Introduction
Buckwheat, an important functional food planted at
high latitudes and in colder climates, is a common dietary
component in East Asian countries (1). It is high in protein, contains a
number of amino acids and is regarded as a popular food globally,
particularly in Asia. Buckwheat protein has high biological value
due to its well-balanced amino acid composition and its high level
of lysine (2,3). Protease inhibitors are widely
distributed in nature and are found in numerous animals, plants and
microorganisms. They are important in maintaining the balance of
proteolytic enzymes in vivo. In addition, they regulate
endogenous proteases during germination and protect plants against
insects and microorganisms (4–6). Of
note, the inhibitors have been found to exhibit anti-carcinogenic
activities and act as cancer-preventive and anti-inflammatory
agents (7–9). Protease inhibitors are capable of
inducing the apoptosis of cancer cells in vitro and they
have drawn attention as potential anti-cancer agents (10). The Bowman-Birk inhibitor family of
proteins attained from soybeans are associated with
anti-inflammatory and anti-carcinogenic activities (11), and are potentially relevant
anti-tumor agents, particularly with regard to colon cancer
(12).
Numerous lines of evidence have suggested that
protease inhibitors may induce apoptosis in various tumor cell
lines; however, the underlying mechanisms of their anti-tumor
activity remain to be elucidated. Induction of tumor cell apoptosis
is a common mechanism of action of cancer therapeutics (13,14).
Caspase-3 is one of the key initiators of apoptosis via the
mitochondrial pathway and an essential factor for the activation of
the caspase cascade (15–17). Recent studies have revealed that
the activation of caspase-9 also induced the activation of the
caspase cascade, triggering apoptotic events and inducing cell
apoptosis (15,18,19).
In addition, another pathway associated with apoptosis is the
extrinsic pathway, which is associated with death receptors,
including Fas. Adaptor molecules are recruited to the receptors
following Fas ligand binding to the Fas death receptor, initiating
the program of apoptosis (16,20,21).
Previous studies by our group revealed that a
trypsin inhibitor from buckwheat was able to markedly inhibit the
proliferation of the IM-9 and K562 cell lines in vitro
(22,23). In order to elucidate whether the
recombinant buckwheat trypsin inhibitor (rBTI) has the same effect
in vivo and which apoptotic pathway is activated following
rBTI treatment, the effect of rBTI treatment on the proliferation
of H22 hepatic carcinoma cells was investigated in vitro and
in vivo.
Materials and methods
Materials
The H22 hepatic carcinoma cell line and the 7702
normal liver cell line were obtained from the Third Hospital of
Shanxi Medical University (Taiyuan, China) and China Radiation
Defense and Preservation Institute (Taiyuan, China). For all
experiments, 4–6 week-old female BALB/c mice (50 mice; 18–22 g)
purchased from the Institute of Laboratory Animal Science, Chinese
Academy of Medical Sciences (Beijing, China) were used. All mice
were group-housed in plastic cages with stainless-steel grid tops
in a room under a 12-h light/dark cycle and fed with distilled
water and food. The general health status of the animals was
monitored daily. An abdominal injection of the H22 cells (0.2 ml
106 cells/ml with 0.1 ml normal saline) was administered
to the BALB/c mice. The cell culture medium was composed of
RPMI-1640 (Gibco-BRL, Invitrogen Life Technologies, Carlsbad, CA,
USA) supplemented with 10% fetal calf serum (Institute of
Hematology, Hangzhou, China), 100 U/ml streptomycin and penicillin
solution (100 U/ml) purchased from the Cell Culture Center of the
Institute of Basic Medical Sciences, Chinese Academy of Medical
Sciences. The cells were incubated at 37°C under humidified
conditions with 5% CO2. MTT was purchased from
Sigma-Aldrich (St. Louis, MO, USA). An Annexin V-fluorescein
isothiocyanate Apoptosis Detection kit was obtained from BD
Pharmingen (San Diego, CA, USA). An apoptosis DNA Ladder Detection
kit was purchased from Nanjing KeyGen Biotech Co., Ltd. (Nanjing,
China). A Cytochrome C Detection kit and Caspase-3, -8 and-9
Colorimetric Assay kits were obtained from BioVision (Mountain
View, CA, USA). The rBTI was prepared in our laboratory by cloning,
expression and one-step affinity purification as described
previously (22,23). All other chemicals used were of
analytical grade. The current study was approved by the Ethics
Committee of Shanxi University (Taiyuan, China).
Cell viability assay
The cell viability was assessed using an MTT assay.
The MTT assay is a colorimetric assay, which measures the
percentage of surviving cells. The H22 and 7702 cell lines were
separately transferred to quadruplicate wells of 96-well microtiter
plates at a density of 5×103 cells/well. The cells were
treated with rBTI at concentrations of (6.25, 12.5, 25 and 50
μg/ml for 12, 20 and 24 h, each group investigated in
triplicate. A total of 20 μl MTT (5.0 mg/ml) was added and
the cells were incubated for 4 h. Following this, 80 μl
dimethyl sulfoxide was added. The color intensity was measured
using a microtiter plate reader (Model 550; Bio-Rad Laboratories,
Inc., Hercules, CA, USA) at 570 nm. The absorbance of the untreated
cells was considered to be 100%. The 50% inhibitory concentration
was the concentration at which a 50% decrease in the optical
density of the drug-treated cells was induced, with respect to
untreated cells.
Morphological observation of nuclei
Following treatment with rBTI (50 μg/ml) for
24 h, the H22 cells were washed with phosphate-buffered saline
(PBS; pH 6.7; Sigma-Aldrich) and centrifuged (2,000 ×g), then
incubated with 4% paraformaldhyde (Sigma-Aldrich) for 10 min.
Subsequently, the cells were collected via centrifugation (2,000
×g). The cells were stained with 0.1 μg/ml DAPI
(Sigma-Aldrich) for 5 min. Fluorescence microscopy was then used to
observe the nuclear morphology.
DNA fragmentation analysis
Following treatment with various concentrations of
rBTI (12.5, 25 or 50 μg/ml), a total of 1×106 H22
cells were collected. The DNA of the H22 cells treated with rBTI
was extracted according to a procedure described in a previous
study by our group (23). The
extracted DNA was analyzed on a 1.0% agarose gel (Sigma-Aldrich)
with the GeneGenius Bio Imaging system (SynGene, Frederick, MD,
USA) and observed under ultraviolet light with an FV1000
fluorescent microscope (Olympus, Tokyo, Japan).
Flow cytometric analysis of cell
apoptosis
Following treatment with rBTI (12.5, 25 or 50
μg/ml) for 24 h, the cells were adjusted to 1×106
in 50 mM PBS at pH 7.6 and centrifuged at 2,000 ×g. A total of 100
μl binding buffer was then added to re-suspend the cells.
Annexin V and propidium iodide were added to the cell suspension
followed by a 20-min incubation, and the cells were then analyzed
using flow cytometry using an FC500 Flow Cytometer (BD Biosciences,
Franklin Lakes NJ, USA) (9).
Measurement of cytochrome C in
mitochondria and cytoplasm
Extraction of cytochrome C. Following treatment with
rBTI (25 or 50 μg/ml) for 24 h, 1×106 H22 cells
were washed with cold PBS. The cells were then centrifuged at 600
×g for 5 min at 4°C. The supernatant was then removed and the cells
were re-suspended in 1.0 ml cytosol extraction buffer. The samples
were incubated on ice for 10 min. The homogenate was then
transferred to a 1.5-ml microcentrifuge tube and centrifuged at 700
×g for 10 min at 4°C. The supernatant was then collected into a
fresh 1.5-ml tube and centrifuged at 10,000 ×g for 30 min at 4°C.
The supernatant was then collected as the cytosolic fraction. The
pellet was re-suspended in 0.1 ml mitochondrial extraction buffer
mix with protease inhibitors, vortexed for 10 sec and saved as the
mitochondrial fraction. After extracting cytochrome C from the
mitochondria and cytoplasm, cytochrome C was electro-phoresed using
15% SDS-PAGE.
Western blot analysis
The protein concentration was determined using a BCA
Protein Assay kit (Lanbao Biotechnology, Shanghai, China) according
to according to the manufacturer’s protocol. The protein was
electrophoresed using 15% SDS-PAGE (as described above) and
transferred onto nitrocellulose membranes (Beyotime Institute of
Biotechnology, Shanghai, China). Following blocking, the membranes
were incubated for 4 h at 37°C with the primary antibody, which was
goat anti-mouse monoclonal anti-cytochrome C (1:400; IM 94638;
Beyotime Institute of Biotechnology), and the secondary antibody
(1:5,000; 572909; Beyotime Institute of Biotechnology), which was
goat anti-rat alkaline phosphatase-conjugated immunoglobulin E. The
blots were visualized using enhanced chemiluminescence detection
reagents (Beyotime Institute of Biotechnology) according to the
manufacturer’s instructions. The SynGene GeneTools analysis
software-version 3.02.00 (SynGene, Frederick, MD, USA) was used to
analyze the images and perform calculations.
Caspase colorimetric activation
assessment
The cells, which were treated with 50 μg/ml
rBTI for 24 h, were collected and prepared via incubation with
extraction buffer for 30 min and centrifugation at 15,000 ×g for 20
min. Subsequently, the supernatant was collected and the protein
concentration was assessed. The protein concentration was
determined using a bicinchoninic acid protein assay kit (Beyotime
Institute of Biotechnology). The extracts were incubated in a
96-well microtitre plate with three types p-nitroanilide
(pNA): Asp-Glu-Val-Asp-pNA for caspase-3, Ile-Glu-Thr-Asp-pNA for
caspase-8 and Leu-Glu-His-Asp-pNA for caspase-9, respectively, for
2 h at 37°C (BioVision). The absorbance was then measured at 405
nm.
Animal experiment
The anti-tumoral activity of rBTI against ascites
development was evaluated in mice. Following the development of
ascites, the peritoneal cavity was washed with 2 ml PBS following
treatment with 2.5 mg/100 g pentobarbital (Westang Biotechnology,
Shanghai, China). The mice were assigned randomly into five groups:
The rBTI treatment groups (four groups) and the control group (n=10
per group). The drug (0.125, 0.25, 0.50 or 1.0 mg/kg; 0.2 ml rBTI)
was administered via abdominal injection daily for eight days. The
ascites volume was calculated using a burette and the cumulative
ascites volume was calculated by adding these values. The total
number of viable H22 cells present in the ascites fluid (peritoneal
wash) was counted using a hemocytometer.
Statistical analysis
SPSS 12.0 for Windows (SPSS, Inc., Chicago, IL, USA)
was used for statistical analysis. All values are expressed as the
mean ± standard error. Comparisons within groups were performed
using a one-way analysis of variance and differences between groups
were determined using Scheffé’s method. P<0.05 was considered to
indicate a statistically significant difference.
Results
rBTI inhibits H22 cell proliferation in a
dose- and time-dependent manner
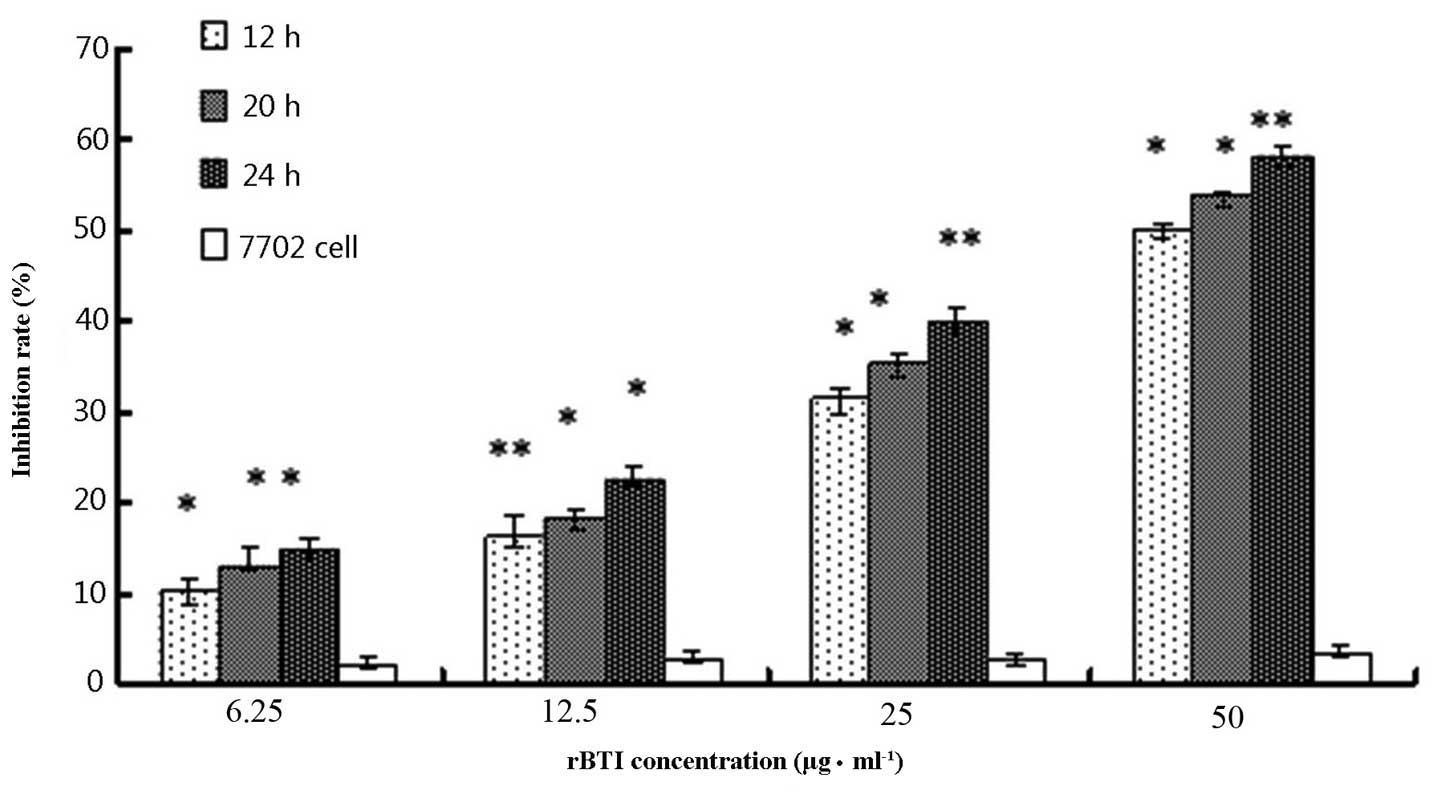
The cell growth inhibitory activity of rBTI was
assessed using a colorimetric MTT assay, as shown in Fig. 1. The growth inhibition rate of rBTI
was 17.8, 27.3, 43.6 and 62.7% at concentrations of 6.25, 12.5, 25
or 50 μg/ml, respectively. The inhibitory effect of rBTI on
the proliferation of H22 cells markedly increased as the
concentration of rBTI increased. Similarly, the inhibition rate was
enhanced with increasing incubation time with the drug. The
inhibitory effect of rBTI on the proliferation of H22 cells
occurred in a dose-and time-dependent manner; however, there were
minimal effects on the 7702 normal liver cell line.
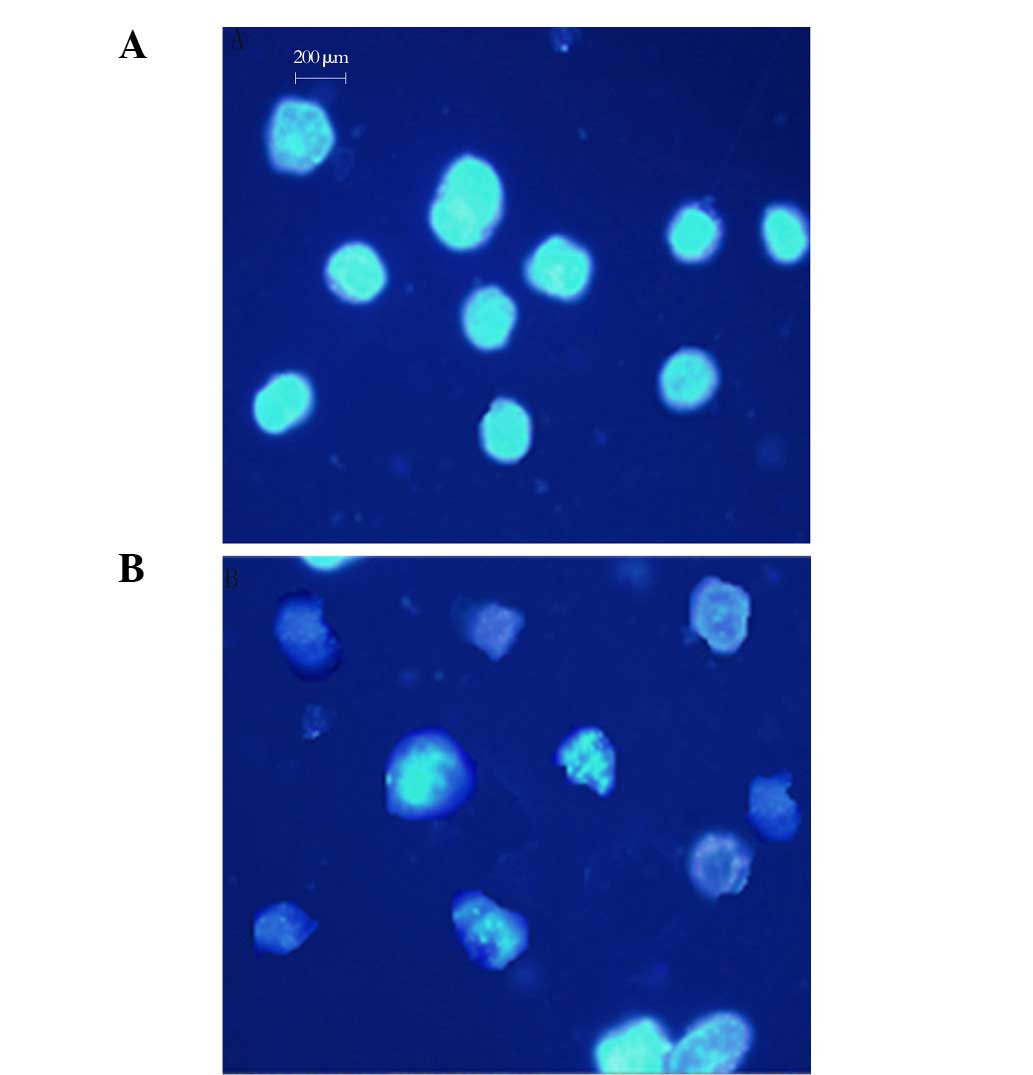
Nuclear morphological observation
The apoptotic induction effect of rBTI on H22 cells
was also investigated. As shown in Fig. 2, following 24 h of 50 μg/ml
rBTI treatment, there were significant changes to the cell
morphology of H22 cells compared with that of the control cells.
The typical characteristics of apoptosis, including membrane
disintegration and apoptotic body formation, were also observed
(Fig. 2B).
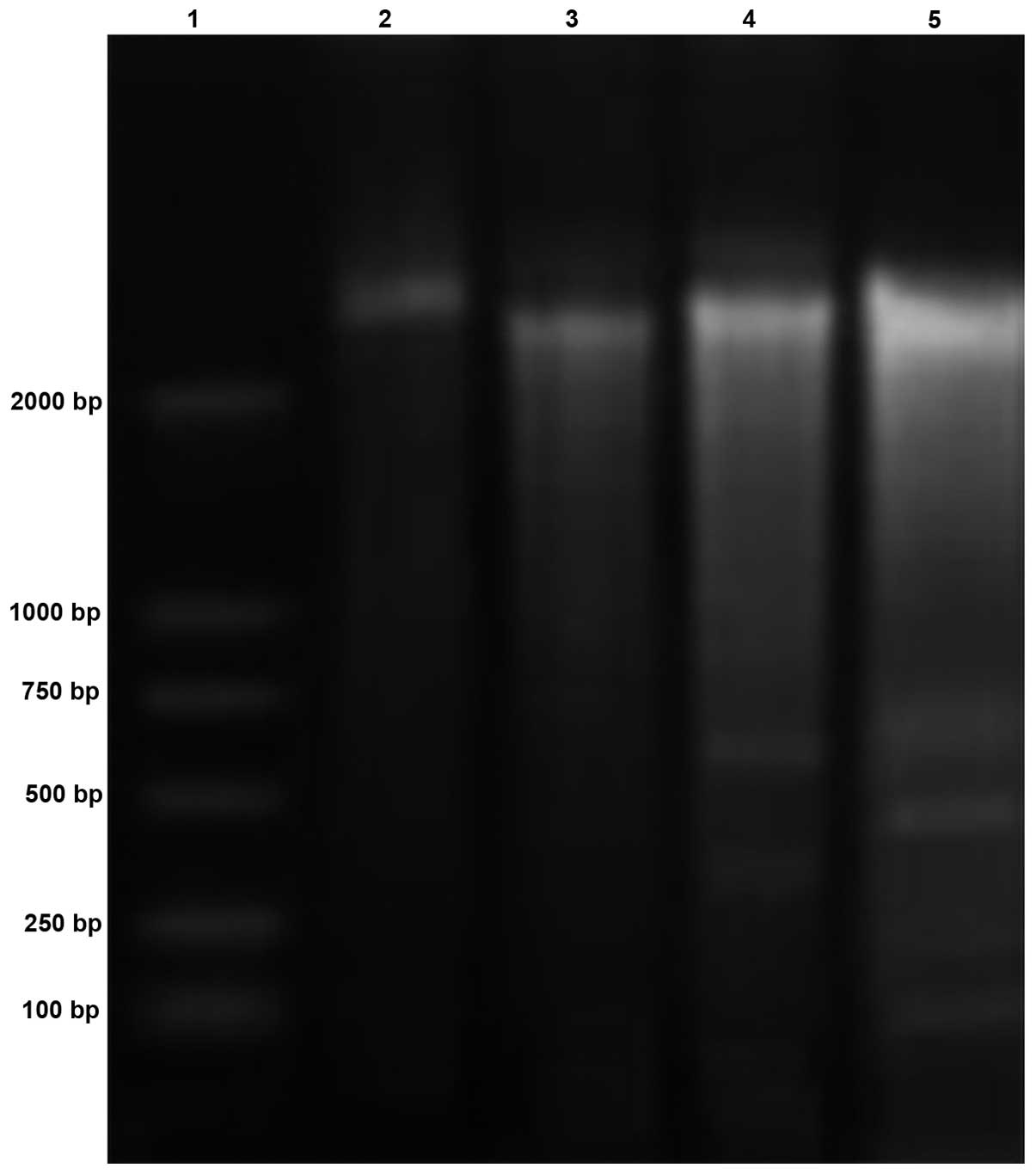
DNA electrophoresis band analysis
The results of the band analysis following DNA
electrophoresis revealed that 200-base pair fragments were present
following treatment with 50 μg/ml rBTI (Fig. 3). The bands of the DNA
electrophoresis presented regularly, similar to a ladder. The band
pattern exhibited typical features of apototic cells.
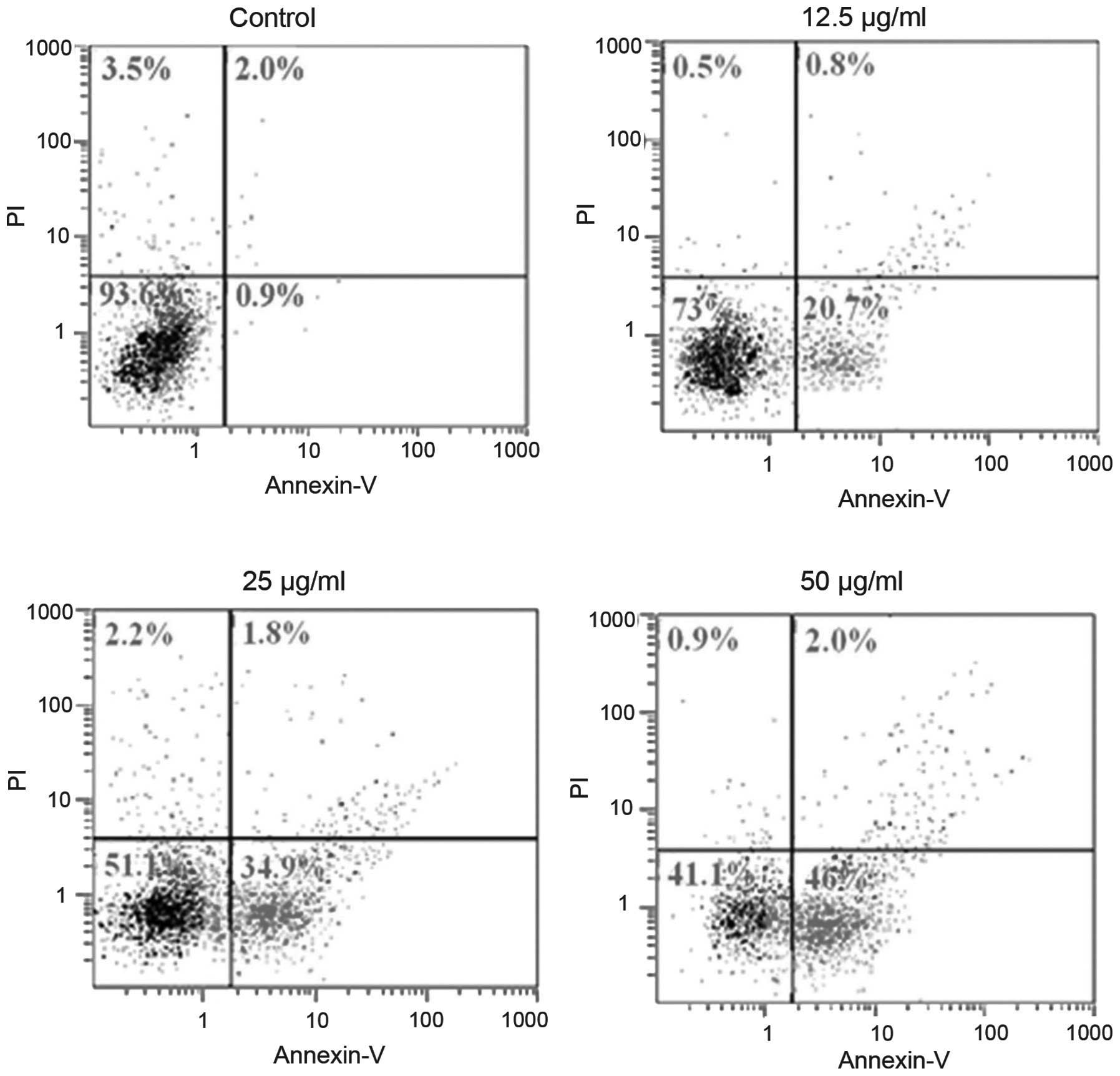
Flow cytometric analysis
Cell apoptosis in the H22 cell line was determined
by flow cytometry with or without treatment with rBTI at various
concentrations (12.5, 25 or 50 μg/ml) for 24 h (Fig. 4). The results revealed that the
ratio of apoptotic cells increased with the increase in rBTI
concentration. Significant differences were observed among each
treatment group and the control.
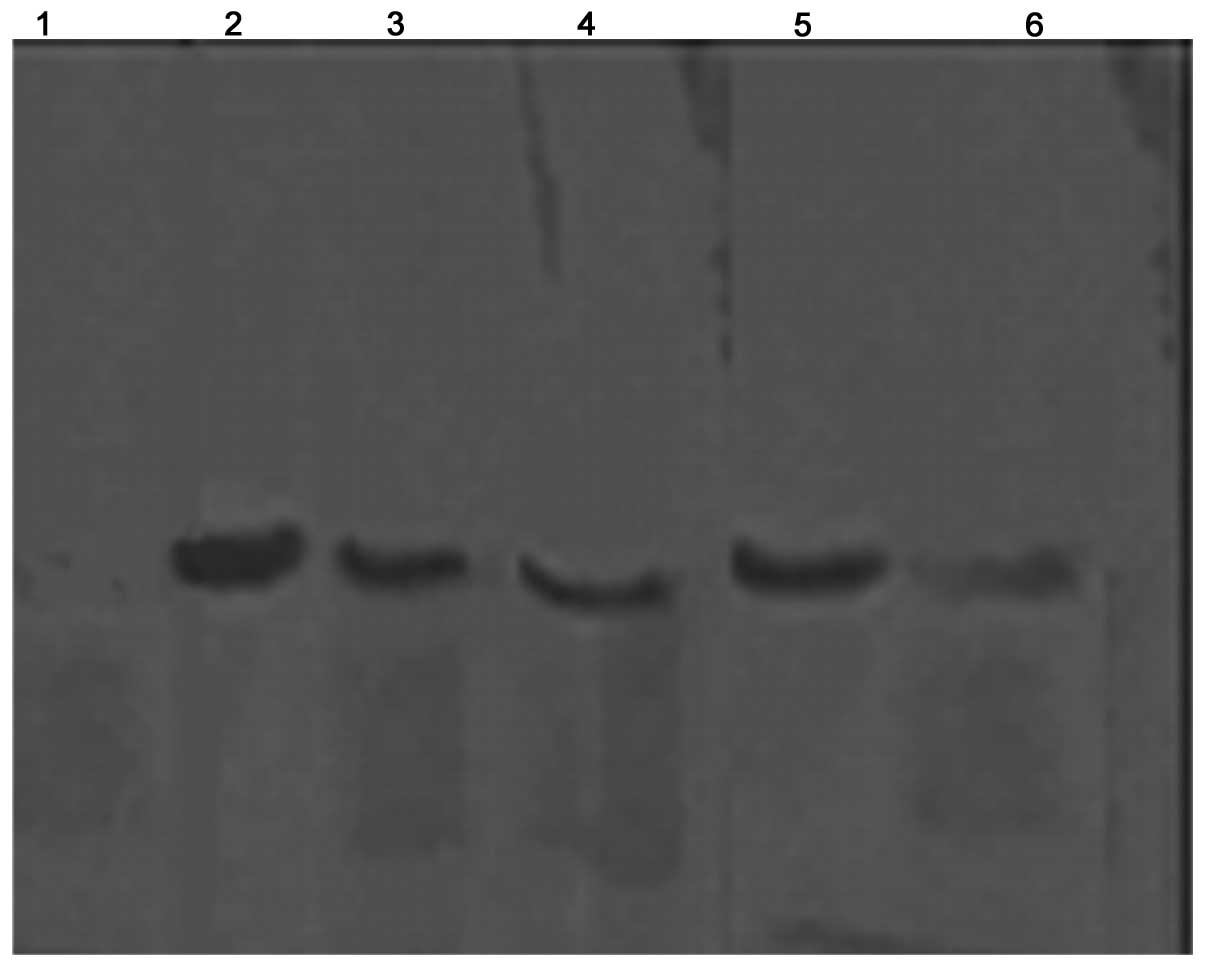
Western blot analysis
Cytochrome C is important in cell apoptosis. It is
located in the inter-membrane space between the two membranes.
Cytochrome C is released from the mitochondria to the cytosol when
the cell undergoes apoptosis. Fig.
5 demonstrates that cytochrome C was released from the
mitochondria into the cytoplasm following treatment with rBTI for
24 h and following an increase in the concentration of rBTI from 25
to 50 μg/ml. The quantities of cytochrome C in the cytosol
were 54.1 and 74.6% of total cytochrome C, respectively.
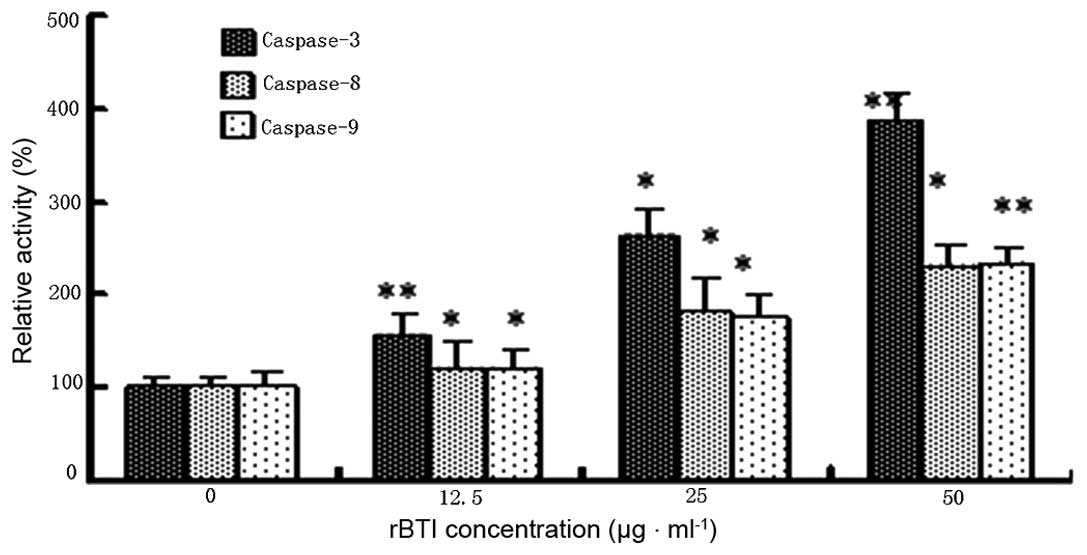
Caspase colorimetric activation
assessment
As shown in Fig. 6,
rBTI-induced apoptosis of H22 cells was predominantly associated
with the activities of caspase-3 in a concentration-dependent
manner. In addition, caspase-8 and caspase-9 were slightly
activated by rBTI. The results supported the hypothesis that rBTI
predominantly induced apoptosis in the H22 cell line via the
mitochondrial-mediated intrinsic apoptotic pathway involving
caspase-3 and caspase-9.
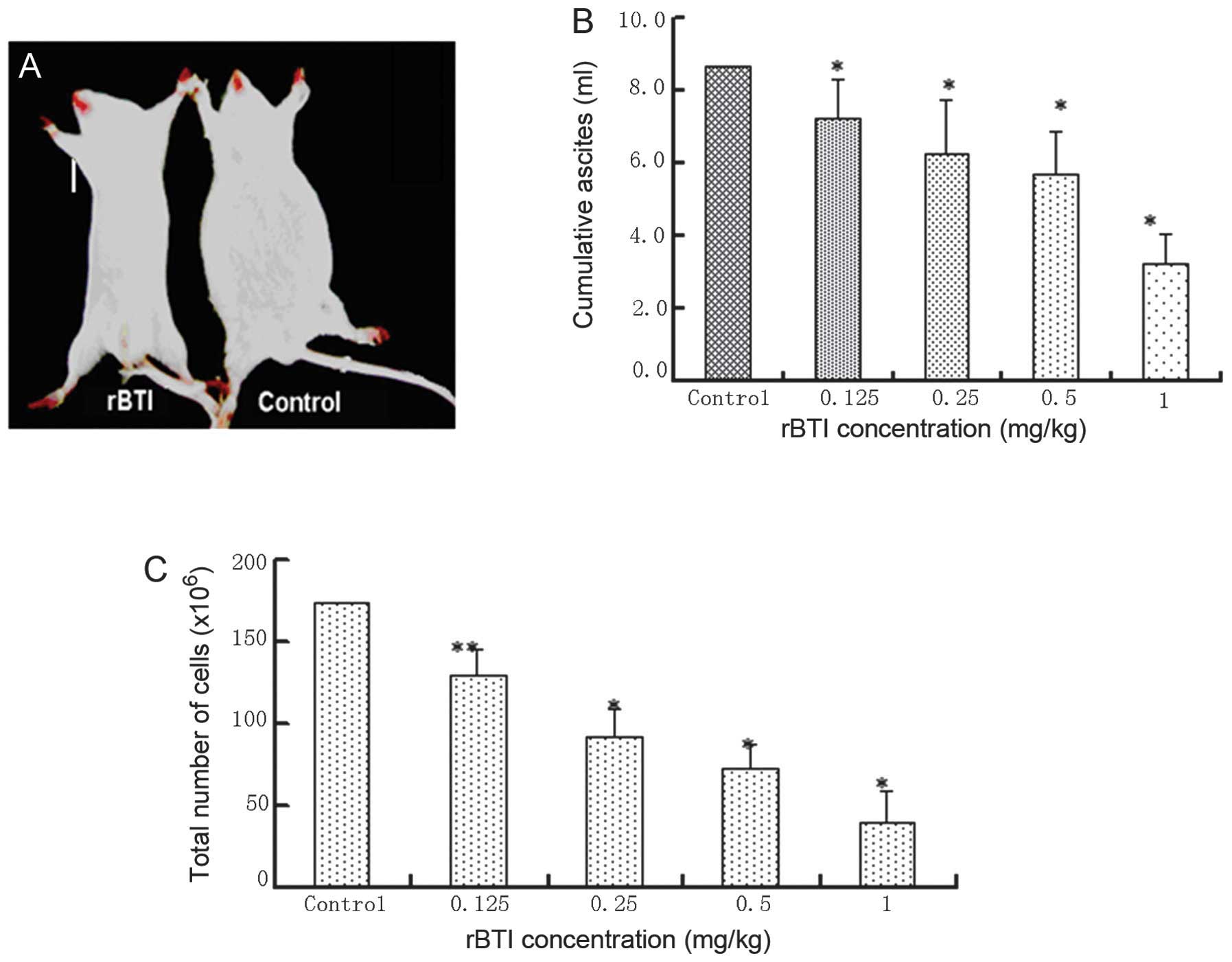
Inhibition of ascites production in mice
following treatment with rBTI
With the development of ascites generation, the
abdominal content increased and mice (7 mice with no treatment, 4
mice treated with 0.125 mg/kg, 4 mice treated with 0.25 mg/kg, 3
mice treated with 0.5 mg/kg and 1 mouse treated with 1 mg/kg) died
throughout the course of the treatment (8 days). By contrast, mice
treated with rBTI at various doses (0.125, 0.25, 0.5 and 1 mg/kg)
exhibited a reduction in visible ascites during the entire
treatment period of 8 days, particularly those treated with rBTI at
the highest concentration of 1 mg/kg (Fig. 7A). The cumulative volume of ascites
was evaluated (Fig. 7B). The
results revealed that ascites production was significantly
suppressed following treatment with rBTI. Similarly, rBTI treatment
led to a significant reduction in the quantity of intraperitoneal
tumor cells (Fig. 7C).
Discussion
A preliminary study by our group revealed the
efficient in vitro anti-tumoral effects of rBTI in K562
cells (23). In the present study,
the potent anti-tumoral activity of rBTI was demonstrated in
vitro and in vivo. The cell proliferation inhibition
rates of rBTI were 17.8, 27.3, 43.6 and 62.7% at concentrations of
6.25, 12.5, 25 or 50 μg/ml, respectively. The inhibitory
effect of rBTI on the proliferation of H22 cells occurred in a
dose-dependent and time-dependent manner. rBTI treatment induced
caspase activation and also resulted in the translocation of
cytochrome C from the mitochondria to the cytosol, as well as the
cleavage of DNA, but no effect on the proliferation of normal liver
cells was observed. This anti-tumoral activity was partly
consistent with the reported biological effects of other inhibitors
extracted from buckwheat seeds, including BWI-1 and BWI-2a, which
were able to inhibit T-acute lymphoblastic leukemia cell growth
in vitro (24). The caspase
family, which is comprised of aspartate-specific cysteine
proteases, is critical in the regulation of apoptosis. The key
biochemical pathways of caspase activation are well known (25). Caspase signaling is initiated and
propagated by proteolytic autocatalysis and the cleavage of
downstream caspases and substrates, including poly adenosine
diphosphate ribose polymerase and phospholipase C-γ1 (26). In particular, caspase-3 is one of
the key executioners of apoptosis, as it is either partially or
completely responsible for the proteolytic cleavage of a number of
key proteins (27). The vast
majority of cell death signals engage the mitochondrial pathway,
where the cysteine protease, caspase-9, is recruited and activated
(28). Activation of caspase-9 is
mediated by the formation of a macromolecular complex, termed the
apoptosome, with the release of cytochrome C from mitochondria
(29). In the present study, it
was first demonstrated that rBTI increases the release of
cytochrome C from the mitochondria. The release of cytochrome C
suggested that rBTI induced apoptosis through a mitochondrial
pathway (Fig. 5), which is
consistent with previous studies (18). In addition, caspase-3, -8 and -9
were activated, which are associated with the mitochondrial
intrinsic apoptotic pathway. However, the underlying mechanisms of
the induction of mitochondrial dysfunction following treatment with
rBTI remain to be elucidated. rBTI may inhibit the synthesis of
proteins, which maintain the mitochondrial membrane permeability as
a protease inhibitor (30). In
addition, it was identified that trypsin certain types of
transmembrane protein have a high homology at the DNA level;
therefore, rBTI possibly combines with the transmembrane protein
and enters the cell, where it induces mitochondrial
dysfunction.
The results of the present study also confirmed that
rBTI is able to significantly suppress the ascites production in
mice. The accumulation of malignant ascites is an important cause
of cancer-associated morbidity and mortality in patients with
peritoneal metastases (31). In
the present study, it was revealed that rBTI was able to
significantly suppress ascites formation in H22 tumor-bearing mice.
The results clearly demonstrated an effect on the inhibition of
proliferation in vitro and in vivo of rBTI. However,
the mechanisms associated with tumor growth inhibition require
further investigation.
In conclusion, rBTI, a novel trypsin inhibitor, was
shown to exhibit anti-tumoral activity in vitro and in
vivo. rBTI exerted its effects in a dose- and time-dependent
manner, while it only had minimal effects on the normal liver cell
line 7702. rBTI induced apoptosis by promoting mitochondrial
dysfunction, thereby leading to caspase activation. The present
study revealed a novel function of rBTI and supported its potential
application in treating malignant ascites. The development of a
trypsin inhibitor as an anti-tumoral agent requires further
investigation.
Acknowledgments
The present study was supported by grants from the
Natural Science Foundation of China (grant nos. 31171659 and
81173574) and Shanxi Provincial Experimental Animal Special Funds
(grant no. 2011K04). The authors gratefully acknowledge the kind
discussions and cooperation of the Central Laboratory of Chinese
Medicine Hospital of Shanxi Province. The authors would also like
to thank all the staff of the Institute of Biotechnology (Taiyuan,
China) for their helpful technical assistance.
References
|
1
|
Li CH, Kobayasshi K, Yoshida Y and Ohsawa
R: Genetic analyses of agronomic traits in Tartary buckwheat
(Fagopyrum tataricum (L.) Gaertn.). Breed Sci. 62:303–309. 2012.
View Article : Google Scholar
|
|
2
|
Pomeranz Y and Robbins GS: Amino acid
composition of buckwheat protein. J Agric Food Chem. 20:270–274.
1972. View Article : Google Scholar
|
|
3
|
Ikeda K and Kishida M: Digestibility of
proteins in buckwheat seed. Fagopyrum. 13:21–24. 1993.
|
|
4
|
Zhang Z, Li YY, Shi XR, et al: Gene
cloning and sequencing of common buckwheat trypsin inhibitor. Acta
Botanica Boreali-Occidentalia Sinica. 25:46–51. 2005.
|
|
5
|
Htay HH, Tsubouchi R, Haneda M, Murakami K
and Yoshino M: Induction of apoptosis of HL60 cells by gallic acid
derivatives. Biomed Res. 23:127–134. 2002. View Article : Google Scholar
|
|
6
|
Gardner PR: Aconitase: Sensitive target
and measure of superoxide. Methods Enzymol. 349:9–23.
2002.PubMed/NCBI
|
|
7
|
Gladysheva IP, Moroz NA, Karmakova TA,
Nemtsova ER, Yakubovskaya RI and Larionova NI: Immunoconjugates of
soybean Bowman-Birk protease inhibitor as targeted antitumor
polymeric agents. J Drug Target. 9:303–316. 2001. View Article : Google Scholar
|
|
8
|
Marschütz MK and Bernkop-Schnürch A: Oral
peptide drug delivery: Polymer-inhibitor conjugates protecting
insulin from enzymatic degradation in vitro. Biomaterials.
21:1499–1507. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Malkowicz SB, Liu SP, Broderick GA, Wein
AJ, Kennedy AR and Levin RM: Effect of the Bowman-Birk inhibitor (a
soy protein) on in vitro bladder neck/urethral and penile corporal
smooth muscle activity. Neurourol Urodyn. 22:54–57. 2003.
View Article : Google Scholar
|
|
10
|
Supuran CT, Casini A and Scozzafava A:
Protease inhibitors of the sulfonamide type: Anticancer,
antiinflammatory, and antiviral agents. Med Res Rev. 23:535–558.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Birk Y: The Bowman-Birk inhibitor.
Trypsin- and chymotrypsin-inhibitor from soybeans. Int J Pept
Protein Res. 25:113–131. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yavelow J, Finlay TH, Kennedy AR and Troll
W: Bowman-Birk soybean protease inhibitor as an anticarcinogen.
Cancer Res. 43(Suppl): 2454s–2459s. 1983.PubMed/NCBI
|
|
13
|
Cryns V and Yuan J: Proteases to die for.
Genes Dev. 12:1551–1570. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hu W and Kavanagh JJ: Anticancer therapy
targeting the apoptotic pathway. Lancet Oncol. 4:721–729. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dias N and Bailly C: Drugs targeting
mitochondrial functions to control tumor cell growth. Biochem
Pharmacol. 70:1–12. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Schulze-Osthoff K, Ferrari D, Los M,
Wesselborg S and Peter ME: Apoptosis signaling by death receptors.
Eur J Biochem. 254:439–459. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Rao RV, Ellerby HM and Bredesen DE:
Coupling endoplasmic reticulum stress to the cell death program.
Cell Death Differ. 11:372–380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Budihardjo I, Oliver H, Lutter M, Luo X
and Wang X: Biochemical pathways of caspase activation during
apoptosis. Annu Rev Cell Dev Biol. 15:269–290. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu X, Kim CN, Yang J, Jemmerson R and
Wang X: Induction of apoptotic program in cell-free extracts:
Requirement for dATP and cytochrome c. Cell. 86:147–157. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thorburn A: Death receptor-induced cell
killing. Cell Signal. 16:139–144. 2004. View Article : Google Scholar
|
|
21
|
Lacour S, Micheau O, Hammann A, Drouineaud
V, Tschopp J, Solary E and Dimanche-Boitrel MT: Chemotherapy
enhances TNF-related apoptosis-inducing ligand DISC assembly in
HT29 human colon cancer cells. Oncogene. 22:1807–1816. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang Z, Li Y, Li C, Yuan J and Wang Z:
Expression of a buckwheat trypsin inhibitor gene in Escherichia
coli and its effect on multiple myeloma IM-9 cell proliferation.
Acta Biochim Biophys Sin (Shanghai). 39:701–707. 2007. View Article : Google Scholar
|
|
23
|
Wang ZH, Gao L, Li YY, Zhang Z, Yuan JM,
Wang HW, Zhang L and Zhu L: Induction of apoptosis by buckwheat
trypsin inhibitor in chronic myeloid leukemia K562 cells. Biol
Pharm Bull. 30:783–786. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park SS and Ohba H: Suppressive activity
of protease inhibitors from buckwheat seeds against human T-acute
lymphoblastic leukemia cell lines. Appl Biochem Biotechnol.
117:65–74. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Stennicke HR and Salvesen GS: Properties
of the caspases. Biochim Biophys Acta. 1387:17–31. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park JW, Choi YJ, Suh SI, Baek WK, Suh MH,
Jin IN, Min DS, Woo JH, Chang JS, Passaniti A, et al: Bcl-2
overexpression attenuates resveratrol-induced apoptosis in U937
cells by inhibition of caspase-3 activity. Carcinogenesis.
22:1633–1639. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kaufmann SH and Hengartner MO: Programmed
cell death: Alive and well in the new millennium. Trends Cell Biol.
11:526–534. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li P, Nijhawan D, Budihardjo I,
Srinivasula SM, Ahmad M, Alnemri ES and Wang X: Cytochrome c and
dATP-dependent formation of Apaf-1/caspase-9 complex initiates an
apoptotic protease cascade. Cell. 91:479–489. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Narayanan S, Surolia A and Karande AA:
Ribosome-inactivating protein and apoptosis: Abrin causes cell
death via mitochondrial pathway in Jurkat cells. Biochem J.
377:233–240. 2004. View Article : Google Scholar
|
|
31
|
Smith EM and Jayson GC: The current and
future management of malignant ascites. Clin Oncol (R Coll Radiol).
15:59–72. 2003. View Article : Google Scholar
|