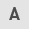Isoalantolactone inhibits constitutive NF-κB activation and induces reactive oxygen species-mediated apoptosis in osteosarcoma U2OS cells through mitochondrial dysfunction
- Authors:
- Published online on: July 30, 2014 https://doi.org/10.3892/or.2014.3368
- Pages: 1585-1593
Abstract
Introduction
Osteosarcoma is the most common form of primary malignant bone cancer in children and adolescents (1), and the 5-year survival rate of osteosarcoma patients is 61.6% (2). Although in the last few decades, multi-agent chemotherapy in combination with surgery has resulted in an improvement in the patient survival rate, which has increased to 70%, osteosarcoma is still characterized by frequent relapse and metastatic disease (3,4). Exploring novel therapeutic agents and their molecular mechanisms are, therefore, necessary for improving the outcome of osteosarcoma treatment.
In the present study, we screened 400 natural compounds against human osteosarcoma U2OS, MG-63 and Saos-2 cells. Several compounds were found cytotoxic, among which, isoalantolactone showed strong antiproliferative effects against osteosarcoma cells. Isoalantolactone has been reported to have a variety of biological effects, including antifungal, antihelmintic, and anticancer activities (5). A previous study by Khan et al showed that isoalantolactone did not exert any acute or chronic cytotoxicity in mouse liver and kidney (6). Although there are a few reports on the anticancer activities of isoalantolactone (6–10), there is no report available on the anti-osteosarcoma effect of isoalantolactone.
Apoptosis plays an important role in multiple steps leading to cell death. Two major pathways are involved in apoptosis: the extrinsic (death receptor) and the intrinsic (mitochondrial) pathways (11). The extrinsic pathways are involved in death receptor-induced intracellular signaling and the cleavage and activation of caspase-8 (12,13). Caspase-8 can then cleave effector caspase-3 to induce apoptosis. The intrinsic cell death pathways are mediated by modulation of Bcl-2 family proteins, mitochondrial membrane potential dissipation and cytochrome c release (14), which subsequently leads to the activation of caspase-3, resulting in apoptosis. To date, no report has addressed the role of isoalantolactone in the extrinsic pathways. The present study was undertaken to investigate the mechanisms of isoalantolactone-induced apoptosis in osteosarcoma. Here, our results demonstrated that isoalantolactone inhibits cell proliferation, triggers S phase and mainly G2/M phase arrest and induces ROS-dependent apoptosis in U2OS cells via a novel mechanism involving inhibition of NF-κBp65.
Materials and methods
Chemicals and antibodies
All of the chemicals were purchased from Sigma unless otherwise stated. Isoalantolactone (purity >98%) was purchased from Tauto Biotech Co., Ltd. (Shanghai, China). Fetal bovine serum (FBS) was purchased from Hangzhou Sijiqing Biological Engineering Materials Co., Ltd. Nuclear protein extraction kit was purchased from KeyGen (Nanjing, China), and the Annexin V-FITC apoptosis detection kit, and Bcl-2, caspase-3 and -8 antibodies, and the caspase inhibitor Z-VAD-FMK were purchased from Beyotime Institute of Biotechnology (Shanghai, China). PARP, Bax, cyclin B1, DR5, FADD, TRADD, RIP and NF-κBp65 antibodies were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). β-actin, secondary anti-mouse and anti-rabbit antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Cell culture and treatments
Human osteosacoma U2OS, MG-63 and Saos-2 cells were maintained in DMEM supplemented with 10% FBS, 100 units/ml penicillin and 100 μg/ml streptomycin at 37°C in a humidified atmosphere with 5% CO2. The cells were treated with various concentrations of drug dissolved in dimethyl sulfoxide (DMSO) with a final concentration <1%. DMSO-treated cells were used as the control.
Cell proliferation assay
The effects of isoalantolactone on cell viability were evaluated by MTT assay as previously described (15). U2OS, MG-63 and Saos-2 cells were treated with various concentrations (0–200 μM) of isoalantolactone for 24 h. Following treatment, the MTT reagent was added (500 μg/ml), and the cells were further incubated at 37°C for 4 h. Subsequently 150 μl DMSO was added to dissolve formazan crystals, and the absorbance was measured at 570 nm in a microplate reader (Thermo Scientific). IC50 values were calculated using GraphPad Prism 5.
Flow cytometric analysis of apoptosis
To determine apoptosis, U2OS cells were treated with 0, 20 or 40 μM isoalantolactone for 24 h. For the caspase inhibitor and ROS inhibitor analyses, the cells were pretreated with 50 μM Z-VAD-FMK and 3 mM NAC for 2 h and then incubated with 40 μM isoalantolactone for 24 h. Following the treatment, the cells were collected, washed with phosphate-buffered saline (PBS) and then re-suspended in binding buffer containing Annexin V-FITC and propidium iodide (PI), and incubated in the dark for 15 min at room temperature. Then, samples were analyzed by flow cytometry (Beckman Coulter EPICS XL) for the percentage of apoptotic cells.
Flow cytometric analysis of the cell cycle
For cell cycle analysis, U2OS cells were treated with 0, 20 or 40 μM isoalantolactone for 24 h. After the treatment, the cells were collected, washed with PBS, and fixed with 70% ethanol at 4°C overnight. After washing twice with PBS, the cells were stained with a solution containing 50 μg/ml PI and 100 μg/ml RNase A for 30 min in the dark at room temperature. Cell cycle profiles were analyzed by flow cytometry (Beckman Coulter EPICS XL).
Flow cytometric analysis of reactive oxygen species (ROS) generation and mitochondrial membrane potential (MMP) in U2OS cells
To determine the ROS generation and MMP, U2OS cells were stained with 2′,7′-dichlorodihydrofluorescein diacetate (DCFH-DA) and Rhodamine 123 (Rho-123) respectively, as previously described (15). Briefly, U2OS cells were incubated with 0, 20 or 40 μM isoalantolactone for 24 h. After the treatment, the cells were harvested, washed with PBS, and then incubated with DCFH-DA (10 μM) or Rho-123 (5 μg/ml) in the dark for 30 min. After washing, the samples were analyzed for the fluorescence of DCF or Rhodamine 123 by flow cytometry.
Immunoblotting and immunoprecipitation
Following drug treatment, adherent and floating cells were collected and centrifuged. Nuclear and cytosolic proteins were extracted using a cytosolic and nuclear extraction kit (KeyGen) according to the manufacturer’s instructions. Total proteins were isolated as previously described (15). Briefly, U2OS cells were harvested, and the cell pellets were re-suspended in lysis buffer and lysed on ice for 30 min. After centrifugation for 15 min, the supernatants were collected, and the protein concentration was measured by NanoDrop 1,000 spectrophotometer (Thermo Scientific, USA). A total of 40 μg protein was electrophoresed on 10–15% SDS-PAGE and transferred to PVDF (Amersham Biosciences, Piscataway, NJ, USA) membranes. After being blocked with 5% non-fat milk for 2 h and washed with TBST, the membranes were incubated overnight at 4°C with DR5 (1:1,000), FADD (1:1,000), TRADD (1:1,000), RIP (1:1,000), NF-κBp65 (1:500), cyclin B1 (1:1,000), caspase-8 (1:500), caspase-3 (1:500), cleaved-PARP (1:1,000), Bcl-2 (1:1,000), Bax (1:1,000) and β-actin (1:500) antibodies, respectively. After washing, the blots were incubated with relevant secondary antibodies (1:5,000) for 1 h at room temperature. Signals were detected using the ECL-Plus chemiluminescence kit (Millipore Corporation) on X-ray film. All of the bands were quantified by densitometry using ImageJ software. For the immunoprecipitation (IP), the cells were incubated with different concentrations of isoalantolactone (0, 20 and 40 μM), and the lysates were incubated with anti-DR5 and protein G beads (YueKe Institute of Biotechnology, Shanghai, China) at 4°C overnight. Immunoblotting was carried out to analyze the association between DR5 and FADD. The beads were washed three times with IP buffer [50 mM Tris-HCl (pH 7.4), 150 mM NaCl and 1% NP40], boiled in 2X SDS loading buffer for 10 min, and then analyzed by immunoblotting.
Real-time PCR
The reverse transcription (RT) reaction was performed using the PrimeScript™ RT Master Mix (Takara) in a final volume of 10 μl containing 0.5 μg total RNA, 2 μl 5X PrimeScript buffer, 0.5 μl Oligo(dT) primer, 0.5 μl PrimeScript RT enzyme and RNase-free water. The RT reaction was performed at 37°C for 15 min and then terminated by heating at 85°C for 5 sec. Real-time quantitative PCR using SYBR®-Green (Roche) was performed with LightCycler 480 Real-Time PCR System. Thermal protocol was as follows: 95°C for 5 min plus 40 cycles at 95°C for 30 sec, 55°C for 30 sec and 72°C for 45 sec, with final elongation of 10 min at 72°C. The cyclin B1 primers were forward, 5′-GGC CAA AAT GCC TAT GAA GA-3′ and reverse, 5′-AGA TGT TTC CAT TGG GCT TG-3′. The GAPDH primers were forward, 5′-CGG AGT CA A CGG ATT TGG TCG TAT-3′ and reverse, 5′-AGC CTT CTC CAT GGT GGT GAA GAC-3′. Human GAPDH was measured as the internal control. Data analyses were performed according to the 2−ΔΔCt method.
Statistical analysis of the data
The results are expressed as means ± standard error of the mean (SEM) and were statistically compared with the control group or within groups using one-way ANOVA followed by Tukey’s multiple comparison test. The level of statistical significance was regarded as P<0.05. All of the experiments were repeated at least three times.
Results
Isoalantolactone inhibits proliferation of U2OS cells
In the present study, we screened natural compounds and their derivatives against human osteosarcoma U2OS, MG-63 and Saos-2 cells to identify putative therapeutic compounds. Isoalantolactone (chemical structure shown in Fig. 1A) exhibited a significant antiproliferative effect on osteosarcoma cell lines in a dose-dependent manner. The IC50 values of isoalantolactone against U2OS, MG-63 and Saos-2 cells were 40, 50 and 40 μM, respectively (Fig. 1B). The cytotoxic effect of isoalantolactone was also assessed by observing morphological changes in the cells under phase-contrast microscopy. As shown in Fig. 1C, the control cells adhered to the well and displayed normal cell morphology. After isoalantolactone treatment, the cells displayed significant morphological changes, and acquired a round and shrunken shape, an increase in floating cells and a reduction in the total number of cells in a time-dependent manner. The data are consistent with the results of the cell proliferation inhibition studies, confirming that isoalantolactone inhibited the proliferation of osteosarcoma cells.
Isoalantolactone induces S and mainly G2/M phase cell cycle arrest in U2OS cells
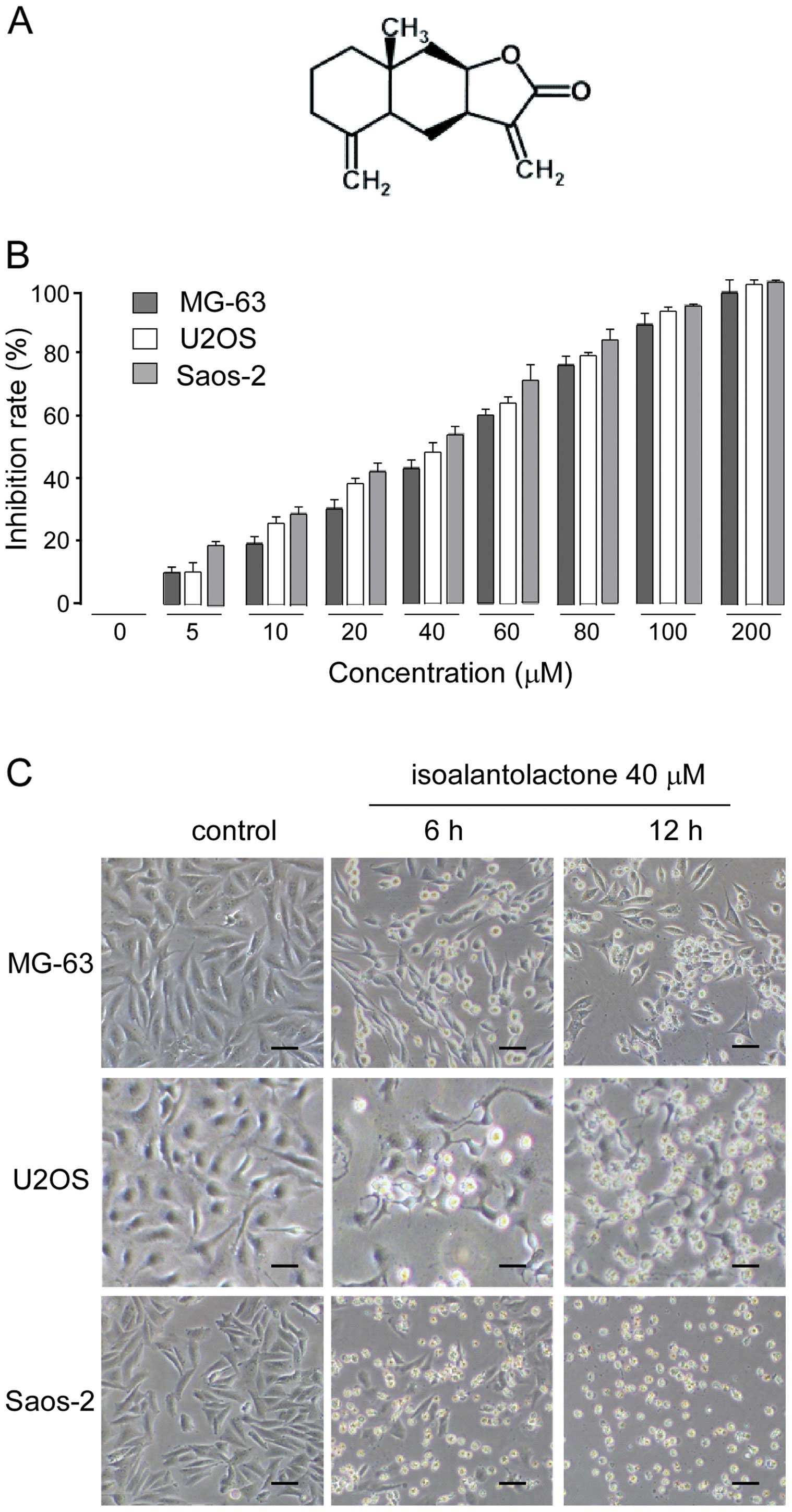
Cell cycle regulation and apoptosis are two major regulatory mechanisms for cell growth. When specific checkpoints during the cell cycle are arrested, apoptotic cell death occurs (16–20). To gain further insight into the mechanisms underlying the cytotoxic effects of isoalantolactone on U2OS cells, we next investigated the effect of this compound on the cell cycle phase profile. As shown in Fig. 2A, the percentages of cells that accumulated in the S phase were 34.8, 40.1 and 48%; in the G2/M phase, 10.1, 25.7 and 29.3%, following treatment with 0, 20 and 40 μM of isoalantolactone for 24 h, respectively. These results indicated that isoalantolactone induced cell cycle arrest at the S and mainly the G2/M phase.
To elucidate the molecular mechanism underlying G2/M phase arrest, we investigated the expression of cyclin B1, a key protein involved in the G2/M phase. U2OS cells were treated with different concentrations of isoalantolactone (0, 20 or 40 μM) for 24 h or with 40 μM isoalantolactone for 0, 12 or 24 h, respectively. The expression of cyclin B1 was assessed by western blotting and real-time PCR. The results demonstrated that isoalantolactone decreased the cyclin B1 expression at the mRNA and protein levels in a dose- and time-dependent manner (Fig. 2C–E).
Isoalantolactone induces apoptosis in U2OS cells
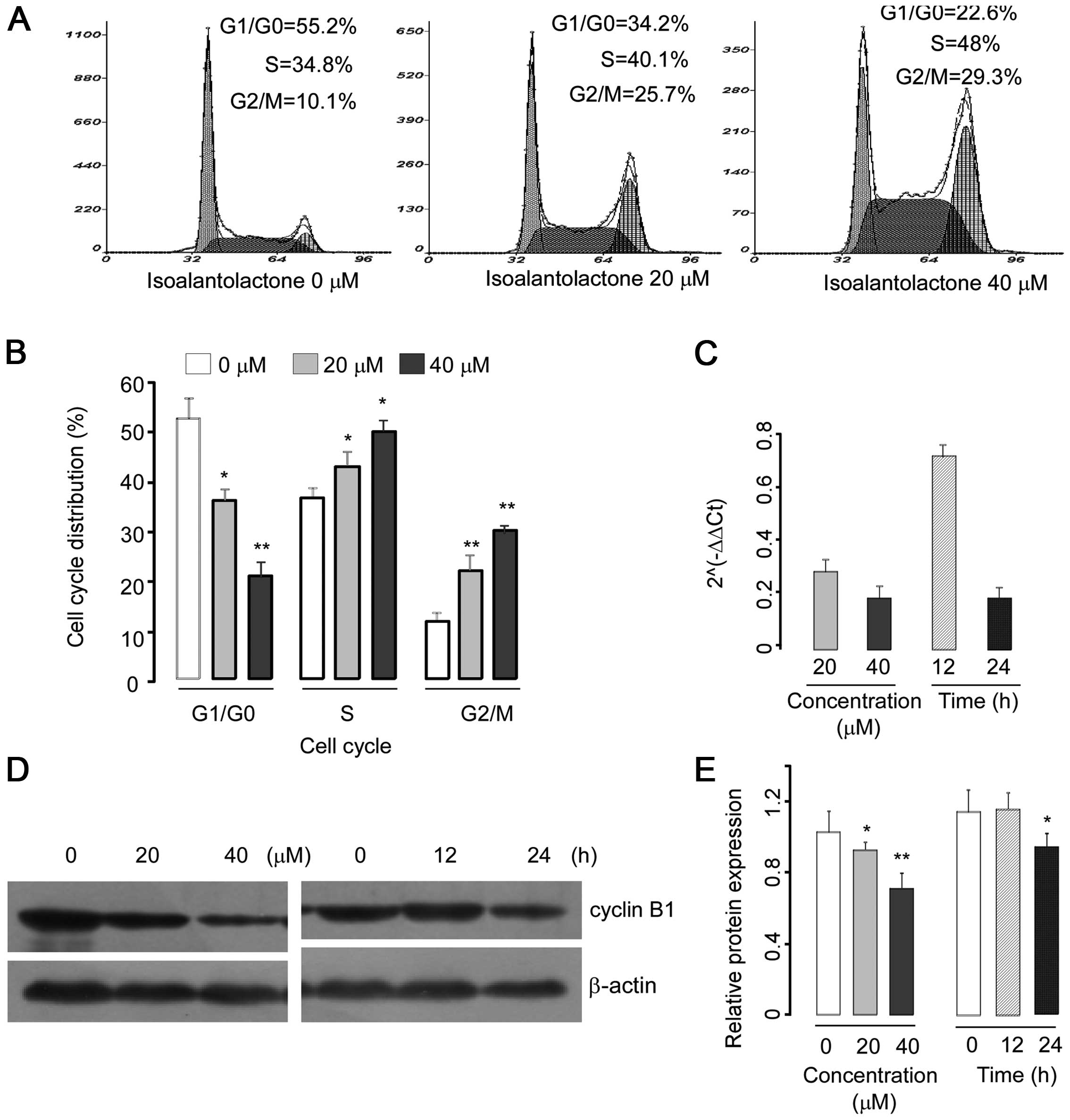
To confirm the nature of isoalantolactone-induced cell death, the treated cells were stained with Annexin V-FITC and PI, and the percentages of apoptotic and necrotic cells were determined by flow cytometry. Our results demonstrated that isoalantolactone triggered apoptosis in a dose-dependent manner (Fig. 3). However, isoalantolactone-induced apoptosis was markedly abrogated when the cells were pretreated with N-acetylcysteine (NAC), a specific ROS inhibitor or Z-VAD-FMK, a caspase inhibitor, suggesting that the apoptosis-inducing effect of isoalantolactone in osteosarcoma cells was mediated by reactive oxygen species and the caspases.
DR5/FADD/caspase-8 pathway is associated with the isoalantolactone- induced apoptosis in U2OS cells
Death receptor 5 (DR5) signals apoptosis through Fas-associating protein with death domain (FADD) and caspase-8 (21). To further explore the mechanism underlying isoalantolactone-induced apoptosis, we examined the effect of isoalantolactone on the death receptor pathways by measuring levels of DR5 and FADD and caspase-8 using western blotting. The data revealed that isoalantolactone increased the expression of DR5, FADD and induced the cleavage of caspase-8 in a dose- and time-dependent manner (Fig. 4A–C). The results suggest that the DR5/FADD/caspase-8 extrinsic pathways are associated with isoalantolactone-induced apoptosis in osteosarcoma U2OS cells. We next evaluated the physical interaction between DR5 and FADD in U2OS cells using Co-IP assay. The results showed that in the control group DR5 and FADD had almost no combination, while after treatment with isoalantolactone the combination between DR5 and FADD was significantly increased (Fig. 4D).
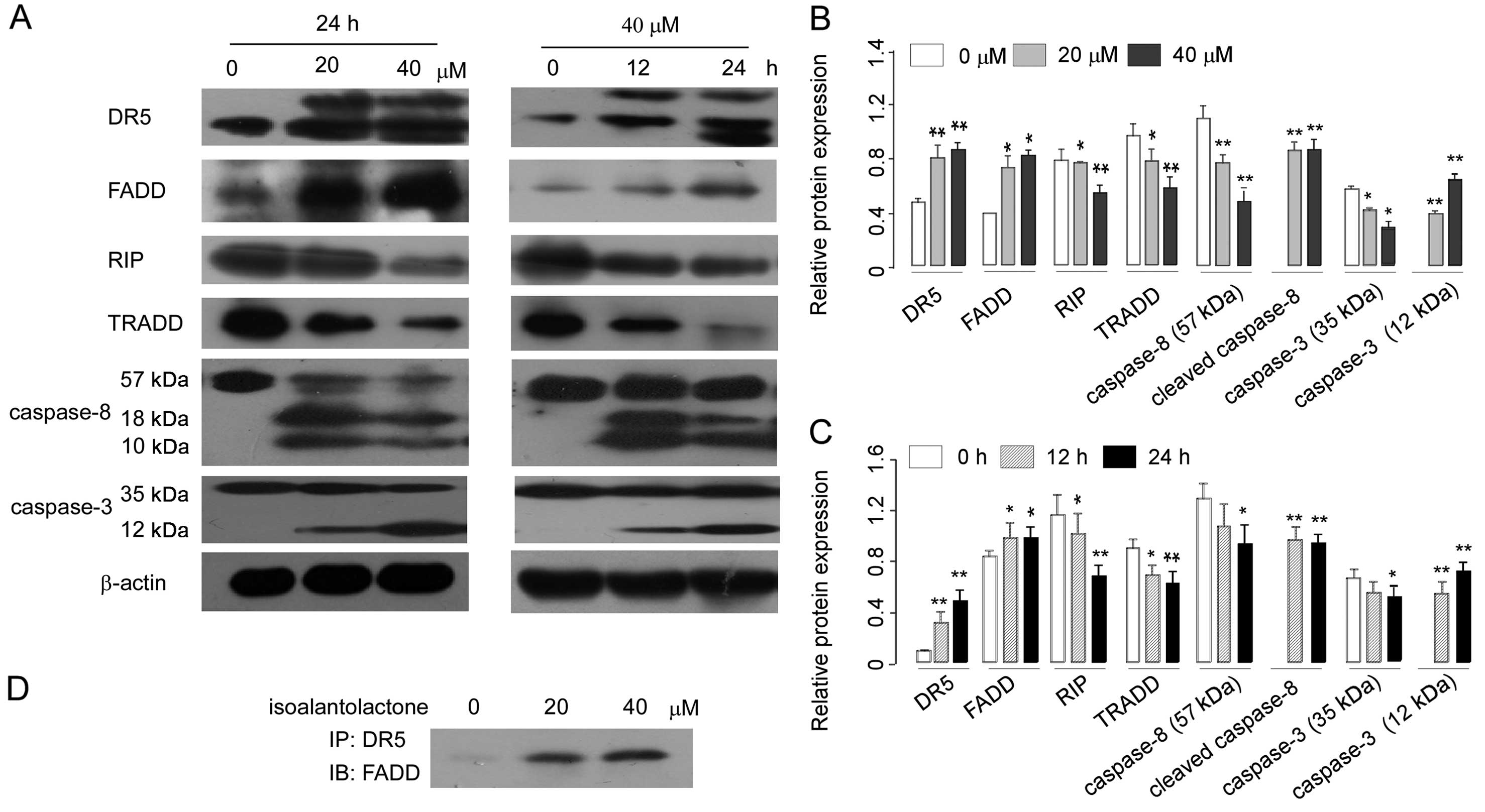
Many sesquiterpene lactone compounds have been reported to induce apoptosis through inhibition of NF-κB activation in various cancer cell lines (22,23), and previous research suggests that DR5 can use a TRADD-dependent pathway to activate NF-κB, which is independent of their ability to induce apoptosis (24). Here, we aimed to ascertain whether the apoptotic effect of isoalantolactone in U2OS cells is associated with NF-κB. For this, levels of TNF-receptor-associated death domain protein (TRADD), receptor-interacting protein (RIP), and nuclear protein NF-κBp65 were assessed in the U2OS cells. Western blot analysis revealed that treatment with isoalantolactone decreased the levels of TRADD and RIP (Fig. 4A–C) and decreased the levels of nuclear NF-κBp65 in a dose-dependent manner (Fig. 6). Further studies are required to examine the association of these proteins after treatment of isoalantolactone.
Effect of isoalantolactone on ROS generation and mitochondrial membrane potential
Recent evidence suggests that ROS serve as messengers for normal signal transductions at lower concentrations; at higher concentrations, however, they become toxic and induce cell death through various signaling pathways (25). As cancer cells contain a higher level of ROS than normal cells, they can be easily poisoned by the phytochemical targeting of ROS metabolism (25,26). Many studies have reported that isoalantolactone-induced apoptosis is linked with ROS generation (5,6). Here, we hypothesized that the effect of isoalantolactone on U2OS cells is associated with ROS. Therefore, we analyzed the level of ROS by flow cytometry. The levels of ROS in the cells treated with 0, 20 and 40 μM isoalantolactone for 24 h were 64.69, 82.54 and 94.58%. Our results demonstrated that isoalantolactone treatment significantly increased ROS generation in the U2OS cells in a dose-dependent manner (Fig. 5A and B).
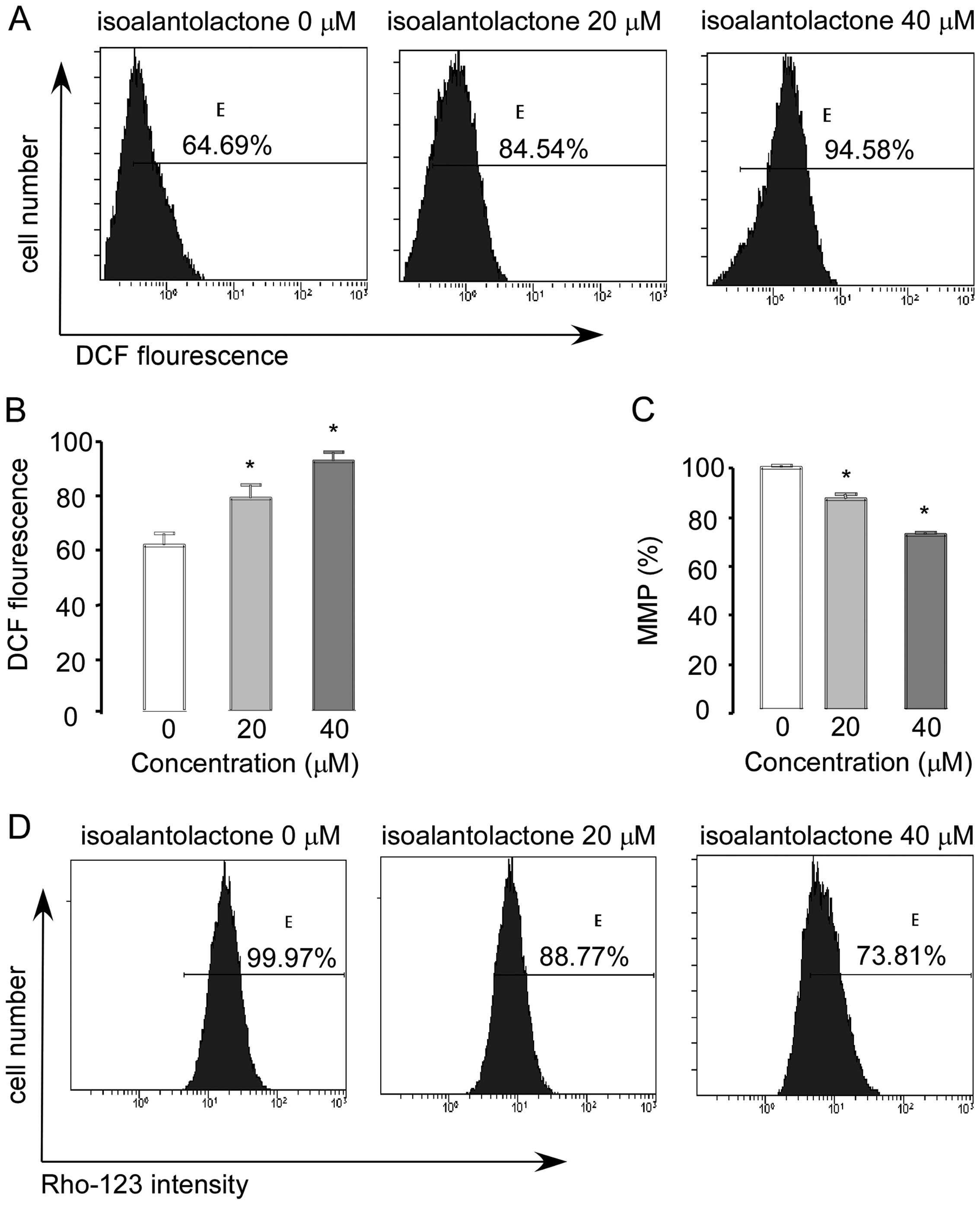
Next, we determined the effect of isoalantolactone on mitochondrial membrane potential in U2OS cells. As shown in Fig. 5C and D, the levels of MMP in the cells treated with 0, 20 and 40 μM isoalantolactone for 24 h were 99.97, 88.77 and 73.81%; Rho123 was significantly dissipated in U2OS cells in a dose-dependent manner (Fig. 5C and D). These findings clearly indicate that isoalantolactone-induced apoptosis is associated with ROS generation and MMP disruption.
Isoalantolactone modulates Bcl-2 family proteins
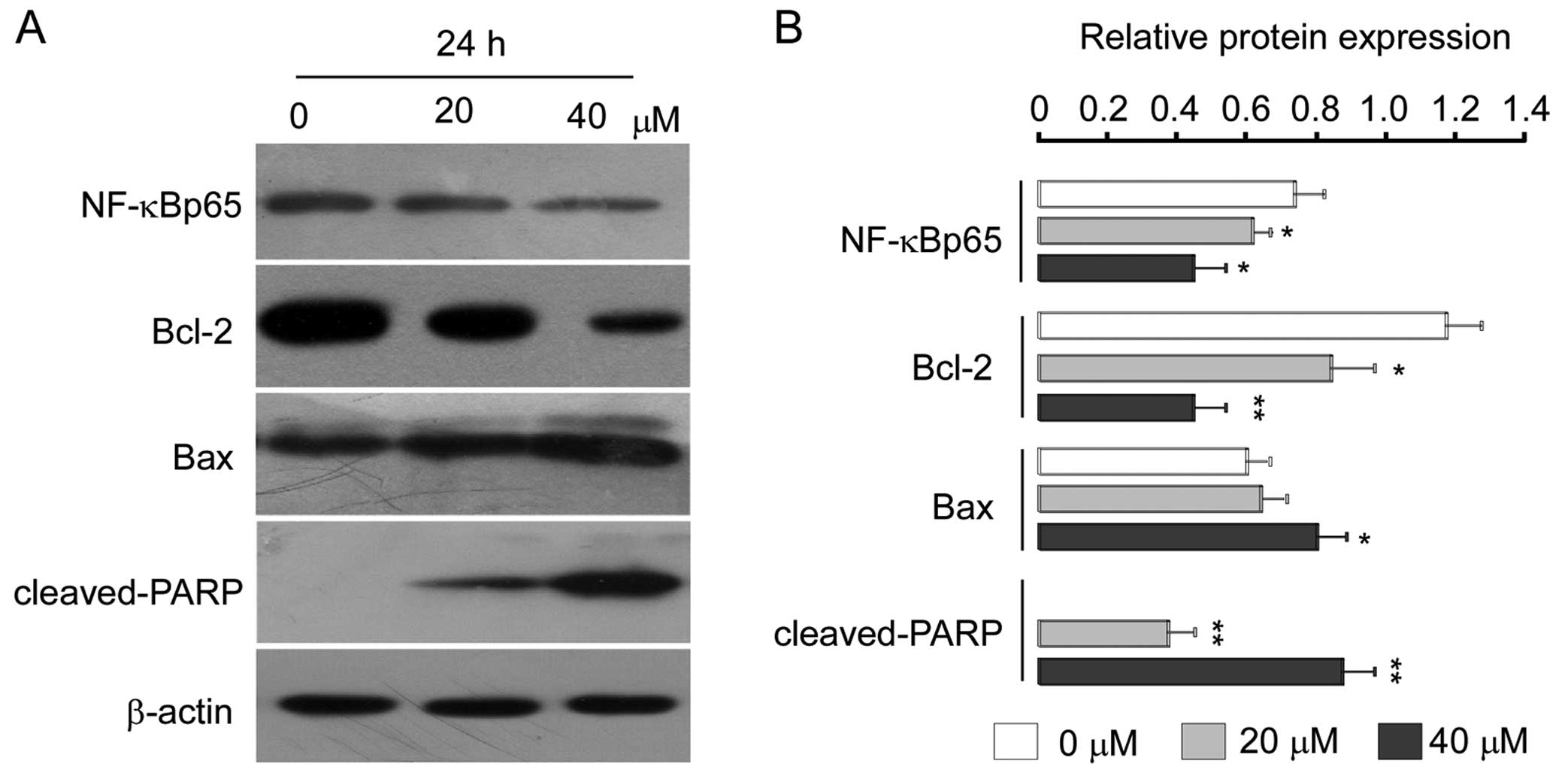
To further characterize this cell-specific apoptotic effect of isoalantolactone in osteosarcoma U2OS cells, we analyzed the levels of Bcl-2 family proteins after treatment with isoalantolactone (0, 20 and 40 μM) in a dose-dependent manner. We observed that expression of anti-apoptotic Bcl-2 was downregulated, whereas the expression level of pro-apoptotic Bax was markedly upregulated (Fig. 6). To examine whether regulation of caspase-3 is necessary for isoalantolactone-induced apoptosis, we investigated the expression level of caspase-3 after treatment with 0, 20 and 40 μM isoalantolactone for 24 h. As shown in Fig. 4A, isoalantolactone treatment was found to result in a significant increase in cleaved caspase-3, and its downstream target, PARP (Fig. 6) in a dose-dependent manner which is indicative of the induction of apoptosis.
Discussion
Osteosarcoma is one of the most common primary malignant tumors. Although neoadjuvant chemotherapy has improved the overall survival of patients over the last several years, the metastatic and toxic effects of chemotherapy still remain a major drawback in the treatment of osteosarcoma patients. The goal of the present study was to identify novel anti-osteosarcoma agents from natural herbal compounds. Isoalantolactone did not exert any acute or chronic cytotoxicity in liver and kidney, and sesquiterpene lactones have drawn considerable attention in pharmacological research due to their anti-neoplastic, anti-parasitic, anti-inflammatory and antitumor activities (27–29). However, its effects on osteosarcoma remained unexplored. In the present study, we found that isoalantolactone strongly inhibited the proliferation of osteosarcoma cells in a dose-dependent manner. Therefore, we selected isoalantolactone compound to study the molecular mechanisms of its antiproliferative effect against osteosarcoma U2OS cells.
Isoalantolactone has been reported to induce cell death in several human cancer cell lines, such as PANC-1 cells (6), human prostate cancer cells (8), and human gastric adenocarcinoma SGC-7901 cells (9) via cell cycle arrest and induction of apoptosis. In line with previous reports, we found that isoalantolactone arrested the cell cycle at the S and mainly the G2/M phase in U2OS cells. Cyclin B1, the key protein in the regulation of G2/M transition, forms a complex with cdc2 which is crucial in the transition from the G2 to the M phase (30). Our results demonstrated that isoalantolactone downregulated the expression of cyclin B1 at the mRNA and protein levels in a dose- and time-dependent manner. These data are in agreement with those of a previous study (31), suggesting that decreased expression of cyclin B1 may be a molecular mechanism through which isoalantolactone induces S and mainly G2/M phase arrest.
DR5 is a member of the tumor necrosis factor receptor family, which plays an important role in the mediation of extrinsic pathways. These receptors possess a cytoplasmic region called the death domain (DD) (32,33). Overexpression of death domain-containing receptors can induce apoptosis in a ligand-independent manner (34,35). Transient transfection of a full-length DR5 construct was found to induce rapid apoptosis (24). Bodmer et al (21) reported that DR5 can recruit FADD through its death effector domain, and FADD binds to caspase-8. Caspase-8 is the most proximal caspase in the cascade of caspases, the activation of which eventually leads to cell death (36). Overall, these studies suggest that activation of the DR5/FADD/caspase-8 extrinsic pathways may have therapeutic value for cancer patients. In the present study, we found that isoalantolactone treatment increased the expression of DR5 and FADD, and induced the binding of FADD to DR5. It also decreased the expression of pro-caspase-8 and increased the cleavage of caspase-8 in U2OS cells. The data clearly demonstrated that isoalantolactone-induced apoptosis is associated with the activation of DR5/FADD/caspase-8 extrinsic pathways. To the best of our knowledge, this is the first report indicating that isoalantolactone- induced apoptosis is associated with DR5/FADD/caspase-8 extrinsic pathways.
Death domains are also found in TRADD and RIP, two cytoplasmic adaptor proteins implicated in the mediation of apoptosis by the death domain-containing receptors (37,38). Overexpression studies have demonstrated that TRADD is required for recruitment of several important molecules, including RIP, which are indispensable for the activation of NF-κB and cell survival (38,39). NF-κB is a transcription factor, which can promote cell survival and proliferation. The constitutive activation of NF-κB is associated with numerous types of cancers (40–42). In the present study, we examined the expression of nuclear protein NF-κBp65 and its regulatory proteins TRADD and RIP. Our data indicated that isoalantolactone inhibits NF-κB activation, and whether this is associated with TRADD and RIP, more research needs to be performed to find the link between these proteins after treatment of isoalantolactone.
Mitochondria play a critical role in anticancer drug-induced apoptosis (43). The Bcl-2 family proteins, which play a pivotal role in the intrinsic apoptosis pathways, include both anti-apoptotic and pro-apoptotic proteins such as Bcl-2 and Bax, respectively. The balance between these two classes of proteins is critical for determining whether a cell undergoes apoptosis or not (44–46). ROS generation can inhibit the anti-apoptotic protein Bcl-2 and activate the pro-apoptotic protein Bax to the outer mitochondrial membrane (47,48). In the present study, we found that isoalantolactone increased ROS generation, and the ROS inhibitor, NAC, markedly reduced isoalantolactone-induced apoptosis in the U2OS cells. Our results revealed that isoalantolactone-induced apoptosis in U2OS cells was ROS-dependent. Moreover, flow cytometric analysis of MMP showed that isoalantolactone led to dissipation of mitochondrial membrane potential, and western blot analysis of apoptosis-related proteins in U2OS cells revealed that isoalantolactone downregulated the expression of Bcl-2 and upregulated the expression of Bax, activated caspase-3 and its downstream substrate PARP. Therefore, we conclude that isoalantolactone promotes opening of the mitochondrial PTP by increasing the Bax/Bcl-2 ratio and the isoalantolactone-induced apoptosis is associated with intrinsic pathways.
In conclusion, our data provide evidence that isoalantolactone inhibits the growth of human osteosarcoma cells by inducing apoptosis. The present study is the first to describe the role of ROS in the induction of apoptosis in osteosarcoma cells. In addition, in the present study, we also found that isoalantolactone upregulated DR5, FADD and cleaved caspase-8, increased the interaction between DR5 and FADD, and inhibited the expression of nuclear NF-κBp65, which, to our knowledge, is the first observation that isoalantolactone upregulates expression of these proteins and inhibits NF-κBp65 in osteosarcoma U2OS cells. The pathway that we have described herein is novel and has not been previously elucidated. Furthermore, isoalantolactone induced apoptosis in U2OS cells by modulating mitochondrial Bcl-2 family proteins and caspase-3. These findings support a prominent insight into how isoalantolactone exerts its cytotoxic effect against osteosarcoma cells. Thus, isoalantolactone may become a potential lead compound for future development of anti-osteosarcoma therapy. Further investigation is needed to validate the contribution of isoalantolactone to tumor therapy in vivo.
References
|
Broadhead ML, Clark JC, Myers DE, Dass CR and Choong PF: The molecular pathogenesis of osteosarcoma: a review. Sarcoma. 2011:9592482011. View Article : Google Scholar : PubMed/NCBI | |
|
Mirabello L, Troisi RJ and Savage SA: Osteosarcoma incidence and survival rates from 1973 to 2004: data from the Surveillance, Epidemiology, and End Results Program. Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Marina N, Gebhardt M, Teot L and Gorlick R: Biology and therapeutic advances for pediatric osteosarcoma. Oncologist. 9:422–441. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Kempf-Bielack B, Bielack SS, Jürgens H, et al: Osteosarcoma relapse after combined modality therapy: an analysis of unselected patients in the Cooperative Osteosarcoma Study Group (COSS). J Clin Oncol. 23:559–568. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Wu M, Zhang H, Hu J, et al: Isoalantolactone inhibits UM-SCC-10A cell growth via cell cycle arrest and apoptosis induction. PLoS One. 8:e760002013. View Article : Google Scholar : PubMed/NCBI | |
|
Khan M, Ding C, Rasul A, et al: Isoalantolactone induces reactive oxygen species mediated apoptosis in pancreatic carcinoma PANC-1 cells. Int J Biol Sci. 8:533–547. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Konishi T, Shimada Y, Nagao T, Okabe H and Konoshima T: Antiproliferative sesquiterpene lactones from the roots of Inula helenium. Biol Pharm Bull. 25:1370–1372. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Rasul A, Khan M, Yu B, et al: Isoalantolactone, a sesquiterpene lactone, induces apoptosis in SGC-7901 cells via mitochondrial and phosphatidylinositol 3-kinase/Akt signaling pathways. Arch Pharm Res. 36:1262–1269. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Rasul A, Di J, Millimouno FM, et al: Reactive oxygen species mediate isoalantolactone-induced apoptosis in human prostate cancer cells. Molecules. 18:9382–9396. 2013. View Article : Google Scholar : PubMed/NCBI | |
|
Seo JY, Park J, Kim HJ, et al: Isoalantolactone from Inula helenium caused Nrf2-mediated induction of detoxifying enzymes. J Med Food. 12:1038–1045. 2009. | |
|
Fulda S and Debatin KM: Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene. 25:4798–4811. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Mühlethaler-Mottet A, Bourloud KB, Auderset K, Joseph JM and Gross N: Drug-mediated sensitization to TRAIL-induced apoptosis in caspase-8-complemented neuroblastoma cells proceeds via activation of intrinsic and extrinsic pathways and caspase-dependent cleavage of XIAP, Bcl-xL and RIP. Oncogene. 23:5415–5425. 2004. | |
|
Kamachi M, Le TM, Kim SJ, Geiger ME, Anderson P and Utz PJ: Human autoimmune sera as molecular probes for the identification of an autoantigen kinase signaling pathway. J Exp Med. 196:1213–1225. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Tang D, Lahti JM and Kidd VJ: Caspase-8 activation and bid cleavage contribute to MCF7 cellular execution in a caspase-3-dependent manner during staurosporine-mediated apoptosis. J Biol Chem. 275:9303–9307. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Khan M, Yi F, Rasul A, et al: Alantolactone induces apoptosis in glioblastoma cells via GSH depletion, ROS generation, and mitochondrial dysfunction. IUBMB Life. 64:783–794. 2012. View Article : Google Scholar : PubMed/NCBI | |
|
Lu YJ, Yang SH, Chien CM, et al: Induction of G2/M phase arrest and apoptosis by a novel enediyne derivative, THDB, in chronic myeloid leukemia (HL-60) cells. Toxicol In Vitro. 21:90–98. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Torres K and Horwitz SB: Mechanisms of Taxol-induced cell death are concentration dependent. Cancer Res. 58:3620–3626. 1998.PubMed/NCBI | |
|
Gamet-Payrastre L, Li P, Lumeau S, et al: Sulforaphane, a naturally occurring isothiocyanate, induces cell cycle arrest and apoptosis in HT29 human colon cancer cells. Cancer Res. 60:1426–1433. 2000.PubMed/NCBI | |
|
Murray AW: Recycling the cell cycle: cyclins revisited. Cell. 116:221–234. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
Orren DK, Petersen LN and Bohr VA: Persistent DNA damage inhibits S-phase and G2 progression, and results in apoptosis. Mol Biol Cell. 8:1129–1142. 1997. View Article : Google Scholar : PubMed/NCBI | |
|
Bodmer JL, Holler N, Reynard S, et al: TRAIL receptor-2 signals apoptosis through FADD and caspase-8. Nat Cell Biol. 2:241–243. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Ghantous A, Gali-Muhtasib H, Vuorela H, Saliba NA and Darwiche N: What made sesquiterpene lactones reach cancer clinical trials? Drug Discov Today. 15:668–678. 2010. View Article : Google Scholar : PubMed/NCBI | |
|
Lyss G, Knorre A, Schmidt TJ, Pahl HL and Merfort I: The anti-inflammatory sesquiterpene lactone helenalin inhibits the transcription factor NF-κB by directly targeting p65. J Biol Chem. 273:33508–33516. 1998. | |
|
Chaudhary PM, Eby M, Jasmin A, Bookwalter A, Murray J and Hood L: Death receptor 5, a new member of the TNFR family, and DR4 induce FADD-dependent apoptosis and activate the NF-κB pathway. Immunity. 7:821–830. 1997.PubMed/NCBI | |
|
Juan ME, Wenzel U, Daniel H and Planas JM: Resveratrol induces apoptosis through ROS-dependent mitochondria pathway in HT-29 human colorectal carcinoma cells. J Agric Food Chem. 56:4813–4818. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Schumacker PT: Reactive oxygen species in cancer cells: live by the sword, die by the sword. Cancer Cell. 10:175–176. 2006. View Article : Google Scholar : PubMed/NCBI | |
|
Gu JQ, Gills JJ, Park EJ, et al: Sesquiterpenoids from Tithonia diversifolia with potential cancer chemopreventive activity. J Nat Prod. 65:532–536. 2002.PubMed/NCBI | |
|
Koch E, Klaas CA, Rüngeler P, et al: Inhibition of inflammatory cytokine production and lymphocyte proliferation by structurally different sesquiterpene lactones correlates with their effect on activation of NF-κB. Biochem Pharmacol. 62:795–801. 2001.PubMed/NCBI | |
|
Nam NH: Naturally occurring NF-κB inhibitors. Mini Rev Med Chem. 6:945–951. 2006. | |
|
Lee SM, Kwon JI, Choi YH, Eom HS and Chi GY: Induction of G2/M arrest and apoptosis by water extract of Strychni Semen in human gastric carcinoma AGS cells. Phytother Res. 22:752–758. 2008. View Article : Google Scholar : PubMed/NCBI | |
|
Rasul A, Yu B, Zhong L, Khan M, Yang H and Ma T: Cytotoxic effect of evodiamine in SGC-7901 human gastric adenocarcinoma cells via simultaneous induction of apoptosis and autophagy. Oncol Rep. 27:1481–1487. 2012.PubMed/NCBI | |
|
Scott FL, Stec B, Pop C, et al: The Fas-FADD death domain complex structure unravels signalling by receptor clustering. Nature. 457:1019–1022. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Guicciardi ME and Gores GJ: Life and death by death receptors. FASEB J. 23:1625–1637. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Gaeta ML, Johnson DR, Kluger MS and Pober JS: The death domain of tumor necrosis factor receptor 1 is necessary but not sufficient for Golgi retention of the receptor and mediates receptor desensitization. Lab Invest. 80:1185–1194. 2000. View Article : Google Scholar | |
|
Sheikh MS and Fornace AJ Jr: Death and decoy receptors and p53-mediated apoptosis. Leukemia. 14:1509–1513. 2000. View Article : Google Scholar : PubMed/NCBI | |
|
Ramaswamy M, Efimova EV, Martinez O, Mulherkar NU, Singh SP and Prabhakar BS: IG20 (MADD splice variant-5), a proapoptotic protein, interacts with DR4/DR5 and enhances TRAIL-induced apoptosis by increasing recruitment of FADD and caspase-8 to the DISC. Oncogene. 23:6083–6094. 2004. View Article : Google Scholar : PubMed/NCBI | |
|
MacEwan DJ: TNF ligands and receptors - a matter of life and death. Br J Pharmacol. 135:855–875. 2002. View Article : Google Scholar : PubMed/NCBI | |
|
Stanger BZ, Leder P, Lee TH, Kim E and Seed B: RIP: a novel protein containing a death domain that interacts with Fas/APO-1 (CD95) in yeast and causes cell death. Cell. 81:513–523. 1995. View Article : Google Scholar : PubMed/NCBI | |
|
Hsu H, Shu HB, Pan MG and Goeddel DV: TRADD-TRAF2 and TRADD-FADD interactions define two distinct TNF receptor 1 signal transduction pathways. Cell. 84:299–308. 1996. View Article : Google Scholar : PubMed/NCBI | |
|
Bharti AC, Donato N, Singh S and Aggarwal BB: Curcumin (diferuloylmethane) down-regulates the constitutive activation of nuclear factor-κB and IκBα kinase in human multiple myeloma cells, leading to suppression of proliferation and induction of apoptosis. Blood. 101:1053–1062. 2003.PubMed/NCBI | |
|
Li Q, Yu YY, Zhu ZG, et al: Effect of NF-κB constitutive activation on proliferation and apoptosis of gastric cancer cell lines. European surgical research. Eur Surg Res. 37:105–110. 2005. | |
|
Shen HM and Tergaonkar V: NFκB signaling in carcinogenesis and as a potential molecular target for cancer therapy. Apoptosis. 14:348–363. 2009. | |
|
Jeong SY and Seol DW: The role of mitochondria in apoptosis. BMB Rep. 41:11–22. 2008. View Article : Google Scholar | |
|
Danial NN: BCL-2 family proteins: critical checkpoints of apoptotic cell death. Clin Cancer Res. 13:7254–7263. 2007. View Article : Google Scholar : PubMed/NCBI | |
|
Frenzel A, Grespi F, Chmelewskij W and Villunger A: Bcl2 family proteins in carcinogenesis and the treatment of cancer. Apoptosis. 14:584–596. 2009. View Article : Google Scholar : PubMed/NCBI | |
|
Wei W, Huang H, Zhao S, et al: Alantolactone induces apoptosis in chronic myelogenous leukemia sensitive or resistant to imatinib through NF-κB inhibition and Bcr/Abl protein deletion. Apoptosis. 18:1060–1070. 2013. | |
|
Felty Q, Xiong WC, Sun D, et al: Estrogen-induced mitochondrial reactive oxygen species as signal-transducing messengers. Biochemistry. 44:6900–6909. 2005. View Article : Google Scholar : PubMed/NCBI | |
|
Jabs T: Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochem Pharmacol. 57:231–245. 1999. View Article : Google Scholar : PubMed/NCBI |