Introduction
Hepatocellular carcinoma (HCC) is one of the most
common malignant tumors in China (1). Although surgical resection remains the
optimum option for HCC, many patients are unresectable due to the
tumor size, metastasis, hepatic functional reserve and/or portal
hypertension. Trans-arterial embolization (TAE), percutaneous
injection of ethanol and radiofrequency ablation have also been
used in the treatment of unresectable patients. However, these
methods only target the local tumors and the high recurrence and
metastatic rate limit the outcome. In addition, HCC is not
sensitive to most chemotherapeutic drugs, such as paclitaxel,
doxorubicin, fuorouracil, cisplatin and mitomycin. Thus, new agents
that are safe and effective are to be identified for the treatment
of HCC.
Findings of recent studies have demonstrated that
many compounds from Traditional Chinese Medicine (TCM) are
effective in the treatment of malignant tumors, including HCC
(2–4). In a previous study, we found that
Ganzaoning granule, a Traditional Chinese Medicine formula, is able
to inhibit diethylnitrosamine-induced hepatocarcinogenesis in rat
(5,6). However, the active ingredients of
Ganzaoning exerting an anti-HCC effect have not been clarified.
Diosgenin, a steroidal saponin, is abundant in a variety of plants,
such as yam (Dioscorea villosa) which is a main drug in
Ganzaoning granule. Results of recent studies have shown that
diosgenin exerts anticancer effects against a wide variety of tumor
cells, including leukemic, gastric, colorectal, and breast cancer
(7–11). However, the anticancer effect of
diosgenin on HCC and the mechanisms has not been completely
elucidated.
Therefore, we investigated the inhibitory effect of
diosgenin on HCC, and the molecular mechanism of the antitumor
effect in this study. Our results showed that diosgenin reduced the
proliferation of Bel-7402, SMMC-7721 and HepG2 cells in a
dose-dependent manner. In addition, diosgenin exerted an
anti-proliferative effect in the three HCC cells by inducing G2/M
cell cycle arrest and apoptosis. Furthermore, diosgenin upregulated
p27 and p21 expression and activated the caspase cascade.
Diosgenin-induced p27 and p21 upregulation was independent of p53.
The results suggest that diosgenin potentially exerts
chemopreventive effects on the relevant cell cycle regulation and
death receptor apoptotic pathways.
Materials and methods
Cell culture and drug treatment
SMMC-7721, Bel-7402, HepG2 HCC cell lines were
cultured in DMEM medium (HyClone, Logan, UT, USA) supplemented with
10% fetal bovine serum (FBS) (Biowest, Nuaille, France). The cells
were cultured at 37°C with 5% CO2. Diosgenin, with a
purity of >98% was purchased from Shanghai Winherb Medical
Technology Co., Ltd. (Shanghai, China) and dissolved in
ethanol.
MTT assay
The cells were seeded in 96-well plates and treated
with diosgenin at concentrations of 0–40 μM for the indicated
time-points. After the exposure period, the media were removed.
Cell viability was measured using the MTT method as previously
described (12). The experiment was
performed in triplicate. The inhibitory rate was calculated as a
percentage using the formula: (1 −
ODdiosgenin/ODcontrol) ×100%.
Cell morphology was observed under an inverted
microscope and the images were obtained at amagnification of
×200.
Cell cycle and apoptosis assay
Flow cytometric analysis was performed to determine
the presence of cell cycle arrest and apoptotic cells. After
treatment with diosgenin for 24 h, the cells were collected by
trypsinisation and washed twice with PBS, fixed in ice-cold 80%
ethanol, and stored overnight at 4°C. For analysis, the cells were
washed with PBS twice, and suspended in 1 ml of cold propidium
iodide (PI) solution. The cells were then incubated on ice for 30
min in the dark and then analyzed using flow cytometry.
FITC-labeled Annexin V/PI staining was performed
according to the manufacturer’s instructions (Keygen, Nanjing,
China). Briefly, 1×106 cells/well were suspended in
buffer containing FITC-conjugated Annexin V/PI at appropriate
concentrations. The samples were analyzed by flow cytometry and
20,000 events from each sample were obtained to ensure adequate
data.
Quantitative RT-PCR
Total RNA was extracted from SMMC-7721 cells after
diosgenin treatment with TRIzol reagent (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer’s instructions. Total RNA (5 μg)
was reverse-transcribed into cDNA using a first-strand cDNA
synthesis kit (FSK-100) (Toyobo, Osaka, Japan). Amplification of
the cDNA was achieved in triplicate using a commercially available
SYBR-Green PCR Master mix (Toyobo). cDNA was amplified under the
following conditions: 95°C for 5 min for denaturation and subjected
to 40 cycles of 95°C for 10 sec, 60°C for 20 sec, and 72°C for 25
sec. The relative expression level of mRNA in each sample was
normalized to its β-actin content. The relative expression levels
of mRNA were calculated as 2−ΔΔCt.
Western blotting
SMMC-7721 cells were seeded in 6-well plates at a
density of 1×106 cells/well with 2 ml completed DMEM
medium. Following diosgenin treatment for the indicated times,
total protein was extracted as previously described (12,13).
The protein concentration was determined using the BCA method.
Equal quantities of proteins were separated by sodium dodecyl
sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred by electroblotting onto a nitrocellulose membrane. The
membrane was blocked with 5% BSA in TBST buffer (20 mM Tris-HCl, pH
7.4, 150 mM NaCl and 0.1% Tween-20) overnight at 4°C. The membrane
was incubated with specific primary antibodies for 2 h and a
secondary antibody for 1 h. The signal was visualized with an
enhanced chemiluminescence kit (ECL) (Thermo Scientific, San Jose,
CA, USA).
Statistical analysis
Data are presented as means ± SD. Statistical
significance was determined using SPSS 17.0 for Windows. The
Student’s t-test was used to compare means for two groups and
one-way ANOVA was performed for multiple comparisons followed by
the Newman-Keuls test for multiple comparisons. Differences were
considered significant when P<0.05.
Results
Diosgenin inhibits HCC cell
proliferation
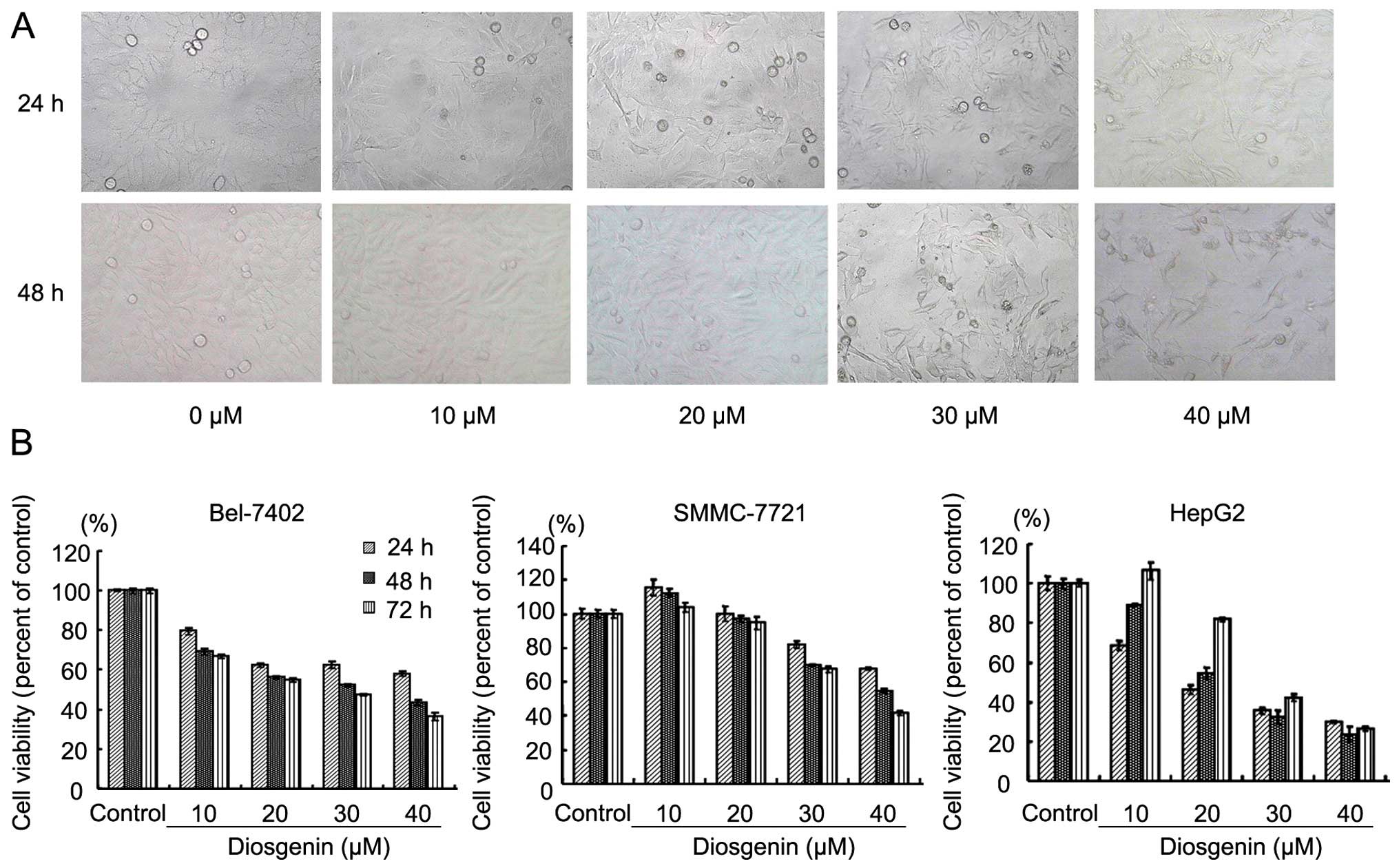
Diosgenin treatment for 24 and 48 h induced changes
in SMMC-7721 cell morphology, including cell shrinkage,
disappearance of tentacles and round-up shapes, indicating cell
damage (Fig. 1A). The effect of
diosgenin was concentration- and time-dependent. We examined the
inhibitory rate of diosgenin in Bel-7721, SMMC-7721 and HepG2 HCC
cells. The results showed that diosgenin treatment significantly
inhibited Bel-7721, SMMC-7721 and HepG2 cell proliferation in a
concentration-dependent manner (Fig.
1B).
Diosgenin induced G2/M cell cycle arrest
and apoptosis of HCC cells
To investigate the mechanisms of diosgenin-induced
growth repression, the flow cytometric analysis was performed.
Diosgenin treatment caused a concentration-dependent increase of
G2/M phase cell population in Bel-7721, SMMC-7721 and HepG2 HCC
cells, indicating diosgenin was able to arrest the cell cycle in
G2/M phase (Fig. 2). The proportion
of G2/M phase cells increased with concentration of diosgenin.
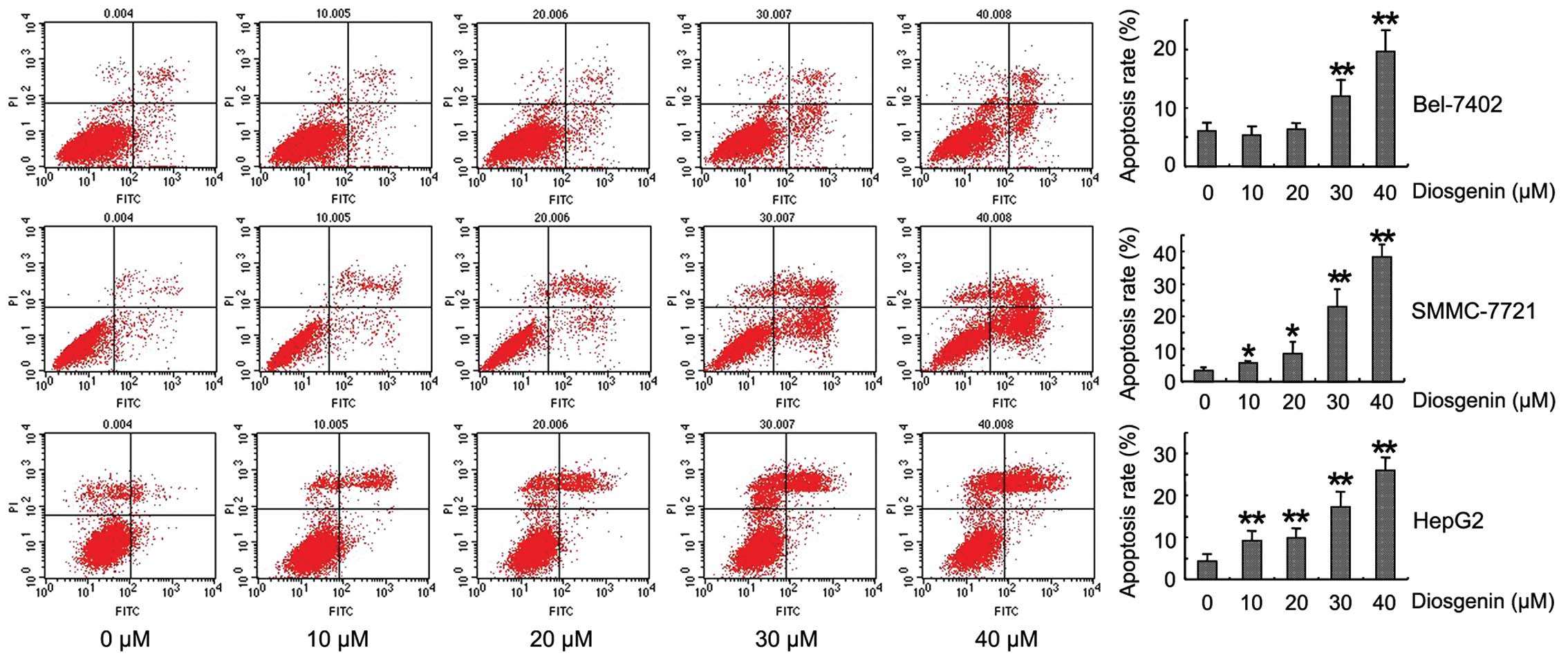
We also determined whether diosgenin-induced HCC
cell proliferation and inhibition involved apoptosis. Flow
cytometry using Annexin V-PI staining was performed. After
treatment with different concentrations of diosgenin for 24 h, the
proportions of apoptotic cells were markedly increased in Bel-7721,
SMMC-7721 and HepG2 HCC cells (Fig.
3), suggesting that diosgenin was able to induce the HCC cell
apoptosis. The effect of diosgenin on HCC cell apoptosis was
concentration-dependent.
Effect of diosgenin on cell cycle-related
proteins
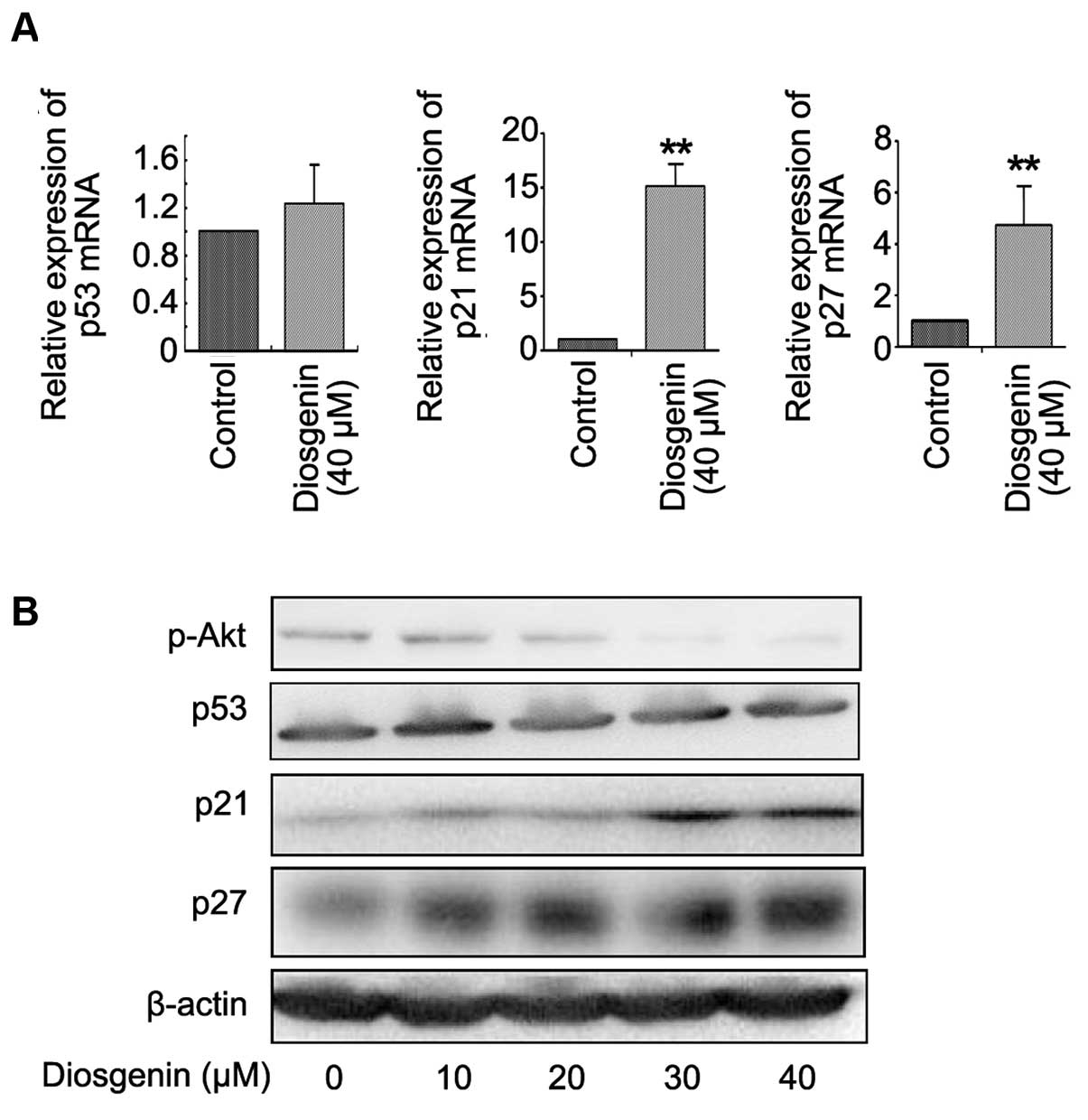
To investigate the mechanisms of diosgenin-induced
cell cycle arrest, we examined the expression of cell cycle-related
proteins by quantitative RT-PCR and western blotting. Diosgenin (40
μM) treatment for 24 h significantly upregulated the p21 and p27
mRNA levels in SMMC-7721 cells (Fig.
4A). Following treatment with diosgenin for 24 h, Akt
phosphorylation was significantly inhibited in SMMC-7721 cells in a
concentration-dependent manner (Fig.
4B). p21 and p27 protein levels were significantly upregulated
after diosgenin treatment in a concentration-dependent manner.
However, diosgenin treatment did not alter the expression of p53 in
the mRNA and protein levels (Fig. 4A
and B).
Effects of diosgenin on cell
apoptosis-related proteins
To determine the signaling pathway responsible for
diosgenin-induced apoptosis in HCC cells, the expression levels of
apoptosis-related proteins were examined subsequently. We first
examined whether diosgenin could alter the balance between
pro-apoptotic protein Bax and anti-apoptotic protein Bcl-2 in
SMMC-7721 cells. The results showed that diosgenin treatment did
not alter Bax and Bcl-2 levels (Fig.
5).
The expression levels of caspase-3, -8 and -9 were
also detected in diosgenin-treated SMMC-7721 cells. After various
concentrations of diosgenin treatment for 24 h, the expression of
caspase-3, -8 and -9 was markedly reduced in a
concentration-dependent manner (Fig.
5), whereas the cleaved caspase-3, -8 and -9 were obviously
increased.
Discussion
In the present study, we investigated the anti-HCC
effect of diosgenin in three HCC cell lines. The results
demonstrated that diosgenin exerted a strong growth inhibitory
activity against Bel-7402, SMMC-7721 and HepG2 human liver cancer
cells. Diosgenin induced G2/M cell cycle arrest and apoptosis in
these cells. Further study showed that the upregulation of cell
cycle-related proteins, p21 and p27, and activation of caspase
cascade may be involved in diosgenin-induced cell cycle arrest and
apoptosis.
Although the anticancer property of diosgenin has
been widely reported in many cancer cells, the anti-HCC effect and
the mechanisms involved in diosgenin have not been extensively
investigated. In the present study, the results showed that
diosgenin inhibited the proliferation of Bel-7402, SMMC-7721 and
HepG2 HCC cells. We also studied the mechanisms of diosgenin at
cellular and molecular levels. It is well recognized that
dysregulation of the cell cycle is a hallmark of tumor cells.
Numerous anticancer drugs play a therapeutic role by inducing cell
cycle arrest. In the present study, we showed that treatment with
diosgenin induced G2/M cell cycle arrest in Bel-7402, SMMC-7721 and
HepG2 cells in a concentration-dependent manner. Although previous
studies showed that diosgenin treatment caused cell cycle arrest in
G1 phase in osteosarcoma cells and C3A hepatoma cells (14,15),
diosgenin also induced G2/M cell cycle arrest in erythroleukemia
HEL cells and human leukemia K562 cells (9,16).
Those results suggest that the action of diosgenin on cell cycle
and its mechanism may be determined by the cell type.
The potent cyclin-dependent kinase inhibitors (CKI)
(17), p21 and p27, negatively
regulate multiple phases of the cell cycle progression (18). After binding to the Cyclin-Cdk
complexes, p21 and p27 inhibit their kinase activities and prevent
cell cycle progression (19). Since
the upregulation of p21 and p27 suppresses the proliferation of
many cancer cells by inducing cell cycle arrests, they are
recognized as important tumor suppressors (20,21).
Diosgenin treatment induced a significant increase of p21 and p27
in SMMC-7721 cells, suggesting that the regulation of p21 and p27
may be involved in diosgenin-induced cell cycle arrest. However,
p53 expression, which is conventionally considered as a regulator
of p21, was not altered by diosgenin, suggesting the
diosgenin-induced upregulation of p21 and p27 is not p53-dependent.
Phosphatidylinositol 3 kinase (PI3K)/Akt pathway is a vital
regulator of cell survival, proliferation and migration (22). Deregulation of the PI3K/Akt
signaling pathway is important in cancer development and has been
suggested as a therapeutic target for cancer (22–24).
The inhibition of PI3K/Akt pathway by its specific inhibitor
LY294002 is able to upregulate the expression of p21 and p27
(25) and lead to cell cycle arrest
(26). Diosgenin is known to
suppress Akt activation in various cell types (27–29).
Our data also showed that diosgenin treatment inhibited Akt
phosphorylation. These results suggest that the inactivation of the
PI3K/Akt pathway accompanied by an increased expression of p21 and
p27 is involved in diosgenin-induced cell cycle arrest in HCC
cells.
Cell growth is also regulated by apoptosis.
Diosgenin-induced G2/M cell cycle arrest provides an opportunity
for HCC cells to undergo apoptotic progression. In this study, flow
cytometry showed that treatment with diosgenin caused
concentration-dependent cell apoptosis. Since activation of the
caspase cascade and its downstream molecules turns on cell
apoptotic death progression, we first assessed the effect of
diosgenin on the caspase cascade, which mediates the death receptor
pathway. Western blotting showed that diosgenin decreased the total
caspase-3, -8 and -9 levels, but increased the cleaved caspase-3,
-8 and -9 levels in a concentration-dependent manner. The Bcl-2
family proteins are crucial for mitochondrial pathway-induced
apoptosis (30). Diosgenin
treatment for 24 h did not alter the expression of pro-apoptotic
protein Bax and anti-apoptotic protein Bcl-2. These results suggest
that diosgenin-induced apoptosis of HCC cells may be mediated by
the death receptor pathway, but not the mitochondrial pathway.
Although Kim et al (31)
reported that diosgenin induced apoptosis in HepG2 cells through
the generation of reactive oxygen species and mitochondrial
pathway, our results may indicate a diverse mechanism of
diosgenin.
In conclusion, our study demonstrated that diosgenin
inhibited HCC cell proliferation by inducing G2/M cell cycle arrest
and apoptosis. The inactivation of Akt, upregulation of p21 and p27
expression and activation of the caspase cascades were involved in
the anti-HCC effect of diosgenin. Therefore, our study may provide
evidence for the anti-HCC effect of diosgenin and elucidate the
underlying mechanisms of diosgenin.
Acknowledgements
This project was supported by grants from the
National Natural Science Foundation of China (no. 81202973) and the
E-Institutes of Shanghai Municipal Education Commission (no.
E-03008).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Paul A, Das S, Das J, et al:
Diarylheptanoid-myricanone isolated from ethanolic extract of
Myrica cerifera shows anticancer effects on HeLa and PC3 cell
lines: signalling pathway and drug-DNA interaction. J Integr Med.
11:405–415. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhai XF, Chen Z, Li B, et al: Traditional
herbal medicine in preventing recurrence after resection of small
hepatocellular carcinoma: a multicenter randomized controlled
trial. J Integr Med. 11:90–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhao L, Zhao AG, Zhao G, et al: Survival
benefit of traditional Chinese herbal medicine (a herbal formula
for invigorating spleen) in gastric cancer patients with peritoneal
metastasis. Evid Based Complement Alternat Med. 2014:6254932014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qian Y, Jin Y, Li Y and Ling C: Inhibitive
effect of 981208 agent on cell proliferating of
hepatocarcinogenesis induced by diethyl nitros-amine in rats. Acad
J Sec Mil Med Univ. 22:558–560. 2001.
|
|
6
|
Qian Y, Jin Y, Li Y, Li B and Ling C:
Experimental study of mechanism of Ganfujian granule inhibiting
hepatocarcinogenesis induced with Diethylnitrosamine in rats. J
Zhejiang Chin Med Univ. 27:56–58. 2003.
|
|
7
|
Mao ZJ, Tang QJ, Zhang CA, et al:
Anti-proliferation and anti-invasion effects of diosgenin on
gastric cancer BGC-823 cells with HIF-1α shRNA s. Int J Mol Sci.
13:6521–6533. 2012. View Article : Google Scholar
|
|
8
|
He Z, Tian Y, Zhang X, et al: Anti-tumour
and immuno-modulating activities of diosgenin, a naturally
occurring steroidal saponin. Nat Prod Res. 26:2243–2246. 2012.
View Article : Google Scholar
|
|
9
|
Liu MJ, Wang Z, Ju Y, Wong RN and Wu QY:
Diosgenin induces cell cycle arrest and apoptosis in human leukemia
K562 cells with the disruption of Ca2+ homeostasis.
Cancer Chemother Pharmacol. 55:79–90. 2005. View Article : Google Scholar
|
|
10
|
Lepage C, Leger DY, Bertrand J, Martin F,
Beneytout JL and Liagre B: Diosgenin induces death receptor-5
through activation of p38 pathway and promotes TRAIL-induced
apoptosis in colon cancer cells. Cancer Lett. 301:193–202. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
He Z, Chen H, Li G, et al: Diosgenin
inhibits the migration of human breast cancer MDA-MB-231 cells by
suppressing Vav2 activity. Phytomedicine. 21:871–876. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Du J, Cheng B, Zhu X and Ling C:
Ginsenoside Rg1, a novel glucocorticoid receptor agonist of plant
origin, maintains glucocorticoid efficacy with reduced side
effects. J Immunol. 187:942–950. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Binbin C, Yinglu F, Juan D and Changquan
L: Upregulation effect of ginsenosides on glucocorticoid receptor
in rat liver. Horm Metab Res. 41:531–536. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li F, Fernandez PP, Rajendran P, Hui KM
and Sethi G: Diosgenin, a steroidal saponin, inhibits STAT3
signaling pathway leading to suppression of proliferation and
chemosensitization of human hepatocellular carcinoma cells. Cancer
Lett. 292:197–207. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Moalic S, Liagre B, Corbiere C, et al: A
plant steroid, diosgenin, induces apoptosis, cell cycle arrest and
COX activity in osteosarcoma cells. FEBS Lett. 506:225–230. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leger DY, Liagre B, Corbiere C,
Cook-Moreau J and Beneytout JL: Diosgenin induces cell cycle arrest
and apoptosis in HEL cells with increase in intracellular calcium
level, activation of cPLA2 and COX-2 overexpression. Int J Oncol.
25:555–562. 2004.PubMed/NCBI
|
|
17
|
Satyanarayana A, Hilton MB and Kaldis P:
p21 inhibits Cdk1 in the absence of Cdk2 to maintain the G1/S phase
DNA damage checkpoint. Mol Biol Cell. 19:65–77. 2008. View Article : Google Scholar :
|
|
18
|
Chan TA, Hwang PM, Hermeking H, Kinzler KW
and Vogelstein B: Cooperative effects of genes controlling the
G(2)/M checkpoint. Genes Dev. 14:1584–1588. 2000.PubMed/NCBI
|
|
19
|
Lee E, Kwak GH, Kamble K and Kim HY:
Methionine sulfoxide reductase B3 deficiency inhibits cell growth
through the activation of p53-p21 and p27 pathways. Arch Biochem
Biophys. 547:1–5. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Masgras I, Carrera S, de Verdier PJ, et
al: Reactive oxygen species and mitochondrial sensitivity to
oxidative stress determine induction of cancer cell death by p21. J
Biol Chem. 287:9845–9854. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yadav V, Sultana S, Yadav J and Saini N:
Gatifloxacin induces S and G2-phase cell cycle arrest in pancreatic
cancer cells via p21/p27/p53. PLoS One. 7:e477962012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pal I and Mandal M: PI3K and Akt as
molecular targets for cancer therapy: current clinical outcomes.
Acta Pharmacol Sin. 33:1441–1458. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Liang J and Slingerland JM: Multiple roles
of the PI3K/PKB (Akt) pathway in cell cycle progression. Cell
Cycle. 2:339–345. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Osaki M, Oshimura M and Ito H: PI3K-Akt
pathway: its functions and alterations in human cancer. Apoptosis.
9:667–676. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Izutani Y, Yogosawa S, Sowa Y and Sakai T:
Brassinin induces G1 phase arrest through increase of p21 and p27
by inhibition of the phosphatidylinositol 3-kinase signaling
pathway in human colon cancer cells. Int J Oncol. 40:816–824.
2012.PubMed/NCBI
|
|
26
|
Georgakis GV, Li Y, Rassidakis GZ,
Medeiros LJ, Mills GB and Younes A: Inhibition of the
phosphatidylinositol-3 kinase/Akt promotes G1 cell cycle arrest and
apoptosis in Hodgkin lymphoma. Br J Haematol. 132:503–511.
2006.PubMed/NCBI
|
|
27
|
Leger DY, Liagre B and Beneytout JL: Role
of MAPKs and NF-κB in diosgenin-induced megakaryocytic
differentiation and subsequent apoptosis in HEL cells. Int J Oncol.
28:201–207. 2006.
|
|
28
|
Shishodia S and Aggarwal BB: Diosgenin
inhibits osteoclastogenesis, invasion, and proliferation through
the downregulation of Akt, I kappa B kinase activation and NF-kappa
B-regulated gene expression. Oncogene. 25:1463–1473. 2006.
View Article : Google Scholar
|
|
29
|
Srinivasan S, Koduru S, Kumar R,
Venguswamy G, Kyprianou N and Damodaran C: Diosgenin targets
Akt-mediated prosurvival signaling in human breast cancer cells.
Int J Cancer. 125:961–967. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zinkel S, Gross A and Yang E: BCL2 family
in DNA damage and cell cycle control. Cell Death Differ.
13:1351–1359. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim DS, Jeon BK, Lee YE, Woo WH and Mun
YJ: Diosgenin induces apoptosis in HepG2 cells through generation
of reactive oxygen species and mitochondrial pathway. Evid Based
Complement Alternat Med. 2012:9816752012. View Article : Google Scholar : PubMed/NCBI
|