Introduction
Lung cancer is one of the most common malignancies
and results in large number of cancer-related deaths in the world.
Approximately 1.6 million new cases are diagnosed annually. More
than 85% of those cases are defined as non-small cell lung cancer
(NSCLC) which is comprised of adenocarcinoma (ADC), squamous cell
carcinoma (SCC), large-cell carcinoma (LLC) and other types
(1). The diagnosis and clinical
treatments of advanced NSCLC remains a great challenge of medical
oncology and the 5-year survival rates is <16%, causing 1.4
million deaths annually (2). Hence,
therapies aimed at understanding the disease development and
progression, inhibiting further recurrences, or retarding the tumor
colony growth are urgently needed. Recently, the understanding of
the molecular mechanisms of NSCLC has progressed significantly.
Several oncogene amplifications and tumor-driven mutations have
been identified such as KRAS, EGFR and ALK (3,4).
However, the biology of NSCLC still remains to be well
characterized.
In cancer cells, aerobic glycolysis is one of the
most important metabolic processes even if there is enough oxygen
(5). This metabolic decision which
divert pyruvate and its precursors undergo other anabolic processes
or convert to lactate for cancer cells is known as the Warburg
effect (6). Also the intermediates
support nucleic, lipid and protein synthesis during the rapid cell
division. So the Warburg effect supplies the high energy demands
for cancer cell and biosynthesis materials. During these molecular
adaptations, high pyruvate concentrations are maintained in cancer
cells, which results in HIF-1α stabilization (7). Then HIF-1α acts as a key mediator that
transcriptionally regulates expression of a vast variety of genes
to promote cell proliferation, migration and metastasis (8–10).
However, the molecular mechanism of increased glycolysis in NSCLC
is largely unknown.
Pyruvate dehydrogenase (PDH) is a mitochondrial
multienzyme complex that catalyzes the oxidative decarboxylation of
pyruvate and is one of the major enzymes responsible for the
regulation of homeostasis of carbohydrate (11,12).
Pyruvate dehydrogenase kinases (PDKs) could regulate PDH
phosphorylation which leads to the inactivation of PDH (13). PDK1 has frequently increased
expression in cancer cells including gastric cancer, colon cancer
and acute myeloid leukemia (14–16).
Besides, Koukourakis et al performed an immunohistochemical
study and found that PDH/PDK pathway was repressed in 73% of NSCLC
(17). However, the precise
mechanisms and functions underlying the aberrant expression of PDK1
in NSCLC are still largely unclear, even controversy exist at
present.
In this study, we used public database and our own
clinical tissue samples to evaluate the expression and significance
of PDK1 in NSCLC. PDK1 was closely associated with advanced T stage
and multivariate analysis demonstrated that upregulated PDK1 was an
independent prognostic factor. Moreover, ectopic expression or
knockdown of PDK1 showed that PDK1 promoted cell proliferation and
inhibited apoptosis via modulating the Warburg effect. These
results provided new insights into the NSCLC progression.
Materials and methods
Ethics and tissue specimens
Tissue microarray containing a total of 132 NSCLC
cases was obtained from Outdo Biotech Co. (Shanghai, China). The
age of these patients ranged from 20 to 84 (mean 60.5) and a 5-year
follow-up was completed. The study was approved by the Ethics
Committee of Heilongjiang Province Hospital.
Cell lines and transfection
NSCLC cell lines, including A549, H1299 and NCI-H460
were obtained form Cell Bank (SIBS, China). All cells were
maintained in Dulbecco's modified Eagle's medium (DEME),
supplemented with 10% fetal bovine serum and antibiotics (100 U/ml
penicillin and 100 µg/ml streptomycin). PDK1 overexpression plasmid
pcDNA3.1-PDK1 and empty plasmid were transiently transfected into
cells using Lipofectamine 2000 (Invitrogen, USA). Lentivirus
containing PDK1 shRNA (RiboBio, China) were used to infect cells to
establish the stable knock-down cell lines.
Immunohistochemical staining
(IHC)
The tissue microarray slides were deparaffinized,
rehydrated in xylene and graded ethanol. Then antigens were
retrieved at 95°C for 30 min. After cooling to room temperature,
the slides were incubated with 3% hydrogen peroxide to block the
endogenous peroxidase activity. Anti-PDK1 antibody was added to the
slides overnight. After washing, the slides were incubated with
HRP-labeled secondary antibody and stained using diaminobenzidine
solution and counterstained with hematoxylin.
The IHC results were measured using a
semi-quantitative score for both intensity and percentage of
positive staining. Briefly, the intensity of staining was scored
as: 1 (no stain), 2 (weak stain), 3 (medium stain), 4 (strong
stain) and the percentage was scored as: 1 (<25%), 2 (25–50%), 3
(50–75%), 4 (>75%). Then, the final score was calculated by
multiplying the two scores and the overall score of 8 was the
cut-off score which classified them into low expression and high
expression groups.
Western blot analysis
Cell lysates were prepared using RIPA buffer with
protease inhibitor and phosphatase inhibitor cocktail. Total
protein (70 µg) was loaded to 10% SDS-PAGE and transferred onto
PVDF membranes. After blocking with 5% BSA, the membranes were
blotted with primary antibody at 4°C overnight. Membranes were then
incubated with peroxidase-conjugated secondary antibody at 37°C.
The blotting was visualized by enhanced chemiluminescence detection
system. The antibodies used in this study were as followed:
anti-PDK1 (10026-1-AP) from ProteinTech Group, Inc. (USA),
anti-p-PDH (ab177461), anti-β-actin (ab8226) from Abcam (Cambridge,
MA, USA) and anti-PDH (#3205) from Cell Signaling Technology (MA,
USA).
RNA extraction and quantitative
real-time PCR (qRT-PCR)
Total RNA was extracted by TRIzol reagent
(Invitrogen) according to the manufacturer's insturctions. The
first-strand cDNA was synthesized by PrimeScript RT Reagent kit
(Takara, Japan). Quantitative real-time PCR was performed by SYBR
Green method in ABI7500 system. The primers used in the study were
as follows: PDK1, forward, 5′-CTCAGGACACCATCCGTTCA-3′; reverse,
5′-ACCATGTTCTTCTAGGCCTTTCAT-3′. β-actin, forward,
5′-TGATGATATCGCCGCGCTC-3′; reverse, 5′-CCATCACGCCCTGGTGC-3′.
β-actin was used as internal control and the level of PDK1 was
normalized using 2−∆∆ct method.
Cell viability assay
For analyzing the viability of PDK1 overexpressing
or knockdown cells, 2×103 cells/well were seeded in
96-well plates. Cell Counting Kit-8 (CCK-8, Dojindo, Japan) was
used to measure the cell viability at 24, 48, 72 and 96 h according
to the manufacturer's insturctions. The absorbance was tested at
450 nm. The experiments were repeated at least three times.
Colony formation assay
Indicated cells were seeded into the 6-well plate at
500 cells per well after 48 hours of transfected with vector, PDK1
plasmid or infected with PDK1 shRNA. The medium was changed every
four days. After 2 weeks of incubation, colonies were fixed with 4%
paraformaldehyde and stained with crystal violet. The colony number
was measured under a microscope. All of the experiments were
performed at least three times.
Apoptosis assay
The apoptosis of cells with PDK1 ectopic expression
or knockdown was analyzed using Annexin V/PI apoptosis detection
kit (BD Bioscience, San Diego, CA, USA). Cells were trypsinized,
collected in PBS, and resuspended in binding buffer. After
incubation with Annexin V/PI for 30 min the proportion of Annexin
V-positive cells were analyzed by FACSCalibur flow cytometer. All
of the experiments were performed at least three times.
Migration assay
To detect cell migration ability, indicated cells
(104 in serum-free medium) were seeded into the top
chamber of Transwell. The bottom chamber contained complete medium
with 20% serum. After 48 h of incubation at 37°C, cells on the
lower membrane were fixed and stained with 0.1% crystal violet and
imaged.
Lactate production, glucose uptake,
oxygen consumption, ATP level detection
For lactate production measurement, cells were
seeded into 96-well plates with phenol red-free medium and
determined using Lactate assay kit (BioVision, CA, USA). For
glucose uptake assay, the 3H-2-deoxyglucose was
detected. Cells were cultured in 6-well plates and
3H-2-deoxyglucose was added to the cells and then washed
with PBS and lysed in SDS. The radioactivity of cell lysates were
detected. For oxygen consumption assay, the consumption rate was
measured with MitoCell system (Strathkelvin Instruments, UK). ATP
levels were measured using CellTiter-Glo Luminescent Viability
Assay kit (Promega).
Statistical analysis
Statistical analysis was performed with SPSS 21.0
software (IBM, Chicago, IL, USA). The correlation between PDK1
expression and clinicopathological parameters was measured by
Chi-square test. Survival rate was measured by Kaplan-Meier method
and the significance was calculated with log-rank test. Univariate
and multivariate analysis were performed by Cox regression model.
Student's t-test was used to compare the groups in the cell
experiments. Data are presented as the mean ± SEM of three
independent experiments. A P-value of <0.05 was considered
statistically significant.
Results
PDK1 is upregulated in NSCLC
tissues
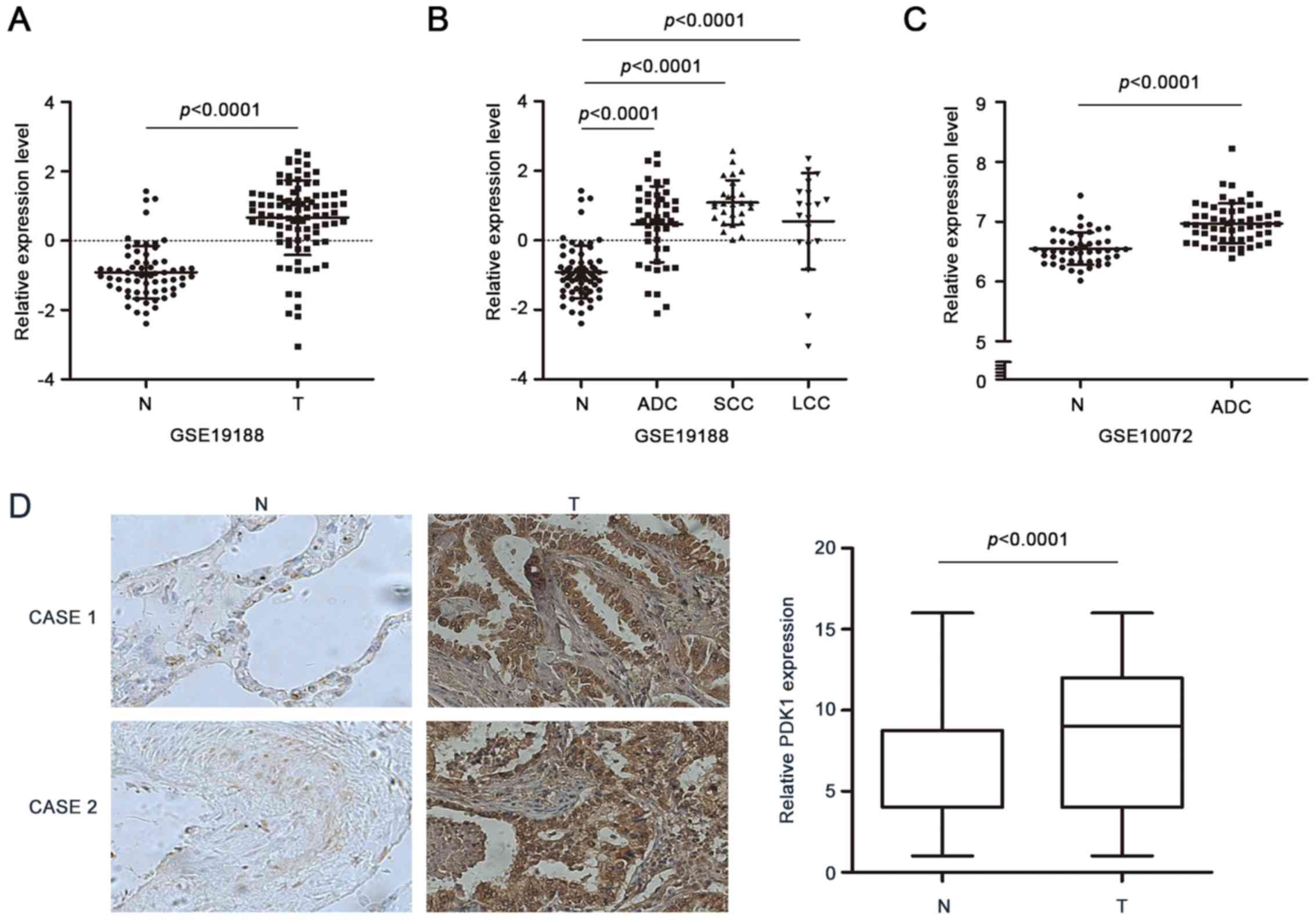
To explore the relationship between the important
enzymes involved in glycolysis and NSCLC progression, we analyzed
the previous studies that identified numerous deregulated genes in
NSCLC and focused on the glycolytic enzymes. We noted that PDK1 was
upregulated in several studies which presented expression profiling
by array. According to the GSE19188, containing a cohort of 91
tumor- and 65 adjacent normal lung tissue samples, PDK1 was
significantly higher in tumor tissues compared to normal tissues
(Fig. 1A). More importantly, the
PDK1 expression in individual subtype of lung cancer, including
adenocarcinoma (ADC), squamous cell carcinoma (SCC) and large cell
carcinoma, was also enhanced (Fig.
1B). Furthermore, we employed another dataset to validate the
hypothesis. As shown in Fig. 1C,
the PDK1 expression in GSE10072 containing 180 cases of paired
tumor samples was in conformity with our observation.
Then we examined the protein level of PDK1 in 132
pairs of NSCLC samples by immunohistochemical staining. The results
revealed that PDK1 levels were significantly higher in tumor
tissues than that in non-tumor tissues (Fig. 1D). We also analyzed the relationship
between PDK1 and clinicopathological features. The results
indicated that PDK1 expression was correlated with T stage
(P=0.019) (Table I). These data
suggested that PDK1 expression increased with the NSCLC
development.
 | Table I.Correlation between PDK1 expression
and clinicopathological parameters of NSCLC. |
Table I.
Correlation between PDK1 expression
and clinicopathological parameters of NSCLC.
|
| PDK1 expression |
|---|
|
|
|
|---|
| Parameters | No. | Low | High | P-value |
|---|
| Overall | 132 | 39 | 93 |
|
| Gender |
|
|
|
|
| Male | 69 | 17 | 52 | 0.196 |
|
Female | 63 | 22 | 41 |
|
| Age (years) |
|
|
|
|
|
<60 | 60 | 10 | 42 | 0.917 |
| ≥60 | 72 | 21 | 51 |
|
| Tumor size (cm) |
|
|
|
|
|
<3 | 37 | 11 | 26 | 0.977 |
| ≥3 | 95 | 28 | 67 |
|
| Tumor
differentiation |
|
|
|
|
| Well | 20 | 6 | 14 | 0.998 |
|
Moderate | 75 | 22 | 53 |
|
| Poor | 37 | 11 | 26 |
|
| pT status |
|
|
|
|
| pT1 | 30 | 14 | 16 | 0.019a |
|
pT2-4 | 102 | 25 | 77 |
|
| pN status |
|
|
|
|
|
pN0 | 85 | 27 | 58 | 0.452 |
|
pN1-N2 | 47 | 12 | 35 |
|
PDK1 is an independent prognostic
factor of NSCLC
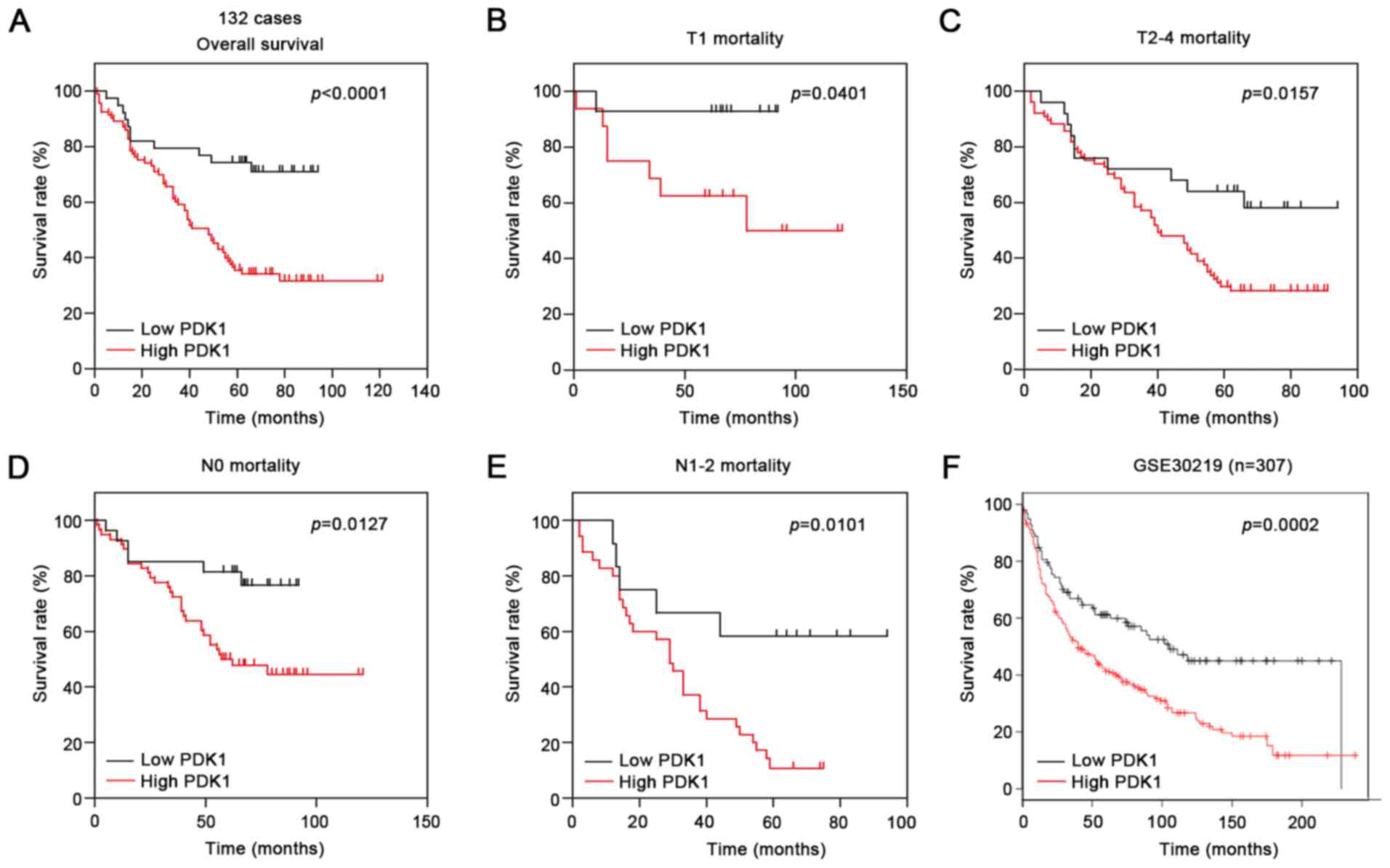
We explored the prognostic significance of PDK1
expression in NSCLC patients. All 132 NSCLC patients were divided
into two groups: high expression (high PDK1, n=93) and low
expression (low PDK1, n=38). Using Kaplan-Meier methods, we
determined that patients with PDK1 high staining revealed
significantly shorter survival rate compared with those with PDK1
low staining (Fig. 2A, P<0.0001,
log-rank test). We also performed univariate analysis to evaluate
the associations between prognosis and clinicopathological features
including age, gender, T stages, N stages, histological
differentiation, tumor size and PDK1 expression. Among these
features, advanced T stages (P=0.003), advanced N stages
(P<0.0001) and PDK1-positivity (P=0.001) were significantly
correlated with poor prognosis. Further multivariate regression
analysis revealed that advanced N stages (P=0.001) and
PDK1-positivity (P=0.001) were independent prognostic factors
(Table II).
| Table II.Univariate and multivariate analysis
for ADC overall survival. |
Table II.
Univariate and multivariate analysis
for ADC overall survival.
|
| Univariate
analysis | Multivariate
analysisa |
|---|
|
|
|
|
|---|
| Parameters | RR | 95% CI | P-value | RR | 95% CI | P-value |
|---|
| Age | 1.147 | 0.721–1.825 | 0.562 | – | – | – |
| Gender | 1.108 | 0.699–1.755 | 0.662 | – | – | – |
| Histological
differentiation | 1.166 | 0.837–1.625 | 0.363 | – | – | – |
| Tumor size | 1.306 | 0.767–2.224 | 0.326 | – | – | – |
| pT status | 3.080 | 1.475–6.432 | 0.003 | 1.754 | 0.795–3.871 | 0.164 |
| pN status | 2.594 | 1.631–4.124 | <0.0001 | 2.262 | 1.376–3.718 | 0.001 |
| PDK1
expression | 3.062 | 1.609–5.829 | 0.001 | 2.879 | 1.500–5.527 | 0.001 |
We then explored the correlations between PDK1
expression and prognosis using various subset analyzes. The results
revealed that PDK1 was associated with shorter survival regardless
of T stages or N stages (Fig.
2B-E). Moreover, we analyzed the PDK1 expression in GSE30219
and demonstrated that PDK1 could be a prognostic factor of
NSCLC.
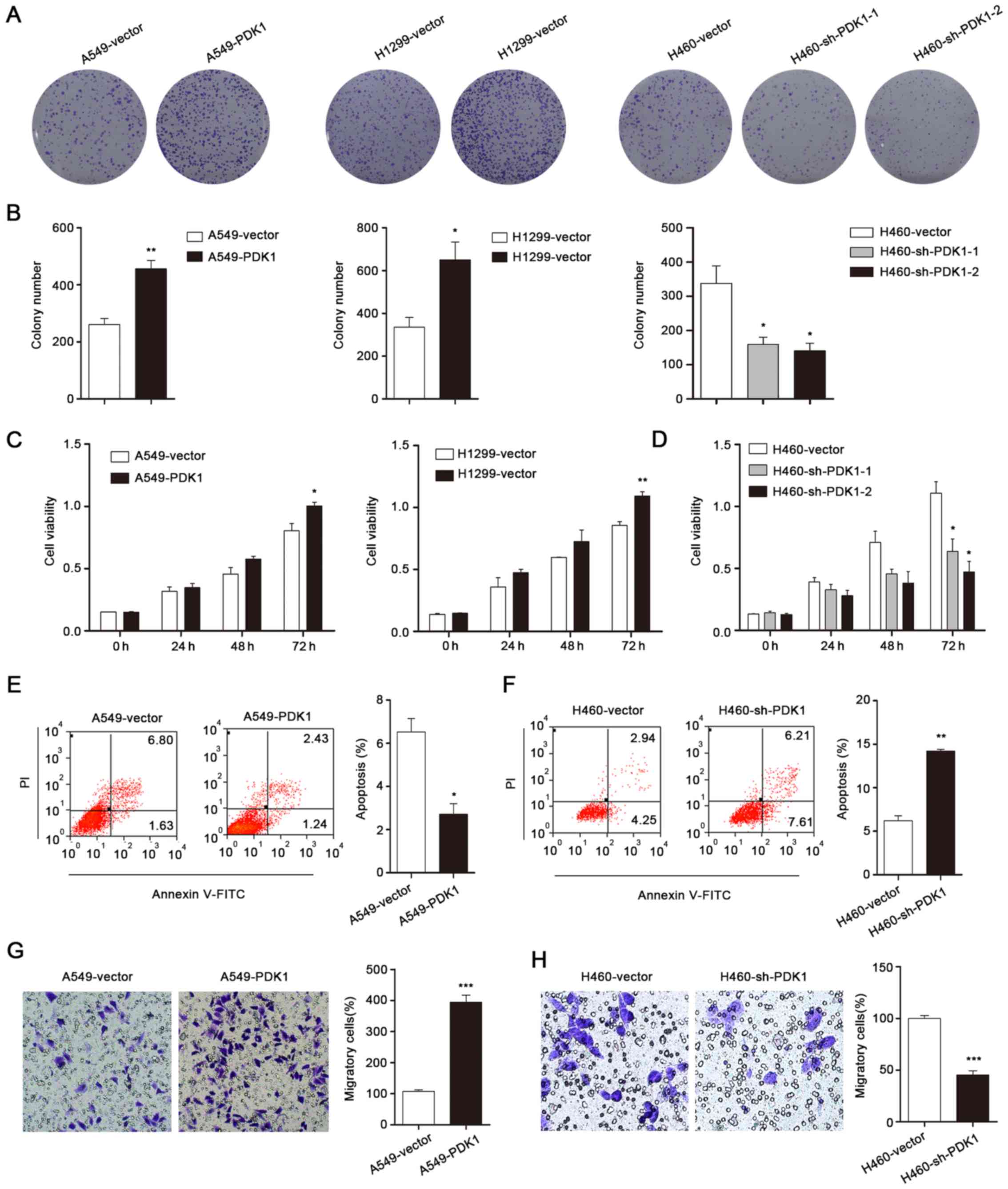
PDK1 promotes cell growth and inhibits
apoptosis
For subsequent experiments, we performed ectopic
expression of PDK1 in A549 and H1299 cell lines and knockdown of
PDK1 in H460 cells. The colony formation assay was carried out to
evaluate the functions of PDK1 on proliferation of NSCLC cell
lines. As shown in Fig. 3A and B,
the statistical results indicated that overexpression of PDK1 could
significantly promote cell growth while knockdown inhibited it.
Moreover, the CCK-8 assay was used to further determine the growth
effect of PDK1. The results of CCK-8 assay is shown in Fig. 3C and D, and were similar with colony
formation assays. Also we performed FACS to explore whether the
growth suppression was caused by apoptosis. Cell apoptosis was
decreased after PDK1 overexpression (Fig. 3E). Conversely, knocking down of PDK1
markedly increased the percentage of Annexin V-positive cells
(Fig. 3F). In addition, we employed
Transwell assay to examine the effect of PDK1 on cell mobility
(Fig. 3G and H). The number of
migratory cells with overexpression of PDK1 was increased
significantly compared to control cells. Collectively, we suggested
that PDK1 played oncogenic roles in NSCLC cells.
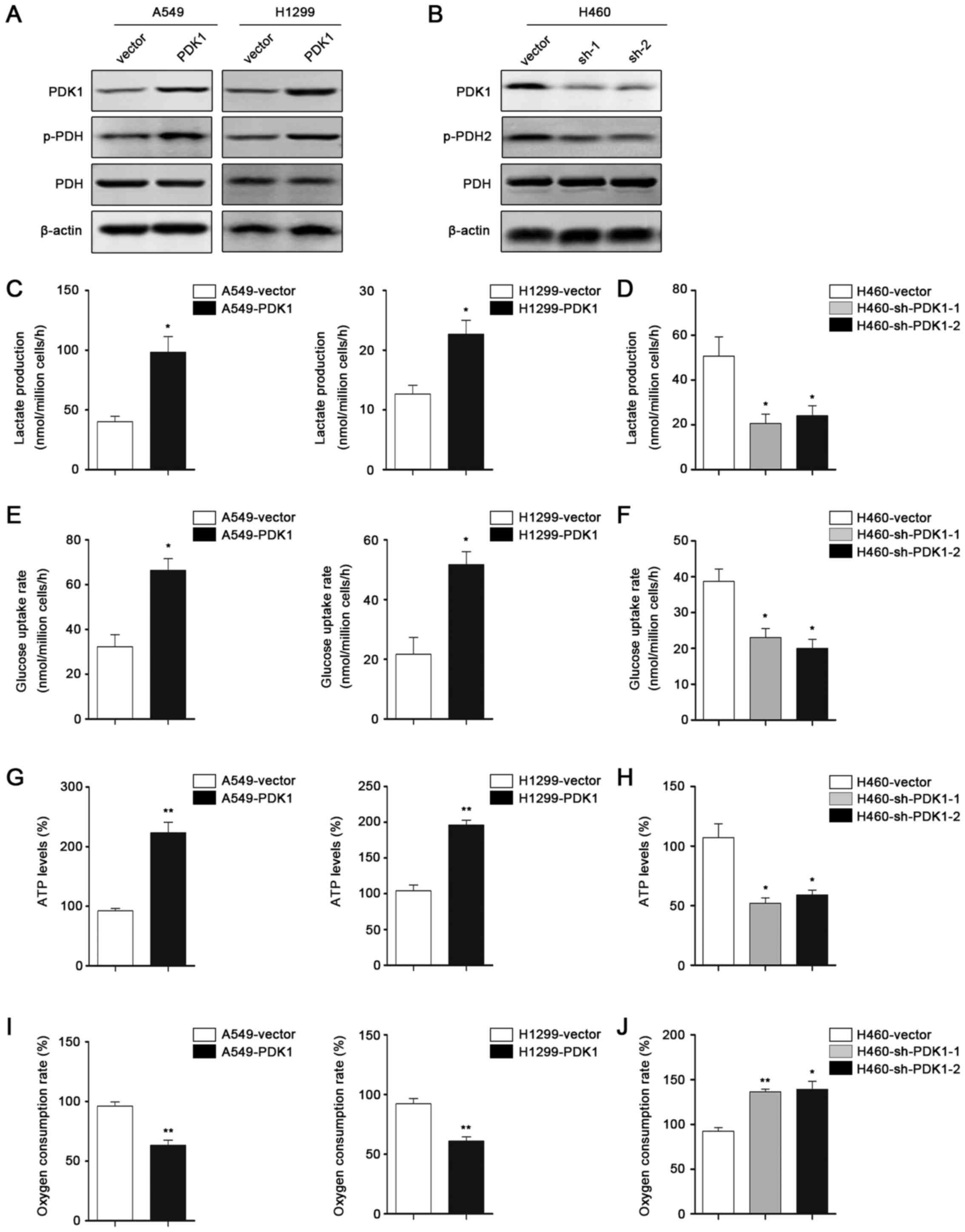
PDK1 modulates Warburg effect of NSCLC
cells
As PDK1 is a key enzyme in modulating tumor cell
glycolysis, we explored the function of PDK1 in regulating the
Warburg effect. Western blot analysis was performed to measure the
PDK1 overexpression or knockdown effects (Fig. 4A and B). Besides, p-PDH expression
was increased by PDK1 phosphorylation. Measurement of metabolic
parameters revealed that overexpression of PDK1 significantly
increased cellular glucose uptake and lactate production (Fig. 4C and E), while knockdown of PDK1
decreased both glucose uptake and lactate production (Fig. 4D and F). Moreover, forced expression
of PDK1 led to increased ATP levels and reduced O2
consumption (Fig. 4G and I).
Contrary results were observed when we knocked down PDK1 (Fig. 4H and J). Collectively, we suggested
that PDK1 could promote Warburg effect to facilitate cancer cell
proliferation.
Discussion
Lung cancer is one of the most common malignant
tumor which leads to over quarter of million deaths in America each
year, and the 5-year survival is unfavorable. The heterogeneity of
lung cancer is obvious, comprising adenocarcinoma, squamous cell
carcinoma, large cell carcinoma and small cell carcinoma (18). Hence, the diagnosis and clinical
treatments are much more difficult. NSCLC is a major subtype of
lung cancer (19). Numerous studies
have reported that metabolic reprogramming is a hallmark of many
tumors, such as hepatocellular carcinoma, colorectal carcinoma,
breast cancer and NSCLC (20,21).
So it is meaningful and urgent to identify new markers and
understand the mechanisms underlying the aberrant metabolism in
cancer cells.
In this study, we analyzed the gene expression
profiles in some published reports (22–24).
Through microarray, we revealed that the expression of PDK1 was
significantly upregulated in NSCLC tissues compared to normal
tissues. Also, the immunohistochemical staining in protein level
supported the mRNA results. In fact, PDK1 was reported to be
overexpressed in various types of cancers including acute myeloid
leukemia and colorectal carcinoma. Han et al reported that
miR-138, which could target PDK1, was found in sera and had
prognostic potential (25). These
results prompted us to further investigate PDK1 as an independent
prognostic factor of NSCLC. Our study implied that high expression
of PDK1 was associated with advanced T stage (75.5 vs 53.3% in
early stage) and poor survival rate. Moreover, we determined the
prognostic value of PDK1 by multivariate regression analysis. High
expression of PDK1 was associated with poor prognosis whether in T
stage or N stages.
PDK1 plays a critical role in regulating aerobic
glycolysis by phosphorylating pyruvate dehydrogenase. Also, other
substrates have been identified which were involved in tumor
progression. In the case of AKT, PDK1 phosphorylated it by
co-localizing in the plasma membrane (26). As for phosphorylation of S6K, SGK
and RSK, the progress is dependent in conformational change
(27,28). As a consequence, the ability of PDK1
to induce multiple key molecules and signaling might participate in
the development and progression of cancer. In this study, we
observed that overexpression of PDK1 increased the cell
proliferation rate and inhibited apoptosis while knockdown of PDK1
had the opposite effect. The impact might be due, at least
partially, to enhanced Warburg effect as we detected.
Warburg effect ws recognized decades ago and has
been paid attention to for a long time (29). The aberrant metabolic reprogramming
could confer tumor cell growth advantage, drug resistance and
metastasis not only by providing some raw molecules and precursors,
but also regulating the reactive oxygen species (ROS) and tumor
microenvironment (30,31). Here, we detected the lactate
production and glucose uptake with PDK1 overexpression or
knockdown. In addition, PDK1 inhibited tricarboxylic acid cycle
(TCA), however, as we observed, ATP level was increased with PDK1
overexpression. This could be due to the increased glucose uptake.
So these results supported that PDK1 regulated aerobic glycolysis
although there are still unknown factors that needed to be further
investigated.
In conclusion, PDK1 was demonstrated to be
overexpressed in NCSLS tissues and promoted cell proliferation and
migration by modulating the Warburg effect. Importantly, high
expression of PDK1 was significantly associated with advanced T
stage and could act as an independent prognostic factor of NSCLC.
Hence, further studies would be focused on the preferential
inhibition of PDK1 as a strategy for NSCLC therapy.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Reck M, Heigener DF, Mok T, Soria J-C and
Rabe KF: Management of non-small-cell lung cancer: Recent
developments. Lancet. 382:709–719. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Gainor JF, Varghese AM, Ou S-HI, Kabraji
S, Awad MM, Katayama R, Pawlak A, Mino-Kenudson M, Yeap BY, Riely
GJ, et al: ALK rearrangements are mutually exclusive with mutations
in EGFR or KRAS: An analysis of 1,683 patients with non-small cell
lung cancer. Clin Cancer Res. 19:4273–4281. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gasparini P, Cascione L, Landi L, Carasi
S, Lovat F, Tibaldi C, Alì G, D'Incecco A, Minuti G, Chella A, et
al: microRNA classifiers are powerful diagnostic/prognostic tools
in ALK-, EGFR-, and KRAS-driven lung cancers. Proc Natl Acad Sci
USA. 112:14924–14929. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fong MY, Zhou W, Liu L, Alontaga AY,
Chandra M, Ashby J, Chow A, O'Connor ST, Li S, Chin AR, et al:
Breast-cancer-secreted miR-122 reprograms glucose metabolism in
premetastatic niche to promote metastasis. Nat Cell Biol.
17:183–194. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang W, Zheng Y, Xia Y, Ji H, Chen X, Guo
F, Lyssiotis CA, Aldape K, Cantley LC and Lu Z: ERK1/2-dependent
phosphorylation and nuclear translocation of PKM2 promotes the
Warburg effect. Nat Cell Biol. 14:1295–1304. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Meijer TW, Kaanders JH, Span PN and
Bussink J: Targeting hypoxia, HIF-1, and tumor glucose metabolism
to improve radiotherapy efficacy. Clin Cancer Res. 18:5585–5594.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Gilkes DM, Bajpai S, Chaturvedi P, Wirtz D
and Semenza GL: Hypoxia-inducible factor 1 (HIF-1) promotes
extracellular matrix remodeling under hypoxic conditions by
inducing P4HA1, P4HA2, and PLOD2 expression in fibroblasts. J Biol
Chem. 288:10819–10829. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li J, Xu Y, Long X-D, Wang W, Jiao HK, Mei
Z, Yin QQ, Ma LN, Zhou AW, Wang LS, et al: Cbx4 governs HIF-1α to
potentiate angiogenesis of hepatocellular carcinoma by its SUMO E3
ligase activity. Cancer Cell. 25:118–131. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pullamsetti SS, Banat GA, Schmall A,
Szibor M, Pomagruk D, Hänze J, Kolosionek E, Wilhelm J, Braun T,
Grimminger F, et al: Phosphodiesterase-4 promotes proliferation and
angiogenesis of lung cancer by crosstalk with HIF. Oncogene.
32:1121–1134. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kaplon J, Zheng L, Meissl K, Chaneton B,
Selivanov VA, Mackay G, van der Burg SH, Verdegaal EM, Cascante M,
Shlomi T, et al: A key role for mitochondrial gatekeeper pyruvate
dehydrogenase in oncogene-induced senescence. Nature. 498:109–112.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sutendra G, Kinnaird A, Dromparis P,
Paulin R, Stenson TH, Haromy A, Hashimoto K, Zhang N, Flaim E and
Michelakis ED: A nuclear pyruvate dehydrogenase complex is
important for the generation of acetyl-CoA and histone acetylation.
Cell. 158:84–97. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sutendra G and Michelakis ED: Pyruvate
dehydrogenase kinase as a novel therapeutic target in oncology.
Front Oncol. 3:382013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hur H, Xuan Y, Kim YB, Lee G, Shim W, Yun
J, Ham IH and Han SU: Expression of pyruvate dehydrogenase kinase-1
in gastric cancer as a potential therapeutic target. Int J Oncol.
42:44–54. 2013.PubMed/NCBI
|
|
15
|
Schell JC, Olson KA, Jiang L, Hawkins AJ,
Van Vranken JG, Xie J, Egnatchik RA, Earl EG, DeBerardinis RJ and
Rutter J: A role for the mitochondrial pyruvate carrier as a
repressor of the Warburg effect and colon cancer cell growth. Mol
Cell. 56:400–413. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qin L, Tian Y, Yu Z, Shi D, Wang J, Zhang
C, Peng R, Chen X, Liu C, Chen Y, et al: Targeting PDK1 with
dichloroacetophenone to inhibit acute myeloid leukemia (AML) cell
growth. Oncotarget. 7:1395–1407. 2016.PubMed/NCBI
|
|
17
|
Koukourakis MI, Giatromanolaki A, Sivridis
E, Gatter KC and Harris AL: Tumor and Angiogenesis Research Group:
Pyruvate dehydrogenase and pyruvate dehydrogenase kinase expression
in non small cell lung cancer and tumor-associated stroma.
Neoplasia. 7:1–6. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li T, Kung H-J, Mack PC and Gandara DR:
Genotyping and genomic profiling of non-small-cell lung cancer:
Implications for current and future therapies. J Clin Oncol.
31:1039–1049. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Cooper WA, Lam DC, O'Toole SA and Minna
JD: Molecular biology of lung cancer. J Thorac Dis. 5:(Suppl 5).
S479–S490. 2013.PubMed/NCBI
|
|
20
|
Shimizu T, Inoue K, Hachiya H, Shibuya N,
Shimoda M and Kubota K: Frequent alteration of the protein
synthesis of enzymes for glucose metabolism in hepatocellular
carcinomas. J Gastroenterol. 49:1324–1332. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chiavarina B, Martinez-Outschoorn UE,
Whitaker-Menezes D, Howell A, Tanowitz HB, Pestell RG, Sotgia F and
Lisanti MP: Metabolic reprogramming and two-compartment tumor
metabolism: Opposing role(s) of HIF1α and HIF2α in tumor-associated
fibroblasts and human breast cancer cells. Cell Cycle.
11:3280–3289. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou J, Aerts J, den Hamer B, van Ijcken W,
den Bakker M, Riegman P, van der Leest C, van der Spek P, Foekens
JA, Hoogsteden HC, et al: Gene expression-based classification of
non-small cell lung carcinomas and survival prediction. PLoS One.
5:e103122010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Landi MT, Dracheva T, Rotunno M, Figueroa
JD, Liu H, Dasgupta A, Mann FE, Fukuoka J, Hames M, Bergen AW, et
al: Gene expression signature of cigarette smoking and its role in
lung adenocarcinoma development and survival. PLoS One.
3:e16512008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Rousseaux S, Debernardi A, Jacquiau B,
Vitte A-L, Vesin A, Nagy-Mignotte H, Moro-Sibilot D, Brichon P-Y,
Lantuejoul S, Hainaut P, et al: Ectopic activation of germline and
placental genes identifies aggressive metastasis-prone lung
cancers. Sci Transl Med. 5:186ra166-186ra166. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Han L, Zhang G, Zhang N, Li H, Liu Y, Fu A
and Zheng Y: Prognostic potential of microRNA-138 and its target
mRNA PDK1 in sera for patients with non-small cell lung cancer. Med
Oncol. 31:1292014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martelli AM, Nyåkern M, Tabellini G,
Bortul R, Tazzari PL, Evangelisti C and Cocco L: Phosphoinositide
3-kinase/Akt signaling pathway and its therapeutical implications
for human acute myeloid leukemia. Leukemia. 20:911–928. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Kang S, Dong S, Gu T-L, Guo A, Cohen MS,
Lonial S, Khoury HJ, Fabbro D, Gilliland DG, Bergsagel PL, et al:
FGFR3 activates RSK2 to mediate hematopoietic transformation
through tyrosine phosphorylation of RSK2 and activation of the
MEK/ERK pathway. Cancer Cell. 12:201–214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Dann SG, Selvaraj A and Thomas G: mTOR
Complex1-S6K1 signaling: At the crossroads of obesity, diabetes and
cancer. Trends Mol Med. 13:252–259. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Heiden MG Vander, Cantley LC and Thompson
CB: Understanding the Warburg effect: The metabolic requirements of
cell proliferation. Science. 324:1029–1033. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Upadhyay M, Samal J, Kandpal M, Singh OV
and Vivekanandan P: The Warburg effect: Insights from the past
decade. Pharmacol Ther. 137:318–330. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang F, Zhang H, Mei Y and Wu M:
Reciprocal regulation of HIF-1α and lincRNA-p21 modulates the
Warburg effect. Mol Cell. 53:88–100. 2014. View Article : Google Scholar : PubMed/NCBI
|