China Medical University (CMU; Shenyang, China). All rats were
housed under specific pathogen-free conditions (22C, 12-h
light/dark cycle and 50–55% humidity) with free access to food
pellets and tap water. The SD rats (body weight range, 200–240 g)
were anesthetized by intraperitoneal injection of chloral hydrate
(300 mg/kg) and received a laminectomy at the 10th thoracic
vertebral body under a surgical microscope. The animals received 80
g/cm blast injury using the Infinite Horizon Impactor (Laboratory
Technology Center of CMU). A 1-µl cell suspension of
1×105 cells was injected into the lesions of the spinal
cord. Subsequently, 5-0 chromic gut sutures were used for suturing
of the incision layer by layer and pressure was placed on the
bladder for urination every day. Ampicillin was administered daily
in order to avoiding infections during the entire study. All
experiments were performed in accordance with the Chinese Community
Guidelines (24).
BBB scoring and specimen
preparation
The functional recovery of the spinal cord was
evaluated using the BBB rating scales (25); this was performed by two observers
in a blinded-manner every week following cell transplantation.
Subsequently, the rats were sacrificed with an overdose of sodium
pentobarbital, followed by trans-cardiac perfusion with 4%
paraformaldehyde fixing for 2 h. The specimens, cut into segments 1
cm in length, were placed in 30% sucrose solution for 48 h. These
entire segments were cryosectioned longitudinally into 6-µm serial
sections and collected on slides. The numbers of GFP-labeled cells
were counted under a fluorescence microscope.
Statistical analysis
Statistical analysis was performed using Statistical
Product and Service Solutions software (SPSS 19.0; IBM Corp,
Armonk, NY, USA). The results are presented as the mean ± standard
deviation. One-way analysis of variance was applied for the
comparison of mean values of different groups and Tukey's test was
used as post hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
NRG1 significantly increases the mRNA
expression level of Snail in a concentration-dependent manner
The effects of different concentrations of NRG1
(0–80 ng/ml) on the mRNA expression level of Snail were detected by
RT-qPCR analysis at 24, 48, and 72 h. As shown in Fig. 2A, NRG1 significantly increased the
mRNA expression of Snail in a concentration- dependent manner
between 0 and 40 ng/ml, with a peak at 40 ng/ml. At the dose of 80
ng/ml NRG1, the expression level of Snail was significantly lower
than that at the dose of 40 ng/ml NRG1. The mRNA expression level
of Snail was significantly higher at 48 h than that at either 24 or
72 h post-NRG1 treatment (P<0.05). These results suggested that
NRG1 at a dose of 40 ng/ml for 48 h led to the most effective
increase on the expression of Snail in BMSC, which was the optimal
condition used in the following experiments.
 | Figure 2.Effects of NRG1 on the expression
levels of Snail and MMP-2. (A) NRG1 concentration-dependently
increased the expression level of Snail. *P<0.05 48. vs. 24 h;
**P<0.05 48, vs. 72 h. Expression levels of (B) Snail and (C)
MMP-2 in the different groups of BMSCs. *P<0.05 BMSC-Sna + NRG
group, vs. BMSC-NC + NRG, BMSC + NRG, BMSC-NC, and BMSC groups;
**P<0.05 BMSC-Sna + NRG group, vs. BMSC-Sna group;
#P<0.05 BSMC-Sna group, vs. BMSC-NC + NRG, BMSC +
NRG, BMSC-NC, and BMSC groups. Data are presented as the mean ±
standard deviation and P<0.05 was considered statistically
significant. BMSC, bone marrow mesenchymal stem cells; Sna, Snail
overexpression plasmid; NRG, neuregulin-1; NC, negative control;
MMP-2, matrix metalloproteinase-2. |
Effects of NRG1 on the mRNA and
protein expression levels of Snail and MMP-2
A total of six groups were included in the study:
Group A (BMSCs transfected with Snail overexpression plasmid plus
NRG1; BMSC-Sna + NRG1), group B (BMSCs transfected with Snail
negative expression plus NRG1; BMSC-NC + NRG1), group C (BMSCs plus
NRG; BMSC + NRG1), group D (BMSCs transfected with Snail
overexpression; BMSC-Sna), group E (BMSCs transfected with Snail
negative expression; BMSC-NC) and group F (BMSCs). As shown in
Fig. 2B, the expression levels of
Snail mRNA in groups A and D were significantly higher than those
in groups B, C, E and F due to transfection with the Snail
overexpression plasmid (P<0.05). There were significant
differences between group A and group D (P<0.05). Furthermore,
the mRNA expression levels of MMP-2 in groups A and D were
significantly higher than those in groups B, C, E and F, which
indicated that the overexpression of Snail stimulated the
expression of MMP-2, and NRG1 further enhanced this positive effect
of Snail (P<0.05; Fig. 2C). As
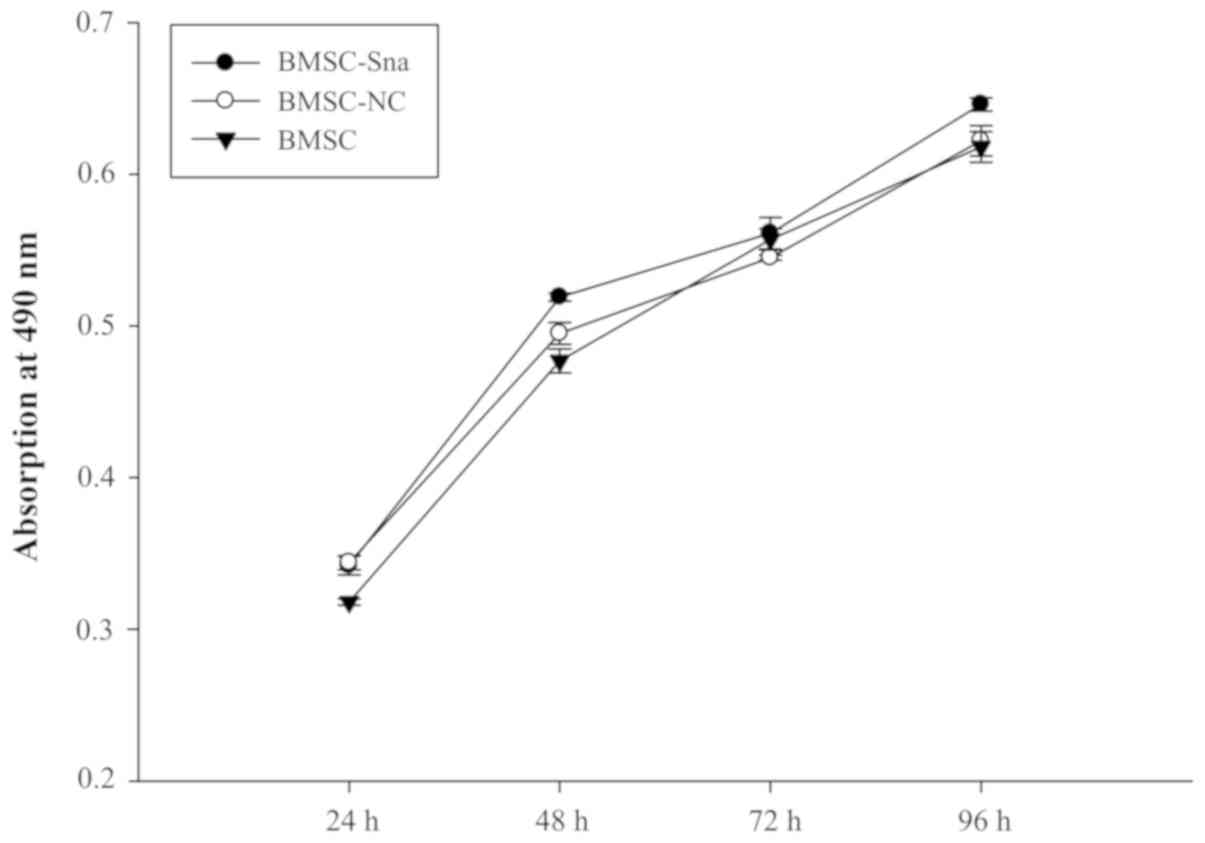
shown in Fig. 3, no significant
differences in cellular viability were observed between groups D, E
and F at different time points (P>0.05). This indicated that the
cellular growth of BMSCs did not significantly alter due to
transfection with the Snail overexpression plasmid. According to
Fig. 4A-C, the results of the
western blot analysis revealed that the expression level of Snail
in group A was significantly higher than those of groups B, C, E
and F (P<0.05) and the expression level of Snail in group was
significantly higher compared with those of groups E and F
(P<0.05). No significant differences were observed between
groups A and D (P>0.05). In addition, the expression levels of
MMP-2 in groups A and D were significantly higher than those of
groups B, C, E and F (P<0.05).
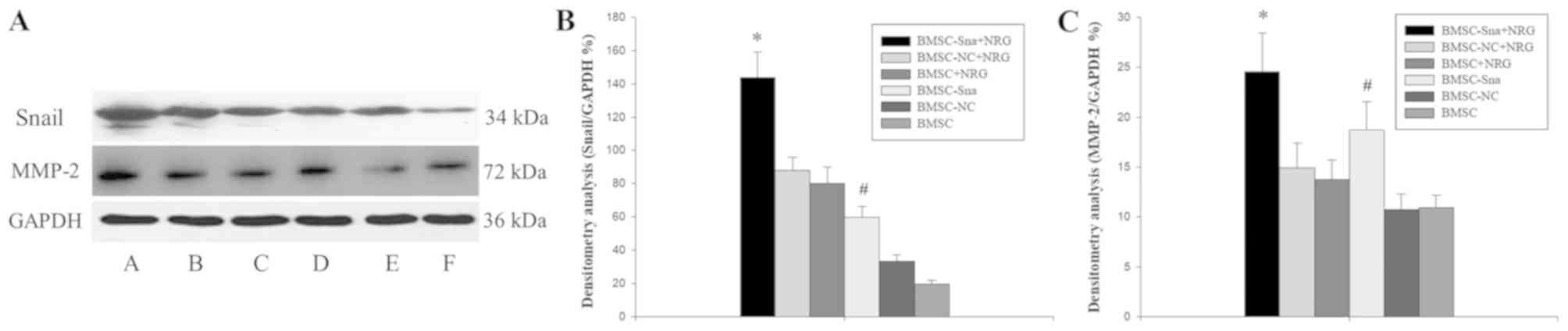 | Figure 4.Western blot analysis results. (A)
Expression of Snail and MMP-2 in the different groups of BMSCs: A,
BMSC-Sna + NRG; B, BMSC-NC + NRG; C, BMSC + NRG; D, BMSC-Sna; E,
BMSC-NC; F, BMSC. Densitometric analysis of (B) Snail and (C)
MMP-2. Data are presented as the mean ± standard deviation and
P<0.05 was considered statistically significant. *P<0.05
BMSC-Sna + NRG group, vs. BMSC-NC + NRG, BMSC + NRG, BMSC-NC, and
BMSC groups; #P<0.05 BMSC-Sna group, vs. BMSC-NC +
NRG, BMSC + NRG, BMSC-NC, and BMSC groups. BMSC, bone marrow
mesenchymal stem cells; Sna, Snail overexpression plasmid; NRG,
neuregulin-1; NC, negative control; MMP-2, matrix
metalloproteinase-2. |
Cell migration of BMSCs in vitro
As shown in Fig. 5A and
B, the numbers of cells crossing the membrane in groups A and D
were significantly higher than those in groups B and C (P<0.05).
Additionally, there were significant differences between groups A
and D in the numbers of cells that crossed the membrane
(P<0.05). However, no cells appeared to cross the membrane in
groups E and F. This suggested that NRG1 enhanced the promoting
effect of Snail on BMSC migration.
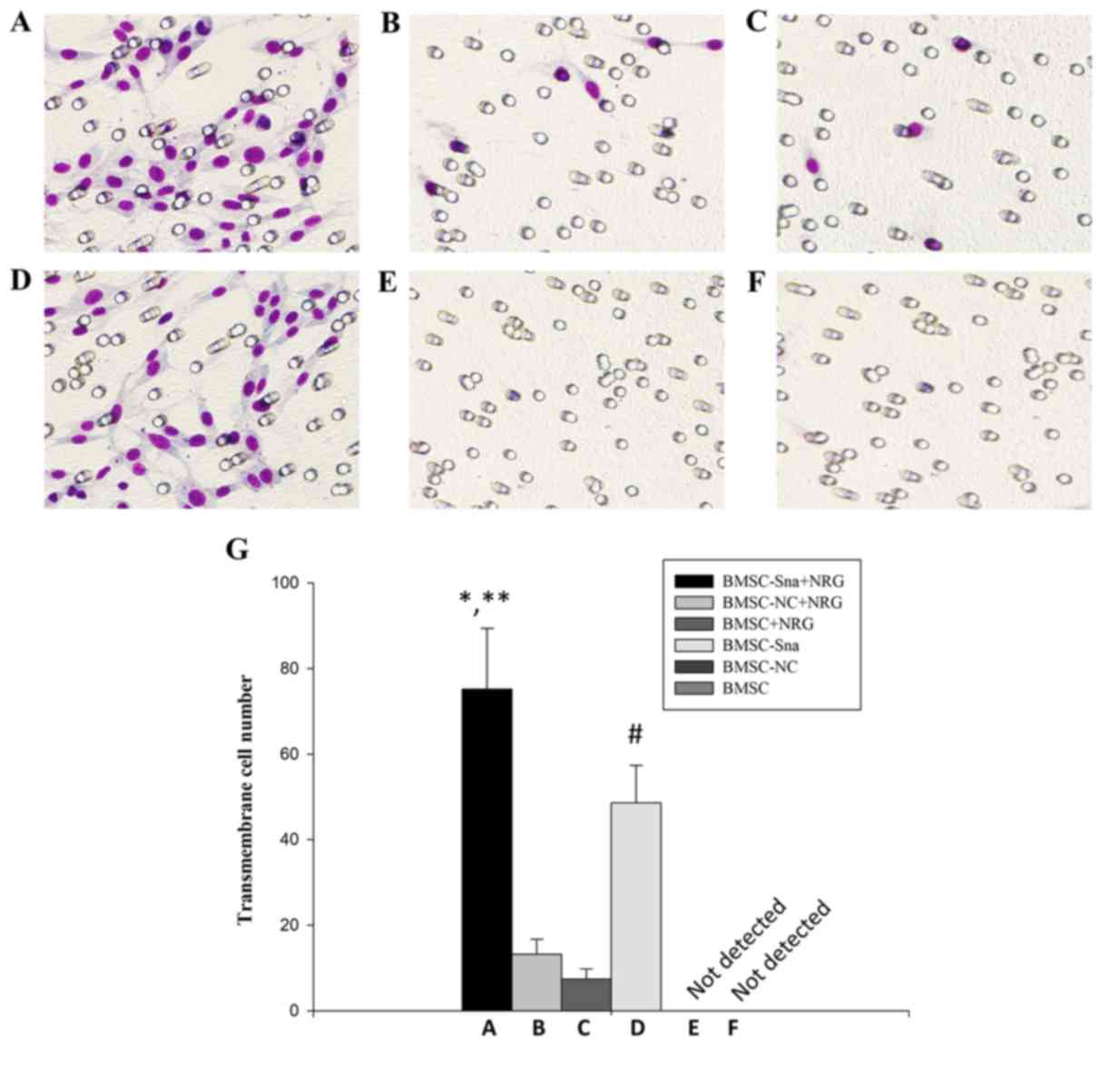 | Figure 5.Cell migration of BMSCs in different
groups. Images under an optical microscope (×100 magnification) of
the (A) BMSC-Sna + NRG group, (B) BMSC-NC + NRG group, (C) BMSC +
NRG group, (D) BMSC + Sna group, (E) BMSC-NC group and (F) BMSC
group. (G) Numbers of transmembrane cells in groups A-F. Data are
presented as the mean ± standard deviation and P<0.05 was
considered statistically significant. *P<0.05 A, vs. B and C;
**P<0.05 A, vs. D; #P<0.05 D, vs. B and C. BMSC,
bone marrow mesenchymal stem cells; Sna, Snail overexpression
plasmid; NRG, neuregulin-1; NC, negative control. |
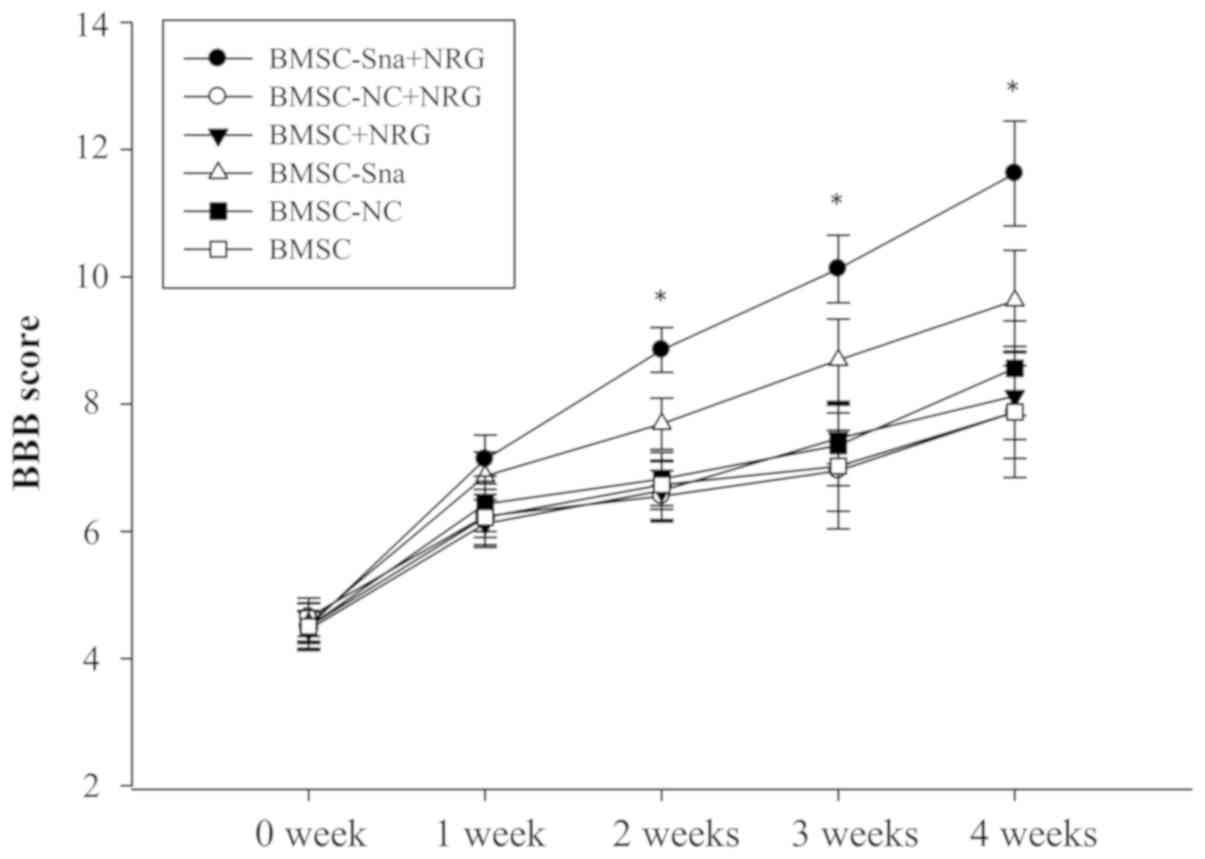
BBB scores and assessment of
functional recovery
Two experts were invited to perform the BBB rating
scale in a blinded-manner at the end of each week between weeks 0
and 4 following cell transplantation. The BBB scores of group D
were significantly higher than those of groups B, C, E and F
between 1 and 4 weeks post-BMSC transplantation (P<0.05). The
BBB score of group A was significantly higher than the scores of
the other groups at different time points, including weeks 2, 3 and
4 (P<0.05; Fig. 6). In
addition, the distribution area of BMSCs in group A was
significantly larger than those of the other groups and the
distribution area of BMSCs in group D was significantly larger than
those of groups E and F (Fig.
7A-G).
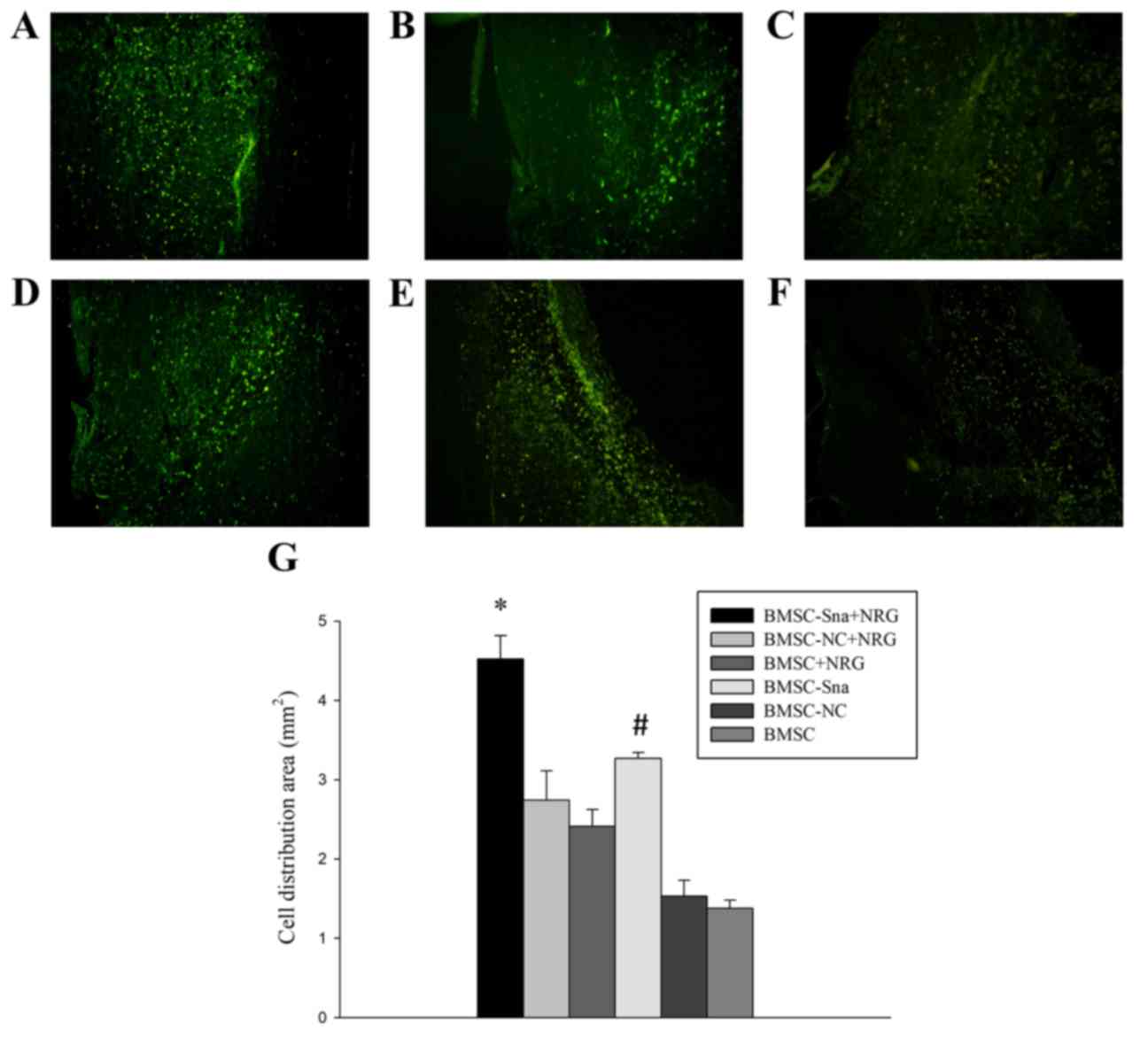 | Figure 7.Morphological observation of BMSCs in
different groups. Images of the spinal cord in the (A) BMSC-Sna +
NRG group, (B) BMSC-NC + NRG group, (C) BMSC + NRG group, (D) BMSC
+ Sna group, (E) BMSC-NC group and (F) BMSC group under a
fluorescent microscope (magnification, ×40). (G) Distribution area
of BMCSs. Data are presented as the mean ± standard deviation and
P<0.05 was considered statistically significant. *P<0.05
BMSC-Sna + NRG group, vs. all other groups; #P<0.05
BMSC-Sna group, vs. BMSC-NC and BMSC groups. BMSC, bone marrow
mesenchymal stem cells; Sna, Snail overexpression plasmid; NRG,
neuregulin-1; NC, negative control. |
Discussion
Currently, substantial efforts are required for
fulfilling the clinical application of stem cell transplantation in
the treatment of SCI (26,27). In particular, the inefficient
migration of BMSCs into the lesions in vivo is one of
biggest problems in these studies. Several previous studies have
suggested that stromal cell-derived factor-1 (28), monocyte chemoattractant protein-1
(29) and interleukin-8 can induce
BMSC migration in vitro. However, there are no reports on
whether NRG1 can induce BMSC migration into lesions of SCI and its
underlying mechanisms.
The application of BMSCs as seed cells is effective
and efficient for cell transplantation in the treatment of SCI due
to lower immunogenicity and no adverse reactions (30). However, Serakinci et al
(31) suggested that the existence
of tumorigenicity was confirmed when BMSCs were transfected with
the telomerase gene in vitro. In addition, Rubio et
al (32) reported that
transgenic BMSCs are safe and effective for 6–8 weeks in
vitro, however, spontaneously emerging mutations were
inevitable in the long-term. Therefore, temporary tumorigenesis
BMSCs were introduced in the present study, which meant that BMSCs
were temporarily modified with gene overexpression or
silencing.
In the present study, it was found that exogenous
NRG1 significantly promoted the expression of Snail in a
concentration-dependent manner <40 ng/ml with a peak at 48 h.
Further experiments were performed under this optimal condition. As
overlapping mechanisms exist between stem cells and tumor cells
(33–37), the transcription factor Snail,
which is important in EMT, is involved in promoting the migration,
invasiveness and metastatic potential of tumor cells (38–41).
Therefore, BMSCs were modified with a Snail-overexpressed plasmid
and changes in the expression of MMP-2 were detected with or
without the addition of exogenous NRG1. The results indicated that
NRG1 significantly promoted the expression of MMP-2 via
upregulating the expression of Snail and contributing to BMSC
migration in vitro. The synergistic effects of NRG1 on Snail
can promote changes of the BMSC cytoskeleton, stimulate the
expression of MMP-2 to degrade extracellular matrix, and enhance
BMSC migration. The results of in vivo experiments in the
present study further showed that BMSCs had a wide and uniform
distribution area in the lesions of the SCI following treatment
with BMSCs modified with Snail and NRG1.
Furthermore, the BBB scores indicated that the
functional recovery of SCI was significantly improved following
transplantation of the BMSCs modified with Snail and NRG1. The
underlying mechanism of this promoting effect may be associated
with the upregulation of MMP-2 and Snail following the addition of
NRG1. The results suggested that NRG1 provided synergistic
improvement of the positive effects of Snail on BMSC migration.
BMSC transplantation offers a novel and promising treatment for SCI
and previous studies have confirmed that BMSC transplantation can
improve neurological deficits effectively and efficiently. However,
there were certain limitations, including the lack of more recent
and advanced techniques, the insufficient number of experimental
animal species and the lack of evaluation of the effect of BMSC
migration following NRG1- or Snail-knockout.
In conclusion, NRG1 upregulated the expression of
Snail in a concentration-dependent and promoted the expression of
MMP-2 at the optimal concentration and time point; this enhanced
the migration of rat BMSCs in vitro. In addition, NRG1 led
to the BMSCs diffusing uniformly in the SCI location and showed
good functional recovery following SCI. This suggests that NRG1 may
be essential in the cell transplantation of modified BSMCs in
treating SCI.
Acknowledgements
Not applicable.
Funding
The present study was sponsored by National Natural
Science Foundation of China (grant no. 81070971), who had no role
in study design, data collection and analysis, decision to publish,
or preparation of the manuscript.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XY, YC and GT were involved in conception and
design, collection and assembly of data, analysis and
interpretation of data, statistical expertise, drafting of the
manuscript, final approval of the manuscript, provision of study
materials and administrative, technical or logistic support;
analysis and interpretation of data, critical revision of the
manuscript for important intellectual content. All authors
contributed equally.
Ethics approval and consent to
participate
This study was approved by the Ethics Committee of
The First Affiliated Hospital of China Medical University
(Shenyang, China). Participants provided written informed consent
to participate in the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Jain NB, Ayers GD, Peterson EN, Harris MB,
Morse L, O'Connor KC and Garshick E: Traumatic spinal cord injury
in the United States, 1993–2012. JAMA. 313:2236–2243. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hofstetter CP, Schwarz EJ, Hess D,
Widenfalk J, El Manira A, Prockop DJ and Olson L: Marrow stromal
cells form guiding strands in the injured spinal cord and promote
recovery. Proc Natl Acad Sci USA. 99:2199–2204. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Qu J and Zhang H: Roles of mesenchymal
stem cells in spinal cord injury. Stem Cells Int. 2017:52513132017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Nakano N, Nakai Y, Seo TB, Homma T, Yamada
Y, Ohta M, Suzuki Y, Nakatani T, Fukushima M, Hayashibe M and Ide
C: Effects of bone marrow stromal cell transplantation through CSF
on the subacute and chronic spinal cord injury in rats. PLoS One.
8:e734942013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Miyahara Y, Nagaya N, Kataoka M, Yanagawa
B, Tanaka K, Hao H, Ishino K, Ishida H, Shimizu T, Kangawa K, et
al: Monolayered mesenchymal stem cells repair scarred myocardium
after myocardial infarction. Nat Med. 12:459–465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wright KT, El Masri W, Osman A, Chowdhury
J and Johnson WE: Concise review: Bone marrow for the treatment of
spinal cord injury: Mechanisms and clinical applications. Stem
Cells. 29:169–178. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Mei L and Nave KA: Neuregulin-ERBB
signaling in the nervous system and neuropsychiatric diseases.
Neuron. 83:27–49. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Birchmeier C and Bennett DL:
Neuregulin/ErbB signaling in developmental myelin formation and
nerve repair. Curr Top Dev Biol. 116:45–64. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lai D, Liu X, Forrai A, Michalicek J,
Ahmed I, Garratt AN, Birchmeier C, Zhou M, Hartley L, Robb L, et
al: Neuregulin 1 sustains the gene regulatory network in both
trabecular and nontrabecular myocardium. Circ Res. 107:715–727.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Simmons LJ, Surles-Zeigler MC, Li Y, Ford
GD, Newman GD and Ford BD: Regulation of inflammatory responses by
neuregulin-1 in brain ischemia and microglial cells in vitro
involves the NF-kappa B pathway. J Neuroinflammation. 13:2372016.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ritch PS, Carroll SL and Sontheimer H:
Neuregulin-1 enhances survival of human astrocytic glioma cells.
Glia. 51:217–228. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Finigan JH, Faress JA, Wilkinson E, Mishra
RS, Nethery DE, Wyler D, Shatat M, Ware LB, Matthay MA, Mason R, et
al: Neuregulin-1-human epidermal receptor-2 signaling is a central
regulator of pulmonary epithelial permeability and acute lung
injury. J Biol Chem. 286:10660–10670. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Brea MS, Díaz RG, Escudero DS, Caldiz CI,
Portiansky EL, Morgan PE and Pérez NG: Epidermal growth factor
receptor silencing blunts the slow force response to myocardial
stretch. J Am Heart Assoc. 5:e0040172016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vermeulen Z, Hervent AS, Dugaucquier L,
Vandekerckhove L, Rombouts M, Beyens M, Schrijvers DM, De Meyer
GRY, Maudsley S, De Keulenaer GW and Segers VFM: Inhibitory actions
of the NRG-1/ErbB4 pathway in macrophages during tissue fibrosis in
the heart, skin, and lung. Am J Physiol Heart Circ Physiol.
313:H934–H945. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Woo RS, Lee JH, Kim HS, Baek CH, Song DY,
Suh YH and Baik TK: Neuregulin-1 protects against neurotoxicities
induced by swedish amyloid precursor protein via the ErbB4
receptor. Neuroscience. 202:413–423. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Miyagawa S, Katsu Y, Watanabe H and Iguchi
T: Estrogen-independent activation of erbBs signaling and estrogen
receptor alpha in the mouse vagina exposed neonatally to
diethylstilbestrol. Oncogene. 23:340–349. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Motoyama AB, Hynes NE and Lane HA: The
efficacy of ErbB receptor-targeted anticancer therapeutics is
influenced by the availability of epidermal growth factor-related
peptides. Cancer Res. 62:3151–3158. 2002.PubMed/NCBI
|
|
18
|
Alvarado D, Ligon GF, Lillquist JS, Seibel
SB, Wallweber G, Neumeister VM, Rimm DL, McMahon G and LaVallee TM:
ErbB activation signatures as potential biomarkers for anti-ErbB3
treatment in HNSCC. PLoS One. 12:e01813562017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wingens M, Walma T, van Ingen H,
Stortelers C, van Leeuwen JE, van Zoelen EJ and Vuister GW:
Structural analysis of an epidermal growth factor/transforming
growth factor-alpha chimera with unique ErbB binding specificity. J
Biol Chem. 278:39114–39123. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Vullhorst D, Ahmad T, Karavanova I,
Keating C and Buonanno A: Structural similarities between
neuregulin 1–3 isoforms determine their subcellular distribution
and signaling mode in central neurons. J Neurosci. 37:5232–5249.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Greenburg G and Hay ED: Epithelia
suspended in collagen gels can lose polarity and express
characteristics of migrating mesenchymal cells. J Cell Biol.
95:333–339. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang Y, Shi J, Chai K, Ying X and Zhou BP:
The role of snail in EMT and tumorigenesis. Curr Cancer Drug
Targets. 13:963–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xia J, Luo M, Ni N, Chen J, Hu Y, Deng Y,
Ji J, Zhou J, Fan X and Gu P: Bone marrow mesenchymal stem cells
stimulate proliferation and neuronal differentiation of retinal
progenitor cells. PLoS One. 8:e761572013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu JZ, Wu TD, Zhang T, Zhao YF, Pang J and
Lu HB: Three-dimensional alteration of microvasculature in a rat
model of traumatic spinal cord injury. J Neurosci Methods.
204:150–158. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
resting in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Svkova E, Jendelova P, Urdzikova L, Lesný
P and Hejcl A: Bone marrow stem cells and polymer hydrogels-two
strategied for spinal cord injury repair. Cell Mol Neurobiol.
26:1113–1129. 2006.PubMed/NCBI
|
|
27
|
Sykova E, Homola A, Mazanec R, Lachmann H,
Konrádová SL, Kobylka P, Pádr R, Neuwirth J, Komrska V, Vávra V, et
al: Autologous bone marrow transplantation in patients with
subacute and chronic spinal cord injury. Cell Transplant.
15:675–687. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hill WD, Hess DC, Martin-Studdard A,
Carothers JJ, Zheng J, Hale D, Maeda M, Fagan SC, Carroll JE and
Conway SJ: SDF-1 (CXCL12) is upregulated in the ischemic penumbra
following stroke: Association with bone marrow cell homing to
injury. J Neuropathol Exp Neurol. 63:84–96. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang L, Li Y, Chen X, Chen J, Gautam SC,
Xu Y and Chopp M: MCP-1, MIP-1, IL-8 and ischemic cerebral tissue
enhance human bone marrow stromal cell migration in interface
culture. Hematology. 7:113–117. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu D, Mahmood A, Wang L, Li Y, Lu M and
Chopp M: Adult bone marrow stromal cells administered intravenously
to rats after traumatic brain injury migrate into brain and improve
neurological outcome. Neuroreport. 12:559–563. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Serakinci N, Guldberg P, Burns JS,
Abdallah B, Schrødder H, Jensen T and Kassem M: Adult human
mesenchymal stem cell as a target for neoplastic transformation.
Oncogene. 23:5095–5098. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rubio D, Garcia-Castro J, Martin MC, de la
Fuente R, Cigudosa JC, Lloyd AC and Bernad A: Spontaneous human
adult stem cell transformation. Cancer Res. 65:3035–3039. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Reya T, Morrison SJ, Clarke MF and
Weissman IL: Stem cells, cancer, and cancer stem cells. Nature.
414:105–111. 2001. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Burkert J, Wright NA and Alison MR: Stem
cells and cancer: An intimate relationship. J Pathol. 209:287–297.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Al-Hajj M, Becker MW, Wicha M, Weissman I
and Clarke MF: Therapeutic implications of cancer stem cells. Curr
Opin Genet Dev. 14:43–47. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Nakagawara A and Ohira M: Comprehensive
genomics linking between neural development and cancer:
Neuroblastoma as a model. Cancer Lett. 204:213–224. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zha YH, He HF, Mei YW, Yin T and Mao L:
Zinc-finger transcription factor Snail accelerates survival,
migration and expression of matrix metalloproteinase-2 in human
bone mesenchymal stem cells. Cell Biol Int. 31:1089–1096. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ikernouchi J, Matsuda M, Furuse M and
Tsukita S: Regulation of tight junctions during the
epithelium-mesenchyme transition: Direct repression of the gene
expression of claudins/occluding by snail. J Cell Sci.
116:1959–1967. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Yang AD, Camp ER, Fan F, Shen L, Gray MJ,
Liu W, Somcio R, Bauer TW, Wu Y, Hicklin DJ and Ellis LM: Vascular
endothelial growth factor receptor-1 activation mediates epithelial
to mesenchymal transition in human pancreatic carcinoma cells.
Cancer Res. 66:46–51. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Gotzmann J, Fischer AN, Zojer M, Mikula M,
Proell V, Huber H, Jechlinger M, Waerner T, Weith A, Beug H and
Mikulits W: A crucial function of PDGF in TGF-beta-mediated cancer
progression of hepatocyers. Oncogene. 25:3170–3185. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Cheng L, Zha Z, Lang B, Liu J and Yao X:
Heregulin-beta 1 promotes metastasis of breast cancer cell line
SKBR3 through upregulation of Snail and induction of
epithelial-mesenchymal transition. Cancer Lett. 280:50–60. 2009.
View Article : Google Scholar : PubMed/NCBI
|