Introduction
In suicide gene therapy, a foreign gene is
introduced into tumor cells and the expression of the gene converts
a non-toxic prodrug into a lethal drug, such as nucleoside kinases
that phosphorylate cytotoxic nucleoside analogs (1). The most extensively studied suicide
genes include the herpes simplex virus type-1 thymidine kinase
(HSV-1 TK) with ganciclovir (GCV) as a prodrug, and the cytosine
deaminase (CD) of Escherichia coli, which converts the
non-toxic antifungal agent fluorocytosine (5-FC) into
5-fluorouracil (5-FU) (2,3). Cells expressing HSV-1 TK phosphorylate
GCV, which is then incorporated into nuclear DNA during DNA
replication and repair. The incorporation of GCV terminates DNA
elongation and causes cell death (4). The killing of adjacent cells due to
the transportation of phosphorylated GCV via gap junctions, known
as the 'bystander effect', is important for successful suicide gene
therapies (5,6).
Previously, we identified a multi-substrate
deoxyribonucleoside kinase (Dm-dNK) from the fruit fly
Drosophila melanogaster and evaluated the possible use of
this enzyme as a suicide gene in vitro and in vivo
(7–11). Dm-dNK phosphorylates a broad
range of substrates including analogs of both purine and pyrimidine
nucleosides, and exhibits a higher activity than previously studied
nucleoside kinases (12,13). When expressed in human cells,
Dm-dNK localizes to the nucleus through a C-terminal nuclear
localization signal (8). We
previously performed mutagenesis of the nuclear localization signal
and investigated the effect of heterologous expression of a
cytosolic Dm-dNK on cancer cells (10). There were no differences between
cytosolic Dm-dNK and nuclear Dm-dNK in regards to
enzyme activity, cellular sensitivity to nucleoside analogs, or
bystander cell killing.
Tanshinone (Tan) IIA is a fat-soluble and
pharmacologically active ingredient of Danshen; a traditional
Chinese medicine used in the treatment of cardiovascular diseases.
It is isolated from the rhizome of a Chinese herb Salvia
miltiorrhiza (10,14). Previous studies have shown that Tan
IIA possessed not only anti-inflammatory (15) and antioxidant properties, but also
anticancer activities in cell culture and animal carcinogenesis
models (16,17). Tan IIA can restore connexin (Cx) 43
by inhibiting the elevated miR-1 expression in ischemic and hypoxic
cardiomyocytes (18,19). Connexins constitute a family of
structurally related transmembrane proteins, including Cx43 and
Cx26, which connect two adjacent cells via gap junctional
intercellular communication (GJIC) (20). Dysfunction of connexins may lead to
defects in cell proliferation, differentiation and localization,
which may be correlated with tumorigenesis (21–23).
Some studies have shown that GJIC is directly involved in the
bystander effect, by which adjacent cells are sensitized to drug
treatment during gene therapy (24–27).
Considering all these facts, Tan IIA may influence the bystander
effect of cancer cells expressing Dm-dNK by regulating the
expression of Cx43 and Cx26.
In the present study, we further characterized
Dm-dNK as a suicide gene when the enzyme was expressed in
different subcellular compartments. We compared the cytotoxicity
and the bystander effects of the nucleoside analogs
(e)-5-(2-bromovinyl)-2′-deoxyuridine (BVDU) and
1-β-D-arabinofuranosylthymine (araT) when the enzyme was expressed
in either the nuclei or the mitochondria. We showed that a
recombinant Dm-dNK with a mitochondrial targeting signal
localized to the mitochondria, retained high enzymatic activity.
The cells expressing mitochondrial Dm-dNK and nuclear
Dm-dNK had similar sensitivities to the nucleoside analogs
and similar bystander effects. The subcellular localization of
Dm-dNK did not affect the sensitivity of the cells to the
nucleoside analogs or the efficiency of bystander cell killing. We
also investigated the influence of Tan IIA on the bystander effect
of cells expressing Dm-dNK, and whether there were any
statistical differences between the Dm-dNK expression in
nuclei and in mitochondria. Furthermore, the present study,
examined the relationship between Tan IIA and the expressions of
Cx43 and Cx26 in order to elucidate the primary mechanism of the
anticancer activities of Tan IIA.
Materials and methods
Cell culture
RetroPack PT67 packaging cells (Clontech, Palo Alto,
CA, USA) and thymidine kinase (TK)-deficient osteosarcoma cells (a
gift from Professor Jan Balzarini, Rega Institute, Leuven, Belgium)
were cultured in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% (v/v) fetal calf serum (Gibco-BRL,
Gaithersburg, MD, USA), 100 U/ml penicillin, and 0.1 mg/ml
streptomycin. Cells were grown at 37°C in a humidified incubator
with a gas phase of 5% CO2.
Construction of retroviral vectors and
subcellular localization of Dm-dNK
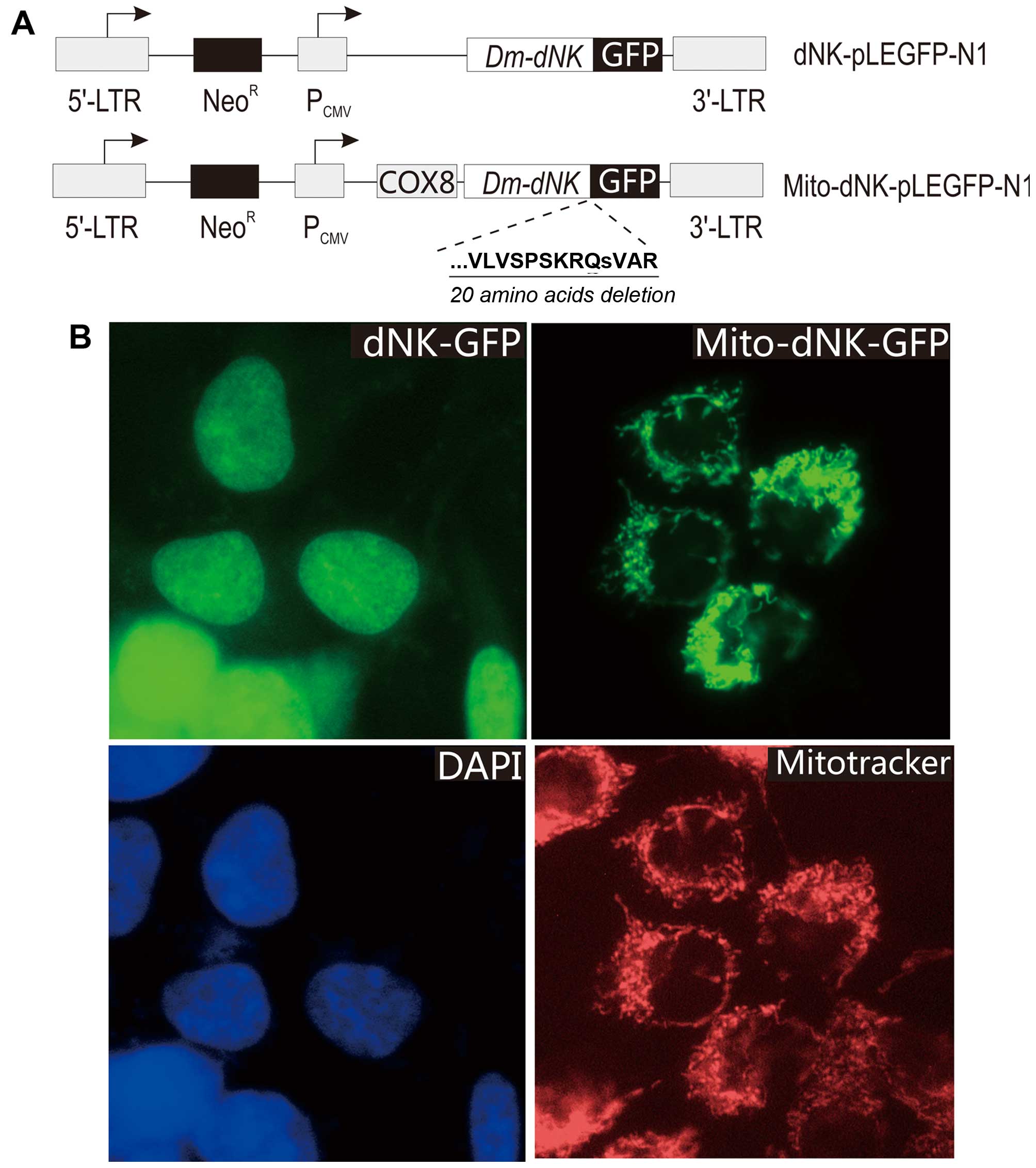
We used the pLEGFP-N1 retroviral vector (Clontech)
to express the Dm-dNK cDNA in fusion with the green
fluorescent protein (dNK-GFP) (Fig.
1). The pLEGFP-N1 with wild-type Dm-dNK was constructed
as previously described (10) and
cloned into the XhoI-BamHI site of the pLEGFP-N1 vector. The
cDNA sequence encoding the 31-amino acid N-terminal mitochondrial
import signal of cytochrome c oxidase subunit VIII was
cloned upstream of Dm-dNK, and C-terminal deletions (20
amino acids deleted) were made using the primer 5′-TCGTCGACTTATG
GATGGCGTCGAATATGCTGCT-3′. Plasmids were purified using the
NucleoBond plasmid purification kit (Clontech). The DNA sequences
of the constructed plasmids were verified using an ABI 310
automated DNA sequencer (Applied Biosystems, Foster City, CA, USA).
The constructed pLEGFP-N1 plasmids were transfected into the PT67
packaging cells using Lipofectamine (Life Technologies, Inc., Grand
Island, NY, USA) according to the protocol provided by the
supplier. The medium from the transfected cells was collected 48 h
after transfection, filtered through a 0.45-mm filter, and diluted
2-fold with fresh medium. The osteosarcoma cells were incubated
with the virus-containing medium for 48 h and cultured continuously
for 3 weeks in the presence of 1.0 mg/ml Geneticin (Gibco-BRL). The
cell nuclei were counterstained with 4′,6-diamidino-2-phenylindole
(DAPI) and the mitochondria were counterstained with MitoTracker
(Invitrogen, Ltd., Paisley, UK). GFP, DAPI and MitoTracker
fluorescence was observed using a Nikon Eclipse E600 microscope
(Nikon, Tokyo, Japan) equipped with a SPOTRT digital camera
(Diagnostic Instruments, Inc., Sterling Heights, MI, USA).
Western blot analysis and enzymatic
assays
Nuclear extracts were prepared as previously
described (8). The mitochondrial
fractions were isolated from transduced osteosarcoma cells as
previously described (28) by
differential centrifugation in lysis buffer (0.3 M mannitol, 0.1%
bovine serum albumin, 2 mM EDTA, 10 mM HEPES, pH 7.4). After cell
homogenization with a glass homogenizer, the suspension was
centrifuged for 10 min at 1,000 × g at 4°C (the supernatant
contained cellular fractions). The supernatant was centrifuged
again at 14,000 × g for 15 min at 4°C. To pellet the mitochondrial
fraction, the supernatant was ultra-centrifuged at 100,000 × g for
60 min. The concentration of the extracted protein was measured
using a BCA protein assay (Kaiji, China). The protein extracts were
separated on a 12% SDS-PAGE gel and electro-transferred to a
nitrocellulose membrane. The membrane was probed for 1 h at room
temperature with a polyclonal anti-GFP antibody (Invitrogen, Ltd.).
Binding of the primary antibody was detected using a secondary
mouse anti-rabbit immunoglobulin (Ig) conjugated to horseradish
peroxidase (Amersham, Arlington Heights, IL, USA). ECL reagents
were used to detect the signals according to the manufacturer's
instructions (Amersham). The enzymatic assays were performed as
previously described (8). For the
reaction, 3 mM [methyl-3H]dThd (Moravek Biochem, Burlington,
Ontario Canada) and 2 mM unlabeled dThd (Sigma-Aldrich, Beijing,
China) were used.
For the analysis of connexin expression, cells were
seeded at 1×104 cells/well in 6-well plates. After 24 h,
the medium was removed and replaced with complete medium containing
Tan IIA (Xi'an Guanyu Bio-tech Co., Ltd., China) at 0, 5, 10 and 20
μM, respectively. After 3 days, cells were washed twice with
PBS and lysed in RIPA buffer (8% SDS, 0.25 m Tris-HCl, pH 6.8, 1 mM
phenylmethylsulfonyl fluoride, 1.0 mg/ml leupeptin, and 10 mg/ml
aprotinin). Total cell extracts were separated on 10% SDS-PAGE
gels. Then western blot analyses were performed using anti-Cx43
(71-0700) and anti-Cx26 (CX-12h10) (both from Zymed, San Francisco,
CA, USA) or anti-actin (ms-1295-po; NeoMarkers, Fremont, CA, USA)
antibodies.
Cell proliferation and bystander killing
assays
araT was obtained from Lilly Research Laboratories.
BVDU was a gift from Professor Jan Balzarini (Rega Institute,
Leuven, Belgium). Cells were plated at 3,000 cells/well in 96-well
plates. Nucleoside analogs were added 24 h after plating and the
medium containing the nucleoside analogs was changed once during
the 4-day incubation. Cell survival was assayed using a
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay (Boehringer Mannheim, Welwyn Garden City, UK) after 4 days of
drug exposure. Each experiment was performed in triplicate.
The assay for bystander cell killing was performed
as previously described (9). Tumor
cells expressing Dm-dNK were mixed at different ratios with
their respective parental cell lines. To promote cell contacts, the
mixed cells were plated in 24-well plates at 3×105
cells/well. The following day, confluent cells were treated with
BVDU. After 24 h of incubation, cells were trypsinized and a 1:100
dilution of the cells was distributed into 96-well plates in five
replicates. Cells were cultured subsequently in the presence of
BVDU for 2–3 days until the cells without BVDU treatment reached
confluence. Cell survival was determined as described above. Each
experiment was performed in triplicate.
Effects of Tan IIA and BVDU on the growth
of mixed cells
Cells expressing dNK-GFP or mito-dNK-GFP were mixed
at a ratio of 1:9 with untransduced cells. For the measurement of
cell viability, the mixtures were seeded at 3×103
cells/well in 96-well plates. After 24 h, the medium was removed
and replaced with complete medium with or without Tan IIA (5, 10,
20 and 40 μM) for another 24 h. Cells were incubated with
BVDU (0.1 μM) for 48 h in the presence of Tan IIA. Then MTT
analysis was performed as described above. Each assay was repeated
3 times.
Statistical analysis
Data are expressed as the mean value ± standard
deviation. All experiments were performed in triplicate. All
statistical analyses were performed using SPSS version 11.0.
Comparisons among all groups were performed with one-way analysis
of variance (ANOVA) and the Student-Newman-Keuls method.
Statistical significance was indicated by a p-value <0.05.
Results
Expression of Dm-dNK in nuclei or
mitochondria in a cancer cell line
To study the effects of phosphorylated nucleoside
analogs in the mitochondria matrix and to compare the sensitivities
of the cells to nucleoside analogs and the efficiency of bystander
cell killing when the enzyme was located in either of the two
subcellular compartments, we aimed to express Dm-dNK
targeted to the mitochondria in TK-deficient osteosarcoma cancer
cell lines. We fused the mitochondrial import signal of cytochrome
c oxidase subunit VIII to the N-terminus of Dm-dNK.
For easy visualization of the subcellular localization of the
protein, we also fused GFP to the C-terminus of Dm-dNK, as
we did with wild-type Dm-dNK using the pLEGFP-N1 retroviral
vector.
To achieve this, vectors harboring replication
deficient retroviral elements were constructed to express either
the wild-type nuclear Dm-dNK (dNK-GFP) or the mitochondrial
targeted Dm-dNK mutant (mito-dNK-GFP) (Fig. 1A). A TK-1-deficient osteosarcoma
cell line was transduced with the recombinant retroviruses. After
selection of stably transfected cells, 90% of the cells exhibited
green fluorescence (Fig. 1B). The
cells transduced with the virus encoding dNK-GFP exhibited
fluorescence in the nucleus, whereas the cells transduced with the
virus encoding mito-dNK-GFP had a dotted fluorescence pattern.
These results were further confirmed by counterstaining with DAPI
and MitoTracker (Fig. 1B),
respectively. A co-localization of GFP and DAPI fluorescence
indicated that the protein was located in the nuclei, and a
co-localization of GFP and MitoTracker fluorescence indicated that
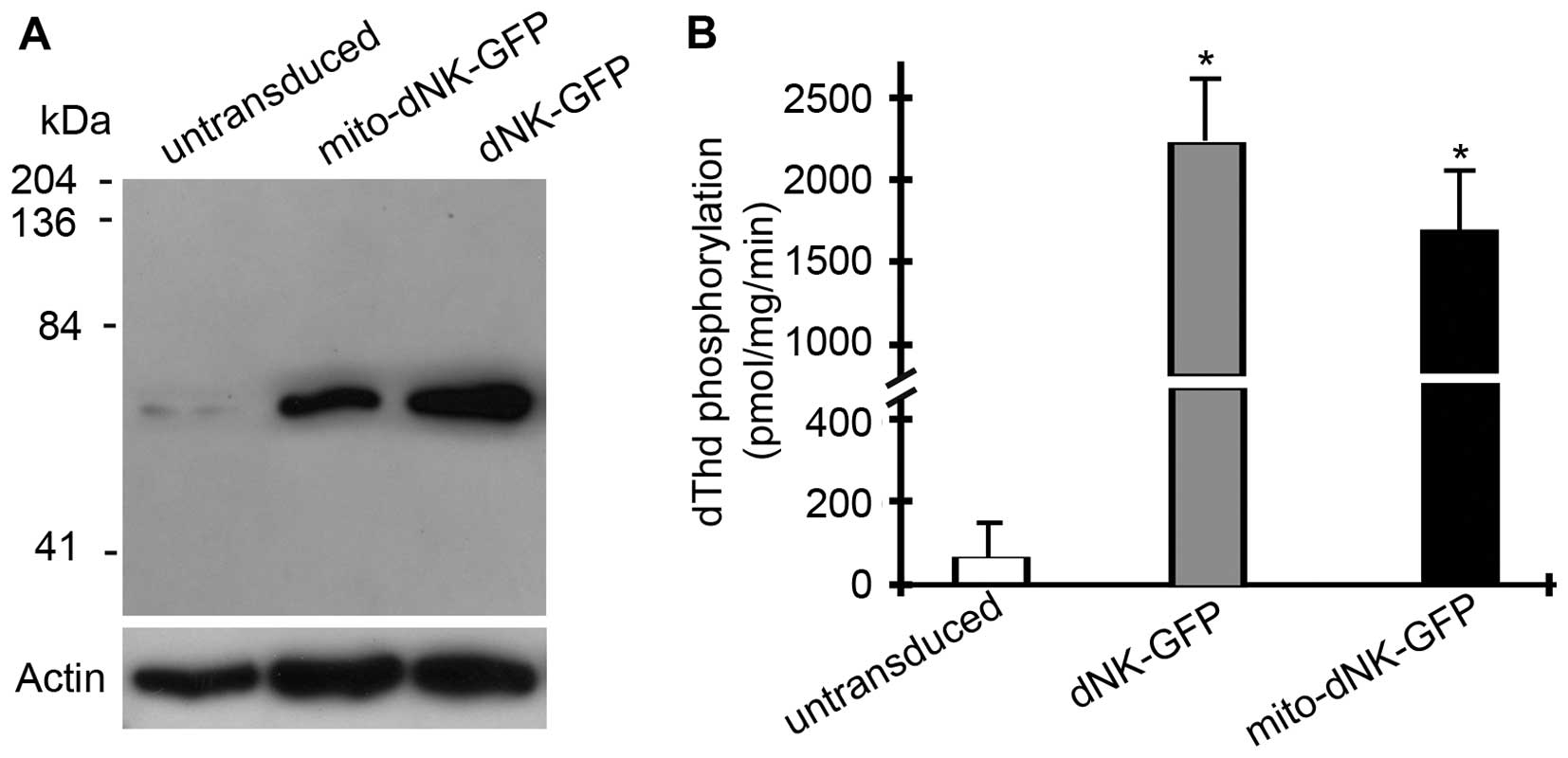
the protein was located in the mitochondria. Western blot analyses
with anti-GFP antibodies also detected the dNK-GFP and mito-dNK-GFP
fusion proteins (60 kDa) in the nuclei and the mitochondria,
respectively (Fig. 2A).
To measure the enzymatic activities of the
Dm-dNK-GFP fusion proteins, we assayed the dThd
phosphorylation activities in the cell extracts (Fig. 2B). The dThd kinase activities
increased ~40-fold in the cells expressing the nuclear dNK-GFP and
~35-fold in the cells expressing the mitochondrial mito-dNK-GFP,
compared with the untransduced parent cells (p<0.01). There was
no significant difference in Dm-dNK activity between the
cells expressing dNK-GFP in the mitochondria or in the nuclei.
Nucleoside analog sensitivity and
bystander cell killing
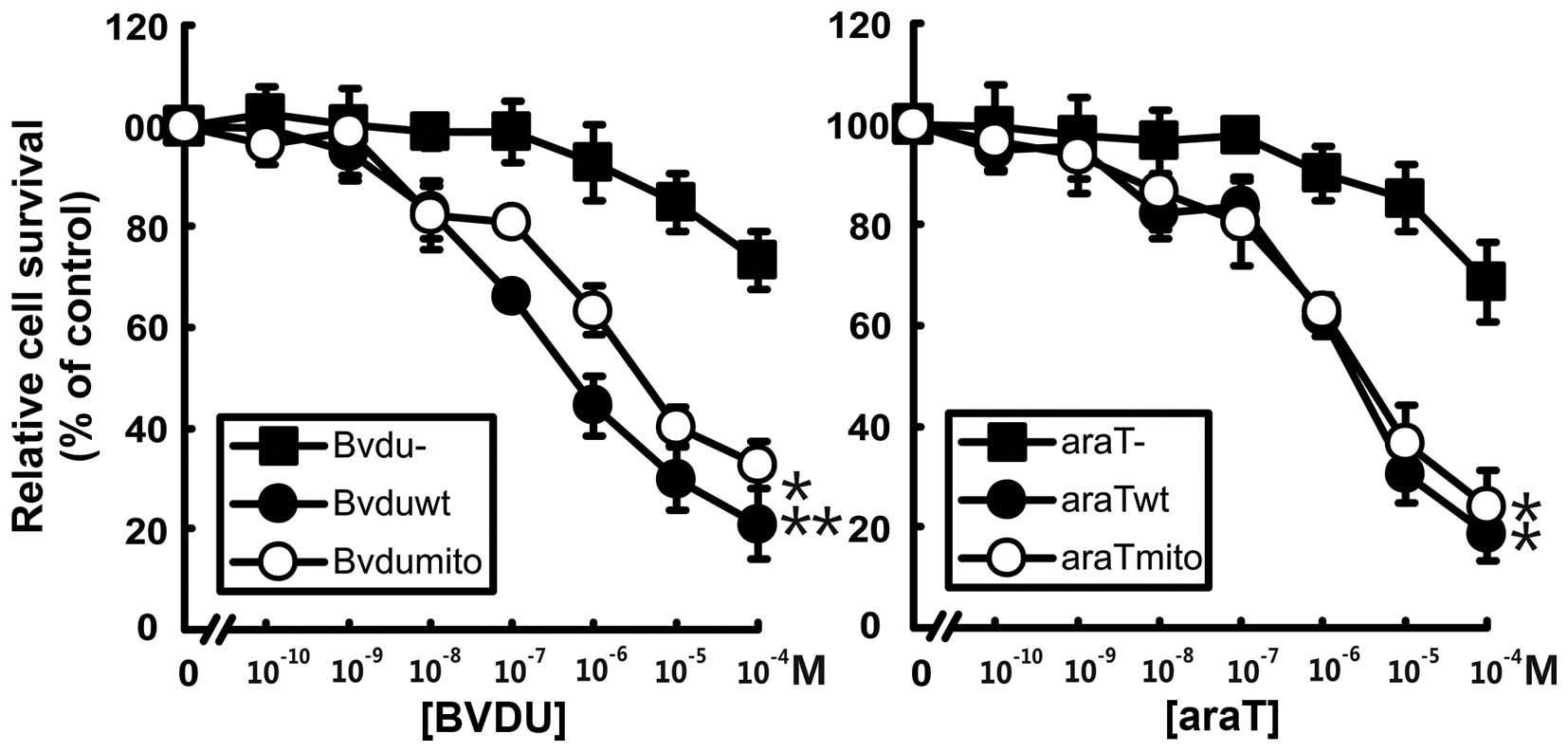
We determined the sensitivities of the transduced
cells to the pyrimidine nucleoside analogs BVDU and araT (Fig. 3). The two
Dm-dNK-GFP-expressing osteosarcoma cell lines were more
sensitive to the nucleoside analogs than untransduced cells. The
cells expressing Dm-dNK in either the nuclei or in the
mitochondria exhibited 100- to 500-fold lower IC50
values to araT and BVDU than the untransduced cells (p<0.01).
There were no differences in sensitivity to the nucleoside analogs
between the cells expressing Dm-dNK in the nuclei or in the
mitochondria.
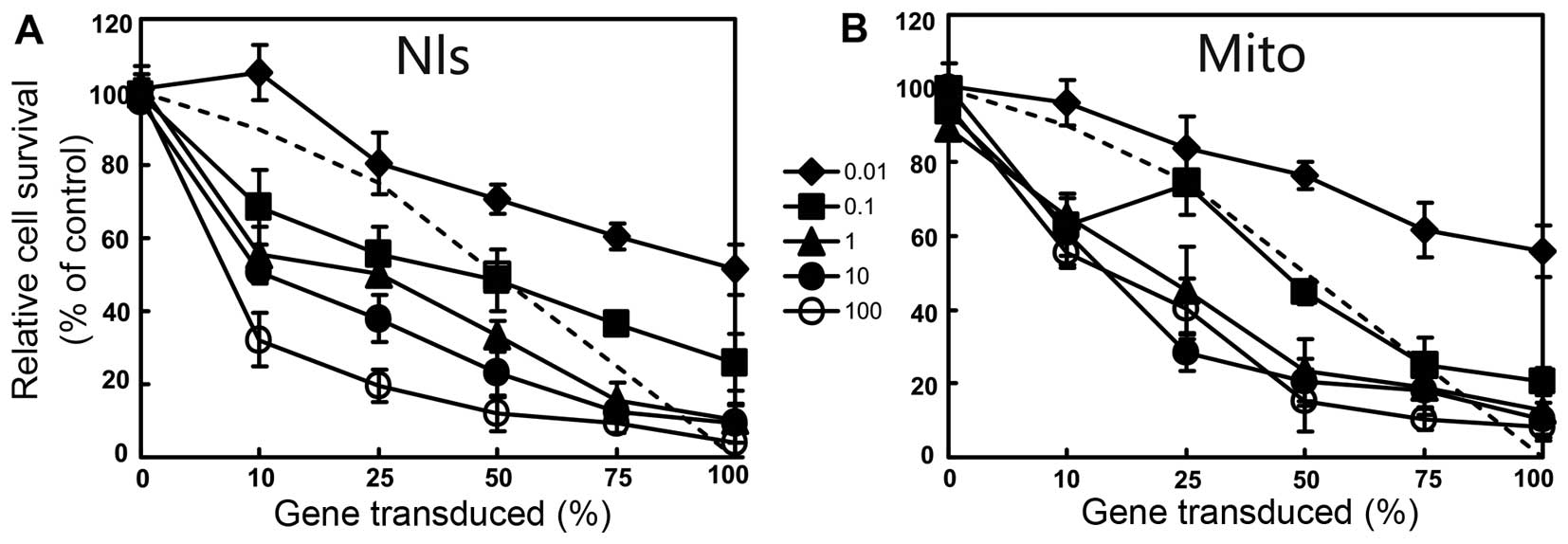
The cells expressing dNK-GFP or mito-dNK-GFP were
mixed at different ratios (0, 10, 25, 50, 75 and 100%) with the
untransduced cells. BVDU was added to the mixed cells at
concentrations from 0.01 to 100 μM and cells were incubated
for 4 days. A bystander effect was observed in the osteosarcoma
cell lines expressing dNK-GFP or mito-dNK-GFP (Fig. 4). For example, we found that 10% of
the cells expressing dNK-GFP and 25% of the cells expressing
mito-dNK-GFP induced 70% cell death in the presence of 100
μM BVDU (Fig. 4). There were
no differences in bystander killing between the cells expressing
Dm-dNK in the nuclei or in the mitochondria, in the presence
of BVDU.
Tan IIA enhances the bystander effect by
increasing the expression of Cx43 and Cx26 in TK-deficient
osteosarcoma cell lines
Previous studies have shown that Tan IIA possessed
not only anti-inflammatory (15)
and antioxidant properties (29),
but also anticancer activities through its influence on GJIC, in
cell experiments and animal carcinogenesis models (16,17).
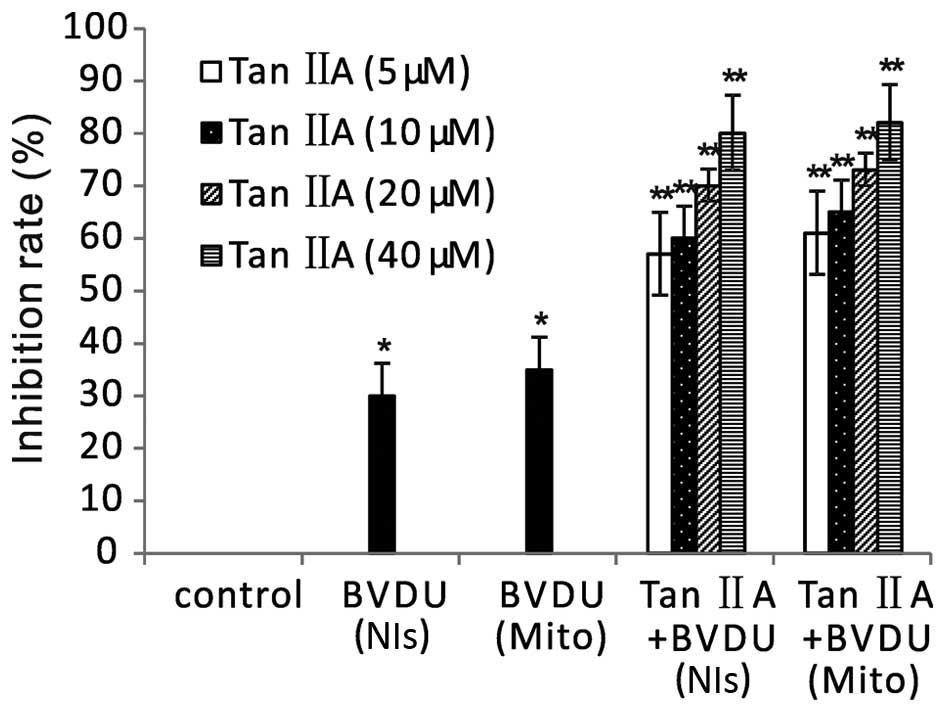
To confirm the bystander effect of Tan IIA on cancer cells, we
performed an MTT assay to assess the viability of a combination of
cells expressing dNK-GFP or mito-dNK-GFP and untransduced cells at
a ratio of 1:9. After BVDU treatment alone, there was a slight
inhibition of viability, and there was no significant difference in
the cells expressing Dm-dNK in the nuclei and in the cells
expressing Dm-dNK in the mitochondria (Fig. 5). Tan IIA and BVDU treatment induced
significantly greater inhibition of viability in mixed cells
compared with BVDU treatment alone (p<0.01). There was no
significant difference between the cells expressing dNK-GFP and the
cells expressing mito-dNK-GFP.
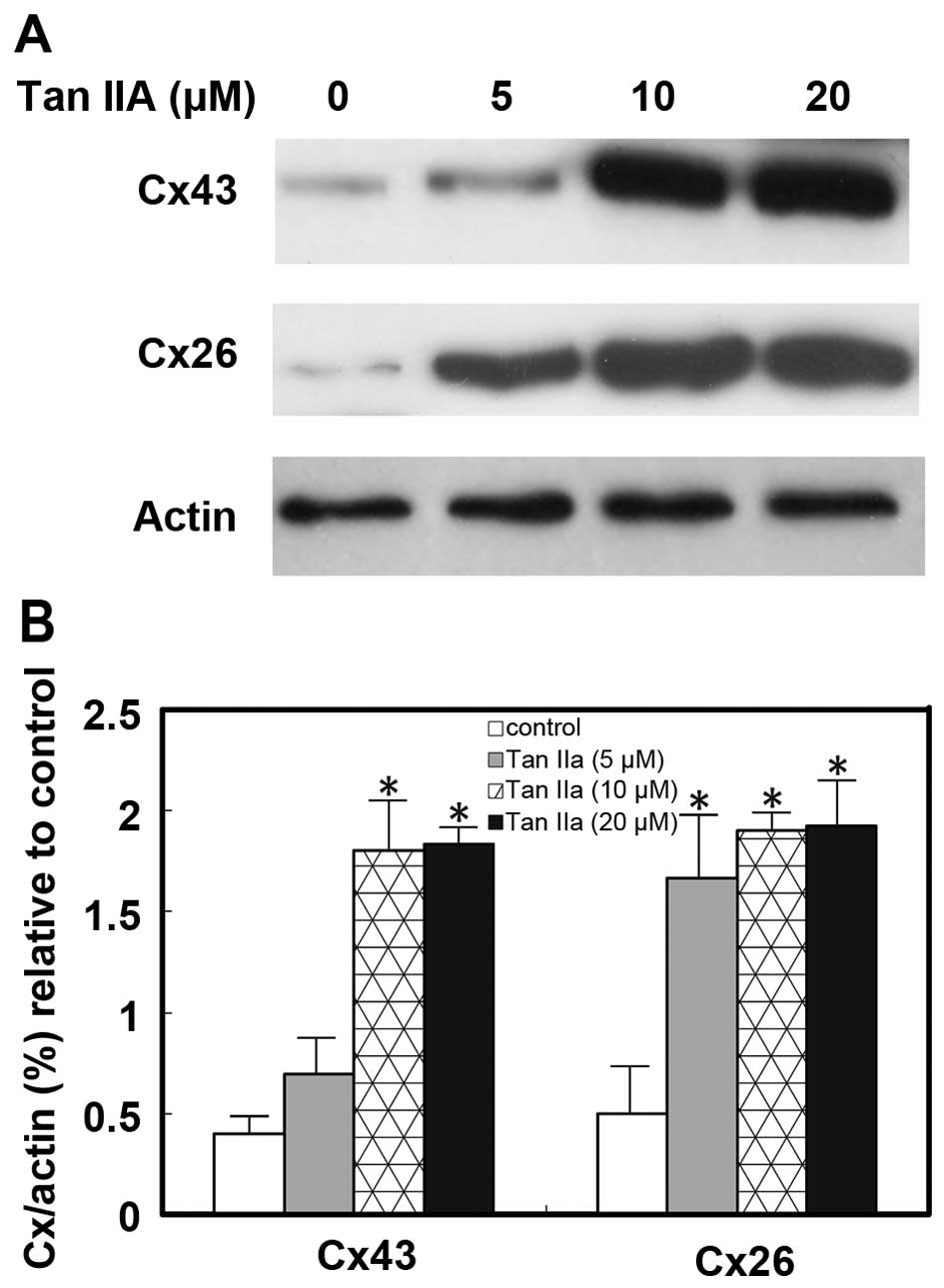
To further ascertain the effect of Tan IIA on GJIC
in TK-deficient osteosarcoma cells, we examined the influence of
Tan IIA on the expression of connexins Cx43 and Cx26. Cells were
cultured and treated with Tan IIA at different concentrations (0,
5, 10 and 20 mM) for 72 h. The results of western blot analysis
showed that the expression levels of Cx43 and Cx26 were
significantly upregulated (p<0.05; Fig. 6). Thus the exposure of TK-deficient
osteosarcoma cells to Tan IIA upregulated the expression of the
important gap junction proteins Cx43 and Cx26.
Discussion
Dm-dNK localizes to cell nuclei when
expressed in human cell lines and this localization is mediated by
a nuclear localization signal in the C-terminal region of the
protein (8). It is likely that the
nuclear localization signal traps the protein in the nuclei and
prevents it from importing to the mitochondria (30). We used a mutant and fused the
mitochondrial import signal to its N-terminus and GFP to the
C-terminus (mito-dNK-GFP), and deleted 20 C-terminal amino acid
residues that contained the nuclear localization signal. An
osteosarcoma cell line stably expressing the mitochondrial
Dm-dNK was generated and exhibited green fluorescence. Our
data showed that Dm-dNK retained its activity when expressed
in the mitochondria, and that the cell lines expressing the enzyme
exhibited increased sensitivity to the tested nucleoside analogs.
These results were the same as those obtained with a mutant of
Dm-dNK generated by cloning a mitochondrial import signal
before Dm-dNK into the pEGFP-N1 vector but with a mutation
of the arginine residue to serine in position 247, to destroy the
nuclear localization signal (30).
Mitochondrial DNA is replicated independently of
nuclear DNA and the dNTP precursor pool is also separated (31,32)
due to the inner mitochondrial membrane between the mitochondrial
matrix and the cytosol (33).
Studies suggest that the majority of deoxyribonucleotides in the
mitochondrial matrix may be trapped and are incorporated into
mitochondrial DNA directly (32,34).
The enzymes involved in mitochondrial DNA replication differ from
those catalyzing nuclear DNA replication, and several features of
DNA replication in these two compartments are also different.
Unlike the replication of nuclear DNA, mitochondrial DNA
replication is independent of the cell cycle and lacks efficient
repair mechanisms (35,36). These features may be beneficial when
the enzymes/nucleoside analogs involved in mitochondrial DNA
replication are used in suicide gene therapies.
We previously found that there are no differences in
enzyme activity, cellular sensitivity to nucleoside analogs, and
bystander cell killing whether the enzyme is expressed in the
cytosol or in the nucleus (10). In
the present study, we showed that whether the Dm-dNK-GFP was
expressed in the nuclei or in the mitochondria also did not affect
cellular sensitivity to cytotoxic nucleoside analogs or the
bystander effect. Our data also showed that the cellular
sensitivity to cytotoxic nucleoside analogs and to bystander cell
killing were not dependent on the subcellular localization of the
enzyme. However, previous results have suggested that due to the
compartmentalized dNTP pools, nucleoside analogs phosphorylated in
the mitochondrial matrix do not induce bystander cell killing
(37). There are important
differences between mitochondrial and nuclear DNA physiology. These
include the non-S-phase restricted replication of mitochondrial DNA
and the less efficient DNA repair systems in mitochondria compared
with nuclear DNA replication (35,36).
These differences suggest that nucleoside analogs targeting nuclear
or mitochondrial DNA may exhibit different pharmacological
profiles. The nucleoside analogs may accordingly, primarily affect
mitochondrial DNA. Bystander cell killing is mediated by the
intercellular transport of phosphorylated nucleoside analogs via
gap junctions in the cell membrane, and gap junctions may also
exist in the inner mitochondrial membrane. An alternative
explanation for this, is that a small amount of phosphorylated
nucleoside analogs are transported from the mitochondrial matrix to
the cytosol and nucleus. Previous studies have suggested that
nucleoside analogs phosphorylated within the mitochondrial matrix
are trapped and fail to be exported to the cytosol or nucleus
(31,34). Certain nucleosides and nucleoside
analogs seem to be preferentially incorporated into both
mitochondrial and nuclear DNA, and there is strong evidence
supporting the communication between the mitochondrial and
cytosolic/nuclear dNTP pools (38).
We cannot presently distinguish between these mechanisms based on
the results obtained. To develop novel treatment strategies, the
cell culture model system described herein will be used to further
study the pharmacological effects of nucleoside analog
phosphorylation in the mitochondria.
Previous studies have shown that Tan IIA has
anticancer activities and can influence the bystander effect, which
is mediated by gap junctional intercellular communication (GJIC)
(16,17). We found that Tan IIA and BVDU
significantly enhanced the bystander cell killing compared with
BVDU treatment alone, and that there was no significant difference
between cells expressing Dm-dNK in nuclei or in
mitochondria. Tan IIA was found to increase the expression of Cx43
by inhibiting the elevated miR-1 expression in ischemic and hypoxic
cardiomyocytes (18,19). We also found in this study that Tan
IIA increased the expression of Cx43 and Cx26. The aberrant
expression and downregulation of Cx43 and Cx26 may lead to the
progression of various cancers (39–41).
The present results revealed the primary mechanism involved in the
enhanced effect of Tan IIA on the bystander effect in cells
expressing Dm-dNK in nuclei and mitochondria. Our study
showed that a safe dosage of Tan IIA exhibited a strong cancer cell
killing effect. However, further studies are needed to examine the
substantial clinical benefit of Tan IIA for clinical practice in
the future.
Acknowledgments
This study was supported by grants from the Hi-Tech
Research Development Program of China (863 Program, 2006AA02Z493)
and the National Natural science Foundation of China (no. 81071900,
81172199 and 81272920).
References
|
1
|
Duarte S, Carle G, Faneca H, de Lima MC
and Pierrefite-Carle V: Suicide gene therapy in cancer: Where do we
stand now? Cancer Lett. 324:160–170. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Culver KW, Ram Z, Wallbridge S, Ishii H,
Oldfield EH and Blaese RM: In vivo gene transfer with retroviral
vector-producer cells for treatment of experimental brain tumors.
Science. 256:1550–1552. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Moolten FL: Tumor chemosensitivity
conferred by inserted herpes thymidine kinase genes: Paradigm for a
prospective cancer control strategy. Cancer Res. 46:5276–5281.
1986.PubMed/NCBI
|
|
4
|
Reardon JE: Herpes simplex virus type 1
and human DNA polymerase interactions with 2′-deoxyguanosine
5′-triphosphate analogues. Kinetics of incorporation into DNA and
induction of inhibition. J Biol Chem. 264:19039–19044.
1989.PubMed/NCBI
|
|
5
|
Dilber MS, Abedi MR, Christensson B,
Björkstrand B, Kidder GM, Naus CC, Gahrton G and Smith CI: Gap
junctions promote the bystander effect of herpes simplex virus
thymidine kinase in vivo. Cancer Res. 57:1523–1528. 1997.PubMed/NCBI
|
|
6
|
Freeman SM, Abboud CN, Whartenby KA,
Packman CH, Koeplin DS, Moolten FL and Abraham GN: The 'bystander
effect': Tumor regression when a fraction of the tumor mass is
genetically modified. Cancer Res. 53:5274–5283. 1993.PubMed/NCBI
|
|
7
|
Ma S, Zhao L, Zhu Z, Liu Q, Xu H,
Johansson M, Karlsson A and Zheng X: The multisubstrate
deoxyribonucleoside kinase of Drosophila melanogaster as a
therapeutic suicide gene of breast cancer cells. J Gene Med.
13:305–311. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zheng X, Johansson M and Karlsson A:
Retroviral transduction of cancer cell lines with the gene encoding
Drosophila melanogaster multisubstrate deoxyribonucleoside kinase.
J Biol Chem. 275:39125–39129. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zheng X, Johansson M and Karlsson A:
Bystander effects of cancer cell lines transduced with the
multisubstrate deoxyribonucleoside kinase of Drosophila
melanogaster and synergistic enhancement by hydroxyurea. Mol
Pharmacol. 60:262–266. 2001.PubMed/NCBI
|
|
10
|
Zheng X, Johansson M and Karlsson A:
Nucleoside analog cytotoxicity and bystander cell killing of cancer
cells expressing Drosophila melanogaster deoxyribonucleoside kinase
in the nucleus or cytosol. Biochem Biophys Res Commun. 289:229–233.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhu Z, Mao L, Zhao L, Sun Z, Wang Z, Xu H
and Zheng X: Synergistic therapeutic effect in gastric cancer cells
produced by oncolytic adenovirus encoding Drosophila melanogaster
deoxyribonucleoside kinase. Cancer Biol Ther. 11:874–882. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Johansson M, van Rompay AR, Degrève B,
Balzarini J and Karlsson A: Cloning and characterization of the
multisubstrate deoxyribonucleoside kinase of Drosophila
melanogaster. J Biol Chem. 274:23814–23819. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Munch-Petersen B, Piskur J and Sondergaard
L: Four deoxynucleoside kinase activities from Drosophila
melanogaster are contained within a single monomeric enzyme, a new
multifunctional deoxynucleoside kinase. J Biol Chem. 273:3926–3931.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Fish JM, Welchons DR, Kim YS, Lee SH, Ho
WK and Antzelevitch C: Dimethyl lithospermate B, an extract of
Danshen, suppresses arrhythmogenesis associated with the Brugada
syndrome. Circulation. 113:1393–1400. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yin X, Yin Y, Cao FL, Chen YF, Peng Y, Hou
WG, Sun SK and Luo ZJ: Tanshinone IIA attenuates the inflammatory
response and apoptosis after traumatic injury of the spinal cord in
adult rats. PLoS One. 7:e383812012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu F, Yu G, Wang G, Liu H, Wu X, Wang Q,
Liu M, Liao K, Wu M, Cheng X, et al: An NQO1-initiated and
p53-independent apoptotic pathway determines the anti-tumor effect
of tanshinone IIA against non-small cell lung cancer. PLoS One.
7:e421382012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang J, Wang X, Jiang S, Yuan S, Lin P,
Zhang J, Lu Y, Wang Q, Xiong Z, Wu Y, et al: Growth inhibition and
induction of apoptosis and differentiation of tanshinone IIA in
human glioma cells. J Neurooncol. 82:11–21. 2007. View Article : Google Scholar
|
|
18
|
Zhang Y, Zhang L, Chu W, Wang B, Zhang J,
Zhao M, Li X, Li B, Lu Y, Yang B, et al: Tanshinone IIA inhibits
miR-1 expression through p38 MAPK signal pathway in post-infarction
rat cardiomyocytes. Cell Physiol Biochem. 26:991–998. 2010.
View Article : Google Scholar
|
|
19
|
Shan H, Li X, Pan Z, Zhang L, Cai B, Zhang
Y, Xu C, Chu W, Qiao G, Li B, et al: Tanshinone IIA protects
against sudden cardiac death induced by lethal arrhythmias via
repression of microRNA-1. Br J Pharmacol. 158:1227–1235. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Söhl G and Willecke K: Gap junctions and
the connexin protein family. Cardiovasc Res. 62:228–232. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dbouk HA, Mroue RM, El-Sabban ME and
Talhouk RS: Connexins: A myriad of functions extending beyond
assembly of gap junction channels. Cell Commun Signal. 7:42009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Klein G: Cancer, apoptosis, and nonimmune
surveillance. Cell Death Differ. 11:13–17. 2004. View Article : Google Scholar
|
|
23
|
Trosko JE and Ruch RJ: Cell-cell
communication in carcinogenesis. Front Biosci. 3:d208–d236.
1998.PubMed/NCBI
|
|
24
|
Mesnil M, Piccoli C, Tiraby G, Willecke K
and Yamasaki H: Bystander killing of cancer cells by herpes simplex
virus thymidine kinase gene is mediated by connexins. Proc Natl
Acad Sci USA. 93:1831–1835. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Elshami AA, Saavedra A, Zhang H,
Kucharczuk JC, Spray DC, Fishman GI, Amin KM, Kaiser LR and Albelda
SM: Gap junctions play a role in the 'bystander effect' of the
herpes simplex virus thymidine kinase/ganciclovir system in vitro.
Gene Ther. 3:85–92. 1996.PubMed/NCBI
|
|
26
|
Wygoda MR, Wilson MR, Davis MA, Trosko JE,
Rehemtulla A and Lawrence TS: Protection of herpes simplex virus
thymidine kinase-transduced cells from ganciclovir-mediated
cytotoxicity by bystander cells: The Good Samaritan effect. Cancer
Res. 57:1699–1703. 1997.PubMed/NCBI
|
|
27
|
Lawrence TS, Rehemtulla A, Ng EY, Wilson
M, Trosko JE and Stetson PL: Preferential cytotoxicity of cells
transduced with cytosine deaminase compared to bystander cells
after treatment with 5-fucytosine. Cancer Res. 58:2588–2593.
1998.PubMed/NCBI
|
|
28
|
Ljubkovic M, Marinovic J, Fuchs A, Bosnjak
ZJ and Bienengraeber M: Targeted expression of Kir6.2 in
mitochondria confers protection against hypoxic stress. J Physiol.
577:17–29. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lin R, Wang WR, Liu JT, Yang GD and Han
CJ: Protective effect of tanshinone IIA on human umbilical vein
endothelial cell injured by hydrogen peroxide and its mechanism. J
Ethnopharmacol. 108:217–222. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Solaroli N, Zheng X, Johansson M,
Balzarini J and Karlsson A: Mitochondrial expression of the
Drosophila melanogaster multisubstrate deoxyribonucleoside kinase.
Mol Pharmacol. 72:1593–1598. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bestwick RK, Moffett GL and Mathews CK:
Selective expansion of mitochondrial nucleoside triphosphate pools
in anti metabolite-treated HeLa cells. J Biol Chem. 257:9300–9304.
1982.PubMed/NCBI
|
|
32
|
Zhu C, Johansson M and Karlsson A:
Incorporation of nucleoside analogs into nuclear or mitochondrial
DNA is determined by the intracellular phosphorylation site. J Biol
Chem. 275:26727–26731. 2000.PubMed/NCBI
|
|
33
|
Dolce V, Fiermonte G, Runswick MJ,
Palmieri F and Walker JE: The human mitochondrial deoxynucleotide
carrier and its role in the toxicity of nucleoside antivirals. Proc
Natl Acad Sci USA. 98:2284–2288. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Berk AJ and Clayton DA: A genetically
distinct thymidine kinase in mammalian mitochondria. Exclusive
labeling of mitochondrial deoxyribonucleic acid. J Biol Chem.
248:2722–2729. 1973.PubMed/NCBI
|
|
35
|
Bogenhagen D and Clayton DA: Mouse L cell
mitochondrial DNA molecules are selected randomly for replication
throughout the cell cycle. Cell. 11:719–727. 1977. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Bogenhagen DF: Repair of mtDNA in
vertebrates. Am J hum Genet. 64:1276–1281. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Sanda A, Zhu C, Johansson M and Karlsson
A: Bystander effects of nucleoside analogs phosphorylated in the
cytosol or mitochondria. Biochem Biophys Res Commun. 287:1163–1166.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ferraro P, Nicolosi L, Bernardi P,
Reichard P and Bianchi V: Mitochondrial deoxynucleotide pool sizes
in mouse liver and evidence for a transport mechanism for thymidine
monophosphate. Proc Natl Acad Sci USA. 103:18586–18591. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ezumi K, Yamamoto H, Murata K, Higashiyama
M, Damdinsuren B, Nakamura Y, Kyo N, Okami J, Ngan CY, Takemasa I,
et al: Aberrant expression of connexin 26 is associated with lung
metastasis of colorectal cancer. Clin Cancer Res. 14:677–684. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ozawa H, Matsunaga T, Kamiya K, Tokumaru
Y, Fujii M, Tomita T and Ogawa K: Decreased expression of
connexin-30 and aberrant expression of connexin-26 in human head
and neck cancer. Anticancer Res. 27:2189–2195. 2007.PubMed/NCBI
|
|
41
|
Chen Y, Hühn D, Knösel T,
Pacyna-Gengelbach M, Deutschmann N and Petersen I: Downregulation
of connexin 26 in human lung cancer is related to promoter
methylation. Int J Cancer. 113:14–21. 2005. View Article : Google Scholar
|