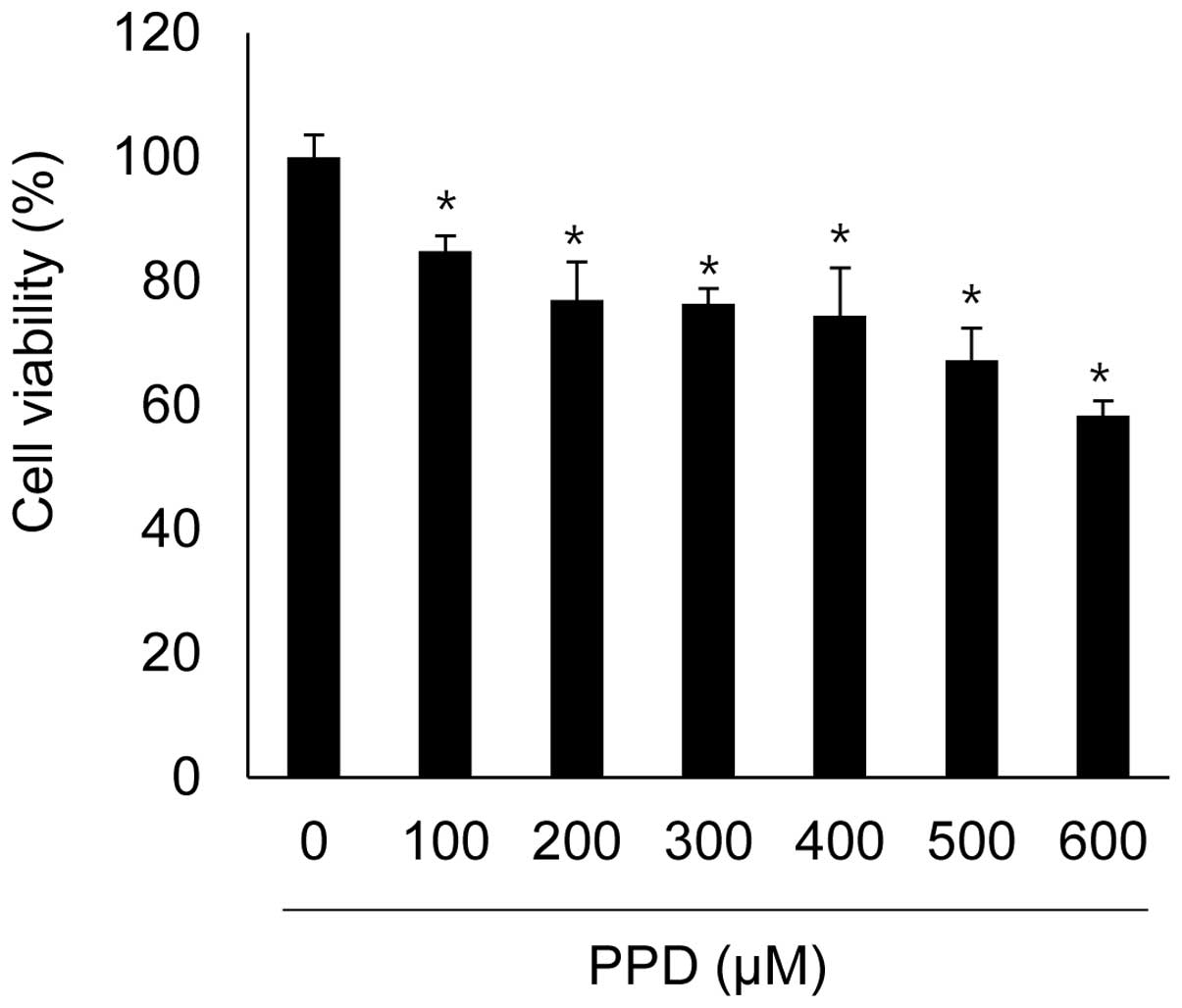
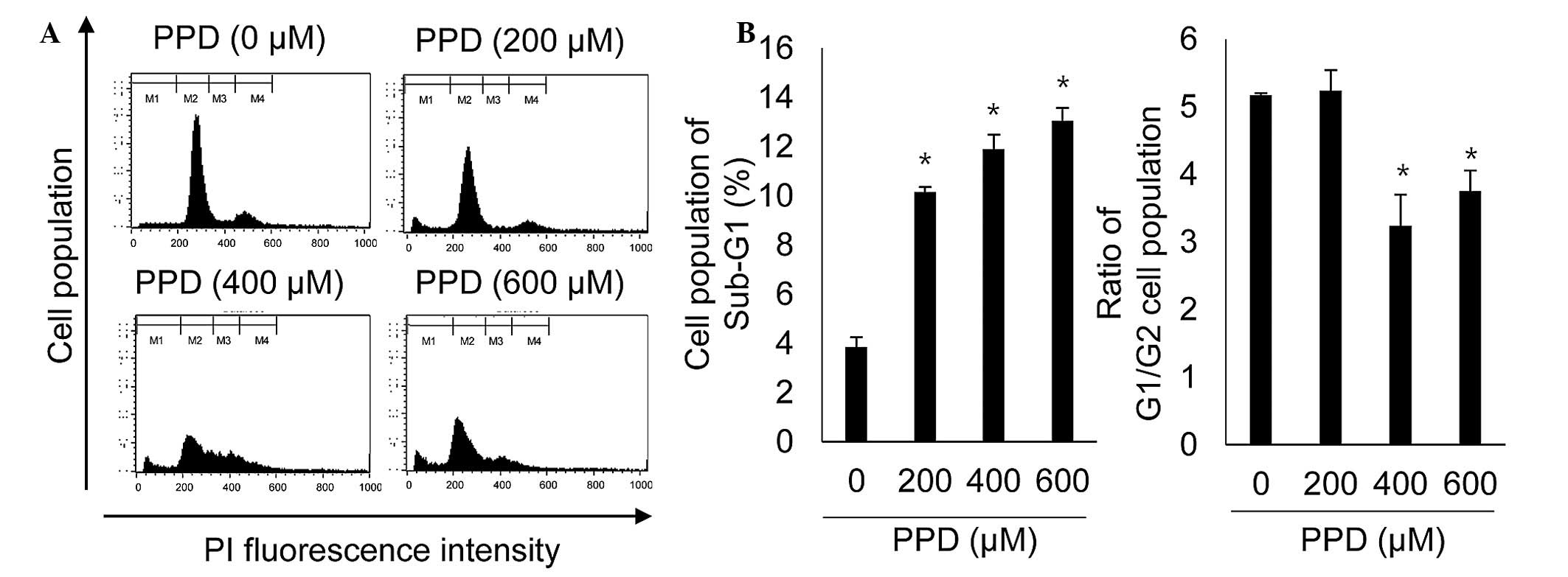
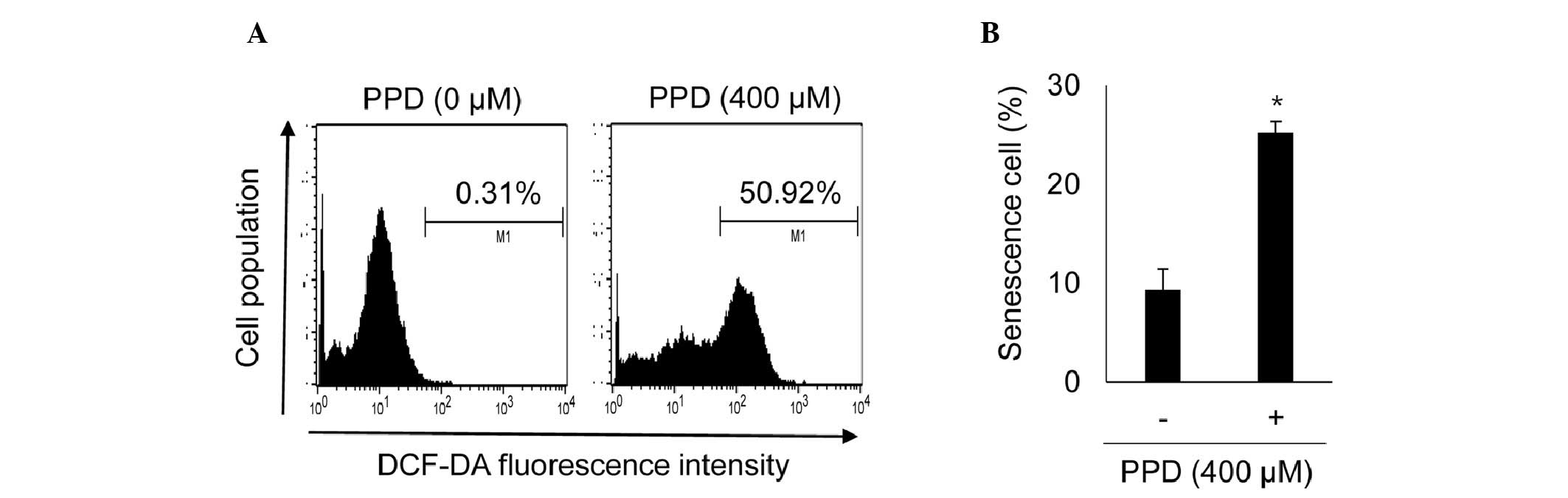
| has-let-7a-3p | CNR1, TFRC, TGFBR1,
F3, LRP2, ID2, VCAM1, SLC1A2 | TCF7L2, JUP,
ITGA2 | FOXO1, ITCH, TLR4,
PDE5A, TGFBR1, F3, CNR1, TCF7L2, MALT1, SGK3, SOX2, CUL1, RHOA,
HIPK2, CUL5, JAK2, IGF1R, MEF2C, LRP6, SMO, SKIL, ECT2, ROCK1,
OPA1, ID1, PKN2, RNF34, CREB1, APP, HIP1, PAK2, RAD21, UBE2B, DSG2,
SOS2 | TLR4, PDE5A, MALT1,
SGK3, MEF2C, SOX2, CUL1, IGF1R, CDC42, PLAG1, HHIP, NRP3, ID4,
CDC73, NR5A2, PROX1, TOB1, RAX6, SNAI2, NR2F2, SALL1, NRP2, UTP20,
EMP2, LRP6, SMO, PDSSB, MTSS1, TGFBR1, F3, VCAM1, LRP2, ID2,
TCF7L2, RHOA, HIPK2, CUL5, JAK2, FOXO1, ITCH, TCF3, JAG1, FZD3 |
| hsa-miR-150-3p | TFCP2L1 | – | INHBA, IL1A, RHOA,
ATG7, BCL3, MECOM, CRH, MPO, MAP3K5, NF1, ARHGEF7 | RHOA, NF1, PHOX2B,
CD164, INHBA, IL1A, MECOM, EVI5, TIMELESS, FER, NDN, FYN, NRP1,
BTRC |
| hsa-miR-1181 | – | – | – | – |
|
hsa-miR-1207-5p | RXRA, TERF1, WNT16,
TMEM115, GNAO1 | DHCR24, TFAP2A,
ATP7A | EDAR, ACAA2, GDNF,
ALX4 DHCR24, TFAP2A, MAP2K5, PDPK1, ATG7, PACS2, TRIO, RXRA, PIM1,
MSX1, IGF1R, ERBB2, PDE1B, BIRC6, CDH1, TERF1, APBB1 | MAFG, FOXO4, PBX1,
RARA, ERBB2, RAC2, CSF1, MXD4, RXRA, DHCR24, TMEM115, IGF1R, PIM1,
MSX1, BIRC6, TFAP2A, WNT16, MAP2K5, TSC2, TENC1, STAT6, MGFEB,
SIX5 |
|
hsa-miR-1226-5p | TBX3, EDNRA | DDR1 | GLI2, DICER1, NF1,
MLLT11, TBX3, EDNRA, PTK2, ING4, SKP2, RTN3, SORT1, PEG10, BCL2L2,
EIF2AK3 | GLI2, DICER1, NF1,
NACC2, IL9R, EMX2, HOXC10, SIX2, TBX3, EDNRA, PTK2, ING4, SKP2,
SIX1, DDR1, TENC1, MLL2, FKTN |
|
hsa-miR-125a-3p | ULK3 | – | RHOA, TAOK1 | RHOA, TAOK1,
TIRAP |
| hsa-miR-128 | EDNRA, MET, CNR1,
SIRT1, F3, MAPK14, MNT, MME, TGFBR1, GRB2 | APC, NGFR,
COL5A1 | NGFR, PIK3R1,
HIPK2, BMI1, CITED2, ERCC1, PHB, SIRT1, TGFBR1, CYLD, SLIT1, APC,
DAPK1, SOS1, BTG2, CASP8, MAP2K4, MAGI3, PDPK1, MNT, CNR1, MAPK14,
FOXO1, NTRK2, TRIM32, MCF2L, WNK3, FOXQ1, PLK2, GSK3B, BCL3 | EIF2S2, FOXO4,
NRP2, GAB1, SIRT1, TGFBR1, EDNRA, F3, ERCC1, PHB, CITED2, BTG2,
RUNX1, SOX11, IRS1, TSC1, NRG1, ASPH, FOXP2, ZEB1, PRKX, JAG1,
RAP1B, FZD3, HGF, HIPK2, FOXO1, BMI1, NTRK2, TRIM32, ADAM10, MNT,
APC, NGFR, PIK3R1, FBXW7, VEFGC, EBI3 |
| hsa-miR-134 | EDN1, CISD2 | COL5A2 | CXCR2, PAWR, TCHP,
BDNF, EDN1, YAP1, STAT5B, PAX8, HIPK2, PDCD7, BARD1, SMAD6, CREB1,
ITSN1 | EDN1, YAP1, CXCR2,
PAWR, CDK13, PKD2, OSMR, KRAS, HIPK2, BDNF, ASCC3, XCL1, STAT5B,
MAGI2 |
|
has-miR-138-2-3p | RPS6KB1, HMGA2,
CHEK2, PNPT1 | CTNNB1, APC,
SHH | CHEK2, APC, NF1,
TP53INP1, FOXO3, MEF2C, SIX1, POLB, HIPK2, SHH, BCL2L1, SLIT3,
HMGA2, HMGA2, RPS6KB1, BECN1, PEG3, RNF152, GSK3B | NF1, HIPK2, BCL2L1,
BECN1, HMGA2, HMGA2, APC, SHH, FOXO3, MEF2C, SIX1, TOB1, OTP, FER,
FRK, PGR, WNT2, LHX9, CD34, E2F3, POU3F2, FGF5, PAX6, KLF5, PLAG1,
STAT3, MBD2, CDK13 |
|
hsa-miR-146b-5p | NOX4, SMC5, CCL5,
LRP2, ATR | APC, ERRFI1,
COL4A3 | CCL5, APC, ADAM17,
CCK, COL4A3, ROBO1, SORT1, SIAH2, TRAF6 | CCK, ERBB4, ROBO1,
FZD3, CCL5, LRP2, NOX4, APC, ADAM17, CD80, HHIP, NRAS, SMAD4,
NUMB |
|
hsa-miR-148b-3p | PTEN, RTN4, SCAP,
UCP3, SERPINE1, MNT, IL15, LRP2, CANX | APC, ATP7A,
COL2A1 | PPARG, MDM4, MITF,
USP7, SULF1, IGF1, ROBO1, SOS1, ETS1, ROCK1, HIPK3, MAX, PTEN,
SERPINE1, CDKN1B, MNT, RTN4, APC, COL2A1, SIK1 | PTEN, SERPINE1,
MNT, IL15, MITF, CDKN1B, E2F7, NRAS, APC, PPARG, SULF1, MDM4, LRP2,
ETS1, IGF1, ROBO1, FLT1, MXD1, CDK13 |
| hsa-miR-17-3p | PTEN, SERP1,
TGFBR1, HMGA2, ICAM1, DLD | BCL11B, | PTEN, TGFBR1, BCL6,
BDNF, HMGA2, BCL11B, MAPK1, PPARG, PIK3CA, TRIM2, MEF2C, DIDER,
TOPOS, WNK3, TIA1 | PTEN, TGFBR1, BCL6,
BDNF, HMGA2, BCL11B, MAPK1, MEF2C, DIDER, TOPOS, PPARG, TSC1,
NF1B |
| hsa-miR-181d | PTEN, ATM, RPS6KB1,
VCAM, TIMP3, MME | – | RPS6KB1, PTEN, ATM,
CUL5, NOTCH2, GATA6, BCL6, BIRC6, CREB1 | RPS6KB1, PTEN, ATM,
CUL5, NOTCH2, GATA6, BCL6, BIRC6, RUNX1, VIP |
| hsa-miR-185-5p | RELA, CDK6, MAPK14,
ACAN | CDSN, EDA | RELA, MAPK14,
BRCA1, AR, MAP2K6, MED1, ERCC3, APP, PAK6, BCL2L2 | RELA, CDK6, MED1,
BRCA1, AR, CDK2, BAI1, INSIG1 |
| hsa-miR-195-5p | BCL2, PDCD4,
MAP2K1, MNT, DLD | WNT7A, ATP7A,
EDA | MNT, BCL2, PDCD4,
WNT7A, NOTCH2, IGF1R, VEGFA, FOXO1, RAF1, BCL2L2, BFAR, SIAH1 | RAF1, CCND1, FBXW7,
E2F3, MNT, BCL2, MAP2K1, IGF1R, HMGA1, WNT7A, NOTCH2, VEGFA, FOXO1,
ABCB1, FKBP1B |
| hsa-miR-197-3p | TERT | DDR1 | TERT, BRCA1, MAPK7,
ING3, PSMD1, MEF2A | BRCA1, DDR1, FTO,
LHX9, ASCC3, PDGFRA, INSL4 |
| hsa-miR-202-3p | NOX4 | TFAP2B, ERRFI1 | TRAF5, YAP1, MEF2C,
RAG1, DICER1, DUSP1, TRAF6 | NOX4, TFAP2B,
DICER1, TRAF5, YAP1, MEF2C, GPC3 |
| hsa-miR-214-5p | CDK6, KL, SLC1A2,
TERF2 | BCL11B, ERRFI1 | BCL11B, COL18A1,
OSR1, RERP, E2F2, TP53I3 | CDK6, BCL11B,
COL18A1, OSR1, DISC1, ABCB1, SMAD4, KLF5 |
| hsa-miR-23a-5p | P2RY2, DCN | TFAP2C, WNT10A | HSPA9, CLIP3,
CARD8, NMT1 | TLR4, FOXC1,
ADAM10, CSF1, MAPRE1, NCK2 |
| hsa-miR-28-5p | – | APC, CDSN | APC, NF1, BAG1,
WNK3 | APC, NF1, SESN1,
TRIM27 |
|
hsa-miR-301a-3p | PTEN, EDN1, CDKN1A,
LRP2, MET CANX, UCP3 | PTGES3, EDA | IRF1, PRNP, RUNX3,
ROBO1, SIK1, RAG1, ZMAT3, TRIM2, SULF1, USP28, SOS2, PAK6, EDN1,
TGFA, MDM4, E2F2, PTEN, CDK5R1, CDKN1A, USP47, ROCK1, ROBO2 | ROBO1, SULF1,
USP28, TSC1, MDM4, PRNP, RUNX3, LIPA, MET, IRF1, TGFA, CDK5R1,
WNT2B, HOXA3, RBFOX2, CDKN1A, PTEN, EDN1, E2F7 |
| hsa-miR-30a-3p | CACYBP, HMGA2, | – | PTEN, HMGA2,
CDKN1B, MEF2C, BIRC6, TRIM2, | CDKN1B, MEF2C,
CREBBP, BLM, BIRC6, SUZ12, |
| hsa-miR-30e-3p | PTEN | – | CREB1, AKAP13,
SOS1 | PTEN, MXD1, ODZ1,
HMGA2 |
| hsa-miR-324-5p | MSH6 | – | MSH6, VDAC1,
PSME3 | CTLA4, FYN,
PBX1 |
| hsa-miR-342-3p | ADRB1, EDRNA, BCL2,
CASP2 | SOX21, EDA, COL5A2,
COL1A2, TCF15 | BRCA1, BCL6,
BCL2L1, E2F1, BCL2, EDNRA, DAPK1, TIA1, NOTCH2, CASP2, ERBB4, PAK2,
DUSP6, DRAM1 | BCL6, BCL2L1, E2F1,
ERBB4, BCL2, NOTCH2, IRAK4, ID4, EDNRA, BRCA1, TACC1, CSF1, PGR,
KAT2B |
| hsa-miR-3651 | MSH6 | – | MSH6, KLHL20,
HIPK1 | HIPK1, PRMT5 |
| hsa-miR-3653 | HMGA2, PTEN,
SOCS2 | TFAP2C | PTEN, HMGA2, SOCS2,
MEF2C, SORT1, CREB | PTEN, HMGA2,
ADAM10, MEF2C, TFAP2C, RUNX1 |
| hsa-miR-3656 | – | – | – | – |
|
hsa-miR-3663-3p | FAS, CASP2, CDKN1A,
SREBF1 | | BCL11B, APC,
ADAMTS2, COL1A1, COL3A1 CASP2, CDKN1A, DUSP2, COMP | BCL11B, APC, USP28,
DSG2, MEF2D, FAS, ARAF, PTH1R, APC, TGFB2, FABP1, CDKN1A, FAS,
LIPG, CD80, BCL11B, USP28, DBN1, VSIG4, IL20RB |
| hsa-miR-369-3p | SIRT1, WRN, MAP2K1,
NUAK1, BRCA2, ADH5 | BCL11B, ATP7A, | HDAC2, WRN, WNT5A,
AHR, CARD11, JAK2, BMP2, XIAP, SATB1, PAWR, SOX2, FGF2, SOS1, OPA1,
NDNF, MEF2D, SOX4, HGF, BIRC3, SLIT3, BRCA2, SIRT1, BCL11B, RASSF6,
NEUROD1 | USP8, CEBPA, ODZ1,
PROX1, CARD11, JAK2, BMP2, XIAP, MEGF10, FGF5, ZEB1, PAX6, SATB1,
PAWR, SOX2, FGF2, SIRT1, MAP2K1, ADAM10, WNT5A, SOX4, AHR, HGF,
NUAK1, BRCA2, HDAC2, CD47, ZEB2, VEGFC, WNT3 |
| hsa-miR-369-5p | – | – | – | – |
| hsa-miR-370 | PNPT1 | APC | APC, SMO, RAF1,
CCL21, BFAR, PIK3CA, AKAP13 | APC, SMO, RAF1,
PRTFDC1, FGF7, SMARCD3, CASK, SBDS, RXRB |
|
hsa-miR-371a-5p | – | LEF1, ATP7A | LEF1, STK4, CITED2,
BARD1 | LEF1, STK4, SOX2,
COL8A1 |
|
hsa-miR-378a-3p | – | – | MNAT1, IGF1R, SKP2,
NAE1 | MNAT1, IGF1R, SKP2,
TOB2 |
| hsa-miR-3926 | INO80C | – | TMX1, BIRC6, SATB1,
IGF1R, CKAP2, | TMX1, BIRC6, SATB1,
IGF1R, CDK7, WDR6, ABCB1, ARNT |
| hsa-miR-409-5p | – | – | CREB1, NAIP | UBE2L3,
FGFR1OP |
| hsa-miR-410 | HMGA2, DCN, TOP2A,
EDN1, SOCS3, CDK1, PTEN, ADM, LRP2, SMC5 | – | BAG1, ELMO2, AHI1,
MAGI3, HMGA2, EDN1 PTEN, SAV1, CD38, TIAL1, MDM2, BMP2, XIAP1,
TBX5, FGF2, ARAF1, TOP2A, ETS1, BDNF, PTK2, PEG3, SNAI1 | HMGA2, CDK1, EDN1,
PTEN, BDNF, PTK2, TIAL1, MDM2, COL4A3, NBN, XIAP, YAP2, ADM, LRP2,
WNT16, EST1, ISL1, NUMB, TOB1, PEX2, E2F7, CBLB, FGF9, KLF5, IRS1,
STAT3, LIFR, FST |
| hsa-miR-423-5p | MSH6, GSN, PML,
ILK, CASP2 | – | CASP2, PML, ILK,
APBB1, MSH6, GSN, LRP5, BAG1 | PML, ILK, PTH1R,
LRP5, BAP1, KDM4C, ELF4 |
| hsa-miR-4271 | HMGA1, SLC6A3,
AMFR | – | FOXO3, EI24, MEF2D,
MAPT, SPN, CBX4, WNT7B, DAPL1, MAPK1, HMOX1, THRA | WNT7B, FOXO3, SPN,
MBD2, HMGA1, MAPK1, HMOX1, MXD11, MLL2, PDGFB, FOXO4, COL4A3BP |
| hsa-miR-4291 | VDR | – | VDR, PIM1, DDX5,
CREB1 | VDR, PIM1, PGF,
IRS1, MCC |
| hsa-miR-431-5p | CANX | TCF7L1 | TNFSF8, SOX9, IRS1,
HIPK3 | TNFSF8, SOX9, IRS2,
NKX6 1 |
| hsa-miR-431-3p | – | – | – | – |
| hsa-miR-4317 | CTNNA1 | APC | CTNNA1, APC,
IRS2 | CTNNA1, APC, IRS2,
ESR1 |
| hsa-miR-487a | SIRT1, CNR1,
MARCH5 | – | SIRT1, CNR1, TMX1,
SGK3 BMI1, SGK3 | SIRT1, TMX1, |
| hsa-miR-500a | MORC3 | – | AHR, BTG1, SULF1,
HDAC2, ERBB3, SRGN, GRID2 | MORC3, AHR, BTG1,
HDAC1, ERBB3, EMP1, SOX11, PRG4, FGF9, PEX2, AKIRIN2, BLM |
| hsa-miR-501-5p | NR3C1, MAP2K1,
TFRC | – | NR3C1, CUL1, MEF2C,
BMI1, ING3, TMBIM4, TRIM39 | NR3C1, CUL1, MEF2C,
BMI1, MAP2K1, BCAT1, NOX1 |
| hsa-miR-505-3p | CHEK1 | – | TBX5, HMGB1 | TBX5, ATP8A2,
IL11 |
|
hsa-miR-513a-5p | PRKCD, NEK6, MORC3,
MAP2K1, CDK6, ACAN, SERP1 | APC, SFN, WNT7A,
TFAP2B | PRKCD, NEK6, APC,
WNT7A, GATA3, XIAP, STAT1, MITF, BCL6, PPARG, ISL1, GZMB, MED1,
HDAC2, SFN, EYA1, TRIO, SOS1, ECE1, VAV2, USP47, HGF, BCL2L11,
WNK3, OCLN, MAL | PRKCD, MORC3, CDK6,
APC, SFN, WNT7A, TFAP2B, BCL6, PCM1, RXRB, PDXK, FOXP2, GATA3,
STAT1, MITF, BTG1, HDAC2, XIAP, ISL1, MAGI2, MAP2K1, XRCC5, PPARG,
E2F7, NFIB, VIP, ATF3, PURA, CSF1, KRAS |
| has-miR-513b | TGFBR1, ID2, SIRT1,
RNT4, ADH5 | BCL11B, TFAP2A,
ATP7A | BCL2L1, PPARG, BMF,
PAK6, TGFBR1, SIRT1, RNT4, PAK1, SNAI2, BCL11B, MOAP1, TCF7L2,
ERBB4, XIAP, CREB1, API5, BCL2L11 | TGFBR1, SIRT1,
MARCKSL1, SNAI2, ERBB4, PAK1, XIAP, LIFR, FER, BCL11B, AGTR1,
GATA2, VIP, BCL2L1, IRF2, ID2, FOXP2, WDR6, VTI1B |
| hsa-miR-539-5p | SMC5, SLC1A2 | TCF7L2, TFAP2A | CUL2, ERBB4, ELMO2,
SET, TCF7L2, TFAP2A, HDAC2, TRIM2, PPARGC1A, SIX4 | ERBB4, FRS2, HDAC2,
PBX1, TCF7L2, CUL2, CD47, DAB2, TFAP2A, VAX1, FKTN |
|
hsa-miR-548c-3p | NR3C1, PTEN, MORC3,
SMC5 | SHH, BCL11B,
TCF7L2 | MITF, ERBB4,
CITED2, XIAP, SGK3, TAOK1, RAG1, HIPK3, NR3C1, PTEN, SHH, TCF7L2,
BTG1, LRP6, GATA6, REST, TIA1 | GATA6, REST, ERBB4,
SGK3, NR3C1, PTEN, MORC3, SHH, ADAM10, FER, HHIP, KRAS, TCF7L2,
BTG1, LRP6, MITF, CITED2, XIAP, SSR1, EREG, SOX11, RUNX1, PURA,
E2F3 |
| hsa-miR-572 | – | – | – | – |
|
hsa-miR-642b-3p | HMGA2, MET, CDKN1A,
PTEN | BCL11B | BCL11B, PDCD10,
WT1, RB1, HMGA2, PTEN, CDKN1A, EIF2AK2, IFNG, MAPK8 | BCL11B, PDCD10,
WT1, RB1, CDKN1A, NR2F2, PROX1, PTEN, EIF2AK2, HMGA2, PAX6, AMBN,
PHOX2B |
| hsa-miR-650 | MADCAM1, SHC1 | – | LRP6, MITF, PIM1,
CUL2, JAK3, MUL1, CSTB, AXL | SHC1, LRP6, MITF,
PIM1, CUL2, JAK3, ERF, FTO |
| hsa-miR-660-5p | – | TFAP2B | TFAP2B, BRCA1,
HIPK1 | TFAP2B, BRCA1,
HIPK1, SF1 |
| hsa-miR-762 | RELA, PML | – | RELA, PML, MAPK1,
MEN1, PAK4, ITCH, BCL6, PPARD, ADD1, PAX7, ITGB2, MYO18A | RELA, PML, MAPK1,
MEN1, PAK4, ITCH, BCL6, PPARD, MMP14, BAP1, FTO, WAR5, FSCN1,
XIRP1. TENC1 |
| hsa-miR-770-5p | HMGA2, CNR1 | RYR1 | HMGA2, CNR1, MED1,
SGK3, XAF1, HELLS, MAP3K1 | HMGA2, BTG1, LRP6,
SGK3, HELLS, CCND2, MED1, PBX1, TSG101, NFIB |
| hsa-miR-874 | DDC | – | ESR1, HIPK2, PAK7,
THRA, SORT1 | RXRB, COMT, BMRP2,
NPR1, ESR1, HIPK2, PAK7, CD276, MXI1 |