Introduction
Ovarian cancer is the most lethal gynecological
malignancy and the fifth most common cause of cancer death among
women, 90% is epithelial ovarian cancer (EOC) (1). The early diagnosis of ovarian cancer
is not perfect so that the majority of patients are diagnosed with
advanced disease, and the five-year survival rate for EOC is
approximately 30% (2). Therefore it
is necessary to find specific diagnostic methods and therapeutic
approaches for ovarian cancer.
B7-H3 is encoded by the CD276 gene, including two
main isoforms, named 4Ig-B7-H3 and 2Ig-B7-H3, which were first
identified in 2001 (3,4). A newly found member of B7/CD28 family,
and was identified as an accessory co-stimulatory molecule after
initial antigen priming in cooperation with a putative counter
receptor (5). Recently, a number of
articles have reported that B7-H3 is highly expressed in many
cancers, including colorectal cancer (6), hepatocellular carcinoma (7), lung cancer (8), gastric cancer (9), breast cancer (10) and prostate cancer (11). In addition, B7-H3 has been shown to
be associated with poor prognosis by the Jak2/Stat3 pathway.
Although the expression of B7-H3 in ovarian cancer was mentioned in
a document, it was verified only at the tissue level that the high
expression of B7-H3 in ovarian cancer and B7-H3-positive tumor
vasculature is associated with high-grade serous histological
subtype, increased recurrence and reduced survival (12). There is no study on the biological
behavior and mechanism of the cells. Thus in this study, we show
how B7-H3 affects the biological behavior of the cells and its
mechanism.
Materials and methods
Patients and tissue samples
Ovarian cancer tissue samples were collected from 41
patients who underwent surgery from 2015 to 2016 in Qilu Hospital
of Shandong University. In addition, 31 normal ovarian tissues were
collected to use as control. None of the patients received
preoperative chemotherapy or radiotherapy before the surgery. All
patients were confirmed by pathological diagnosis. The present
study was approved by the Ethics Committee of Qilu Hospital.
Cell lines and cell culture
The human ovarian cancer cell lines A2780, SKOV3,
and HO8910 were purchased from the American Type Culture Collection
(ATCC; Manassas, VA, USA). A2780 and HO8910 were cultured in the
medium of Roswell Park Memorial Institute (RPMI)-1640 (Hyclone
Laboratories, Logan, UT, USA), while SKOV3 in the medium of
Micro-5A (Gibco-Invitrogen, Grand Island, NY, USA). All of them
were supplemented with 10% fetal bovine serum (FBS; Gibco, Sydney,
Australia) and incubated at 37°C in a humidified incubator
supplemented with 5% CO2.
Antibodies and reagents
Rabbit anti-human Jak2, Phospho-Jak2 (Tyr1007),
signal transducer and activator of transcription 3 (Stat3),
phospho-Stat3 (Tyr705), Bcl-2 and Bax antibodies were purchased
from Abcam (Cambridge, MA, USA). Mouse anti-human Bcl-xl, caspase-8
and MMP-2 were obtained from Santa Cruz Biotechnology (Dallas, TX,
USA). Rabbit anti-human B7-H3, cleaved caspase-8 and the GAPDH were
purchased from Cell Signaling Technology (CST) (Danvers, MA, USA).
All of the above are monoclonal antibodies. Tryphostins AG490 was
from Abcam and prepared at a concentration of 100 mmol/l stock
solution in dimethyl sulfoxide (DMSO).
Immunohistochemical analysis
Polink-2 Plus Polymer HRP Detection System
(ZSGB-BIO, Beijing, China) was used. Paraffin tissues were cut into
4-µm sections. Section were de-paraffinized in xylene, re-hydrated
through a graded ethanol series, then repaired by citric acid
(microwave boiling method). H2O2 (3%) was
incubated for 10 min to block endogenous peroxidase activity. PBS
(phosphate-buffered saline) was used for washing three times for 3
min each. After blocking endogenous peroxidase activity, the slides
were incubated with primary antibody overnight at 4°C. The next
day, sections were washed in PBS three times for 5 min each.
Secondary antibody was applied for 30 min at 37°C. DAB was used for
coloring. Finally slides were counterstained with water rinsing,
staining, dehydration, transparent and mounting. Quantification was
recorded as follows: <10% positive cells, 0; 10–25%, 1; 26–50%,
2; >50% positive cells, 3. Sections with a final score of 0–1
was classified as negative, and ≥2 was considered positive.
Immunofluorescence staining
To find the B7-H3 expression location, the A2780 and
SKOV3 cells were seeded in a 24-well plate at 37°C overnight. Then,
after three washes with PBS, the cells were fixed with 4%
paraformaldehyde for 15 min at room temperature, and permeabilized
with 0.2% Triton 150–200 µl in PBS for 10 min. After that, the
B7-H3 (1:200 dilution) antibody was incubated at 4°C for overnight
and secondary antibody (1:200) for 1 h. Finally, cells were stained
with 4,6-diamidino-2-phenylindole (DAPI) for 5 min in the dark at
room temperature. The fluorescence images were observed using a
fluorescence microscope (Olympus, Tokyo, Japan).
Silencing and overexpression of
B7-H3
Silencing or overexpression of B7-H3 sequences were
designed by GenePharma Co, Ltd. (Shanghai, China). The cells were
seeded in 6-well plates, 24 h after the cell attachment, the virus
liquid was mixed in the culture medium. The cells were screened
with puromycin dihydrochloride (2 µg/ml; Amresco, Solon, OH, USA)
72 h later. After screening for 5–7 days, the cell line with stable
overexpression of B7-H3 (HO8910-B7-H3-EGFP) and their control cell
lines (HO8910-NC) were obtained. The A2780 and SKOV3 cells were
seeded in a 6-well plate with 3×104/ml cell per well, 24
h later, cells were transfected with 50 nM of the Sh-B7-H3 or
control sequences using Lipofectamine-2000 (Invitrogen Life
Technologies). The transfected cells were harvested 48 h
post-transfection for the follow-up experiments, with A2780-NC,
A2780-sh-B7-H3, SKOV3-NC and SKOV3-sh-B7-H3.
Western blot assay
The cells were washed 3 times with PBS and then
lysed on ice for approximately 30 min. The pyrolysis solution is
composed of radio immunoprecipitation assay buffer (RIPA),
phenylmethylsulfonyl fluoride (PMSF) and NaF. The cells were then
lysed with ultrasound. Cells were centrifuged at 12000 rpm at 4°C,
then supernatant were drained, loading buffer was added and heated
for 5 min in metal bath. The protein concentrations were measured
by using the BCA Protein Assay kit (Beyotime, Jiangsu, China).
Total protein (30–50 µg) was separated by SDS-polyacrylamide gel
electrophoresis (SDS-PAGE) in 10% gels (Invitrogen) and transferred
to PVDF membranes (ImmobilonP; Millipore, Bedford, MA, USA). After
blocking with 5% skim milk for 2 h, the membranes were cut into
strips and incubated with the indicated primary antibodies
overnight at 4°C. The next day, the membranes were washed 3 times
with TBS-T (20 mM Tris, pH 7.4, 137 mM NaCl, 0.05% Tween-20) and
then indicated with secondary antibodies at room temperature for
1–2 h. Ultimately, the immunoreactive protein bands were detected
by enhanced chemiluminescence (ECL) using ImageQuant LAS 4000 (GE
Healthcare Life Sciences, Logan, UT, USA). The results were
analyzed by ImageJ software (NIH, Bethesda, MD, USA).
Cell proliferation assay
The cell proliferation was evaluated by MTT assay.
In brief, cells were seeded in 96-well plates at 2000 cells per
well. At indicated time-points, the cells in the 96-well plate were
incubated with 20 µl MTT at 37°C for 4 h. The cell growth was
monitored every 24 h for up to 5 days. Absorbance was measured for
each well at a wavelength of 490 nm using a microplate reader
(Infinite 2000; Tecan, Männedorf, Switzerland).
Cell migration and invasion
assays
For the in vitro migration and invasion
assays, 3–5×104 cells of the A2780, HO8910 or SKOV3
derived cell cultures, in serum-free RPMI and Micro-5A medium, were
seeded into the top of 8 µm pore size transwell chambers (Costar,
Cambridge, MA, USA), which contained 100 µl of Matrigel (1:8
dilution in serum-free medium; Corning Inc., Corning, NY, USA) or
not. Then the chambers were put into the 24-well plates which
included culture medium with 10% serum. After incubation at 37°C
for 24 h in a 5% CO2 atmosphere, the cells were stained
with 0.1% crystal violet and then washed 3 times with PBS. The
images were captured by the Olympus IX51 inverted microscope. The
number of migrating and invading cells on the chambers were counted
in 5 random fields per chamber at ×200 magnification of each
group.
Cell cycle and apoptosis assays
To analyze the effect of B7-H3 on the different
phases of the cell cycle and apoptosis, flow cytometry was used.
Cells were harvested from each sample then fixed with cold 75%
ethanol at 4°C overnight. The cells were washed 3 times and stained
for 30 min in propidium iodide (PI) staining solution in the dark.
The cell cycle was detected by FACSCalibur flow cytometer (both
from BD Biosciences, Franklin Lakes, NJ, USA) and analyzed by
ModFit LT software. For apoptosis, cells were harvested and
processed as described in the Annexin V-fluorescein isothiocyanate
(FITC) Apoptosis Detection kit I manual (BD Transduction
Laboratories, BD Biosciences) and analyzed by flow cytometry (BD
LSR II).
Statistical analysis
The data were analyzed using GraphPad Prism version
5.01 (GraphPad Software Inc., San Diego, CA, USA). The experiment
was performed a minimum of three times, and all data are shown as
means with standard deviations (SDs). The data were analyzed for
statistical significance using an unpaired Student's t-test or a
Chi-squared test. P-value at <0.05 difference was considered to
be statistically significant.
Results
B7-H3 expression in clinical specimens
and the location of B7-H3 in ovarian cancer cell lines
In order to find out the expression of B7-H3 in
ovarian cancer and the location of B7-H3, we used
immunohistochemistry and immunofluorescence methods.
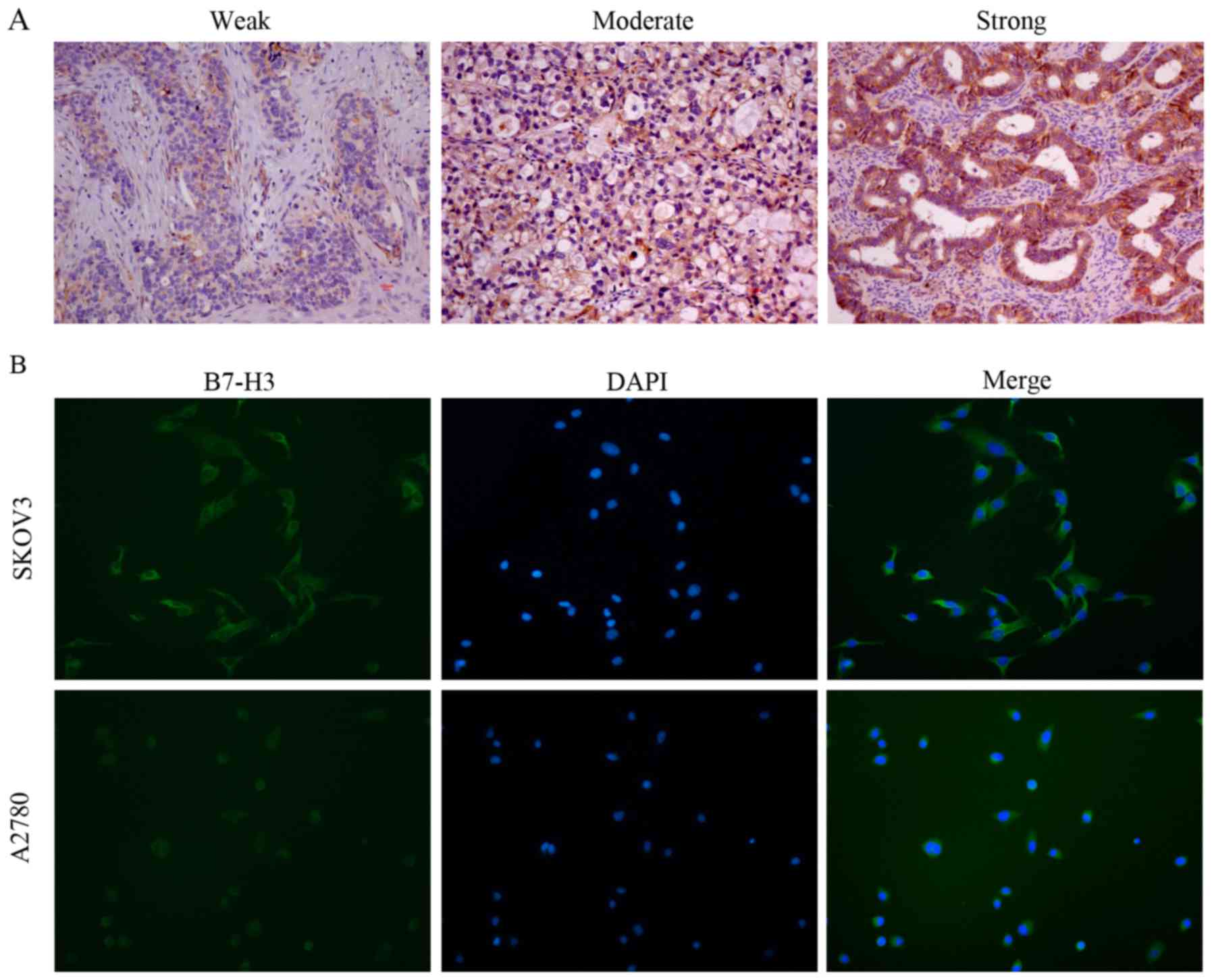
Immunohistochemistry was performed to detect the expression of
B7-H3 in 41 cases of ovarian cancer tissues and 31 cases of normal
ovarian tissues. Although no expression or low expression of B7-H3
can be found in ovarian cancer and there are some weak expression
in normal tissues (Fig. 1A), the
results showed that the expression of B7-H3 in ovarian cancer was
significantly higher than that in normal tissues (P<0.001)
(Table I). Immunohistochemistry
revealed that B7-H3 was expressed in the cytoplasm of the tissue,
and a few were expressed in the interstitial tissue. In order to
clarify the localization of B7-H3, immunofluorescence was utilized
and the results showed that B7-H3 was mostly expressed in the
cytoplasm of ovarian cancer cells (Fig.
1B).
 | Table I.Relationship between B7-H3 expression
on tumor cells and clinicopathological factors. |
Table I.
Relationship between B7-H3 expression
on tumor cells and clinicopathological factors.
|
|
| Expression of
B7-H3 |
|
|---|
|
|
|
|
|
|---|
| Factors | No. | Negative | Positive | P-value |
|---|
| B7-H3 |
|
|
|
<0.0001 |
| Ovarian
cancer tissue | 41 | 11 | 30 |
|
| Normal
ovarian tissue | 31 | 24 | 7 |
|
| Age (years) |
|
|
| 0.195 |
| ≤50 | 16 | 2 | 14 |
|
|
>50 | 25 | 9 | 16 |
|
| Size (cm) |
|
|
| 0.903 |
| ≤5 | 18 | 5 | 13 |
|
|
>5 | 23 | 6 | 17 |
|
| Histology |
|
|
| 0.920 |
| Serous
CA | 28 | 7 | 21 |
|
|
Endometrioid CA | 7 | 1 | 6 |
|
| Vascular
Invasion |
|
|
| 0.300 |
| − | 36 | 11 | 25 |
|
| + | 5 | 0 | 5 |
|
| Distant
metastasis |
|
|
| 0.041 |
| − | 14 | 7 | 7 |
|
| + | 27 | 4 | 23 |
|
|
Differentiation |
|
|
| 1.000 |
|
Low | 2 | 0 | 2 |
|
|
Moderate/high | 33 | 8 | 25 |
|
| Clinical stage |
|
|
| 0.057 |
|
I/II | 13 | 6 | 7 |
|
|
III/IV | 28 | 5 | 23 |
|
B7-H3 expression in relation to
patient clinicopathological factors
The relationship between clinicopathological factors
and B7-H3 expression in patients with ovarian cancer is shown in
Table I. Our data suggest that
B7-H3 expression is associated with distant metastasis of ovarian
cancer (P=0.041), whereas it is not correlated with patient age
(P=0.195), tumor size (P=0.903), tumor histology type (P=0.920),
differentiation degree (P=1.000), clinical stage (P=0.057), and
vascular invasion (P=0.300). However, in this study the number of
cases is too small, therefore, in order to further clarify the
relationship between B7-H3 and the clinicopathological
characteristics of patients the number of samples should be
increased.
Silencing B7-H3 weakens cell
proliferation
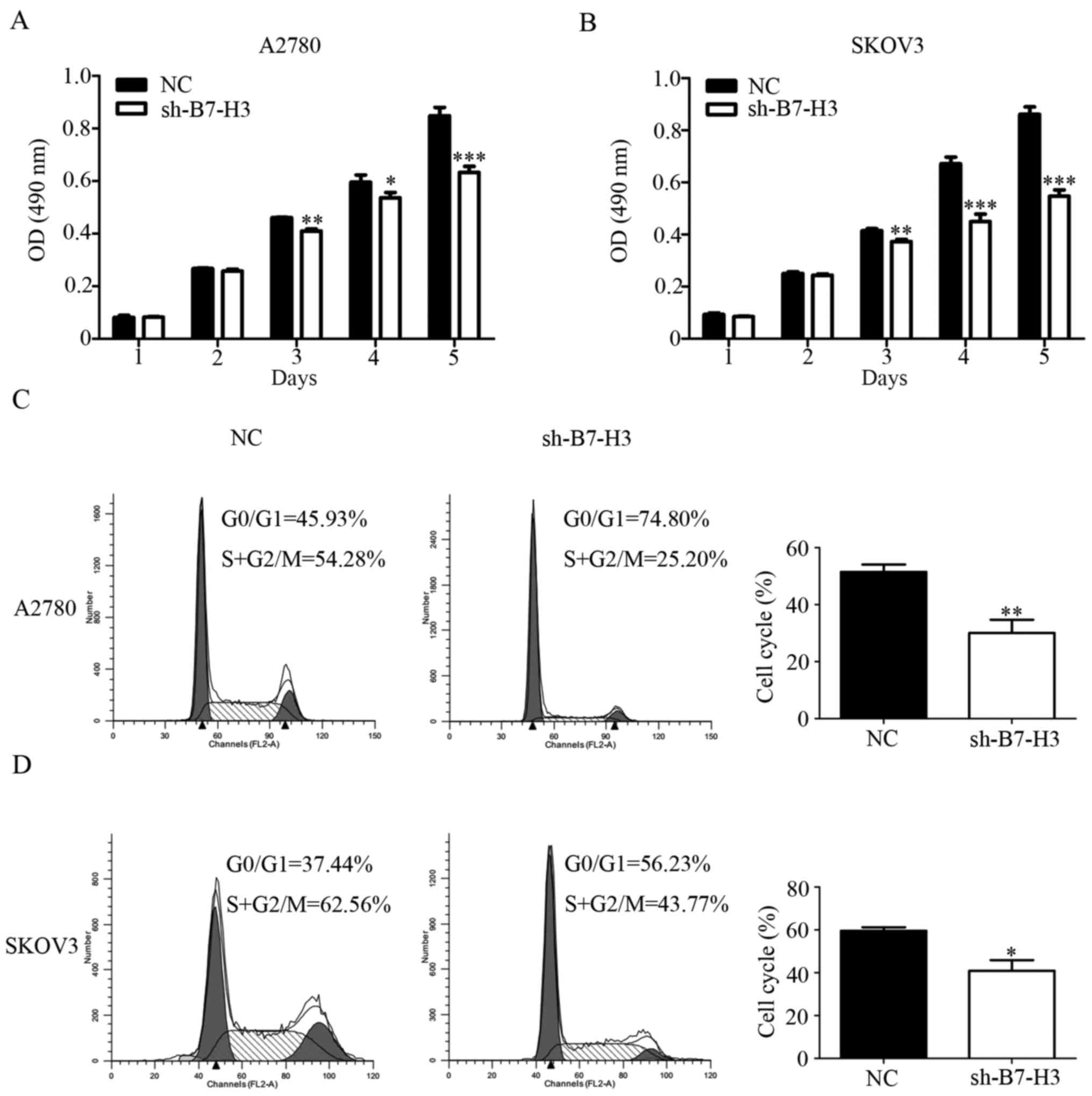
To characterize the role of B7-H3 in A2780 and SKOV3
cell growth we measured the cell proliferation rate in vitro
by MTT assay (11). The experiment
was divided into two groups: NC group and sh-B7-H3 group. The
results suggest that the cell viability of sh-B7-H3 group was
weakened at 3, 4 and 5 days compared with the NC group (Fig. 2A and B). Similar results were also
found in the cell cycle (Fig. 3C and
D). Compared with the control group, A2780 and SKOV3 cell lines
that interfered with B7-H3 decreased the S and G2/M phases of the
cell cycle, which was a period of cell proliferation ability. Thus,
B7-H3 molecule expressed in ovarian cancer cells might play an
important role in regulating the colony formation ability.
Silencing B7-H3 increases cell
apoptosis
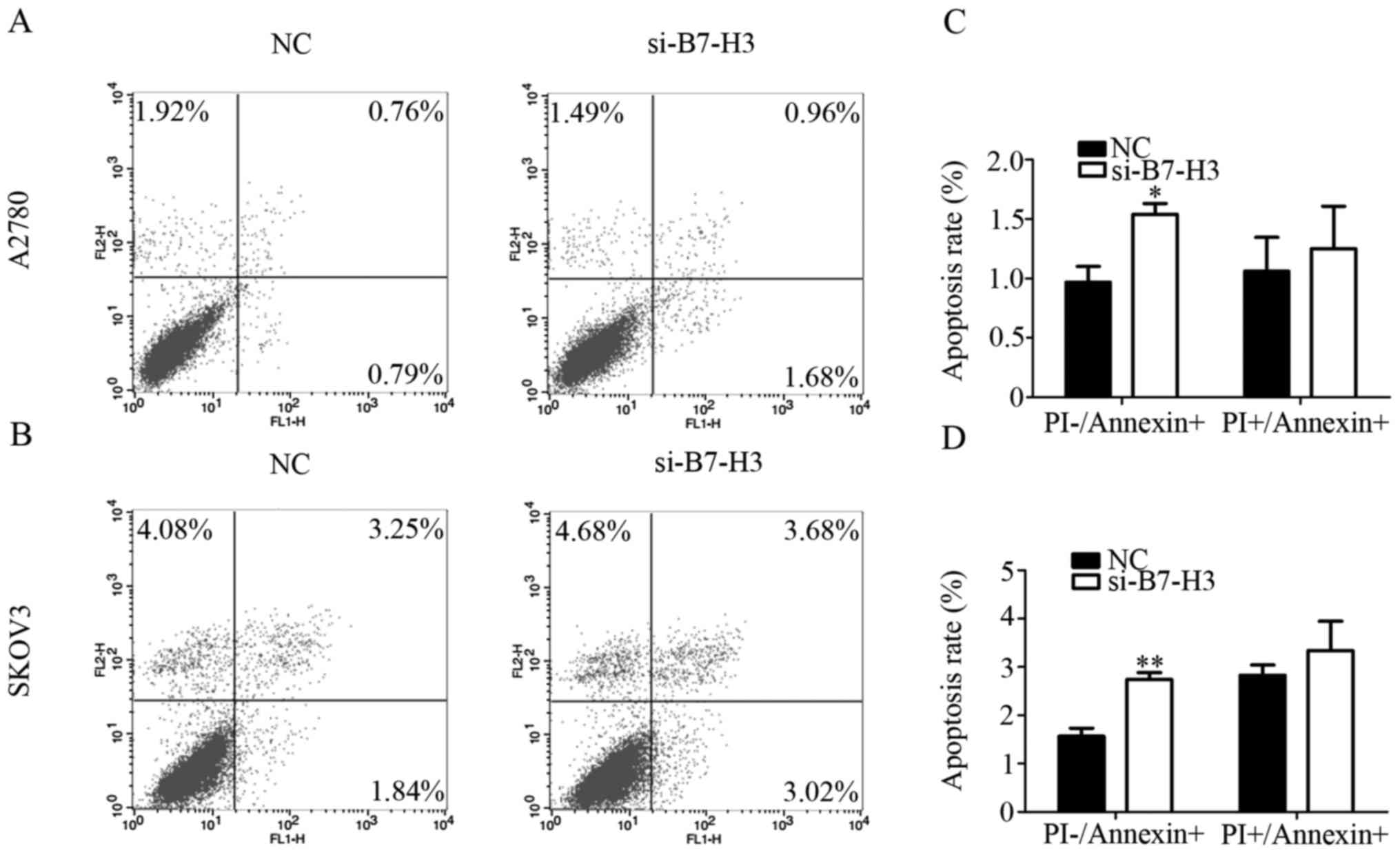
To verify the effect of B7-H3 on cell apoptosis
after B7-H3 interference, we performed flow cytometry. The extent
of apoptosis was investigated by measuring the amount of Annexin V
stained cells, a marker for early stage apoptosis. The amount of
all reagent-positive cells, which reflect the late stage apoptosis,
were also measured. We found that when B7-H3 affected the early
stage apoptosis of A2780 and SKOV3 cells were increased, and the
late stage apoptosis although increased, was P>0.05 (Fig. 3A and B). Therefore, B7-H3 mainly
promotes apoptosis of cells through the early stage apoptosis. In
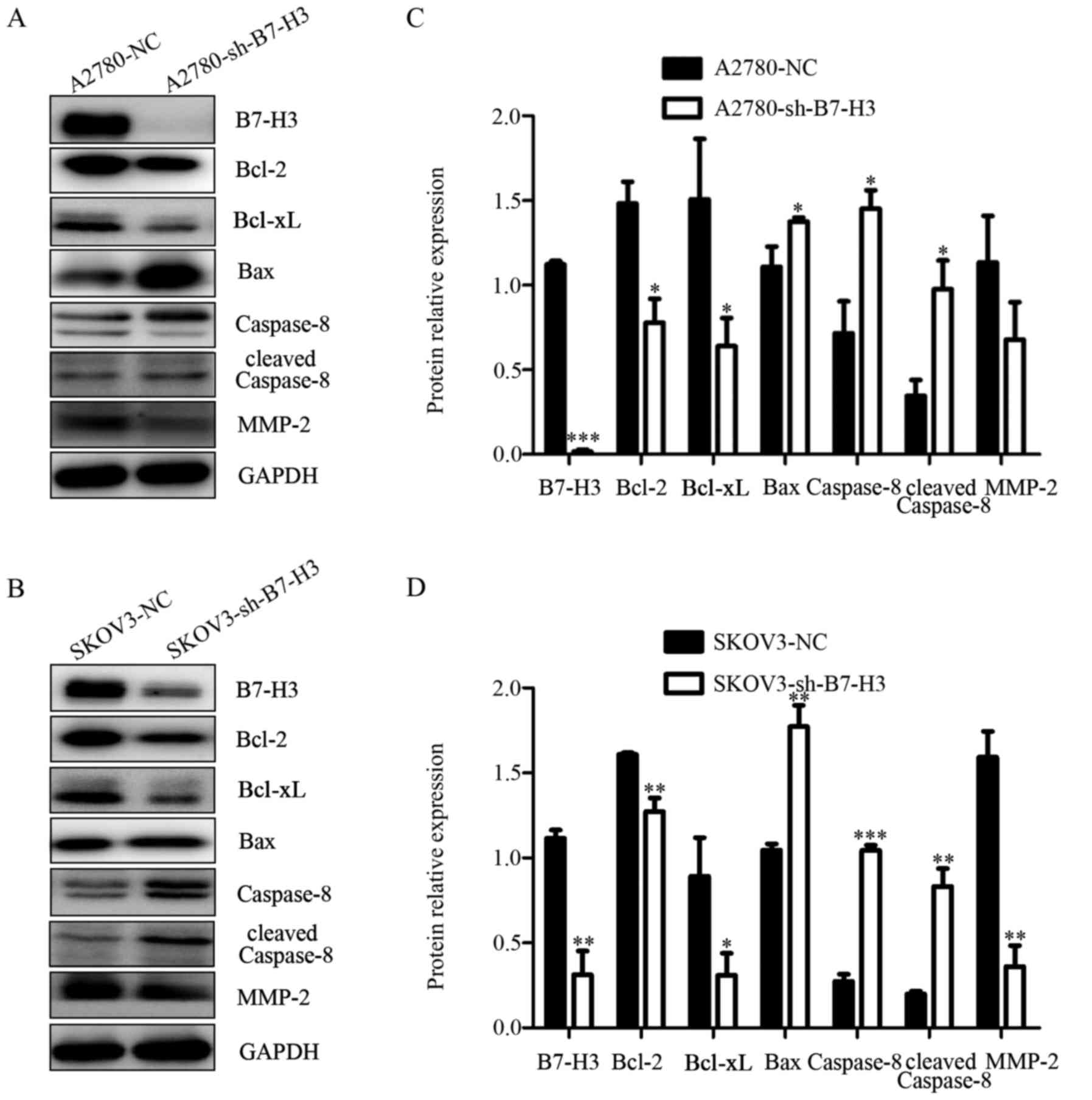
addition, we observed the changes of apoptosis related proteins by
the western blot method with the B7-H3 knockout. The results showed
that with the silence of B7-H3 the expression of apoptosis
regulator proteins of the Bcl-2 family, including the
anti-apoptotic proteins Bcl-2 and Bcl-xl decreased, while
expression of the pro-apoptotic protein Bax, caspase-8 and cleaved
caspase-8 increased (Fig. 4A and
B). The expression of B7-H3 in ovarian cancer cells is closely
related to apoptosis related molecules, which suggests that
silencing B7-H3 may promote apoptosis of ovarian cancer cells.
Silencing B7-H3 reduced the migration
and invasion potential of ovarian cancer cells
We next further assessed the influence of B7-H3 on
ovarian cancer cell migration and invasion by transwell assays. As
shown in the results, compared with the control groups, the number
of cells in migration passing through the chamber was significantly
reduced in A2780-sh-B7-H3 group and SKOV3-sh-B7-H3 group (Fig. 5A and B). The same phenomenon was
observed in the cell invasion test. Silencing B7-H3 can impede cell
migration and inhibit cell invasion via downregulating the
expression of MMP-2 (13). Thus, we
measured the invasion related proteins by western blotting and
found that MMP-2 were lower in A2780-sh-B7-H3 and SKOV3-sh-B7-H3
cells than in the NC groups (Fig. 4A
and B). Based on these results, we come to the conclusion that
the downregulation of B7-H3 expression could suppress cellular
migration and invasion in human ovarian cancer cells.
Overexpression of B7-H3 enhances
ovarian cancer cell invasion, migration and proliferation in
vitro
We described above the effect of silencing B7-H3 in
ovarian cancer cells. In order to further verify the role of B7-H3
in ovarian cancer cells, HO8910 cell line was overexpressed by
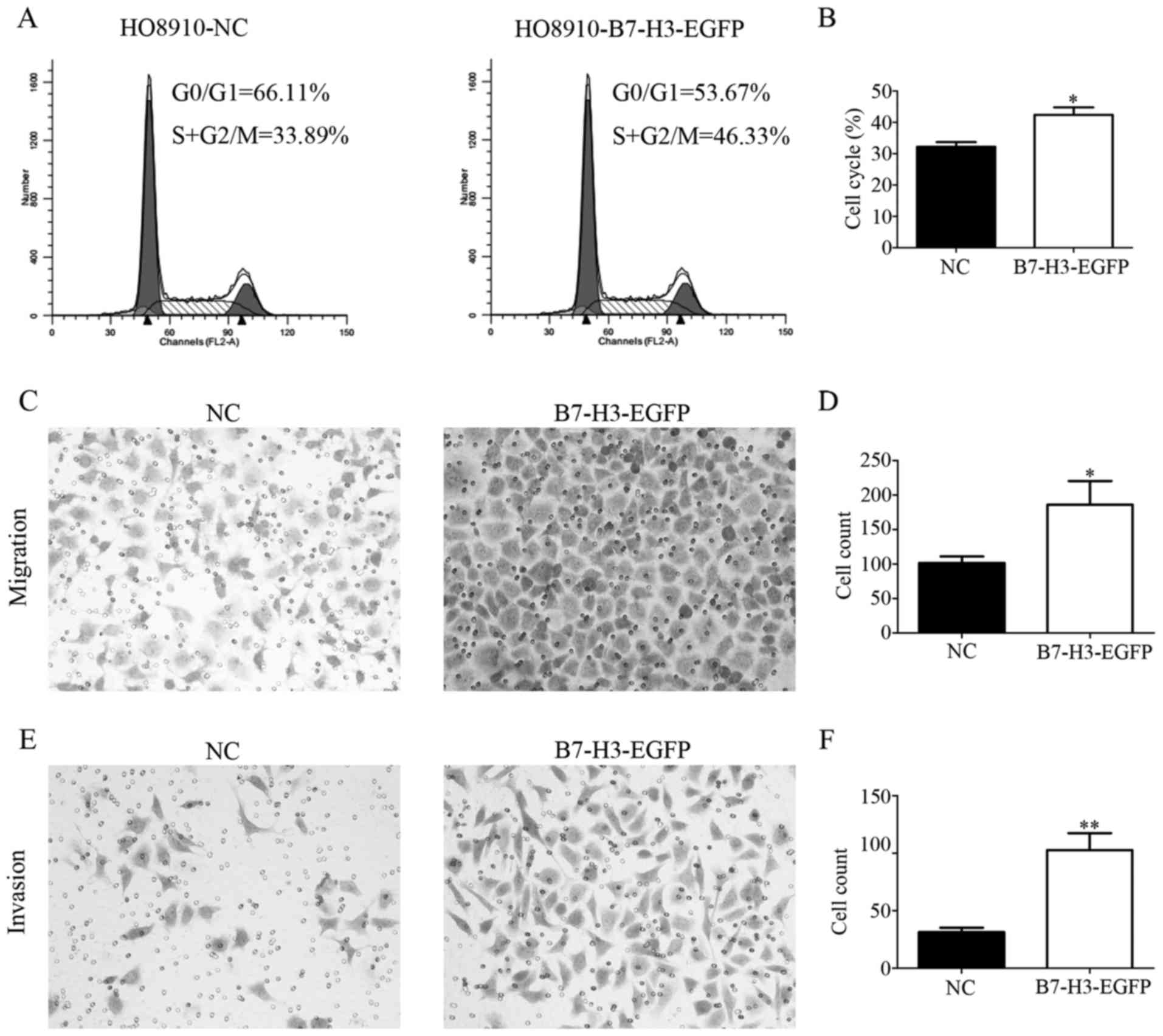
lentivirus. Using cell cycle assay, we observed that the growth of
HO8910-B7-H3-EGFP was increased compared to NC cells (Fig. 6A and B). Besides, we performed
transwell assay and the result showed that overexpression of B7-H3
significantly increased the ability of cell migration and invasion
(Fig. 6C and E). The expression of
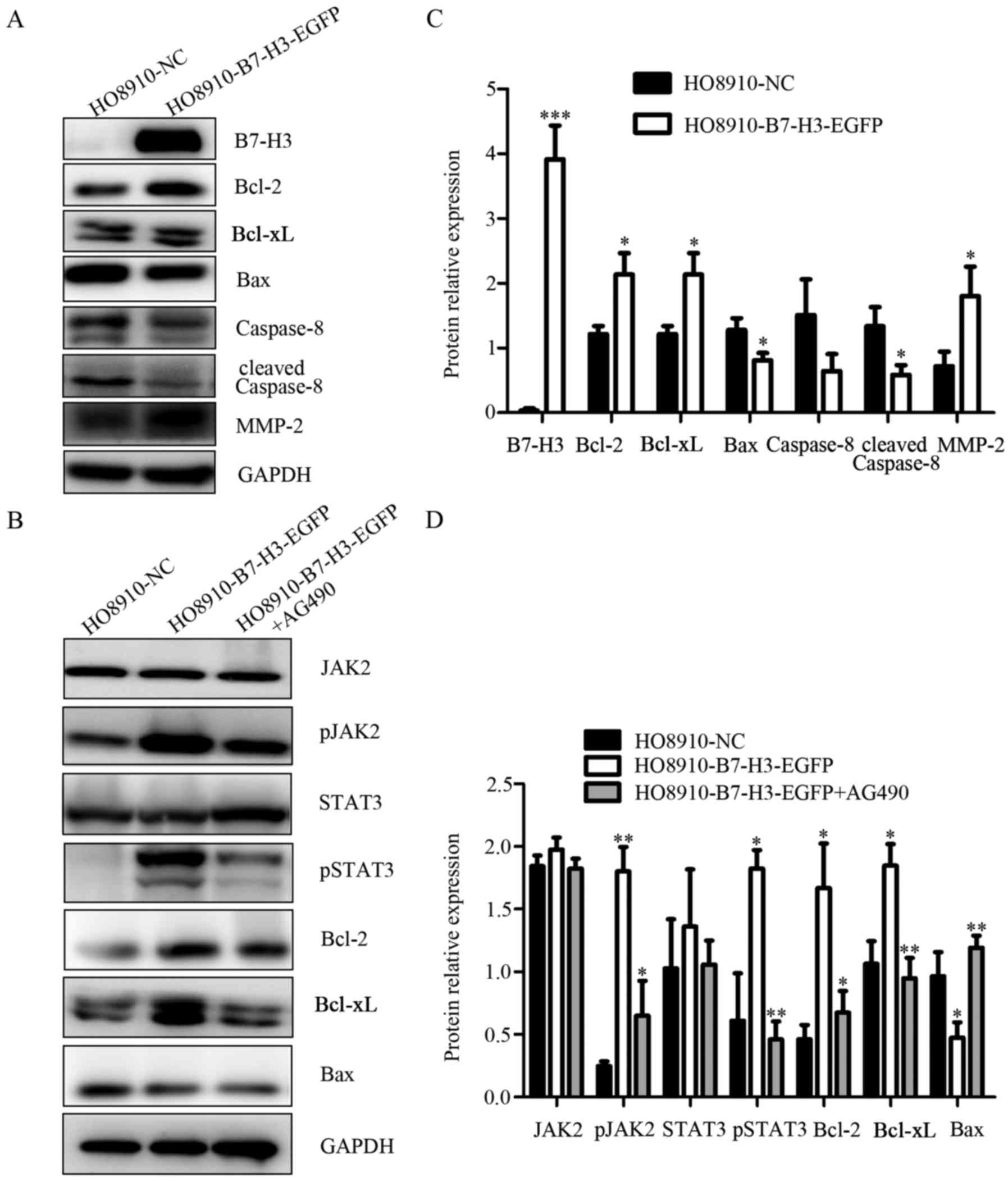
related protein MMP-2 was elevated with the overexpression of B7-H3
(Fig. 7A). The expression of B7-H3
and the anti-apoptotic proteins were positively correlated in CRC
cell lines (6). Therefore, we found
that the expression of protein Bcl-2, Bcl-xl added while Bax,
caspase-8, cleaved caspase-8 reduced by upregulation of B7-H3
expression (Fig. 7A). To summarize,
overexpression of B7-H3 inhibits apoptosis and enhanced cell
invasion, migration and proliferation in ovarian cancer cells.
Overexpression of B7-H3 affects the
Jak2-Stat3 pathway
The role of B7-H3 in ovarian cancer cells is
described above, therefore, we investigated which signaling pathway
was involved in this process. The Jak2/Stat3 pathway has been
reported to be the key in cell migration, invasion and metastasis,
and inhibition of Jak2/Stat3 signaling induced CRC cell apoptosis,
cell arrest and reduced tumor cell invasion (14–16).
Thus, it was analyzed whether this pathway could be affected by
B7-H3 in ovarian cancer. HO8910-B7-H3-EGFP cells were treated with
AG490 with a concentration of 100 mmol/l. After 48 h, the protein
expression of related molecules in Jak2/Stat3 pathway was detected
by western blotting. As shown in the chart, the phosphorylation
levels of Jak2 and Stat3 increased with the overexpression of
B7-H3. However, when the cells were treated with AG490, the
expression decreased correspondingly. Furthermore, anti-apoptotic
proteins Bcl-2 and Bcl-xl increased while expression of the
pro-apoptotic protein Bax was reduced following upregulated
expression of B7-H3 and after joining the AG490 the results were
exactly the opposite (Fig. 7B).
These findings indicated that B7-H3 may participate and influence
the Jak2/Stat3 pathway in ovarian cancer cells. However, whether
B7-H3 affects this pathway through activation or other ways, or
not, needs further experimentation.
Discussion
B7-H3, as an immunoregulatory molecule, playing
different roles in different types of human cancers (17). The role of B7-H3 in tumor immunity
is complicated as both T cell co-stimulatory and co-inhibitory
effects have been shown (18). In
this study, we provide evidence that B7-H3 may be associated with
tumor progression in ovarian cancer. In other words, B7-H3 probably
plays a negative regulatory role in ovarian cancer.
In the past studies, only few scattered stromal
cells in non-neoplastic ovarian tissues expressed B7-H3 (3,19,20).
Only one recent investigation showed that most ovarian cancers
express B7-H3 (12). In this study,
we further demonstrated that B7-H3 is highly expressed in patients
with ovarian cancer, mainly in adenocarcinoma, but has no
expression or low expression in normal epithelial ovarian tissues.
The correlation between B7-H3 and the age, clinical stage,
prognosis and other factors of patients with ovarian cancer was
analyzed. The data suggest that B7-H3 expression is only associated
with distant metastasis of ovarian cancer. We also found that B7-H3
was mainly expressed in the cytoplasm of ovarian cancer cells. To
date, there are no published studies looking at the ability of
B7-H3 to affect the growth of human ovarian cancer cell lines in
vitro (21). Therefore, this
study provides the basis how B7-H3 affects the biological behavior
of ovarian cancer cell lines. Our study demonstrated that B7-H3 is
closely associated with tumor progression in ovarian cancer cells.
Silencing B7-H3 can attenuate the A270, SKOV3 cell lines of
proliferation, migration and invasion and increase cell apoptosis,
and induce changes in related molecules. On the contrary,
overexpression of B7-H3 can increase the proliferation, migration
and invasion of HO8910 cell line, and decrease the rate of
apoptosis. The changes of protein molecules suggest that B7-H3 may
be a potential therapeutic target for ovarian cancer.
The Jak2-Stat3 pathway plays a significant role in
biological function on various human cancers. Zhang et al
confirmed that the Jak2-Stat3 signaling pathway played an important
role in regulating the anti-apoptotic ability of B7-H3 (6). Silenced B7-H3 expression suppresses
migration and invasion of HCC cells via Jak2/SAtat3/Slug signaling
pathway, which was proved by Kang et al (5). Stat3 regulates cellular
differentiation, proliferation, migration and survival as a
cytoplasmic transcription factor (22–25),
which was regulated by its upstream molecule Jak2. Jak2-Stat3 are
reported to be the downstream signaling pathways that adjust the
ability of B7-H3 to regulate the expression of Bcl-2, Bcl-xl, Bax
and other molecules (6). In the
present study, we found that after overexpression of B7-H3 the
phosphorylation levels of Jak2 and Stat3 increased. However, after
treatment with AG490, which is the inhibitor of Jak2, the
expression of anti-apoptotic proteins Bcl-2 and Bcl-xl and
pro-apoptotic protein Bax also changed. The data indicate that
B7-H3 enhanced proliferation and reduces apoptosis probably by
influencing Jak2-Stat3 pathway. But how B7-H3 specifically
influenced this pathway, needs further experiments to verify the
mechanism.
In conclusion, results of this study identified that
B7-H3 is related to tumor progression in ovarian cancer and
probably can be used as an indicator in clinic in the future. The
limitations are that we did not conduct animal experiments in this
research due to the lack of experimental funds and the limitations
of laboratory conditions. We consider this aspect well worth
studying. As mentioned above, the specific role of B7-H3 and its
mechanism need further studies.
Acknowledgements
The present study was funded by the National Natural
Science Foundation of China (NSFC; 81572559), the Science and
Technology Developing Planning of Shandong Province (2014GH218029)
and the National Science and Technology Project of China
(2015BAI13B05).
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2012. CA Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Chapoval AI, Ni J, Lau JS, Wilcox RA,
Flies DB, Liu D, Dong H, Sica GL, Zhu G, Tamada K, et al: B7-H3: A
costimulatory molecule for T cell activation and IFN-gamma
production. Nat Immunol. 2:269–274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Dong C, Nurieva RI and Prasad DV: Immune
regulation by novel costimulatory molecules. Immunol Res. 28:39–48.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kang FB, Wang L, Jia HC, Li D, Li HJ,
Zhang YG and Sun DX: B7-H3 promotes aggression and invasion of
hepatocellular carcinoma by targeting epithelial-to-mesenchymal
transition via JAK2/STAT3/Slug signaling pathway. Cancer Cell Int.
15:452015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang T, Jiang B, Zou S-T, Liu F and Hua
D: Overexpression of B7-H3 augments anti-apoptosis of colorectal
cancer cells by Jak2-STAT3. World J Gastroenterol. 21:1804–1813.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kang FB, Wang L, Li D, Zhang YG and Sun
DX: Hepatocellular carcinomas promote tumor-associated macrophage
M2-polarization via increased B7-H3 expression. Oncol Rep.
33:274–282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jin Y, Zhang P, Li J, Zhao J, Liu C, Yang
F, Yang D, Gao A, Lin W, Ma X, et al: B7-H3 in combination with
regulatory T cell is associated with tumor progression in primary
human non-small cell lung cancer. Int J Clin Exp Pathol.
8:13987–13995. 2015.PubMed/NCBI
|
|
9
|
Dai W, Shen G, Qiu J, Zhao X and Gao Q:
Aberrant expression of B7-H3 in gastric adenocarcinoma promotes
cancer cell metastasis. Oncol Rep. 32:2086–2092. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sun J, Guo YD, Li XN, Zhang YQ, Gu L, Wu
PP, Bai GH and Xiao Y: B7-H3 expression in breast cancer and
upregulation of VEGF through gene silence. Onco Targets Ther.
7:1979–1986. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yuan H, Wei X, Zhang G, Li C, Zhang X and
Hou J: B7-H3 over expression in prostate cancer promotes tumor cell
progression. J Urol. 186:1093–1099. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zang X, Sullivan PS, Soslow RA, Waitz R,
Reuter VE, Wilton A, Thaler HT, Arul M, Slovin SF, Wei J, et al:
Tumor associated endothelial expression of B7-H3 predicts survival
in ovarian carcinomas. Mod Pathol. 23:1104–1112. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang W, Wang Y, Wang J, Dong F, Zhu M,
Wan W, Li H, Wu F, Yan X and Ke X: B7-H3 silencing inhibits tumor
progression of mantle cell lymphoma and enhances chemosensitivity.
Int J Oncol. 46:2562–2572. 2015.PubMed/NCBI
|
|
14
|
Du W, Hong J, Wang YC, Zhang YJ, Wang P,
Su WY, Lin YW, Lu R, Zou WP, Xiong H, et al: Inhibition of
JAK2/STAT3 signalling induces colorectal cancer cell apoptosis via
mitochondrial pathway. J Cell Mol Med. 16:1878–1888. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xiong H, Chen ZF, Liang QC, Du W, Chen HM,
Su WY, Chen GQ, Han ZG and Fang JY: Inhibition of DNA
methyltransferase induces G2 cell cycle arrest and apoptosis in
human colorectal cancer cells via inhibition of JAK2/STAT3/STAT5
signalling. J Cell Mol Med. 13:3668–3679. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiong H, Zhang ZG, Tian XQ, Sun DF, Liang
QC, Zhang YJ, Lu R, Chen YX and Fang JY: Inhibition of JAK1,
2/STAT3 signaling induces apoptosis, cell cycle arrest, and reduces
tumor cell invasion in colorectal cancer cells. Neoplasia.
10:287–297. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang Z-S, Zhong M, Bian Y-H, Mu Y-F, Qin
S-L, Yu M-H and Qin J: MicroRNA-187 inhibits tumor growth and
invasion by directly targeting CD276 in colorectal cancer.
Oncotarget. 7:44266–44276. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
zur Hausen H: Papillomaviruses and cancer:
From basic studies to clinical application. Nat Rev Cancer.
2:342–350. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sica GL, Choi IH, Zhu G, Tamada K, Wang
SD, Tamura H, Chapoval AI, Flies DB, Bajorath J and Chen L: B7-H4,
a molecule of the B7 family, negatively regulates T cell immunity.
Immunity. 18:849–861. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tringler B, Zhuo S, Pilkington G, Torkko
KC, Singh M, Lucia MS, Heinz DE, Papkoff J and Shroyer KR: B7-h4 is
highly expressed in ductal and lobular breast cancer. Clin Cancer
Res. 11:1842–1848. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fauci JM, Straughn JM Jr, Ferrone S and
Buchsbaum DJ: A review of B7-H3 and B7-H4 immune molecules and
their role in ovarian cancer. Gynecol Oncol. 127:420–425. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Liu H, Tekle C, Chen Y-W, Kristian A, Zhao
Y, Zhou M, Liu Z, Ding Y, Wang B, Mælandsmo GM, et al: B7-H3
silencing increases paclitaxel sensitivity by abrogating Jak2/Stat3
phosphorylation. Mol Cancer Ther. 10:960–971. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Levy DE and Darnell JE Jr: Stats:
Transcriptional control and biological impact. Nat Rev Mol Cell
Biol. 3:651–662. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Levy DE and Inghirami G: STAT3: A
multifaceted oncogene. Proc Natl Acad Sci USA. 103:10151–10152.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Schlessinger K and Levy DE: Malignant
transformation but not normal cell growth depends on signal
transducer and activator of transcription 3. Cancer Res.
65:5828–5834. 2005. View Article : Google Scholar : PubMed/NCBI
|