Introduction
Lung cancer, characterized by high incidence and
mortality rates, is a malignant tumor, which threatens human
health. It is reported that the incidence and mortality rates have
increased significantly in several countries in the last 50 years
(1,2). The etiology of lung cancer remains to
be fully elucidated, and there is substantial data showing a close
association between long-term heavy smoking and the occurrence of
lung cancer (2). Non-small cell
lung cancer (NSCLC) accounts for ~80% of all lung cancer cases, and
~75% of patients are diagnosed in the middle-late stage (3,4).
Therapies for NSCLC include surgery, radiation and chemotherapy;
however, the 5-year survival rate remains low. In addition, the
side effects and pain from treatment cause substantial physical and
mental trauma to patients. In previous years, the screening of
compounds from medicinal plants for the treatment of various
diseases has received increased attention. For example, the
anticancer drug, paclitaxel (Taxol), obtained from Taxus
species has been extensively investigated for several years
(5). Therefore, obtaining
anticancer compounds from medicinal plants is important in cancer
treatment.
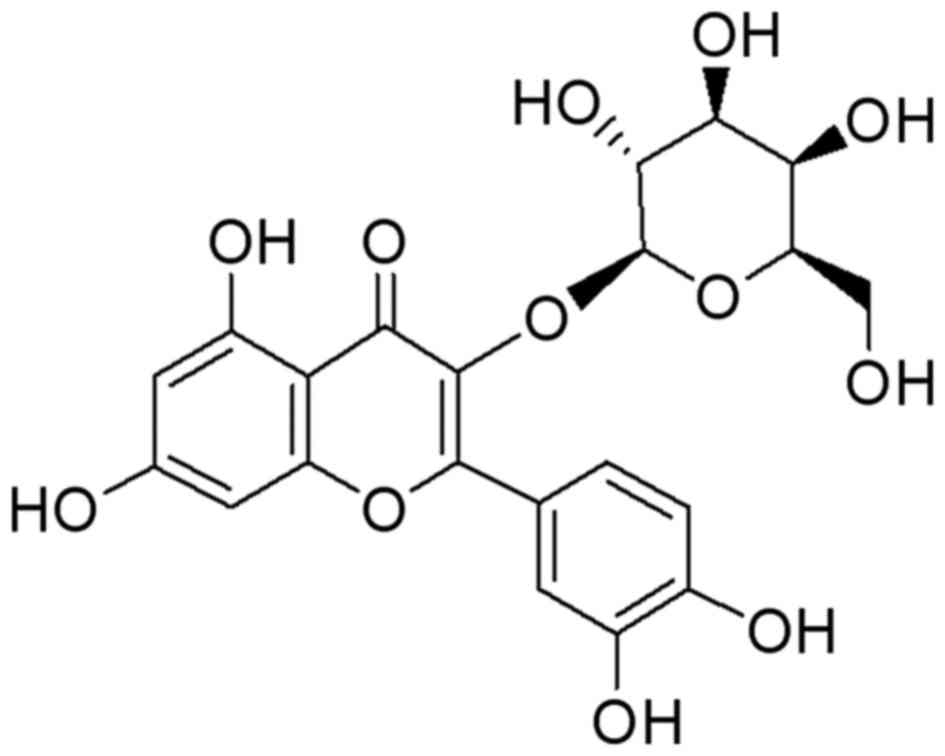
Hyperoside (HY; Fig.
1) is a major pharmacologically active component from
Prunella vulgaris L. and Hypericum perforatum, which
exerts a wide variety of biological activities, including
antioxidant, antihyperglycemic, anticancer, anti-inflammatory and
cardiovascular protective effects (6–9).
However, the underlying molecular mechanisms remain to be fully
elucidated. The present study aimed to determine the anticancer
effect of HY and identifying the possible mechanism involved. In
the present study, HY was identified as an anticancer agent, which
induced the apoptosis of human NSCLC A549 cells though the
mitochondria apoptotic pathway.
Materials and methods
Cell culture
A549 cells were obtained from the Shanghai Cell
Bank, Chinese Academy of Sciences (Shanghai, China). The cells were
cultured in DMEM (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Invitrogen; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 µg/ml streptomycin, and incubated in a humidified
atmosphere at 37°C with 5% CO2.
HY preparation
HY with a purity of 98.78% was obtained as a canary
yellow needle-shaped crystal (YUANYE Technological Co. Ltd.,
Shanghai, China). It was dissolved in an appropriate volume of
dimethylsulfoxide (DMSO) and diluted to the desired concentrations
(10, 50 and 100 µM) prior to utilization, with the final
concentration of DMSO maintained below 0.5%.
Cell viability
A standard tetrazolium bromide (MTT) assay was used
to assess cell viability. Briefly, the cells (5×103
cells/well) were seeded in 96-well plates. Following treatment with
HY (0, 10, 50 and 100 µM) for 24 h at 37°C, 50 ml of MTT
(Sigma-Aldrich; Merck Millipore, Darmstadt, Germany) solution (2
mg/ml in PBS) was added to each well and the plates were incubated
for an additional 4 h at 37°C. The medium was then removed and the
cells were incubated with 200 µl DMSO in the dark for 30 min at
37°C to dissolve violet crystals. The absorbance was read at 570 nm
on an automatic microplate reader with DMSO as the blank control.
All assays were performed with five replicates and repeated at
least three times.
Cell apoptosis
In order to determine cell apoptosis,
5×104 cells were trypsinized, washed with cold
phosphate-buffered saline (PBS) and resuspended in binding buffer
according to the manufacturer's protocol of the Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
Detection kit (cat. no. KGA106; Nanjing KeyGen Biotech Co., Ltd.,
Nanjing, China). FITC-Annexin V and PI were added to the fixed
cells for 20 min in darkness at room temperature. Subsequently,
Annexin V binding buffer was added to the mixture prior to
quantifying the fluorescence using a FACSort flow cytometer (BD
Biosciences, Franklin Lakes, NJ, USA). Cell apoptosis was analyzed
using Cell Quest version 3.0 (BD Biosciences).
Mitochondrial membrane potential
(MMP)
Rhodamine-123 (Rho-123) dye (Sigma-Aldrich; Merck
Millipore) was used to detect alterations in MMP. The cells
(5×104 cells/well) were cultured in a 24-well plate.
Following treatment with HY (0, 10, 50 and 100 µM) for 24 h, the
cells were washed with PBS, incubated with Rho-123 (10 mg/ml) for
30 min at 37°C and subsequently subjected to flow cytometry.
Separation of mitochondrial and
cytosolic fractions
A mitochondria/cytosol fractionation kit (Abcam,
Cambridge, UK) was used to prepare the mitochondrial and cytosolic
fractions from the cells, in accordance with the manufacturer's
protocol. Following treatment for 12 h with the different
concentrations of HY, the cells were harvested, washed by PBS and
centrifuged at 600 × g for 5 min at 4°C. Following re-suspension
with cytosolic extraction buffer, the cells were homogenized on ice
for 40 passes with a grinder for 10 min. The homogenate was
subsequently centrifuged at 700 × g for 10 min at 4°C, supernatant
was carefully collected, and the sediment was saved. Following
centrifugation at 10,000 × g for 30 min at 4°C, the supernatant was
collected as the cytosolic fraction and stored at −80°C. The
sediment was re-suspended with mitochondrial extraction buffer,
vortexed and stored as the mitochondrial fraction at −80°C.
Western blot analysis
The cultured cells were harvested and washed twice
with PBS, and lysed in ice-cold radioimmunoprecipitation assay
buffer (Beyotime Institute of Biotechnology, Shanghai, China) with
freshly added 0.01% protease inhibitor cocktail (Sigma-Aldrich;
Merck Millipore) and incubated on ice for 30 min. The cell lysate
was centrifuged at 13,000 × g for 10 min at 4°C, following which
the supernatant was run on a 10% SDS-PAGE gel and transferred
electrophoretically onto a polyvinylidene fluoride membrane (EMD
Millipore, Billerica, MA, USA). The blots were blocked with 5% skim
milk, followed by incubation with antibodies against phosphorylated
(P)-c-Jun N-terminal kinase (P-JNK; cat. no. ab4821; 1:500; Abcam),
JNK (cat. no. ab154902; 1:200; Abcam), P-P38 (cat. no. ab47363;
1:500; Abcam), P38 (cat. no. ab170099; 1:1,000; Abcam), GAPDH (cat.
no. ab9485; 1:2,500; Abcam), apoptosis-inducing factor (AIF; cat.
no. ab1998; 1:500; Abcam), cytochrome c (cat. no. ab13575;
1:500; Abcam), caspase-3 (cat. no. ab2171; 1:500; Abcam) and
caspase-9 (cat. no. ab32539, 1:1,000; Abcam) at 4°C overnight. The
blots were then incubated with goat anti-mouse or anti-rabbit
secondary antibody (Beyotime Institute of Biotechnology, Shanghai,
China) at room temperature for 1 h and visualized using SuperSignal
West Dura Extended Duration substrate (Thermo Fisher Scientific,
Inc.) and detected using a DNR Bio-Imaging System (DNR Bio-Imaging
Systems, Ltd., Jerusalem, Israel). Protein density levels were
determined by Image Analysis software version 4.0.3.2 (Scion Co.,
Ltd., Frederick, MD, USA) and the densitometric analysis of blots
from three experiments.
Statistical analysis
All results are presented as the mean ± standard
deviation and the data were analyzed using the SPSS 13.0
statistical package (SPSS, Inc., Chicago, IL, USA). Data for
multiple comparisons were subjected to one-way analysis of variance
followed by Dunnett's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
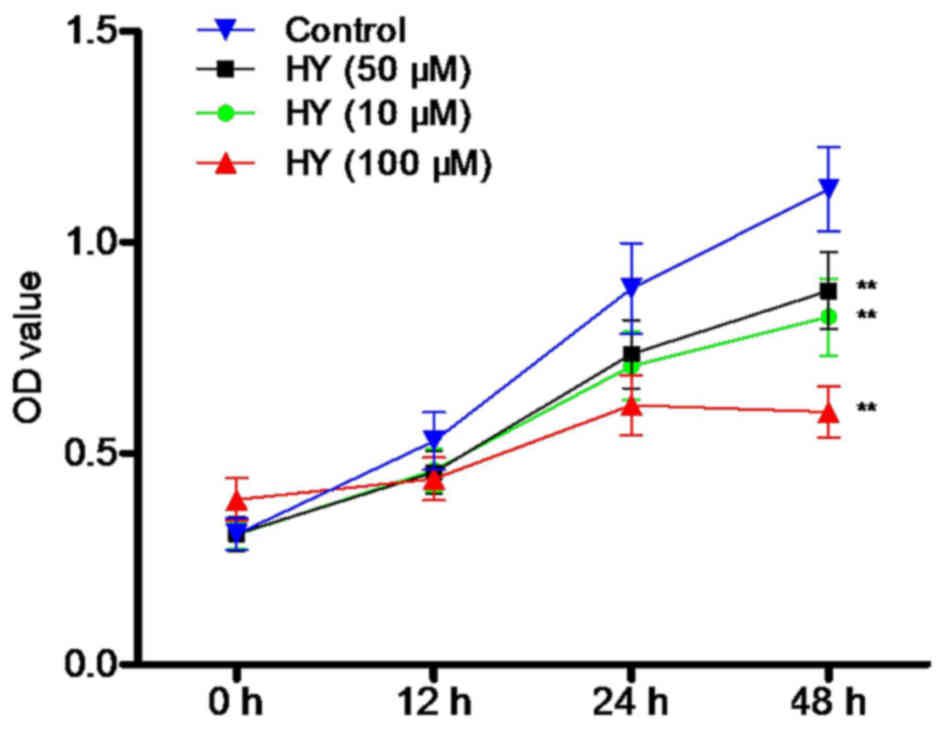
HY suppresses the viability of A549
cells
Following treatment of the cells with various
concentrations of HY (0, 10, 50 and 100 µM) for 12, 24 and 48 h,
cell viability was determined using an MTT assay. As shown in
Fig. 2, HY inhibited the viability
of the A549 cells in a time- and dose-dependent manner. Cell
proliferation was markedly decreased, compared with that in the
control group.
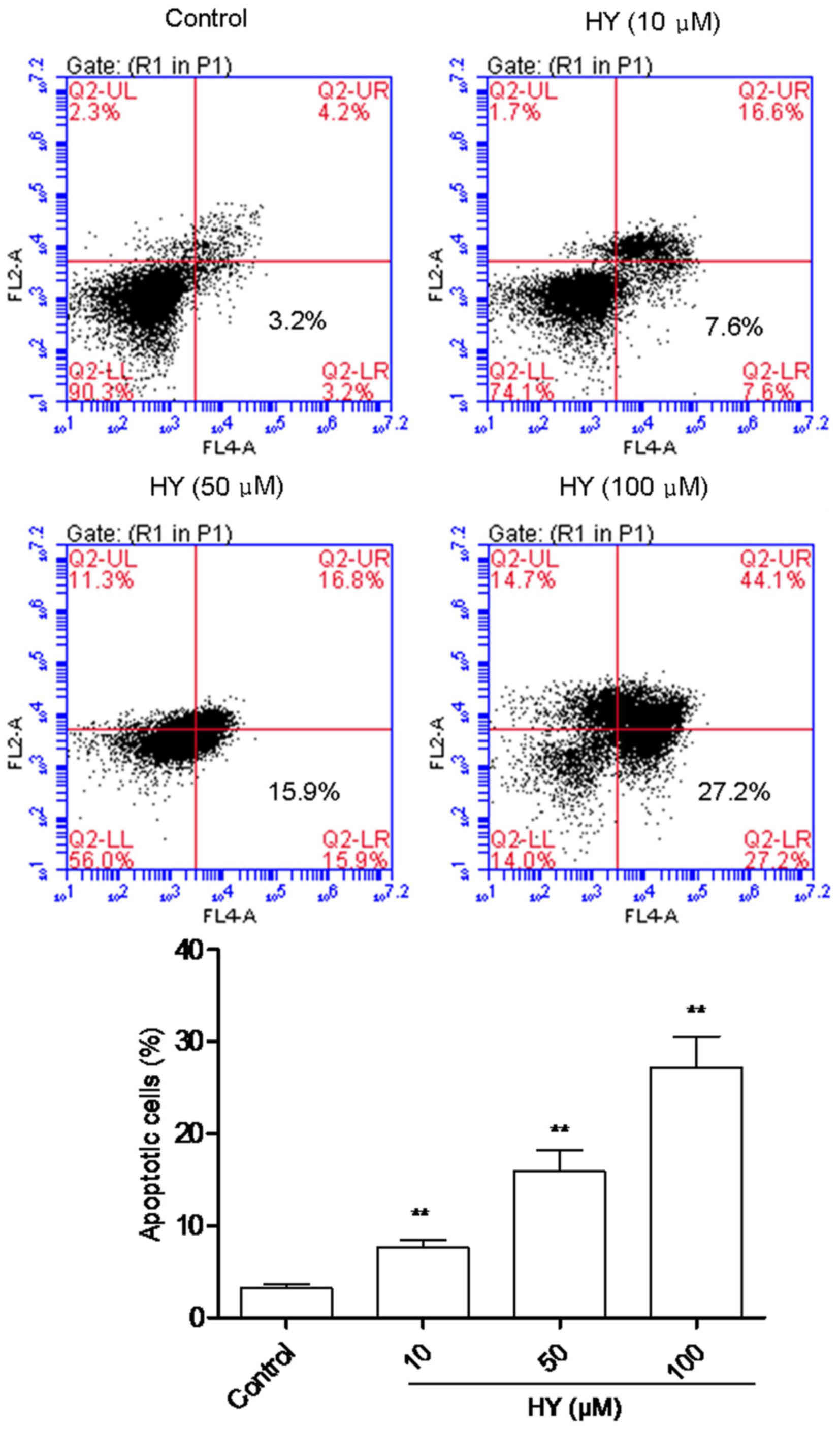
HY induces the apoptosis of cells
In order to quantify cellular apoptosis, a flow
cytometric assay was performed to analyze the proportion of
apoptotic cells dyed with PI and Annexin V, to enable evaluation of
the integrity of cell membrane and the externalization of
phosphatidylserine. As shown in Fig.
3, analysis of the cell population revealed distinct sets
within the population. The majority of cells were alive in the
control group (Fig. 3). However,
when the cells were incubated with various concentrations of HY for
24 h, there was a notable increase in the percentage of apoptotic
cells, and this occurred in a dose-dependent manner.
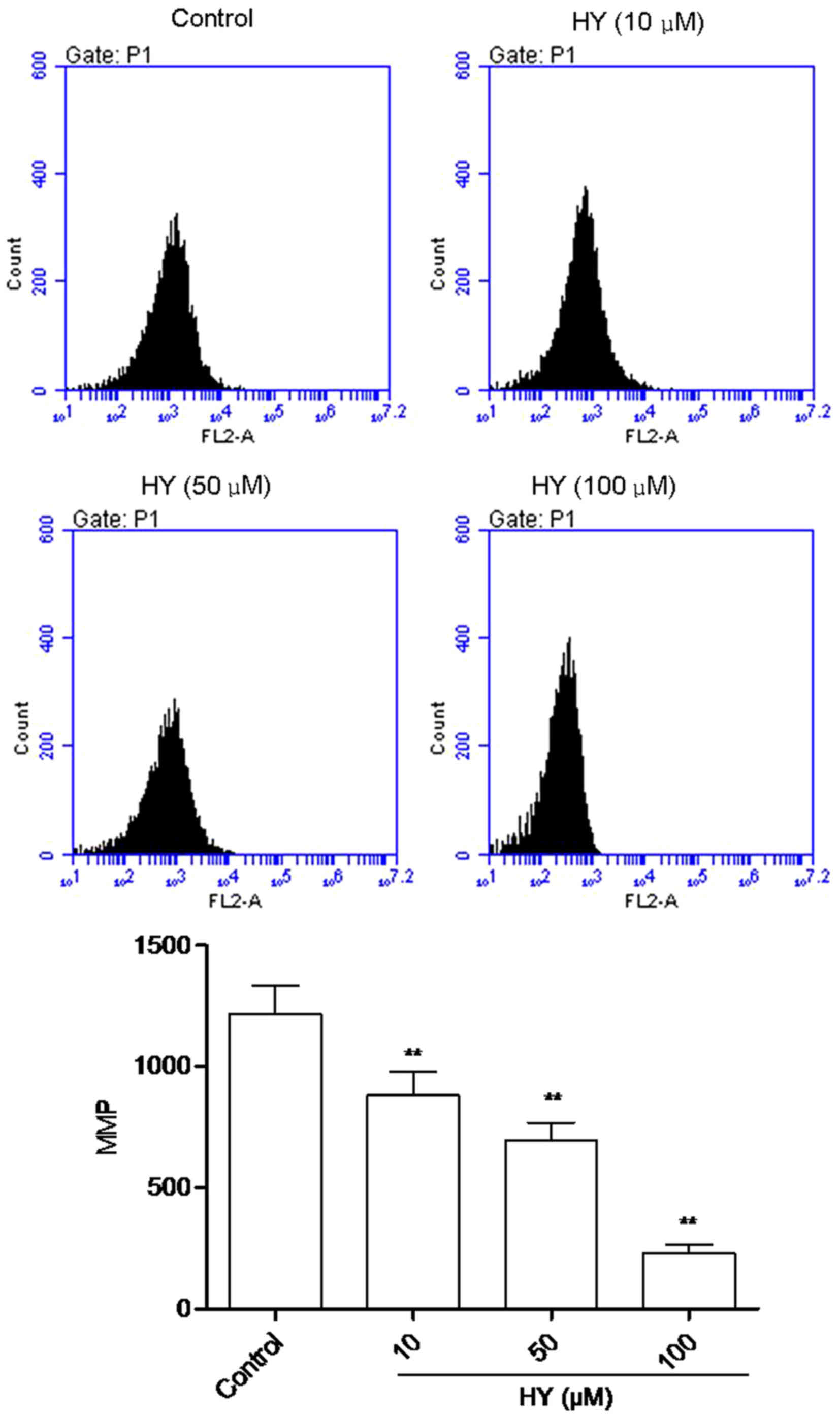
HY enhances MMP dissipation
The fluorescent intensity (FI) of Rho123 is
positively correlated with the MMP. As shown in Fig. 4, high intensity fluorescence was
visible in the control group of cells. Following HY treatment for
24 h, FI decreased in a dose-dependent manner.
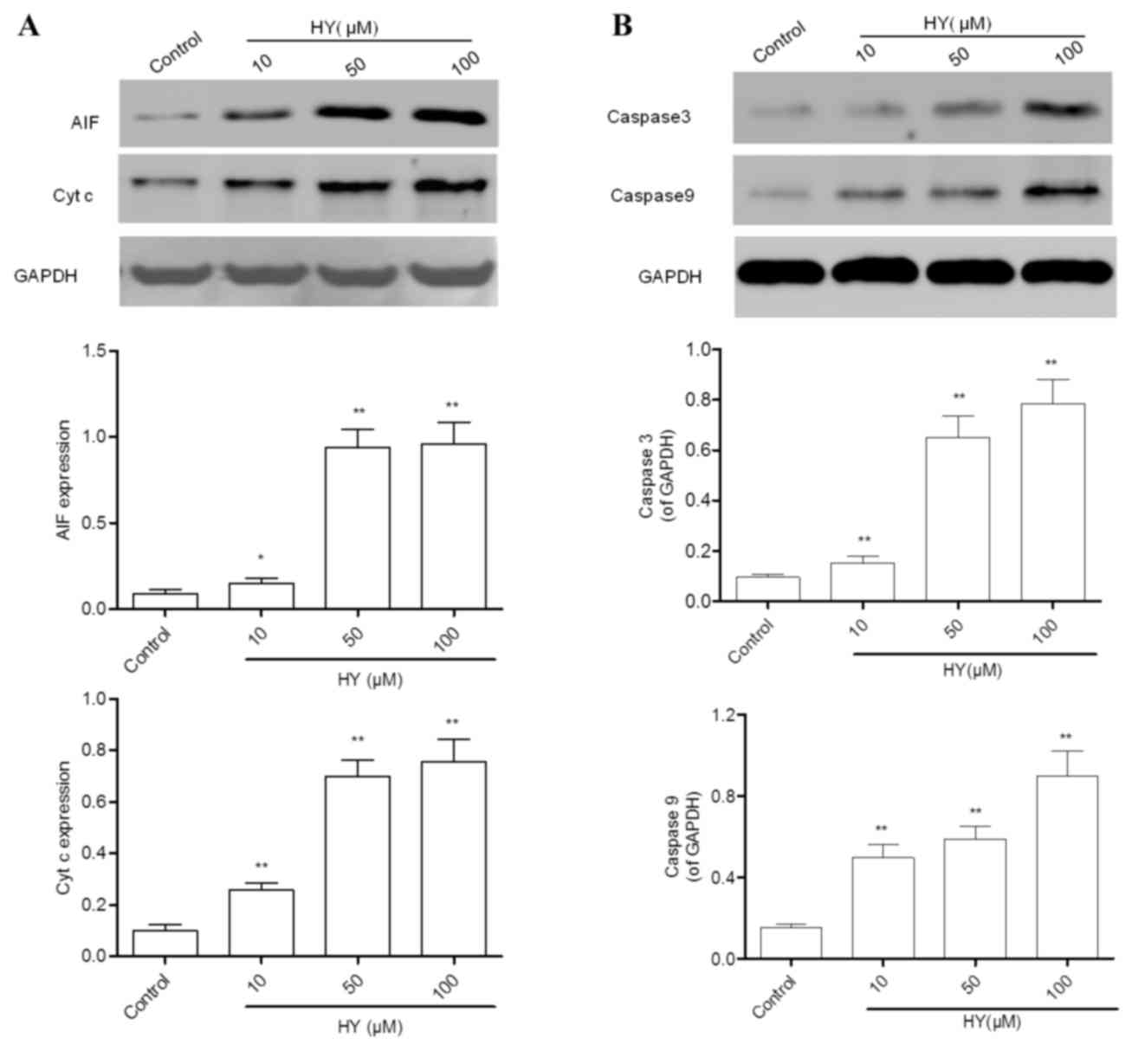
HY induces the release of cytochrome c
and AIF
Following MMP dissipation, the key event of
mitochondria-dependent apoptosis is the release of mitochondrial
cytochrome c and AIF into cytosol. Therefore, these two
parameters were evaluated using western blot analysis. The results
revealed that the expression levels of cytochrome c and AIF
(Fig. 5A) were lower in the
control group of cells, and were significantly elevated by HY
treatment for 24 h.
HY activates caspase-3 and
caspase-9
Caspase-3, a common downstream apoptosis effector,
can be activated by caspase-9 or caspase-8 and cleaved into the
active fragment. In the present study, the protein expression
levels of caspase-3 and caspase-9 were detected using western blot
analysis. The expression levels of caspase-3 and caspase-9 in the
control group were low (Fig. 5B).
However, treatment with HY for 12 h elevated their expression
levels in a dose-dependent manner.
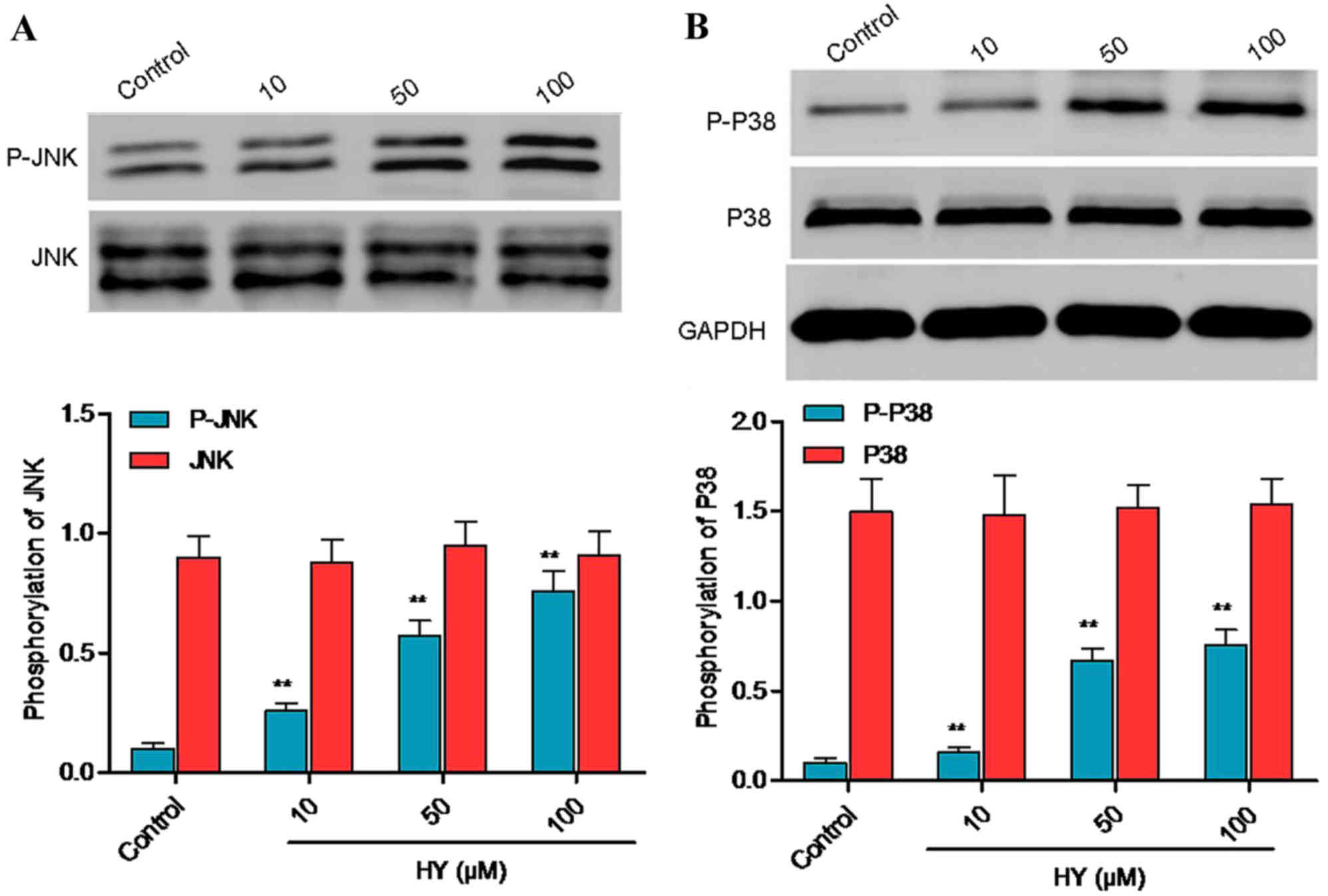
HY facilitates protein phosphorylation
of p38 mitogen-activated protein kinase (MAPK) and JNK
Cell apoptosis is closely associated with activation
of the JNK and p38 MAPK signaling pathways, which is characterized
by increased protein phosphorylation of p38 MAPK and/or JNK.
Following treatment with HY for 3 h, the protein phosphorylation
levels of p38 MAPK and JNK were analyzed using western blot
analysis (Fig. 6A and B).
Treatment with HY for 3 h increased the protein phosphorylation of
p38 MAPK and JNK, and there were significant differences between
the HY-treated groups and the control group.
Discussion
Higher plants produce an abundance of substances,
which have been a reliable source of pharmaceuticals. In the
present study, it was found that HY effectively inhibited the
proliferation of human NSCLC A549 cells in a dose- and
time-dependent manner. Furthermore, Annexin V/PI staining revealed
that treatment with HY (10, 50 and 100 µM) for 12 h increased the
percentage of apoptotic cells in a dose-dependent manner. These
results indicated that HY induced apoptosis of the A549 cells.
MAPK pathways are generally recognized as the signal
transducers of apoptosis in tumor cells, closely associated with
responses to extracellular stimuli, including small molecular
compounds (10–13). It is known that the MAPK system is
composed of extracellular signal-regulated kinases, JNK and p38 MAP
kinase. The JNK and p38 MAPK pathways can be activated by genotoxic
agents and by the cytokine-mediating stress response, inducing
growth inhibition and apoptosis in the cells (14). The induction of A549 cell apoptosis
by 2-methoxy-1,4-naphthoquinone (MNQ) is independent of cell cycle
arrest, and is mediated by the JNK and p38 MAPK signaling pathways.
These signaling pathways have been found to be stimulated by
oxidative DNA damage caused by the increased generation of reactive
oxygen species (ROS) in the MNQ-treated A549 cells (14). It has been reported that capsaicin
induces SMMC-7721 cell apoptosis through the generation of
intracellular ROS, dissipation of MMP, and activation of the JNK
and p38 MAPK pathways (15). Based
on the above evidence, it is clear that p38 MAPK and JNK pathways
are crucial in tumor cell apoptosis. In the present study, western
blot analysis was used to identify the phosphorylation of p38 MAPK
and JNK. The results revealed that HY significantly increased the
phosphorylation levels of p38 MAPK and JNK, indicating the
simultaneous activation of these two signaling pathways.
Activation of the intrinsic mitochondrial pathway
can elicit the release of several apoptotic factors, including
cytochrome c and AIF, which results in mitochondrial
apoptosis and activates the mitochondrial apoptotic pathway. The
release of cytochrome c facilitates the formation of the
apoptosome-containing adaptor, apoptotic protease-activating
factor-1, and another initiator, caspase-9, in the presence of
dATP, following which caspase-3 is activated leading to cell
apoptosis (16,17). AIF can elicit cell apoptosis
independently of the activation of caspase-3 (18). The enhanced protein expression of
activated caspase-3, a downstream effector caspase, indicated that
HY triggered caspase-dependent apoptosis, which can be activated by
the mitochondrial apoptotic pathway and/or the death receptor
pathway. The present study also found that HY facilitated the loss
of MMP, and the release of cytochrome c and AIF into the
cytosol, and enhanced the protein expression of caspase-9. This
indicated that HY induced apoptosis of the A549 cells via the
mitochondrial apoptotic pathway.
In conclusion, the present study demonstrated that
HY elevated the protein phosphorylation levels of p38 MAPK and JNK,
disrupted MMP, facilitated the release of cytochrome c and
AIF into cytoplasm, activated capase-9 and caspase-3, and led to
apoptosis of the A549 cells. As a result, HY may offer potential as
a useful and beneficial natural agent against lung cancer.
Acknowledgements
The present study was supported by Scientific and
Technologic Innovation Action of Shanghai Municipal Science and
Technology Commission (grant no. 15401971800).
References
|
1
|
Zhang J, Liu J, Chen J, Li X, Wu Y, Chen
H, Wu W, Zhang K and Gu L: Angiotensin receptor blockers (ARBs)
reduce the risk of lung cancer: A systematic review and
meta-analysis. Int J Clin Exp Med. 8:12656–12660. 2015.PubMed/NCBI
|
|
2
|
Qiu L, Lan L, Feng Y, Huang Z and Chen Y:
Pulmonary actinomycosis imitating lung cancer on (18)F-FDG PET/CT:
A case report and literature review. Korean J Radiol. 16:1262–1265.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Dong QZ, Wang Y, Tang ZP, Fu L, Li QC,
Wang ED and Wang EH: Derlin-1 is overexpressed in non-small cell
lung cancer and promotes cancer cell invasion via EGFR-ERK-mediated
up-regulation of MMP-2 and MMP-9. Am J Pathol. 182:954–964. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Rao JS, Gondi C, Chetty C, Chittivelu S,
Joseph PA and Lakka SS: Inhibition of invasion, angiogenesis, tumor
growth and metastasis by adenovirus-mediated transfer of antisense
uPAR and MMP-9 in non-small cell lung cancer cells. Mol Cancer
Ther. 4:1399–1408. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Yue W, Ming QL, Lin B, Rahman K, Zheng CJ,
Han T and Qin LP: Medicinal plant cell suspension cultures:
Pharmaceutical applications and high-yielding strategies for the
desired secondary metabolites. Crit Rev Biotechnol. 36:215–232.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ku SK, Zhou W, Lee W, Han MS, Na M and Bae
JS: Anti-inflammatory effects of hyperoside in human endothelial
cells and in mice. Inflammation. 38:784–799. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Sukito A and Tachibana S: Isolation of
hyperoside and isoquercitrin from Camellia sasanqua as antioxidant
agents. Pak J Biol Sci. 17:999–1006. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang J, Zhao F, Yu X, Lu X and Zheng G:
Pharmacokinetics of eupalinolide A eupalinolide B and hyperoside
from Eupatorium lindleyanum in rats by LC/MS/MS. J Chromatogr B
Analyt Technol Biomed Life Sci. 995–996:1–7. 2015. View Article : Google Scholar
|
|
9
|
Yan Y, Feng Y, Li W, Che JP, Wang GC, Liu
M and Zheng JH: Protective effects of quercetin and hyperoside on
renal fibrosis in rats with unilateral ureteral obstruction. Exp
Ther Med. 8:727–730. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al-Mahdi R, Babteen N, Thillai K, Holt M,
Johansen B, Wetting HL, Seternes OM and Wells CM: A novel role for
atypical MAPK kinase ERK3 in regulating breast cancer cell
morphology and migration. Cell Adh Migr. 9:483–494. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jung HG, Kim HH, Paul S, Jang JY, Cho YH,
Kim HJ, Yu JM, Lee ES, An BJ, Kang SC and Bang BH:
Quercetin-3-O-β-d-glucopyranosyl-(1→6)-β-d-glucopyranoside
suppresses melanin synthesis by augmenting p38 MAPK and CREB
signaling pathways and subsequent cAMP down-regulation in murine
melanoma cells. Saudi J Biol Sci. 22:706–713. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chang D, Wang YC, Bai YY, Lu CQ, Xu TT,
Zhu L and Ju S: Role of P38 MAPK on MMP activity in photothrombotic
stroke mice as measured using an ultrafast MMP activatable probe.
Sci Rep. 5:169512015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huo YN, Chen W and Zheng XX: ROS, MAPK/ERK
and PKC play distinct roles in EGF-stimulated human corneal cell
proliferation and migration. Cell Mol Biol (Noisy-le-grand).
61:6–11. 2015.PubMed/NCBI
|
|
14
|
Ong JY, Yong PV, Lim YM and Ho AS:
2-Methoxy-1,4-naphthoquinone (MNQ) induces apoptosis of A549 lung
adenocarcinoma cells via oxidation-triggered JNK and p38 MAPK
signaling pathways. Life Sci. 135:158–164. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bu HQ, Cai K, Shen F, Bao XD, Xu Y, Yu F,
Pan HQ, Chen CH, Du ZJ and Cui JH: Induction of apoptosis by
capsaicin in hepatocellular cancer cell line SMMC-7721 is mediated
through ROS generation and activation of JNK and p38 MAPK pathways.
Neoplasma. 62:582–591. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lee HH, Park C, Jeong JW, Kim MJ, Seo MJ,
Kang BW, Park JU, Kim GY, Choi BT, Choi YH and Jeong YK: Apoptosis
induction of human prostate carcinoma cells by cordycepin through
reactive oxygen species-mediated mitochondrial death pathway. Int J
Oncol. 42:1036–1044. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hsieh YJ, Chang CJ, Wan CF, Chen CP, Chiu
YH, Leu YL and Peng KC: Euphorbia formosana root extract induces
apoptosis by caspase-dependent cell death via Fas and mitochondrial
pathway in THP-1 human leukemic cells. Molecules. 18:1949–1962.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Niemi NM and MacKeigan JP: Mitochondrial
phosphorylation in apoptosis: Flipping the death switch. Antioxid
Redox Signal. 19:572–582. 2013. View Article : Google Scholar : PubMed/NCBI
|