Introduction
Lung cancer is the most common cancer worldwide, as
well as the major cause of cancer-related deaths (1). Among all cases, ~80% are classified as
non-small cell lung cancer (NSCLC), among which adenocarcinoma
accounts for more than 50%. There have been several therapies that
have been developed for lung adenocarcinoma; however, the mortality
rate of lung adenocarcinoma has not largely improved (2). Therefore, it is of great importance to
search for prognostic biomarkers and novel targets to increase the
therapeutic effect.
Livin, an inhibitor of apoptosis protein (IAP), is
selectively overexpressed in certain tumors including lung
adenocarcinoma (3–5), and is correlated with resistance to
chemotherapy (6,7) and radiotherapy (8). Such as other IAP family proteins,
Livin is able to suppress apoptosis by binding certain caspases via
a baculoviral IAP repeat (BIR) domain and a COOH terminal RING
finger domain (9,10). Furthermore, various studies have
demonstrated that Livin plays an important role in tumorigenesis,
metastasis (11) and proliferation
(6) of lung adenocarcinoma cell
lines. However, whether Livin expression is associated with
clinical characteristics and prognosis of lung adenocarcinoma
remains elusive. Moreover, little information has been focused on
the regulation of Livin.
MicroRNAs (miRNAs) are small, non-coding RNA
molecules that post-transcriptionally regulate protein expression
by complementary base pairing (12,13).
miRNAs can contribute to tumorigenesis, cell migration,
invasiveness and metastasis via various signaling pathways, which
suggests their potential as therapeutic targets in cancer (14,15).
With the development of genetic sequencing technology,
differentially expressed miRNAs (DEMs) in lung cancer tissues vs.
non-cancerous lung tissues have been detected (16). Nevertheless, the miRNAs that
potentially target Livin RNA in lung adenocarcinoma are still
unclear.
In the present study, the expression level of Livin
in lung adenocarcinoma tissues vs. adjacent tissues was detected
and the correlation between the expression of Livin and the
clinical outcome was investigated. Bioinformatics analysis
suggested that 2 out of 61 DEMs, namely miR-198 and miR-515-5p,
were more likely to interact with Livin. Then, we identified the
interaction between miRNA-198 and Livin using human lung
adenocarcinoma cell line A549. Furthermore, we found that forced
expression of miR-198 suppressed tumor cell proliferation and
induced apoptosis of lung adenocarcinoma via the downregulation of
Livin and the activation of caspase-3. Finally, the effects of the
downregulation of Livin and the overexpression of miR-198 on A549
cell sensitivity to the cisplatin was detected using Cell Counting
Kit-8 (CCK-8) assays.
Materials and methods
Tissue samples
The paraffin section of lung adenocarcinoma and
their pericarcinous tissues (2 cm away from the tumor edge) were
obtained from patients who had undergone surgery at the Department
of Thoracic Surgery of The First Hospital of China Medical
University from July 2004 to June 2009. The follow-up data of 90
cases were cut-off in August 2014, and the overall survival for the
corresponding patients was calculated from the day of surgery to
the day of death or to the last follow-up. The experiments were
approved by the Ethics Committee of the First Hospital of China
Medical University.
Immunohistochemistry staining
To assess the relationship between the expression of
Livin and prognosis, formalin-fixed paraffin-embedded sections from
specimens of lung adenocarcinoma and the adjacent tissues were
evaluated by immunohistochemistry staining. The 4–5 µm
paraffin-embedded slides were deparaffinized with xylene and
rehydrated in graded alcohols. After high-pressure antigen
retrieval, the activity of endogenous peroxidase was blocked with
3% hydrogen peroxide in ethanol for 10 min. Subsequently, the
slides were washed with phosphate-buffered saline (PBS) and reacted
with a rabbit primary antibody against Livin (dilution 1:500;
Abcam, San Francisco, CA, USA) for 1 h. Then, incubation followed
with an HRP-labeled anti-rabbit IgG (Nichirei, Tokyo, Japan) for 30
min at room temperature. The sections were stained with
3,3-diaminobenzidine (DAB) used as the chromogen. Finally the
sections were counterstained with hematoxylin.
The degree of immunostaining was evaluated and
scored by two independent pathologists blinded to the clinical
information. Both cytoplasmic and nuclear immunostaining was
considered to be positive staining. The proportion of positive
cells was categorized as following: 5%, score 0; >6–25%, score
2; >26–50%, score 3; and >51%, score 4. The other score was
set on the basis of the extent of staining: no staining as 1; light
yellow staining as 2; medium brown staining as 3; brown staining as
4. The average of the two scores was interpreted as either low
expression (average <4) or high expression (average ≥4).
Bioinformatics analysis
The information of miRNA microarray was downloaded
from the NCBI Gene Expression Omnibus (GEO; http://www.ncbi.nlm.nih.gov/geo/) under series
GSE18692 and GPL4717. The detailed experimental design of the
microarray was previously elaborated (16). The known Homo sapien miRNAs
registered in miRBase (release 21) were selected to perform
analysis of miRNAs with differential expression using the Limma
package. Statistically significant differences in DEMs between
cancer and control groups were considered to exist at an adjusted
p-value <0.05 and |logFC| >1. The statistical tests were
carried out with the R program version 3.2.2 (http://www.r-project.org/). Two miRNA-target gene
databases, TargetScan (release 6.2) and MicroCosm 5, were used to
predict the miRNAs suppressing Livin.
Cell culture
The human lung adenocarcinoma cell line A549 was
obtained from the Cell Bank of the Institute of Biochemistry and
Cell Biology of the Chinese Academy of Sciences (Shanghai, China)
and was propagated in RPMI-1640 medium supplemented with 10% fetal
bovine serum (FBS) (both from Invitrogen, Carlsbad, CA, USA), 100
IU/ml penicillin and 100 µg/ml streptomycin, at 37°C in an
atmosphere of 5% CO2.
Synthetic RNA oligonucleotides and
transient transfection
All the miRNA inhibitors, miRNA mimics, nonsense
sequence as miRNA negative control (NC), short hairpin RNA of Livin
(sh-Livin) and short hairpin RNA negative control (NC-shRNA) were
chemically synthesized by GenePharma (Shanghai, China). All of the
transfections in the present study were transient, using JetPRIME
reagent (PolyPlus Transfections SA, Illkirch, France) according to
the manufacturers protocol. The cells were not harvested for the
subsequent assays until 48 h after RNA oligonucleotide
transfection.
RNA extraction and quantitative
reverse transcription-PCR (qRT-PCR)
Total RNA was extracted from A549 using TRIzol
reagent (Takara, Otsu, Japan). Complementary DNA (cDNA) of miR-198
and miR-515-5p were obtained with TransScript® miRNA
First-Strand cDNA Synthesis (TransGen Biotech, Beijing, China). The
transfection efficiency of miRNAs was assessed by quantitative
real-time PCR (qPCR) with SYBR-Green qPCR Master Mix (Takara) on an
ABI 7500 Fast System thermocycler (Applied Biosystems, Foster City,
CA, USA). All the experiments were conducted in accordance with the
manufacturer's instructions. Triplicate reactions were performed
and the data were normalized to U6 and calculated with the
2−ΔΔCt method. The involved primers are described as
follows: miR-198 forward, 5′-GCCAACTGGTCCAGAGGG-3′; miR-515-5p
forward, 5′-TTCTCCAAAAGAAAGCACTTTCTG-3′; U6 forward,
5′-CGCTTCACGAATTTGCGTGTCAT-3′; the universal reverse primers of
miRNAs from the kit.
Protein extraction and western blot
analysis
Forty-eight hours after transfection, the cells were
lysed using cell lysis buffer supplemented with protease inhibitors
and phenylmethylsulfonyl fluoride (PMSF) (Beyotime, Zhejiang,
China). Total proteins were extracted by centrifugation at 12,000 ×
g and 4°C for 20 min. The protein concentration was assessed using
the BCA protein assay kit (Beyotime) and equal amounts of total
proteins were separated in 10% SDS-PAGE and transferred onto
nitrocellulose membranes. The membranes were blocked for 1 h with
5% non-fat milk powder in Tris-Buffered saline containing 0.5%
Tween-20, and then incubated with a primary antibody overnight at
4°C, followed by washing and incubation with a secondary antibody
for 2 h at room temperature. Finally, the membranes were detected
by enhanced chemiluminescence (ECL) plus western blot detection
reagents (Thermo Fisher Scientific, Inc., Waltham, MA, USA). The
antibodies (Thermo Fisher Scientific, Inc.) were used according to
the manufacturers instructions, and were as follows: primary
antibodies against GAPDH (ab9485), Livin (ab97350), caspase-3
(ab32351) and the HRP-conjugated secondary antibody (ab6721).
Dual-luciferase assays
To confirm whether miR-198 or miR-515-5p can
interact with Livin mRNA, pGLO Dual-Luciferase miRNA Target
Expression Vector pmirGLO-wt-Livin (wild-type) and pGLO-mut-Livin
(mutant type) of miR-198, pGLO-wt-Livin (wild-type) and
pGLO-mut-Livin (mutant type) of miR-515-5p were constructed by
GeneChem (Shanghai, China). The pGLO Dual-Luciferase miRNA Target
Expression Vector plasmids (mutant or wild-type) and the internal
control Renilla luciferase plasmid pRL-TK were cotransfected
with miRNAs (miR-198 and miR-515-5p mimics, or NC) with a ratio of
20:1, using JetPRIME reagent into A549. The luciferase activity of
firefly and Renilla luciferase was assessed 24 h after
cotransfection, using the Dual-Luciferase® Reporter
Assay System (Promega, Madison, WI, USA), complying with the
manufacturer's protocol. To aid comparisons, values for cells with
the NC + mut-Livin 3′-UTR group were set equal to 1.
Cell proliferation assays
Cell proliferation was detected using CCK-8 (KeyGen
Biotech, Jiangsu, China). The A549 cells in the logarithmic phase
with or without transfection were seeded in 96-well plates at a
density of 5×103 cells/well. CCK-8 (10 µl/well) was
added at various time-points (0, 24, 48 and 72 h) and incubated at
37°C for 2 h. The absorbance of each well at 450 nm was measured
using a microplate reader (Bio-Rad, Hercules CA, USA) to assess the
number of viable cells. To decrease random errors, all of the
procedures were replicated at least three times.
Cell cycle distribution and apoptosis
analysis
Flow cytometric analyses were utilized to assess
cell apoptosis and cell cycle distribution using a flow cytometer
(Bio-Rad). The A549 cells were synchronized before the cell cycle
assays. After 48 h of transfection, the cells, which were cultured
in 6-well plates and prepared for apoptosis examination and cell
cycle distribution, were harvested by trypsin and detected using an
Annexin V-FITC/PI apoptosis detection kit and a Cell Cycle
Detection kit (both from KeyGen Biotech) respectively, according to
the manufacturers recommendations. Each experiment was performed at
least in triplicate.
Drug sensitivity assays
Before each experiment, the chemotherapeutic drug
cisplatin (F.H. Faulding & Co, Mulgrave Victoria, Australia)
was dissolved in 0.9% sodium chloride, and then diluted with fresh
medium. The concentration of cisplatin was set as 0.01, 0.1 and 1
µmol/l. The cell proliferation rate 48 h after transfection and
drug treatment was detected by CCK-8 assay kits. Furthermore, the
cell proliferation rate treated with 1 µmol/l of cisplatin was
assessed every 24 h for a period of 72 h.
Statistical analysis
The results of immunohistochemical staining were
analyzed using Chi-square test, Kaplan-Meier analysis, Spearman
rank correlation and Cox regression analysis. The result of the
Kaplan-Meier survival analysis was compared with the log-rank test.
Other experiments were analyzed using Student's t-test and
expressed as the mean ± standard deviation (SD). Statistical
analyses were processed using SPSS 23.0 software (SPSS, Inc.,
Chicago, IL, USA) and p<0.05 was considered as statistically
significant.
Results
Livin is overexpressed in lung
adenocarcinoma tissues
The expression of Livin in cancerous and
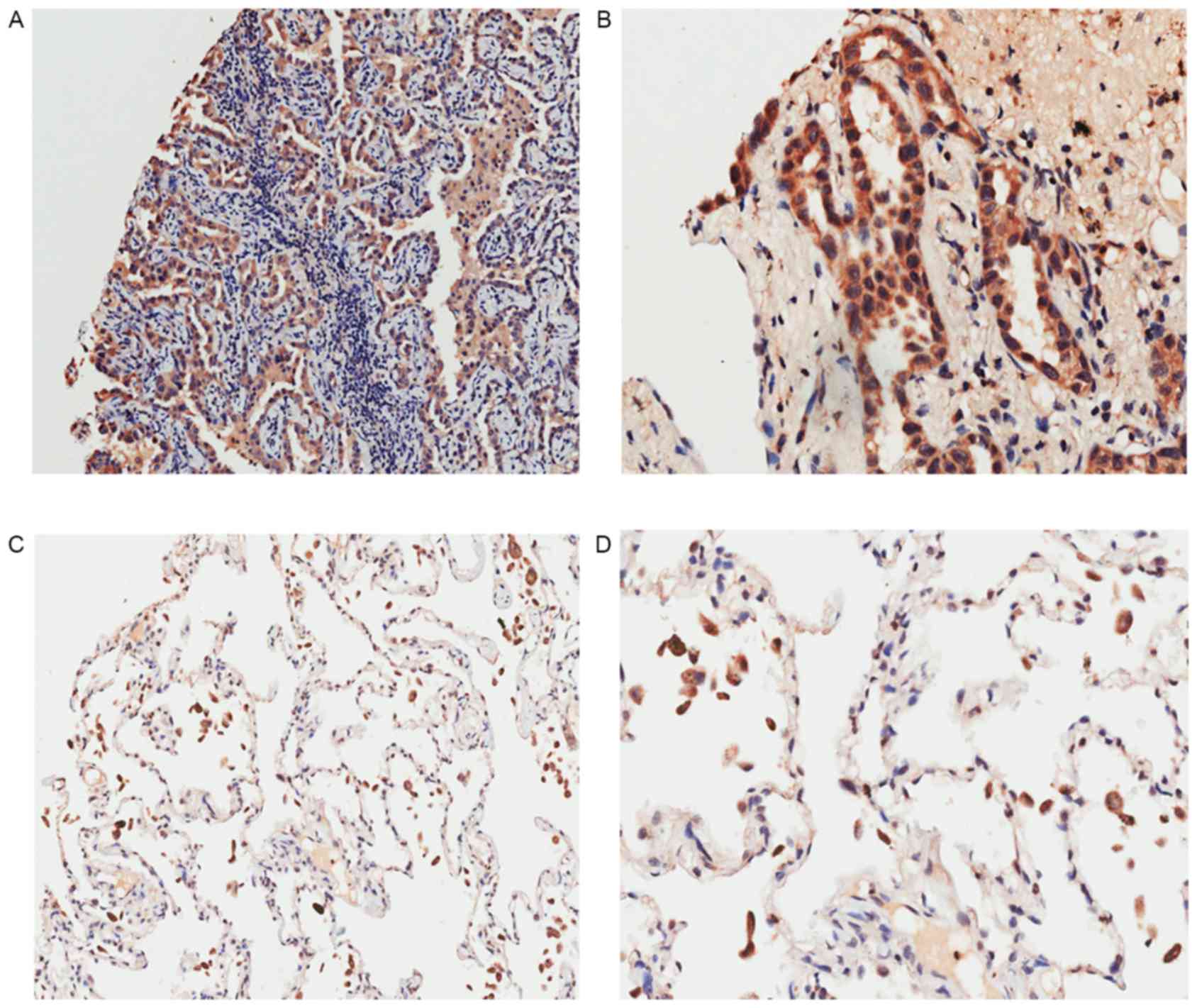
pericancerous tissues is shown in Fig.
1 and Table I. According to the
immunohistochemical results, Livin was mainly expressed in the
cytoplasm and notably overexpressed in cancerous tissues
(p<0.05). Spearman's rank correlation analysis was performed to
display the correlations between the expression of Livin and the
clinicopathological characteristics such as sex, age,
tumor-node-metastasis (TNM) stage and pathological grade (Table II). Notably, it was determined that
the expression of Livin in patients who suffer from lymph node
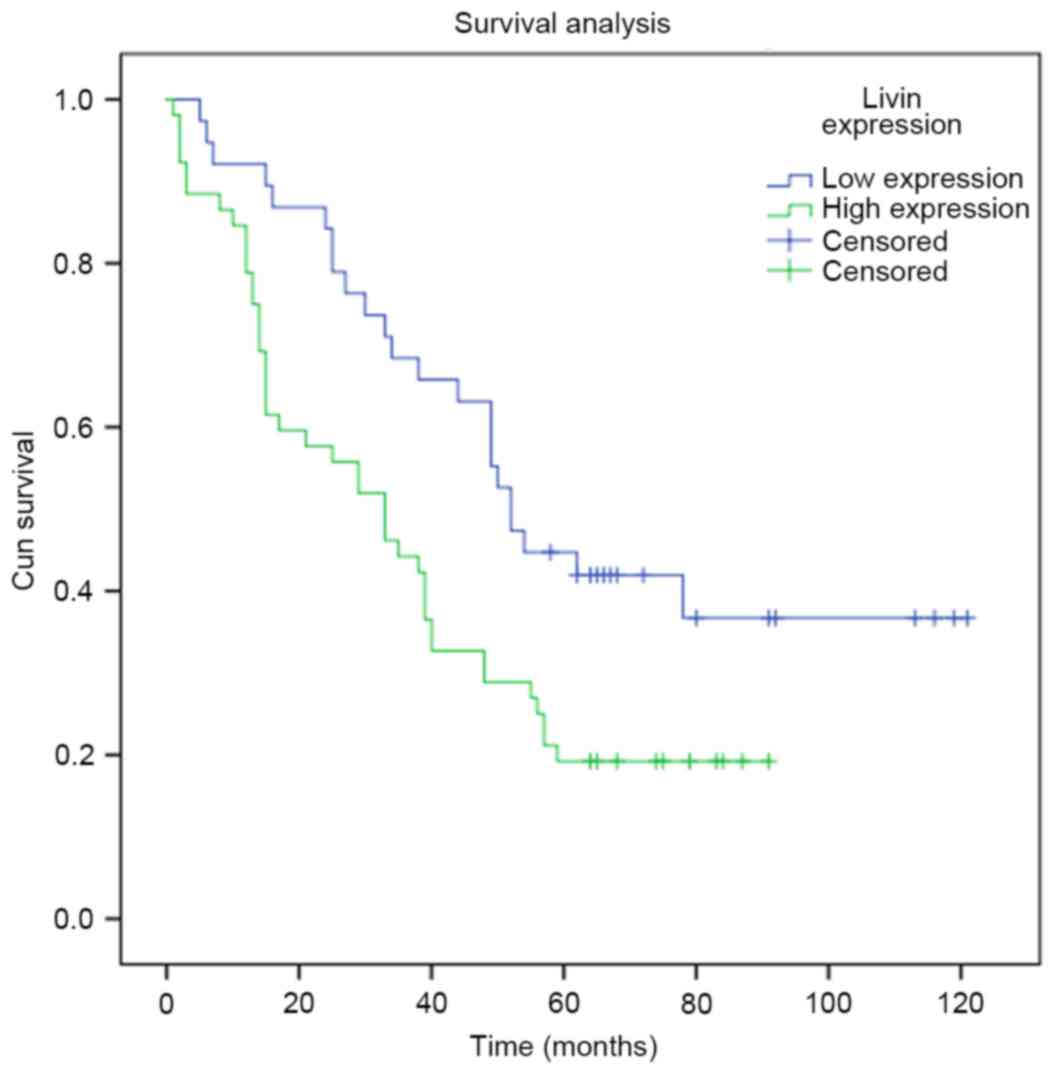
metastasis was elevated (p<0.05). Survival analysis revealed
that lung adenocarcinoma patients with lower expression of Livin
had a higher survival rate than those with higher Livin expression
(Fig. 2; p<0.05). Moreover,
univariate analyses indicated that Livin expression, along with T,
N and TNM stage and grade, all play important roles in the
prognosis of lung adenocarcinoma. Furthermore, these variables with
statistical significance in the univariate analyses were included
in a multivariate regression analysis, and Livin expression and N
stage were identified as significant independent prognostic factors
of lung adenocarcinoma prognosis (Table III).
 | Table I.Differential expression of Livin in
cancerous and pericancerous tissues. |
Table I.
Differential expression of Livin in
cancerous and pericancerous tissues.
|
|
| Livin expression |
|
|
|---|
|
|
|
|
|
|
|---|
| Tissues | n | High(%) | Low(%) | Chi-square value | P-value |
|---|
| Cancerous | 90 | 52 (57.8) | 38 (42.2) | 34.545 |
<0.0001a |
| Pericarcinous | 90 | 14 (15.6) | 76 (84.4) |
|
|
| Table II.Correlation between Livin expression
and clinicopathological characteristics. |
Table II.
Correlation between Livin expression
and clinicopathological characteristics.
|
| Livin
expression |
|
|
|
|---|
|
|
|
|
|
|
|---|
| Variables | High | Low | Total | rs | P-value |
|---|
| Sex |
|
|
| −0.059 | 0.579 |
|
Female | 25 | 16 | 41 |
|
|
|
Male | 27 | 22 | 49 |
|
|
| Age (years) |
|
|
| −0.104 | 0.327 |
|
≤60 | 26 | 15 | 41 |
|
|
|
>60 | 26 | 23 | 49 |
|
|
| Grade |
|
|
| 0.161 | 0.129 |
| 1 | 4 | 8 | 12 |
|
|
| 2 | 43 | 27 | 70 |
|
|
| 3 | 5 | 3 | 8 |
|
|
| T stage |
|
|
| 0.087 | 0.413 |
| T1 | 9 | 8 | 17 |
|
|
| T2 | 28 | 22 | 50 |
|
|
| T3 | 11 | 6 | 17 |
|
|
| T4 | 4 | 2 | 6 |
|
|
| N stage |
|
|
| 0.239 | 0.023a |
| N0 | 19 | 23 | 42 |
|
|
| N1 | 16 | 8 | 24 |
|
|
| N2 | 12 | 6 | 18 |
|
|
| N3 | 5 | 1 | 6 |
|
|
| M stage |
|
|
| 0.033 | 0.755 |
| M0 | 50 | 37 | 87 |
|
|
| M1 | 2 | 1 | 3 |
|
|
| TNM stage |
|
|
| 0.175 | 0.099 |
| I | 16 | 15 | 31 |
|
|
| II | 11 | 13 | 24 |
|
|
|
III | 23 | 9 | 32 |
|
|
| IV | 2 | 1 | 3 |
|
|
| Table III.Univariate and multivariate analyses
of the factors correlated with overall survival of lung
adenocarcinoma patients. |
Table III.
Univariate and multivariate analyses
of the factors correlated with overall survival of lung
adenocarcinoma patients.
|
| Univariate
analysis | Multivariate
analysis |
|---|
|
|
|
|
|---|
| Variables | HR | 95% CI | P-value | HR | 95% CI | P-value |
|---|
| Livin
expression | 0.466 | 0.281–0.773 | 0.003a | 0.568 | 0.339–0.950 | 0.031a |
| Sex | 1.393 | 0.866–2.243 | 0.172 |
|
|
|
| Age | 1.107 | 0.690–1.776 | 0.674 |
|
|
|
| T stage | 1.541 | 1.135–2.092 | 0.006a | 1.170 | 0.799–1.715 | 0.419 |
| N stage | 1.788 | 1.404–2.276 |
<0.001a | 1.565 | 1.107–2.211 | 0.011a |
| M stage | 0.700 | 0.171–2.863 | 0.620 |
|
|
|
| TNM stage | 1.562 | 1.223–1.995 |
<0.001a | 1.031 | 0.679–1.564 | 0.886 |
| Grade | 1.969 | 1.218–3.182 | 0.006a | 1.697 | 0.953–3.022 | 0.072 |
Prediction of putative miRNAs
targeting Livin
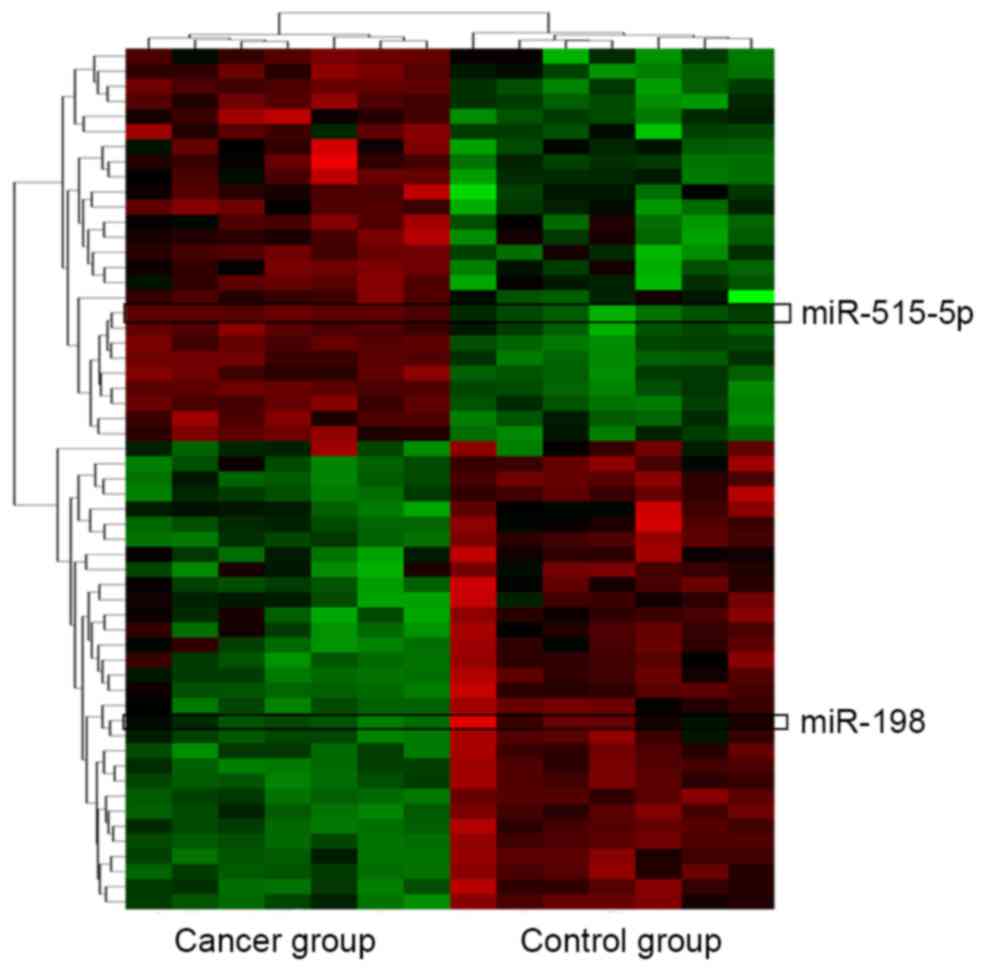
Forty-five putative miRNAs generated by two
databases, were identified as the possible miRNAs targeting Livin
(Table IV). Among the 45 miRNAs,
only two of them, miR-198 and miR-515-5p were significantly
differentially expressed in lung adenocarcinoma tissues vs.
non-cancerous lung tissues according to the miRNA microarray
analysis (Fig. 3).
| Table IV.The possible miRNAs targeting
Livin. |
Table IV.
The possible miRNAs targeting
Livin.
| hsa-miR-940 | hsa-miR-323-5p |
|
hsa-miR-548b-3p | hsa-miR-24 |
| hsa-miR-198 | hsa-miR-324-5p |
| hsa-miR-25* | hsa-miR-148b* |
| hsa-miR-214 | hsa-miR-652 |
| hsa-miR-455-3p | hsa-miR-525-5p |
|
hsa-miR-516a-5p | hsa-miR-582-5p |
| hsa-miR-296-3p | hsa-miR-183 |
| hsa-miR-141 | hsa-miR-139-5p |
| hsa-miR-188-5p | hsa-miR-486-3p |
| hsa-miR-515-3p | hsa-miR-551a |
| hsa-miR-552 | hsa-miR-598 |
| hsa-miR-572 | hsa-miR-601 |
| hsa-miR-593* | hsa-miR-541 |
| hsa-miR-596 | hsa-miR-296-5p |
| hsa-miR-149* | hsa-miR-432* |
|
hsa-miR-548d-3p | hsa-miR-615-3p |
| hsa-miR-942 | hsa-miR-654-5p |
| hsa-miR-874 | hsa-miR-660 |
| hsa-miR-222* | hsa-miR-7 |
| hsa-miR-519e | hsa-miR-512-3p |
| hsa-miR-423-5p | hsa-miR-589* |
| hsa-miR-662 |
|
Livin is a direct target of
miR-198
The aberrant expression of miR-198 and miR-515-5p in
lung adenocarcinoma was reported as previously described (17,18).
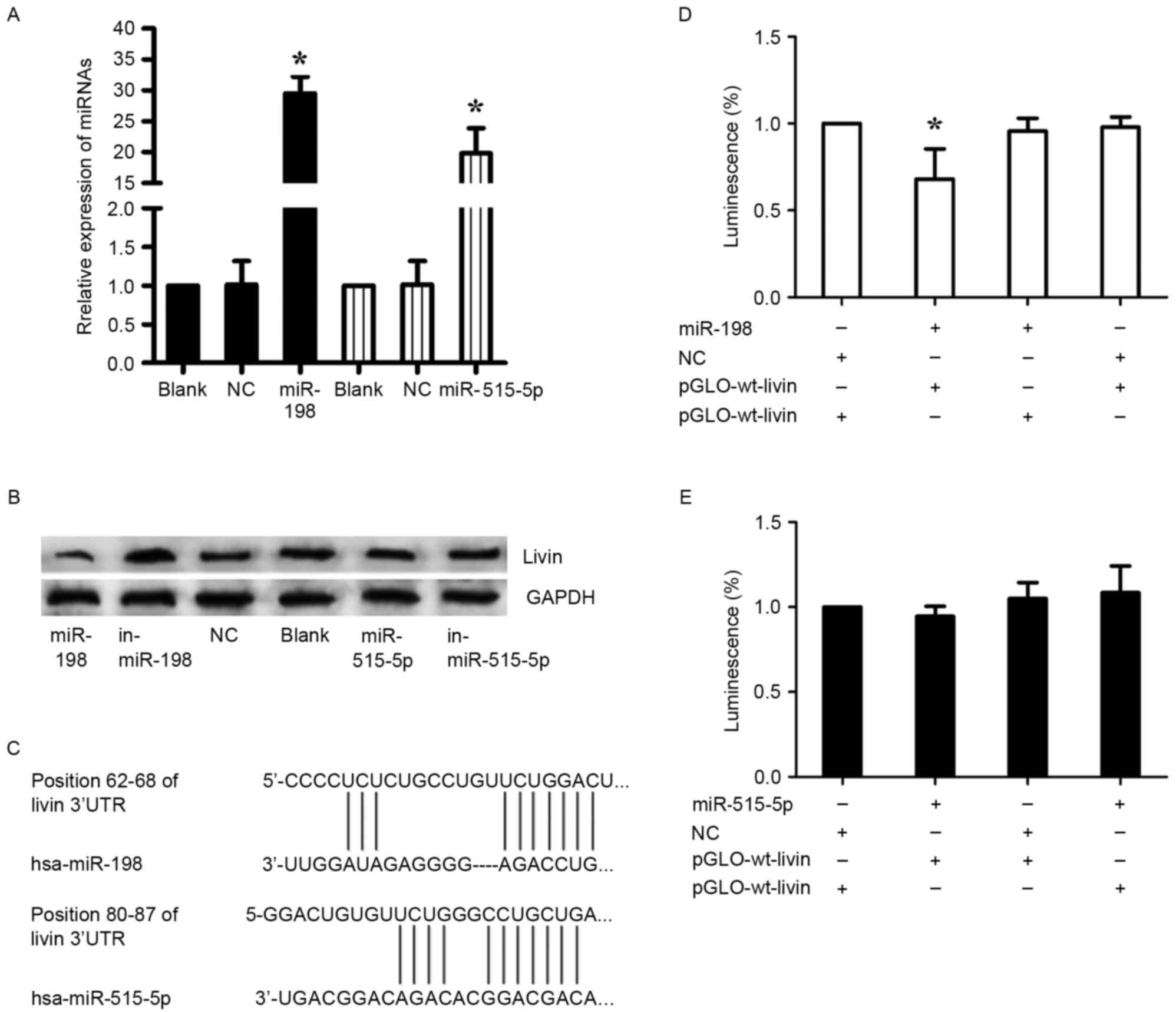
To confirm the prediction, miR-198 mimics and inhibitor
(in-miR-198), miR-515-5p mimics and inhibitor (in-miR-515-5p),
nonsense sequence as NC were transfected into A549 cells. Before
the transfection, the transfection efficiency was detected by
RT-qPCR (Fig. 4A). In consequence,
we found that miR-198 expression revealed a completely contrary
trend to Livin expression, whereas this phenomenon was not observed
in the regulation of miR-515-5p, inferring that miR-198 may be a
specific miRNA targeting Livin in A549 cells (Fig. 4B). Furthermore, luciferase reporter
assays were employed to illuminate the interaction of miR-198 or
miR-515-5p with Livin mRNA, and the predicted binding sites are
shown in Fig. 4C. Either
pGLO-wt-Livin or pGLO-mut-Livin of miR-198 and pRL-TK were
cotransfected into A549 cells with miR-198 or NC. As with miR-198,
the luciferase reporter assays of miR-515-5p were performed. A
significant decrease of the relative luciferase activity was
observed between the miR-198 + pGLO-wt-Livin compared to the other
groups (p<0.05) (Fig. 4D). In
stark contrast, there was no significant change in the relative
luminescence intensities in the miR-515-5p + luciferase reporter
gene groups (Fig. 4E). Therefore,
it is miR-198, but not miR-515-5p that bound directly to the 3′-UTR
of Livin mRNA and negatively regulated the expression of Livin in
A549 cells.
Impact of miR-198 restoration on the
proliferation, apoptosis and cell cycle of A549 cells
The effects of overexpressing or silencing Livin in
A549 cells have been previously demonstrated in detail (10,11).
To investigate the role of miR-198 in A549 cells, miR-198 and NC
were separately transfected into 6-well plates and the CCK-8 assays
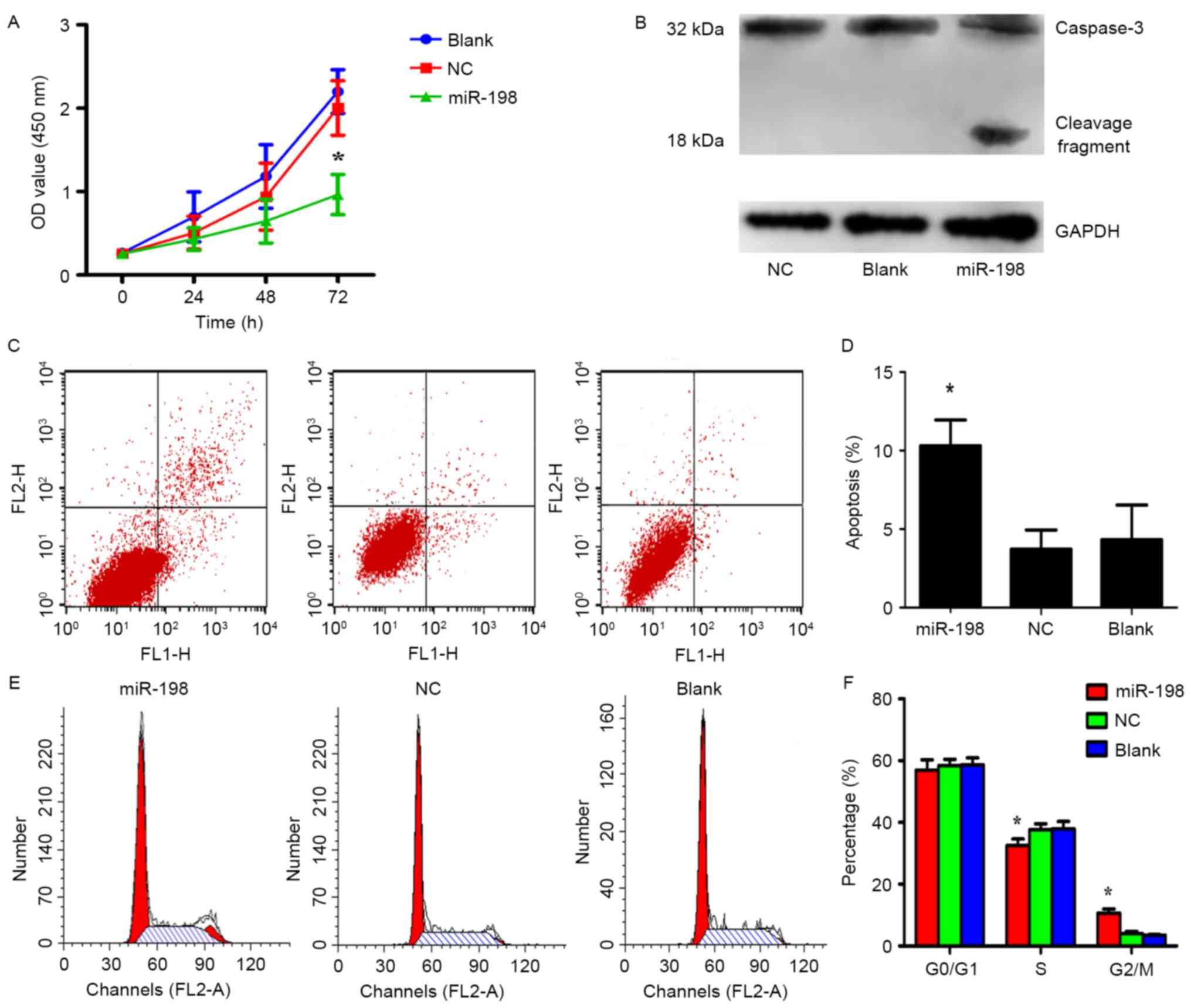
were performed to detect cell proliferation. The results revealed
that the overexpression of miR-198 greatly inhibited cell
proliferation in comparison to the control group, respectively
(Fig. 5A; p<0.05). In order to
elucidate the mechanism of miR-198 in the inhibition of cell
proliferation, apoptosis and cell cycle assays were performed with
miR-198 or NC transfected A549 cells. Flow cytometric analysis
indicated that the percentage of apoptotic cells in the miRNA-198
group was significantly higher than that in both control groups
(Fig. 5C and D; p<0.05).
Furthermore, we found that the miR-198 group had a lower percentage
of S phase cells and a higher ratio of G2/M phase cells than the NC
or blank groups (Fig. 5E and F;
p<0.05). This result indicated that miR-198 induced cell cycle
arrest in the G2/M phase. Nevertheless, proliferation which is
determined by the S + G2/M phases appeared to not be affected,
possibly due to the synchronization of the cells before the
experiments. From this point of view, the diminished cell growth
was attributed to apoptosis due to synchronization of the cells
prior to the experiment. These data demonstrated that low
expression of miR-198 exerts tumor-promoting effects in A549 cells,
therefore their re-expression may benefit epigenetic cancer
therapy, consistent with the outcomes of silencing Livin.
Livin has been demonstrated to be capable of
inhibiting apoptosis by binding to caspase family members directly
with its BIR domain, and then suppressing caspase activities
(9). In order to further ascertain
whether miR-198 induces apoptosis through downregulation of Livin,
miR-198 was overexpressed in A549 cells and the expression of
caspase-3, a member of the caspase family, was observed. Western
blot analysis revealed that the activation of caspase-3
significantly increased in the miR-198 groups, implying that
miR-198 may inhibit cell proliferation by decreasing the expression
of Livin which activates caspase-3 directly (Fig. 5B).
In addition, we determined the correlation between
Livin and lymph node status, which had been reported by Lin et
al. Thus, we attempted to explore whether miR-198 was related
to cell migration and invasion. Preliminary experiments revealed
that miR-198 did not markedly impact cell migration and invasion,
therefore, we finally chose to investigate proliferation and
apoptosis, which may be primarily associated with miR-198.
The effect of miR-198 overexpression
and silencing of Livin on cisplatin chemosensitivity in A549
cells
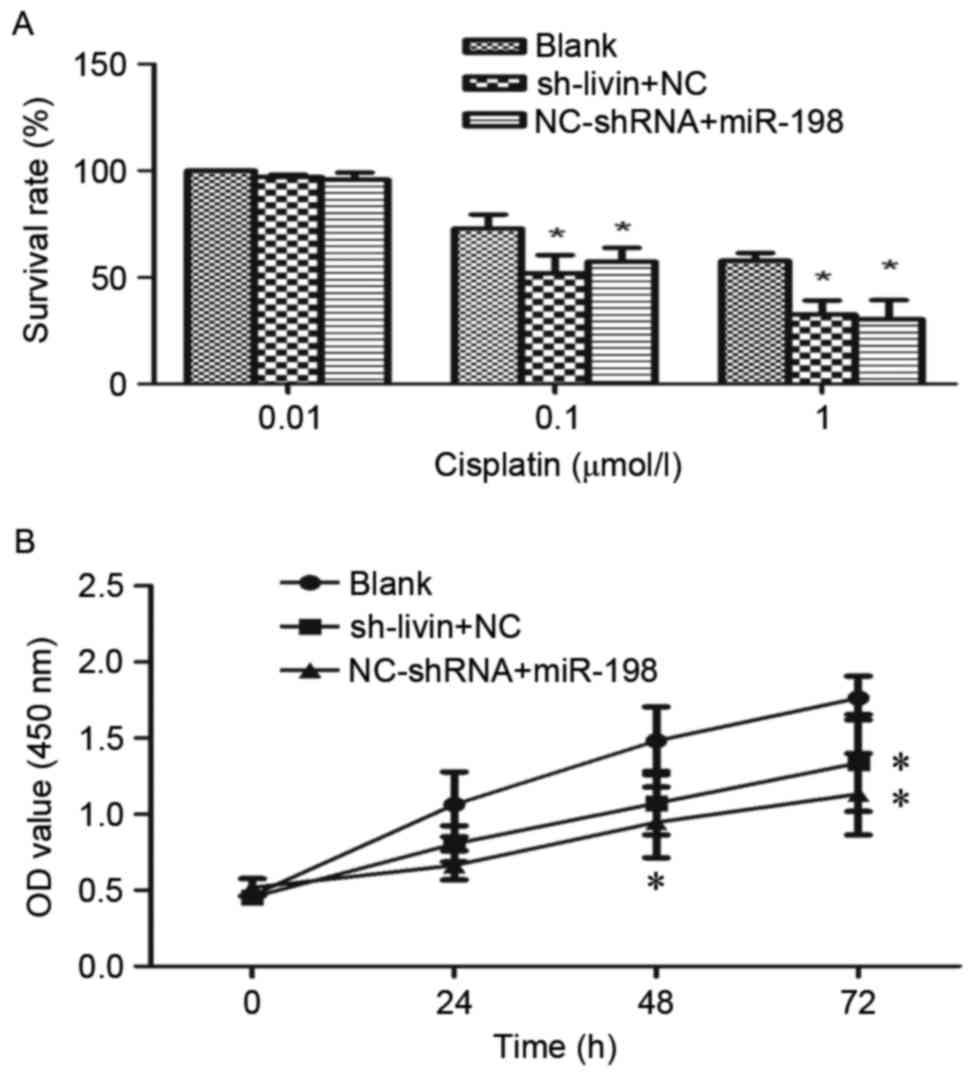
Livin RNA interference has been identified to
enhance the chemosensitivity to chemotherapeutic drugs in NSCLC
cells (6,7). To investigate whether miR-198 ectopic
expression could increase the sensitivity of A549 cells to
cisplatin, sh-Livin or NC-shRNA were cotransfected with miR-198 or
NC into A549 cells. The proliferation rate of these cells was
assessed by CCK-8 assays. As shown in Fig. 6A and B, the proliferation rate in
dose- and time-dependent manners over a 72-h treatment period were
observed. On the one hand, from the dose-survival bar chart, it was
evident that cisplatin had a dose-dependent suppressive effect on
cell growth. Both miR-198 and sh-Livin may enhance the
chemosensitivity of A549 cells to cisplatin significantly, and have
a similar efficiency (Fig. 6A). On
the other hand, we found that the proliferation rate was
significantly slower in sh-Livin + NC and NC-shRNA + miR-198 group
by the end of the 72-h treatment time (Fig. 6B). Therefore, the combination of
chemotherapy and silencing of Livin or miR-198 ectopic expression
may enhance the treatment of cisplatin-resistant lung
adenocarcinoma patients.
Discussion
To the best of our knowledge, this is the first
study elucidating the relationship between Livin and the
clinicopathological features in lung adenocarcinoma patients. Livin
is a member of the IAPs family, and is also known as baculoviral
IAP repeat-containing protein 7 (BIRC7). A high level of Livin has
been found in various tumors, including NSCLC (3,5,9) and is
considered of great importance in the regulation of cell
proliferation, metastasis, the cell cycle and apoptosis in lung
adenocarcinoma via the JNKI signaling pathway, the VEGF and MMP
pathways or the activity of the caspase family (11,19,20).
Notably, our results revealed that a high expression of Livin in
cancer tissues was related with the lung adenocarcinoma patient
metastasis. Moreover, it has been demonstrated that silencing of
Livin in lung adenocarcinoma cell lines enhances the sensitivity of
chemotherapy or radiotherapy (6–8,21).
Livin takes part in the networks involved in the regulation of cell
functions and influences the curative effect of lung adenocarcinoma
through cross-communication, making it promising for targeted
therapy. In the present study, immunohistochemical staining
analysis revealed that the expression of Livin may be associated
with the prognosis and lymph node metastasis of lung
adenocarcinoma. From this perspective, Livin may function as a
therapeutic agent as well as a prognostic indicator for lung
adenocarcinoma. However, the number of patients in the present
study may be insufficient to draw an definite conclusion. In
consideration of this limitation, more samples from lung
adenocarcinoma patients at various stages and their follow-up data
should be collected. However, it may be necessary to perform in
vivo experiments and further investigate the underlying
mechanisms of Livin.
Nonetheless, to date, only Yuan et al has
reported that the transcriptional signaling pathway of Livin is
regulated by β-catenin/TCF signaling, and there is no more research
concerning the upstream regulation of Livin in lung adenocarcinoma
(22). To explore the molecular
mechanism of the aberrant expression of Livin, we employed
databases and the results of microRNA (miRNA) microarray to predict
the miRNAs involved with Livin regulation. The data indicated that
miR-198 and miR-515-5p may be miRNAs that can influence the
expression of Livin. The lung adenocarcinoma cell line A549 was
selected for the experiments due to the fact that it has the
highest expression of Livin compared with other lung adenocarcinoma
cell lines (3). The present study,
demonstrated that Livin is a target of miR-198 and low expression
of miR-198 may be responsible for the upregulation of Livin in lung
adenocarcinoma. Furthermore, it is worth noting that the effect of
miR-198 upregulation is not only consistent with that of Livin
downregulation, but can also activate caspase-3, which is one of
the key factors in apoptotic pathways and can be inhibited by
Livin. Moreover, we found a new phenomenon where miR-198 can
contribute to chemosensitivity by neutralizing Livin. In fact,
miR-198 has been found to be closely related with lung carcinoma
(17,23), and many oncogenes have been
identified as targets of miR-198, such as MCL1 (24), ROCK1 (25), FUT8 (26) and FGFR1 (23).
In general, we demonstrated that Livin, an apoptotic
pathway suppressor, can enhance the growth of lung adenocarcinoma,
which may be partly due to the downregulation of miR-198. Livin has
the potential to be a biomarker for predicting the prognosis of
lung adenocarcinoma and focusing on the miR-198/Livin/caspase-3
regulatory network may become a promising strategy for drug
resistant lung adenocarcinoma therapy.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (grant no. 81372876), and the
Liaoning Province Natural Science Foundation (grant no.
2013021041).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Diaz-Garcia CV, Agudo-Lopez A, Perez C,
López-Martín JA, Rodríguez-Peralto JL, de Castro J, Cortijo A,
Martínez-Villanueva M, Iglesias L, García-Carbonero R, et al:
DICER1, DROSHA and miRNAs in patients with non-small cell lung
cancer: Implications for outcomes and histologic classification.
Carcinogenesis. 34:1031–1038. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hariu H, Hirohashi Y, Torigoe T, Asanuma
H, Hariu M, Tamura Y, Aketa K, Nabeta C, Nakanishi K, Kamiguchi K,
et al: Aberrant expression and potency as a cancer immunotherapy
target of inhibitor of apoptosis protein family, Livin/ML-IAP in
lung cancer. Clin Cancer Res. 11:1000–1009. 2005.PubMed/NCBI
|
|
4
|
Li J, Chen P, Li XQ, Bao QL, Dai CH and Ge
LP: Elevated levels of survivin and livin mRNA in bronchial
aspirates as markers to support the diagnosis of lung cancer. Int J
Cancer. 132:1098–1104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tanabe H, Yagihashi A, Tsuji N, Shijubo Y,
Abe S and Watanabe N: Expression of survivin mRNA and livin mRNA in
non-small-cell lung cancer. Lung Cancer. 46:299–304. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhuang L, Shen LD, Li K, Yang RX, Zhang
QY, Chen Y, Gao CL, Dong C, Bi Q, Tao JN, et al: Inhibition of
livin expression suppresses cell proliferation and enhances
chemosensitivity to cisplatin in human lung adenocarcinoma cells.
Mol Med Rep. 12:547–552. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yuan D, Liu L, Xu H and Gu D: The effects
on cell growth and chemosensitivity by livin RNAi in non-small cell
lung cancer. Mol Cell Biochem. 320:133–140. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sun JG, Liao RX, Zhang SX, Duan YZ, Zhuo
WL, Wang XX, Wang ZX, Li DZ and Chen ZT: Role of inhibitor of
apoptosis protein Livin in radiation resistance in nonsmall cell
lung cancer. Cancer Biother Radiopharm. 26:585–592. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kasof GM and Gomes BC: Livin, a novel
inhibitor of apoptosis protein family member. J Biol Chem.
276:3238–3246. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Crnkovic-Mertens I, Muley T, Meister M,
Hartenstein B, Semzow J, Butz K and Hoppe-Seyler F: The
anti-apoptotic livin gene is an important determinant for the
apoptotic resistance of non-small cell lung cancer cells. Lung
Cancer. 54:135–142. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lin X, Li HR, Lin XF, Yu ME, Tu XW, Hua
ZD, Lin M, Xu NL, Han LL and Chen YS: Silencing of Livin inhibits
tumorigenesis and metastasis via VEGF and MMPs pathway in lung
cancer. Int J Oncol. 47:657–667. 2015.PubMed/NCBI
|
|
12
|
Lau NC, Lim LP, Weinstein EG and Bartel
DP: An abundant class of tiny RNAs with probable regulatory roles
in Caenorhabditis elegans. Science. 294:858–862. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lee RC and Ambros V: An extensive class of
small RNAs in Caenorhabditis elegans. Science. 294:862–864.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kim VN and Nam JW: Genomics of microRNA.
Trends in genetics. TIG. 22:165–173. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Farazi TA, Spitzer JI, Morozov P and
Tuschl T: miRNAs in human cancer. J Pathol. 223:102–115. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Puisségur MP, Mazure NM, Bertero T,
Pradelli L, Grosso S, Robbe-Sermesant K, Maurin T, Lebrigand K,
Cardinaud B, Hofman V, et al: miR-210 is overexpressed in late
stages of lung cancer and mediates mitochondrial alterations
associated with modulation of HIF-1 activity. Cell Death Differ.
18:465–478. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu S, Zhang G, Li P, Chen S, Zhang F, Li
J, Jiang C, Chen X, Wang Y, Du Y, et al: miR-198 targets SHMT1 to
inhibit cell proliferation and enhance cell apoptosis in lung
adenocarcinoma. Tumour Biol. 37:5193–5202. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pardo OE, Castellano L, Munro CE, Hu Y,
Mauri F, Krell J, Lara R, Pinho FG, Choudhury T, Frampton AE, et
al: miR-515-5p controls cancer cell migration through MARK4
regulation. EMBO Rep. 17:570–584. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen YS, Li HR, Lin M, Chen G, Xie BS, Xu
NL and Lin LF: Livin abrogates apoptosis of SPC-A1 cell by
regulating JNKI signaling pathway. Mol Biol Rep. 37:2241–2247.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dubrez-Daloz L, Dupoux A and Cartier J:
IAPs: More than just inhibitors of apoptosis proteins. Cell Cycle.
7:1036–1046. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sun J, Liao R, Chen Z, Wang Z, Zhang Q and
Hu Y: Study on enhancing sensitivity of SPC-A1 cells to
chemotherapy by Livin isoform-specific gene silencing. Zhongguo Fei
Ai Za Zhi. 10:461–465. 2007.(In Chinese). PubMed/NCBI
|
|
22
|
Yuan D, Liu L and Gu D: Transcriptional
regulation of livin by beta-catenin/TCF signaling in human lung
cancer cell lines. Mol Cell Biochem. 306:171–178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang J, Zhao H, Xin Y and Fan L:
MicroRNA-198 inhibits proliferation and induces apoptosis of lung
cancer cells via targeting FGFR1. J Cell Biochem. 115:987–995.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Gigante M, Pontrelli P, Herr W, Gigante M,
D'Avenia M, Zaza G, Cavalcanti E, Accetturo M, Lucarelli G,
Carrieri G, et al: miR-29b and miR-198 overexpression in
CD8+ T cells of renal cell carcinoma patients
down-modulates JAK3 and MCL-1 leading to immune dysfunction. J
Transl Med. 14:842016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang S, Zhao Y and Wang L: MicroRNA-198
inhibited tumorous behaviors of human osteosarcoma through directly
targeting ROCK1. Biochem Biophys Res Commun. 472:557–565. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang M, Wang J, Kong X, Chen H, Wang Y,
Qin M, Lin Y, Chen H, Xu J, Hong J, et al: MiR-198 represses tumor
growth and metastasis in colorectal cancer by targeting fucosyl
transferase 8. Sci Rep. 4:61452014. View Article : Google Scholar : PubMed/NCBI
|