Introduction
Vascular endothelial growth factor (VEGF), also
known as vascular permeability factor, is an endothelial
cell-specific mitogen and an angiogenic inducer (1). The activation of VEGF induces
mitogenic permeability of the vascular endothelium; thus, VEGF has
an important role in various physiological and pathological
modulations, including cyclical blood vessel proliferation,
longitudinal bone growth, endochondral bone formation, wound
healing, cardiovascular diseases, ocular disorders, rheumatoid
arthritis and psoriasis (2–4).
Specifically, VEGF is a critical factor in tumor angiogenesis; VEGF
is highly expressed in various tumorous tissues, and the activation
of VEGF often promotes tumor growth (5). The VEGF family of proteins in mammals
is encoded by five different genes: VEGFA, VEGFB, VEGFC, FIGF, and
placenta growth factors (PGF-1 and PGF-2) (6). VEGFA has the most profound effects on
stimulating endothelial cell proliferation, survival, and
differentiation. Alternative splicing of the VEGFA gene yields five
different isoforms: VEGFA-120, VEGFA-144, VEGFA-164, VEGFA-188, and
VEGFA-205; each differing in biological activity, however they are
primarily associated with angiogenesis (7–10).
The VEGFA isoform regulates cell proliferation through two
membrane-bound receptor tyrosine kinases, VEGFR1 and VEGFR2, which
bind VEFGA with high affinity.
VEGF binds two related receptor tyrosine kinases
(RTKs), VEGF receptor-1 (VEGFR-1) and VEGFR-2. VEGFR-1 and VEGFR-2
have similar extracellular and transmembrane domains containing a
tyrosine kinase sequence (11,12).
A spliced, soluble form of VEGFR-1 (soluble Flt-1), once activated,
leads to the inhibition of downstream pathways of VEGF. Since the
binding site for VEGF (and placental growth factor) is primarily
located in the immunoglobulin-like domain (13), it has been proposed that VEGFR-1
may not be a primary receptor, but a decoy receptor to prevent VEGF
from binding to VEGFR-2 (14).
In the present study, a recombinant VEGFR-1 was
constructed for expression in vitro and in vivo. The
aim of this study was to examine the exact role of VEGFR-1 in
regulating VEGF-induced angiogenesis.
Materials and methods
Generation of recombinant human VEGFR1
domains (D)1-3/Fc (rhVEGFR-1)
cDNA (National Center for Biotechnology Information
reference NM_002019.3) encoding the human VEGFR-1D1-3 was cloned
using chemical synthesis and overlap extension polymerase chain
reaction. The primers used were as follows: Forward,
5′-GGAATTCCGATATCA CCATGGTCAGCTACTGGGAC-3′ and reverse, 5′-CGG
GATCCCGACTTACCTGTTTTATCATATATATGCACTGA G-3′. The cDNA sequences
were fused with a human immunoglobulin G (IgG) 1Fc fragment with an
intron included (provided by Dr H.Z. Liu, Beijing Institute of
Basic Medical Sciences) and inserted into a cloning vector
pcDNA3.1(+) (Invitrogen Life Technologies, Carlsbad, CA, USA),
resulting in pcDNA3.1-rhVEGFR-1.
Expression of rhVEGFR-1
Cell transfection
The Chinese hamster ovary (CHO)-K1 cells were
maintained at 37°C and under 5% CO2 in Dulbecco’s
modified Eagle’s medium (DMEM)/F12 (1:1) (HyClone, Logan, UT, USA)
containing 10% fetal bovine serum (HyClone). The cells were
transfected with pcDNA3.1-rhVEGFR-1 plasmid with Lipofectamine™
2000 reagent (Invitrogen Life Technologies). The stable,
higher-expression recombinant CHO (rCHO) cells were identified
using ELISA (Fangcheng Biotechnology Company, Beijing, China).
rCHO cell culture and purification of
rhVEGFR-1
The selected rCHO cell line was cultured in roller
bottles with DMEM/F12 (1:1) supplemented with 2% newborn calf serum
until the cells were ~90% confluent. The DMEM/F12 (1:1) was
replaced every two days. rhVEGFR-1 was purified from the cell
culture supernatant using rProtein A Sepharose™ Fast Flow (GE
Healthcare Life Sciences, Pittsburgh, PA, USA) affinity
chromatography. The supernatant was adjusted to pH 6.8 in 20 mM
Na2HPO4 and applied to protein A column (GE
Healthcare Life Sciences) by ÄKTAprime plus (GE Healthcare Life
Sciences), and rhVEGFR-1 protein was eluted using 0.5 M arginine,
pH 3.5 (15,16). The eluted protein was neutralized
immediately using 1/10 volume of 1 M Tris-HCl, pH 9.0.
Western blot analysis and protein
identification
Western blot analysis
The protein samples were separated using 10 and 6%
SDS-PAGE under reducing or non-reducing conditions and stained
using Coomassie Brilliant Blue R-250, prior to being transferred
onto a polyvinylidene difluoride membrane (Millipore, Billerica,
MA, USA) by electroblotting. The rhVEGFR-1 (1:50,000) was used as
the primary antibody and horseradish peroxidase (HRP)-conjugated
goat anti-human IgG antibody (1:20,000) was used as a secondary
antibody (BGI-GBI Biotech Co., Ltd., Beijing, China. The Immobilon™
Western Chemiluminescent HRP Substrate (Millipore) was used to
visualize the bound antibody and the final data were recorded using
X-Omat BT Film (Kodak, Atlanta, GA, USA).
Protein identification using nanoscale
liquid chromatography coupled with tandem high-definition mass
spectrometry (nanoLC-HDMS MS/MS)
The amino acid sequence of purified rhVEGFR-1 was
analyzed using nanoLC-HDMS MS/MS on a nanoACQUITY™ Ultra
Performance LC system (Waters Corp., Milford, MA, USA) and a Synapt
HDMS system, with nanospray ion source (Waters Corp.) in the
National Center of Biomedical Analysis (Beijing, China).
ELISA
Prior to measuring the association and dissociation
constants for the binding of rhVEGFR-1 to VEGF, the affinity was
first confirmed using ELISA. A 96-well plate was coated with
VEGF-165 and proteins were added to each line with a
1/2-concentration decrement. The maximum molar ratio of each
protein was ~1/6 to the coated VEGF. The plate was incubated at
37°C for 1 h and then the HRP-conjugated goat-anti-human IgG was
added (1:5,000), followed by 3,3′,5,5′-tetramethyl benzidine
dihydrochloride (Sigma, St. Louis, MO, USA). The plate was then
further incubated at 37°C. The optical density was measured at 450
nm using an ELISA microplate reader (Bio-Rad, Hercules, CA,
USA).
Surface plasmon resonance (SPR)
analysis
All procedures were performed using the Biacore 3000
system (Biacore AB, Uppsala, Sweden) in running buffer (10 mM Tris,
100 mM NaCl and 0.005% Tween-80, pH 7.5) at 25°C. The VEGF-165
(PeproTech, London, UK) was covalently linked to the carboxylated
dextran matrix of the sensor chip CM5 (Biacore AB). Kinetic
experiments were performed by injecting a series of concentrations
of rhVEGFR-1 (200, 100, 50, 25 and 12.5 nM, diluted in running
buffer, with or without 10 mM MgCl2) into the sensor
chip CM5. Baselines were regenerated with a 20-μl first injection
pulse of 10 mM glycine (pH 1.5) and a second injection pulse of
Borate 8.5 (10 mM disodium tetraborate and 1 M NaCl, pH 8.5;
Biacore AB), resulting in <1% loss of baseline per injection.
Dissociation and association rate constants (kd and ka,
respectively) were obtained using BIAevaluation 4.0 software
(Biacore AB), and the equilibrium dissociation constant, KD, values
were calculated by ka/kd.
Proliferation assay
The proliferation metrics of the human microvascular
endothelial cell line HMEC-1, provided by Professor Ding Jian
(Shanghai Institute of Materia Medica, Chinese Academy of Sciences,
Shanghai, China), were determined by suspending the cells at
2×105 cells/ml in ice-cold endothelial cell medium (ECM;
ScienCell Research Laboratories, Carlsbad, CA, USA) containing all
the necessary growth factors and serum supplements. Approximately
50 μl cell suspension was seeded into each well of a 96-well tissue
culture plate. Upon attachment to bottom of the plate, the ECM was
replaced with rhVEGFR-1, ranging between 0 and 800 ng/ml, and
rhVEGF-165 (28 ng/ml preincubation) with continuous incubation at
37°C under 5% CO2 for five days. Approximately 50 μl 5
μg/ml Calcein AM (Invitrogen Life Technologies) was then added to
each well, and the plate was incubated for an additional 30–60 min
under the same conditions. The fluorescence intensities at
excitation/emission wavelengths of 485/530 nm were analyzed using a
fluorescence plate reader (Thermo Fisher Scientific, Inc., Waltham,
MA, USA).
Migration assay
HMEC-1 were suspended in ECM at a density of
4×105 cells/ml. A total of 0.25 ml suspension was placed
into a Millicell chamber (Millipore), while 600 μl culture medium,
containing between 0 and 800 ng/ml rhVEGFR-1 and rhVEGF-165 (28
ng/ml preincubation), was added into the bottom well and incubated
at 37°C under 5% CO2 for 12 h. Following incubation, the
cells were removed from the top chamber. The insert plate was
transferred to another 24-well plate containing 0.5 ml/well Calcein
AM (Invitrogen Life Technologies) that was prepared in Hank’s
Balanced Salt Solution at a concentration of 5 μg/ml, and further
incubated at 37°C under 5% CO2 for an additional 30–60
min. Migrated HMEC-1 were detected and quantified using a
fluorescence plate reader (Thermo Fisher Scientific) with
excitation/emission wavelengths of 485/530 nm.
Chorioallantoic membrane (CAM) assay
Fertilized eggs from White Leghorn chickens
(Laboratory Animal Center, Beijing, China) were washed with
Benzalkonium Bromide (1:1,000) and incubated blunt-end-up in a
standard egg incubator at 37.8±0.5°C and 60–80% relative humidity.
After seven days of development, the eggs to be windowed were dried
using 75% ethanol. An electric engraving tool was used to make a
circular window measuring 15–20 mm in diameter in the air sac. The
round shell caps were aseptically removed and the shell membranes
were washed with warm, sterile phosphate-buffered saline (PBS). The
caps were then completely detached from the shell and the CAM was
removed using fine tweezers. Sterile 6-mm-diameter blank
concentration disks were first soaked with rhVEGFR-1 (ranging
between 0 and 25 pM in 5 μl PBS) and then aseptically transplanted
onto the CAMs. Bovine serum albumin (100 ng) was used as the
negative control. Shell windows were then covered with hyalo
adhesive tape, and the eggs were further incubated at 37.8±0.5°C
and 60–80 % relative humidity. Four days after transplantation, the
eggs were fixed in methanol. The CAMs were excised around the disks
and images were captured (17).
Directed in vivo angiogenesis assay
(DIVAA)
The nude mice were subcutaneously implanted with
semi-closed silicone cylinders (angioreactors). The angioreactors
(DIVAA Inhibition Assay, catalog no. 3450-048-IK) were purchased
from Trevigen Inc. (Gaithersburg, MD, USA). Angioreactors were
filled with 18 μl extracellular matrix premixed with angiogenic or
anti-angiogenic factors. The vascularization within the
angioreactors was quantified using an intravenous injection of
fluorescein isothiocyanate-dextran prior to the recovery of the
angioreactor, followed by spectrofluorimetry. The use of
immunofluorescence to examine the angioreactors showed the invading
angiogenic vessels at different developmental stages. The minimally
detectable angiogenic response occurs nine days subsequent to
implantation and with the addition of >50 ng/ml (P<0.01) of
either fibroblast growth factor-2 or VEGF. The present study was
approved by the Ethics Committee of the Chinese PLA General
Hospital (Beijing, China).
Statistical analysis
Statistical analysis was performed using SPSS
version 13.0 software (SPSS Inc., Chicago, IL, USA). The data are
shown as the means ± standard error of the mean, and were analyzed
by a one-way analysis of variance. A P<0.05 was considered to
indicate a statistically significant difference.
Results
Cloning of rhVEGFR-1
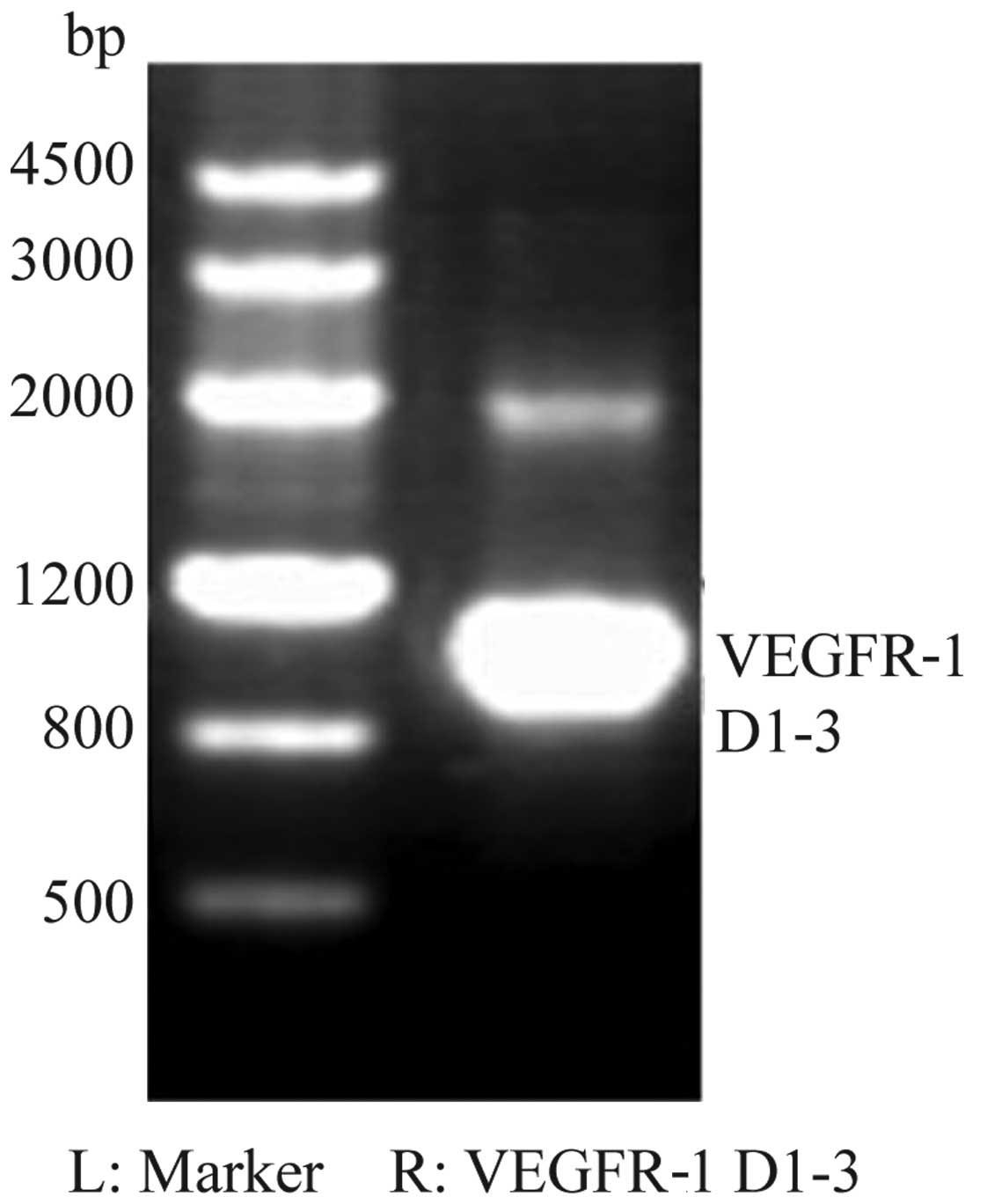
For the cDNA of VEGFR-1, the products on the agarose
gels were observed to be ~1,030-bp long, which was as expected
(Fig. 1). The synthesized sequence
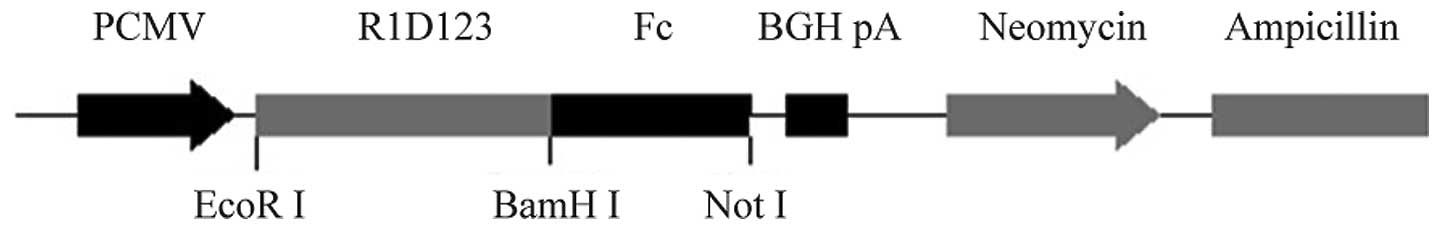
was then fused with human IgG1Fc (intron included) fragment and
cloned into the expression plasmid, resulting in pcDNA3.1-rhVEGFR1
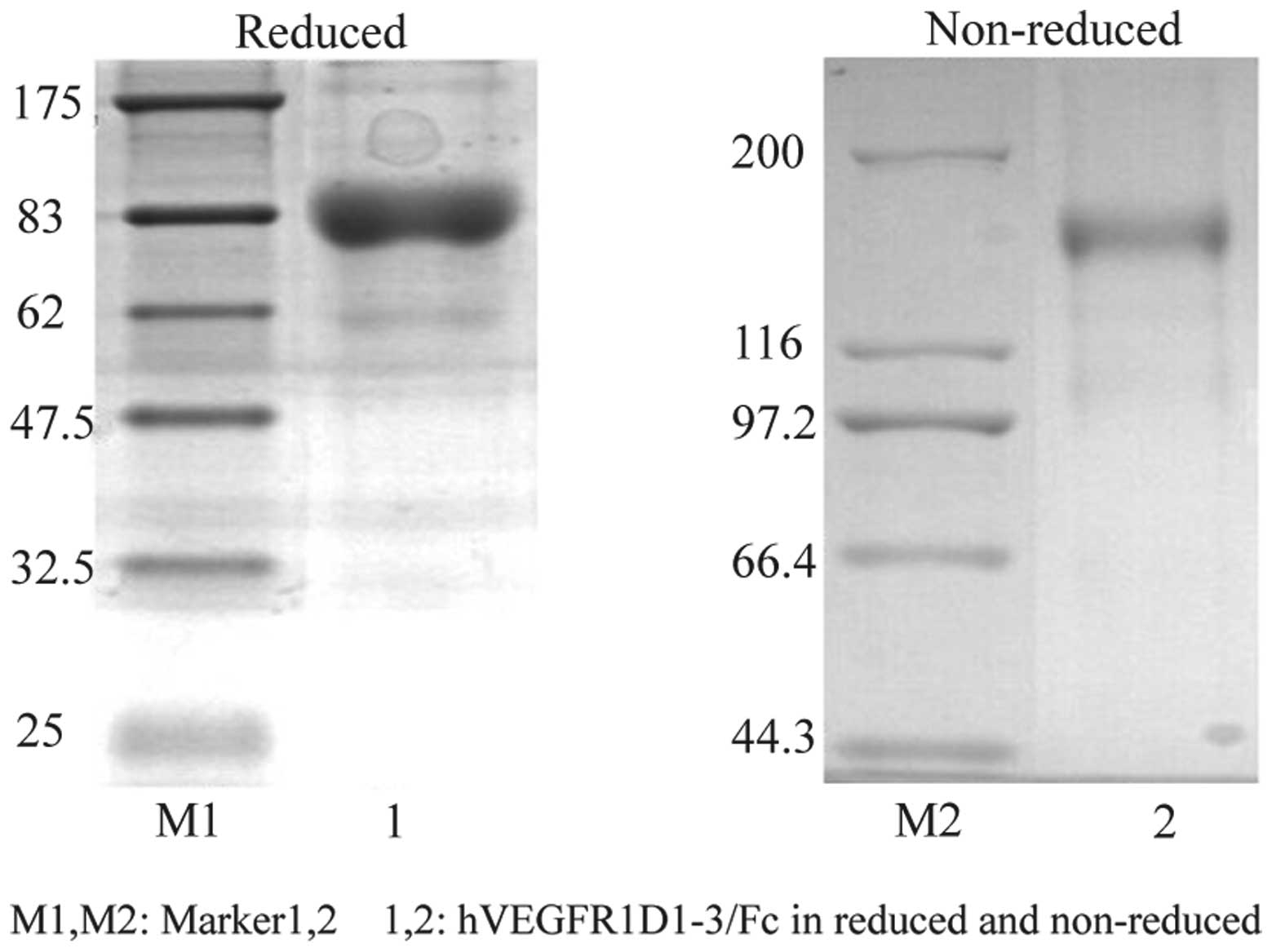
(Fig. 2). Following a 12-day
selective culture, the transfected cells formed clones. After a
further 12 days, the clones were screened for rhVEGFR-1 using ELISA
and confirmed using western blot analysis and peptide sequencing
(Fig 3). The evaluation of the
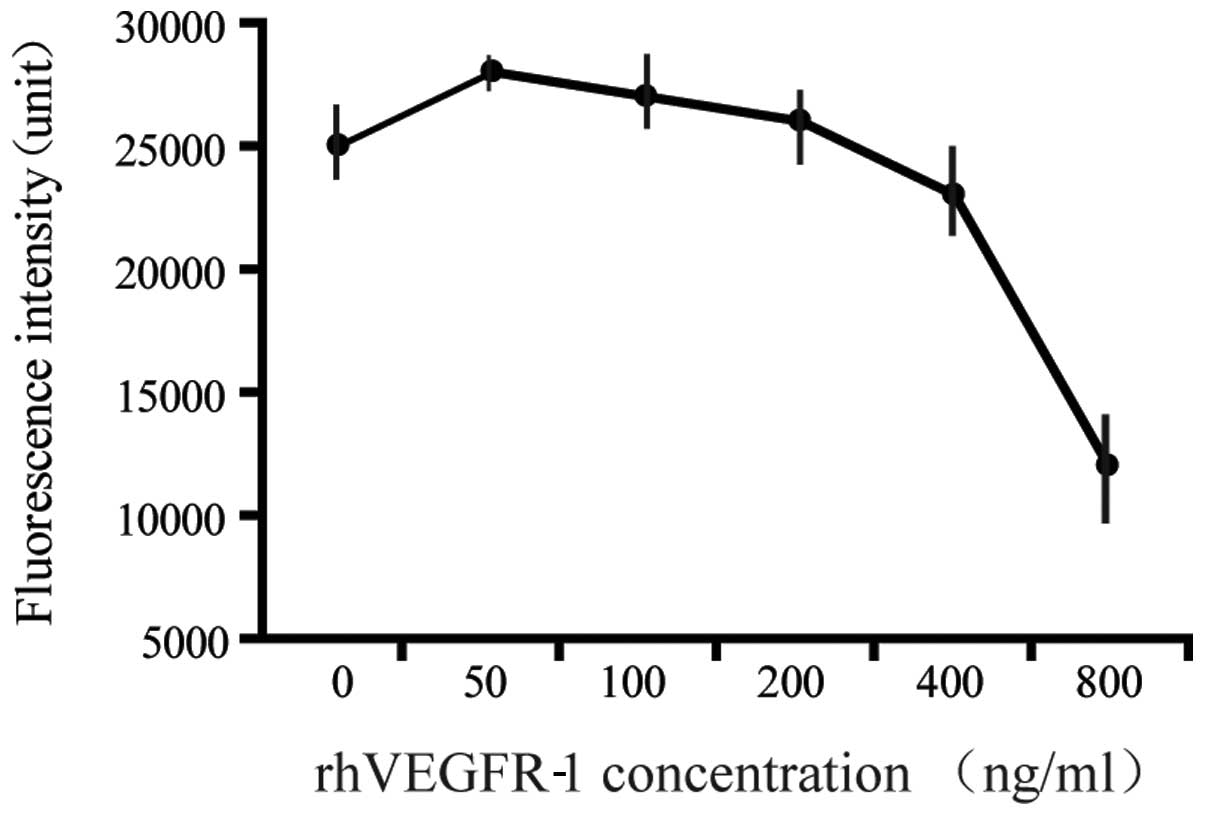
inhibitory effect of rhVEGFR-1 was performed by mixing the receptor
at concentrations ranging between 0 and 800 ng/ml with 28 ng/ml
rhVEGF-165 (Table I and Figs. 4 and 5).
 | Table IEffect of Mg2+ on the
affinity constant for the binding of rhVEGFR-1 to VEGF-165. |
Table I
Effect of Mg2+ on the
affinity constant for the binding of rhVEGFR-1 to VEGF-165.
| Mg2+
(mM) | ka (1/Msec) | kd (1/sec) | KA (1/M) | KD (M) |
|---|
| 0 |
2.60×105 |
7.42×10−5 |
3.5×109 |
2.85×10−10 |
| 10 |
1.37×105 |
5.07×10−5 |
2.7×109 |
3.71×10−10 |
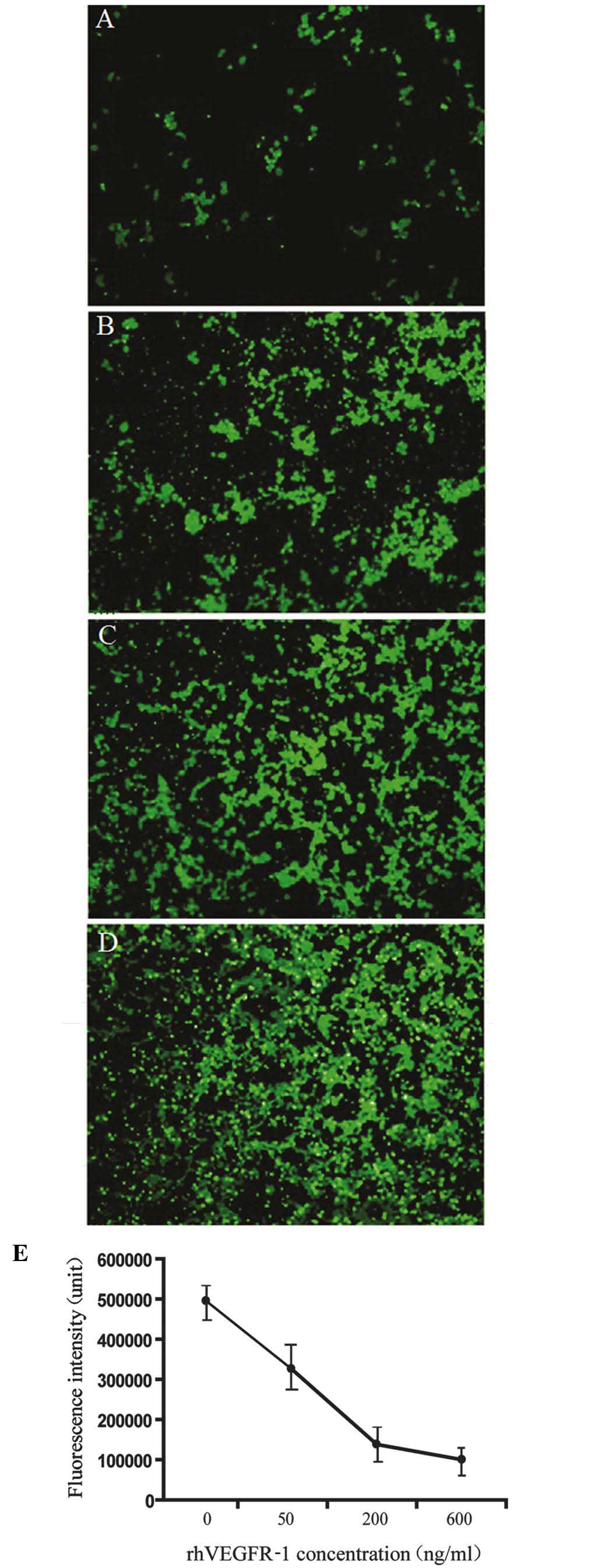
rhVEGFR-1 inhibits VEGF-induced HMEC-1
migration
An in vitro assay was performed using HMEC-1
on an angiogenesis cell migration plate. The cells were placed in
the top chamber, and the medium containing rhVEGF-165 (28 ng/ml)
premixed with rhVEGFR-1 (0–800 ng/ml) was added in the bottom
chamber. Following incubation for 12 h, rhVEGFR-1 was observed to
inhibit the cell migration with a linear correlation at
concentrations of 0–200 ng/ml, and the VEGFR-1 binding sites were
saturated at the range 200–800 ng/ml (Fig. 6).
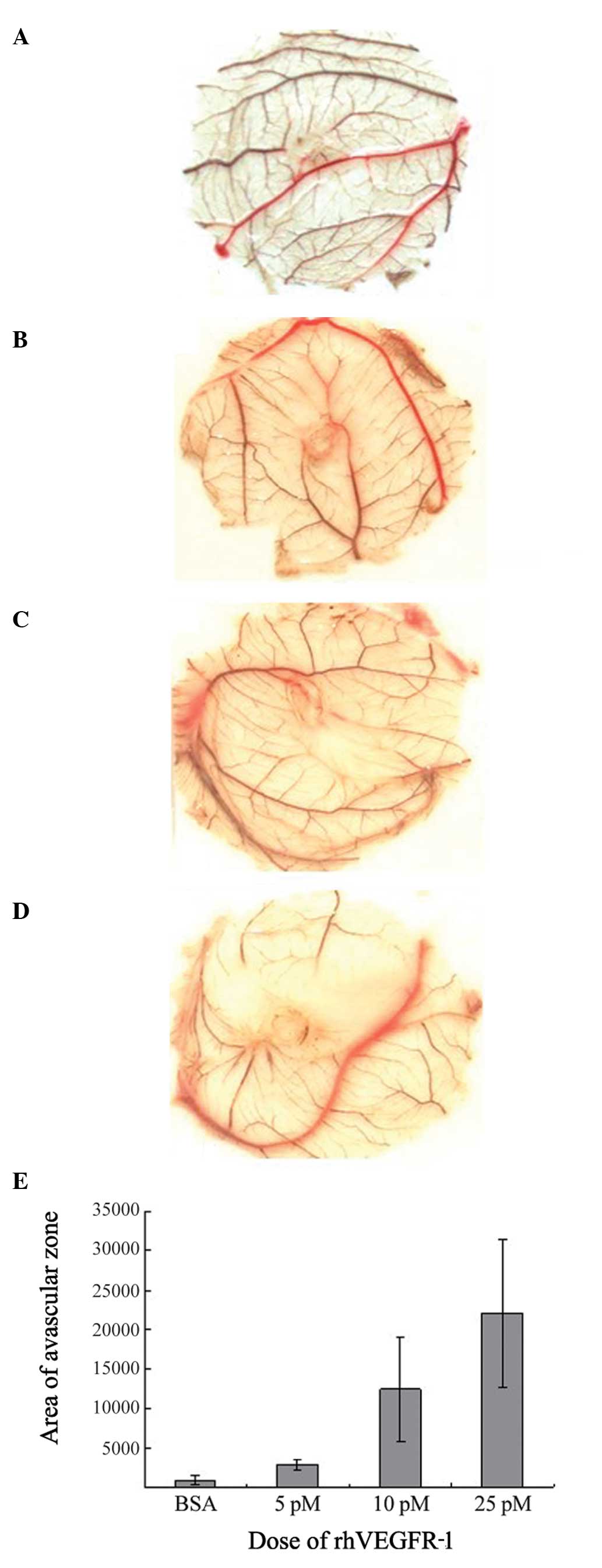
rhVEGFR-1 inhibits CAM angiogenesis
In order to determine the effect of rhVEGFR-1 on
angiogenesis, the area of the avascular zone in chicken embryos was
analyzed. It was found that it increased in size following the
application of rhVEGFR-1 in a dose-dependent manner, confirming the
inhibitory effect of rhVEGFR-1 on CAM angiogenesis (P<0.01;
Fig. 7).
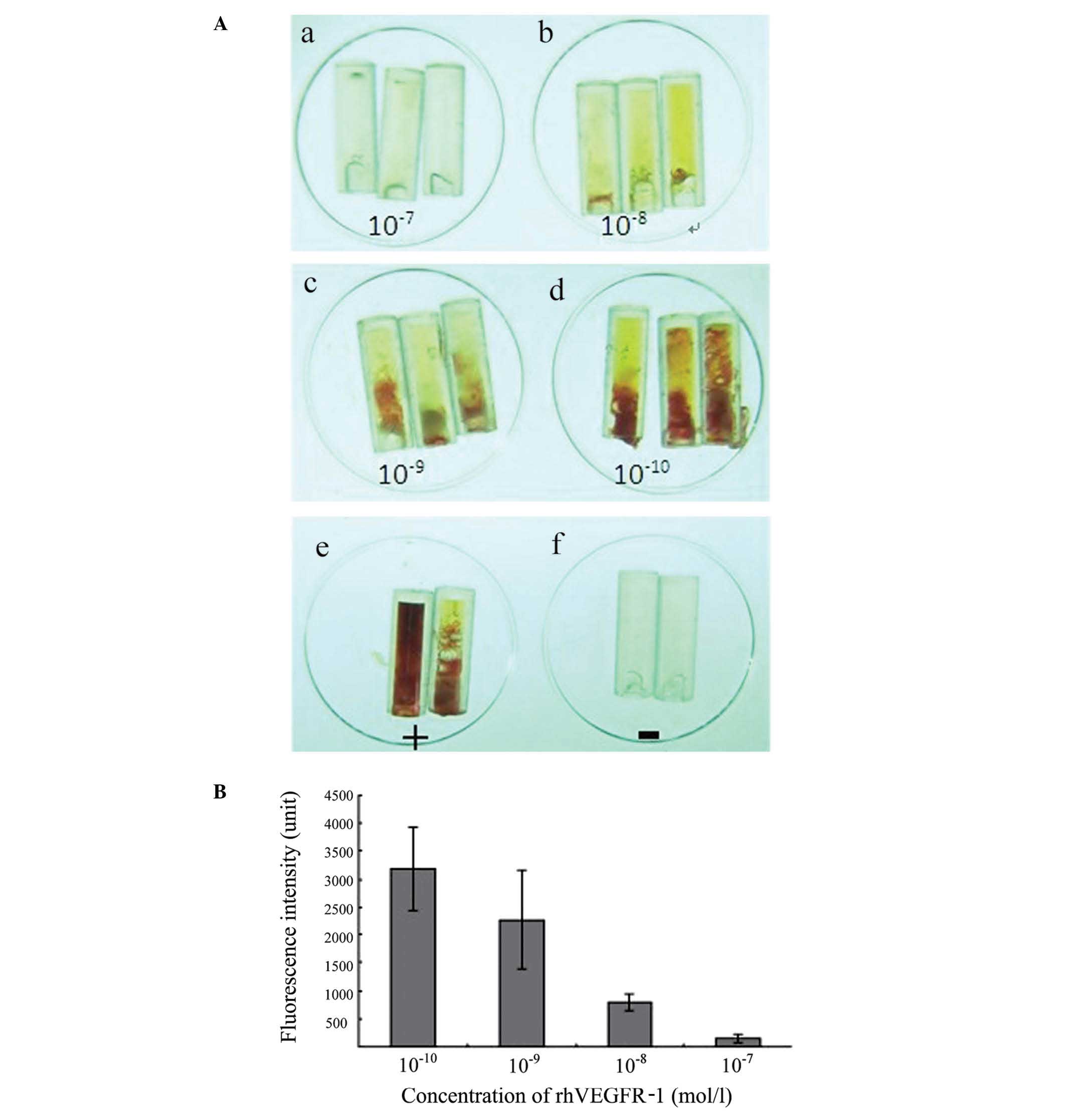
rhVEGFR-1 inhibits VEGF-induced
angiogenesis in vivo
In an in vivo experiment, angiogenesis was
analyzed in nude mice. It was found that rhVEGFR-1 inhibited
VEGF-induced angiogenesis in a dose-dependent manner. The
application of 160 μg/ml rhVEGFR-1 was shown to almost completely
block angiogenesis in nude mice (Fig.
8).
Discussion
The VEGF pathway is important in the regulation of
embryonic vascular development and tumor angiogenesis. Blockade of
the VEGF pathway effectively inhibits tumor angiogenesis and growth
in preclinical models. Thus, targeting VEGF may lead to novel
therapies for clinical application. Data has also suggested that
the potential functional roles of VEGF are associated with its
receptors, VEGF1 and VEGF2. The results of the present study
suggest that VEGF-121, VEGF-165, VEGF-C and VEGFR-2 may all be
involved in regulating carcinoma cell survival, proliferation (via
the autocrine signaling mechanism) and migration (via the paracrine
signaling mechanism).
The cation Mg2+ is an important
supplement in ECM. Notably, it was observed that rhVEGFR-1 bound to
VEGF-165 with high affinity without the obvious influence of
Mg2+, and the KD value of rhVEGFR-1 binding to VEGF-165
was different to that found in previous studies (13,18,19).
This may be explained by the fact that the ELISA, Scatchard
analysis and 125I-VEGF-165 competition analysis employed
in previous studies all had multiple steps and may have had poor
reproducibility. In the present study, the SPR analysis run on
Biacore had just one step and relatively high reproducibility,
suggesting that the data from the present study are more
reliable.
The dose-effect analysis revealed that rhVEGFR-1
could not block VEGF-165 activity with concentrations <200
ng/ml. The results suggested a molar ratio of 2:1 of rhVEGFR-1 to
homodimeric VEGF-165 to be necessary to reach a minimum inhibition.
A ratio of 8:1 appeared to be sufficient for maximum
inhibition.
In the in vivo experiment, the DIVAA
developed by Guedez et al (20) was found to be extremely sensitive.
Without any assistance, newly formed vessels were observed in the
angioreactors 12 days subsequent to transplantation.
In conclusion, in the present study an
hVEGFR1D1-3/Fc fusion protein was constructed to obtain stable
expression in an rCHO cell line. NanoLC-HDMS MS/MS was used to
confirm the purity of the proteins. The affinity constant for the
binding of rhVEGFR-1 to VEGF-165 was determined using SPR for the
first time and was found to be accurate. The rhVEGFR-1 showed
anti-angiogenic activity in cultured cells, in chicken embryos and
in nude mice. The present study demonstrated the important role of
VEGFR-1 in regulating VEGF-induced angiogenesis.
References
|
1
|
Leung DW, Cachianes G, Kuang WJ, Goeddel
DV and Ferrara N: Vascular endothelial growth factor is a secreted
angiogenic mitogen. Science. 246:1306–1309. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferrara N, Gerber HP and LeCouter J: The
biology of VEGF and its receptors. Nat Med. 9:669–676. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Breen EC: VEGF in biological control. J
Cell Biochem. 102:1358–1367. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Carmeliet P: Angiogenesis in health and
disease. Nat Med. 9:653–660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Carmeliet P: VEGF as a key mediator of
angiogenesis in cancer. Oncology. 69(Suppl 3): 4–10. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Clauss M: Molecular biology of the VEGF
and the VEGF receptor family. Semin Thromb Hemost. 26:561–569.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Claesson-Welsh L and Welsh M: VEGFA and
tumour angiogenesis. J Intern Med. 273:114–127. 2013. View Article : Google Scholar
|
|
8
|
Geng L, Chaudhuri A, Talmon G, Wisecarver
JL and Wang J: TGF-Beta suppresses VEGFA-mediated angiogenesis in
colon cancer metastasis. PLoS One. 8:e599182013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shen K, Ji L, Lu B and Wang Z: c-Jun
N-terminal kinase mediated VEGFR2 sustained phosphorylation is
critical for VEGFA-induced angiogenesis in vitro and in vivo. Cell
Biochem Biophys. 64:17–27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Weijts BG, Bakker WJ, Cornelissen PW,
Liang KH, et al: E2F7 and E2F8 promote angiogenesis through
transcriptional activation of VEGFA in cooperation with HIF1. EMBO
J. 31:3871–3884. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Shibuya M, Yamaguchi S, Yamane A, Ikeda T,
Tojo A, Matsushime H and Sato M: Nucleotide sequence and expression
of a novel human receptor-type tyrosine kinase gene (flt) closely
related to the fms family. Oncogene. 5:519–524. 1990.PubMed/NCBI
|
|
12
|
Terman BI, Carrion ME, Kovacs E, Rasmussen
BA, Eddy RL and Shows TB: Identification of a new endothelial cell
growth factor receptor tyrosine kinase. Oncogene. 6:1677–1683.
1991.PubMed/NCBI
|
|
13
|
Wiesmann C, Fuh G, Christinger HW, et al:
Crystal structure at 1.7 A resolution of VEGF in complex with
domain 2 of the Flt-1 receptor. Cell. 91:695–704. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Park JE, Chen HH, Winer J, Houck KA and
Ferrara N: Placenta growth factor. Potentiation of vascular
endothelial growth factor bioactivity, in vitro and in vivo, and
high affinity binding to Flt-1 but not to Flk-1/KDR. J Biol Chem.
269:25646–25654. 1994.PubMed/NCBI
|
|
15
|
Ejima D, Yumioka R, Tsumoto K and Arakawa
T: Effective elution of antibodies by arginine and arginine
derivatives in affnity column chromatography. Anal Biochem.
345:250–257. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Arakawa T, Philo JS, Tsumoto K, et al:
Elution of antibodies from a Protein-A column by aqueous arginine
solutions. Protein Expr Purif. 36:244–248. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shi ML, Duan HF, Xu ZP, Hu XW and Chen HP:
Optimization of applying chick chorioallantoic membrane to
angiogenesis assay. Letters in Biotech. 19:566–568. 2008.(In
Chinese).
|
|
18
|
Herley MT, Yu Y, Whitney RG and Sato JD:
Characterization of the VEGF binding site on the Flt-1 receptor.
Biochem Biophys Res Commun. 262:731–738. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Holash J, Davis S, Papadopoulos N, et al:
VEGF-Trap: a VEGF blocker with potent antitumor effects. Proc Natl
Acad Sci USA. 99:11393–11398. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Guedez L, Rivera AM, Salloum R, et al:
Quantitative assessment of angiogenic response by the directed in
vivo angiogenesis assay. Am J Pathol. 162:1431–1439. 2003.
View Article : Google Scholar : PubMed/NCBI
|