Transforming growth factor-β (TGF-β) signaling forms
a complex web in the progression of cancer. There is substantial
evidence indicating that downregulated TGF-β signaling in tumor
initiation. However, in certain tumors, TGF-β appears to have the
ability to exert a tumor-promoting effect depending on cellular
context (1–3). Accordingly, TGF-β signaling has been
considered as a tumor suppressor and a promoter of tumor
progression (4,5). It acts as a tumor suppressor by
inhibiting cell proliferation through repressing the expression of
c-Myc and certain cyclin-depenent kinase inhibitors (CDKIs) and
through the secretion of anti-angiogenic factors (6,7). It
functions as a tumor promoter through the stimulation of matrix
deposition, perturbation of immune function and induction of
epithelial-mesenchymal transition (EMT) (8). The TGF-β super-family is a large
group of structurally associated proteins including TGF-β, nodal,
activin, lefty, bone morphogenetic proteins and growth and
differentiation factor. There are numerous cellular
context-dependent factors tightly implicated in the balance of
TGF-β signaling, thus TGF-β signaling forms a complicated network
in cancer cells (5).
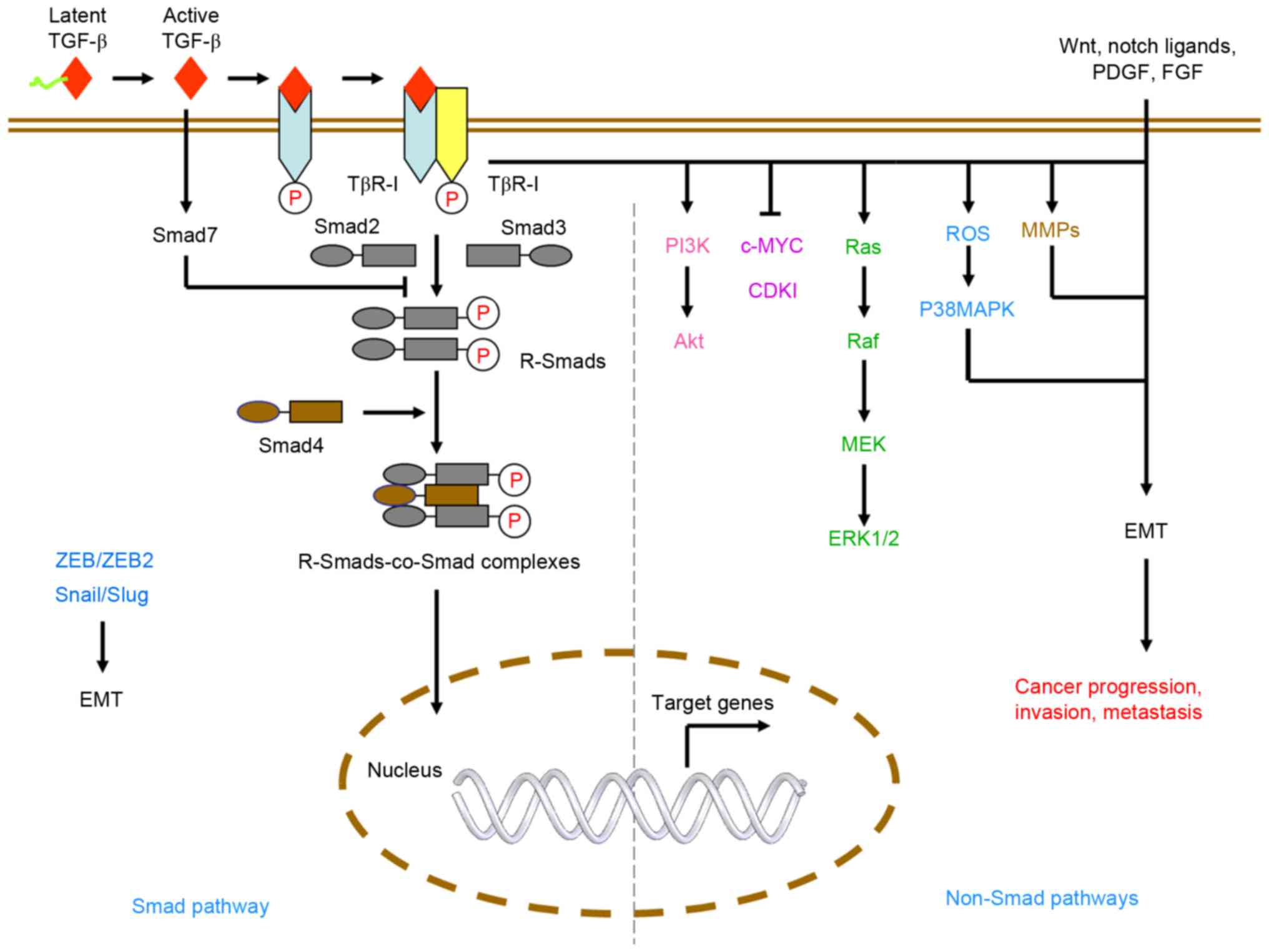
TGF-β signaling is transduced through Smad and
non-Smad pathways. These pathways are mediated by TGF-β ligands,
type1 and type2 receptors and Smad or non-Smad proteins, including
Akt, extracellular signal-regulated kinase (ERK)1/2 and p38
mitogen-activated protein kinase (MAPK; Fig. 1). In mammals, there are three types
of TGF-β (TGF-β1, 2 and 3), which are encoded by different genes
and which function through the same receptor signaling systems
(9). Of these, TGF-β1 is most
frequently upregulated in tumor cells (10,11).
However, the fate of cells following TGF-β1 treatment is often
determined by cellular context and experimental conditions
(12–14).
The effects of TGF-β signaling are mediated by three
TGF-β ligands (TGF-β1-3) through TGF-β type1 (TGFβR1) and type2
receptors TGFβR2 (21–23). TGFβR1 recruits and phosphorylates
receptor-regulated Smads (R-Smads) when phosphorylated. TGF-β
ligands can bind to TGFβR2 with high affinity once activated by
metalloproteinases. Binding of the ligands causes the formation of
a heterotetrameric active receptor complex, which results in the
phosphorylation of TGFβR1 by TGFβR2. The phosphorylation of R-Smads
by TGFβR1 form heteromeric complexes with the common partner Smad
(co-Smad; Smad4 in mammals) (17)
and these R-Smads-co-Smad complexes translocate into the nucleus to
regulate the expression of target genes with other DNA-binding
transcription factors.
TGF-β can also activate non-canonical signaling
pathways, also termed non-Smad pathways. For example, TGF-β1 is
known to activate the Erk/MAPK (24,25)
pathway and the phosphoinositide 3-kinase (PI3K)/Akt (26–28)
pathway. These non-Smad pathways work independently or together
with Smad complexes to regulate the functions of TGF-β. For
example, the activation of Akt signaling by TGF-β1 has been shown
to promote cell proliferation (28). TGF-β receptors activate MMPs,
p38MAPK and Zinc finger E-box-binding homeobox 1 (ZEB1), ZEB2,
Snail and Slug, leading to EMT, which is required for cancer cell
invasion and metastasis. TGF-β can also regulate target gene
expression through the MAPK signaling pathway (29,30).
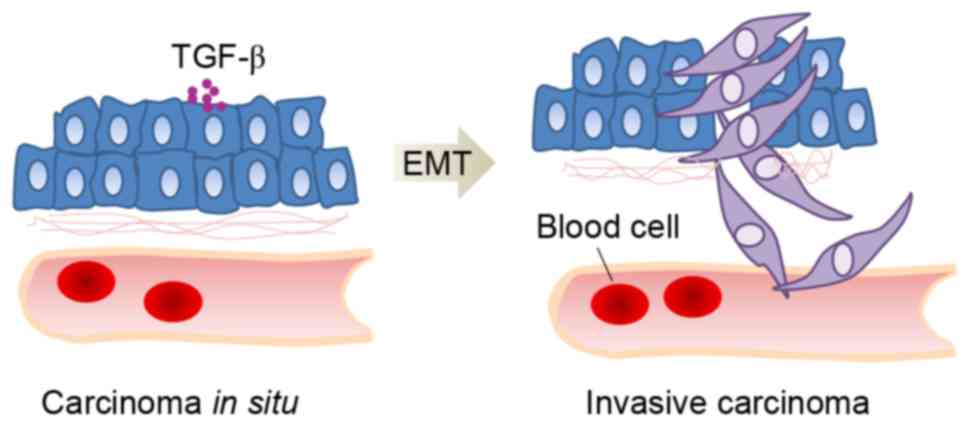
EMT is a key step in the progression of cancer
invasion and metastasis, characterized by reduced epithelial marker
and elevated mesenchymal marker expression (36,37).
The EMT process is often associated with upregulation of TGF-β
signaling and TGF-β drives EMT through the Smad-mediated or
non-Smad-mediated reprogramming of gene expression (Fig. 2) (38). TGF-β receptors can induce the
expression of ZEB1, ZEB2, Snail and Slug through the Smad pathway
and mediate MMPs and p38MAPK through non-Smad pathways, which both
lead to EMT (39). ROS are also
important in TGF-β-induced EMT, primarily through the activation of
MAPK and through subsequent ERK-directed activation of the Smad
pathway in proximal tubular epithelial cells (40). TGF-β signaling not only mediates
EMT, but it is also required for distant metastases. A previous
study demonstrated that TGF-β in the breast cancer microenvironment
promoted pulmonary metastases of breast cancer cells (41).
Advances in cancer stem cell (CSC) research have
indicated that the TGF-β signaling pathway is essential for the
maintenance of CSCs. CSCs have been reported to be responsible for
the recurrence of disease following anticancer therapy (42). Certain extracellular and
intracellular signals allow cancer progenitors to dynamically
revert to a stem cell state. Evidence has shown that TGF-β induced
the expression of CD133, a CSC marker, in liver cancer cell lines
and increased tumor initiating ability in mice, compared with the
milder and transient effect of interleukin-6 (43). Chanmee et al found that the
over production of hyaluronan allowed cancer progenitors to revert
to stem cell states via Twist and the TGF-β-Snail signaling axis
(44). Furthermore,
tumor-associated macrophages (TAMs) in the tumor microenvironment
promoted CSC-like properties via TGF-β1-induced EMT in HCC
(45). MicroRNA (miR)-106b is
significantly upregulated in CD44(+) cells and the inhibition of
miR-106b shows suppression of the TGF-β/Smad signaling pathway and
decreases self-renewal capacity and cell invasiveness (46). These findings suggest that TGF-β
signaling is associated with cancer cell stemness.
There are epigenetic differences in TGF-β
signaling-related genes between cancer stem-like cells and
differentiated cancer cells. It was previously reported that the
methylation levels of genes coding TGF-β signaling-related proteins
can regulate breast cancer stem cell differentiation (47). Evidence suggests that several
members of the TGF-β pathway are targeted by long non-coding RNAs,
key epigenetic mediators and this may, at least in part, explain
the association between cancer cell stemness and TGF-β signaling
(48). These findings may assist
in the development of novel agents, which can specifically control
increases and decreases in the TGF-β signaling pathway in CSCs and
thereby provide a novel avenue for the prevention and treatment of
malignant cancer (48).
Acute inflammation is an essential component of the
wound-healing response to stimuli. However, chronic inflammation
favors the accumulation of mutations and epigenetic aberrations in
normal cells, thereby promoting malignant transformation (49,50).
This process is mediated by chemokines, cytokines and growth
factors secreted by stromal components of the tumor
microenvironment. Among those secreted factors, the TGF-β subfamily
has been shown to generate a favorable immune microenvironment for
tumor growth. TGF-β can impair anticancer immune responses in
several ways, including immune cell inhibition and the elimination
of major histocompatibility complex class I and II (51). It can also induce TAMs and generate
ROS with genotoxic activity (52–54).
TAMs are primarily a macrophage subpopulation with an M2 phenotype.
TAM-derived factors may enhance the invasiveness of tumor cells by
enhancing their adhesion to extracellular matrix in the tumor
stroma (55,56). Evidence from clinical and
epidemiological studies has shown that TAM density is positively
associated with poor survival rates in several types of cancer
(56,57). The involvement of ROS signaling in
tumor progression has also been recognized (58,59).
Previous studies have suggested that the increased generation of
ROS in tumor cells may affect certain redox-sensitive molecules,
leading to mutations and genetic instability, cellular
proliferation and metastasis (60,61).
The ability of tumor cells to induce the formation
of blood vessels is crucial for tumor growth, invasion and
metastasis. TGF-β is a key mediator of angiogenesis in the tumor
microenvironment, contributing to angiogenesis by inducing
proangiogenic factors (62).
TGF-β can induce a proangiogenic environment and
stimulate tumor-associated angiogenesis. Elevated expression levels
of TGF-β have been linked to increased microvessel density in
certain tumor types (63). The
mechanism of angiogenesis stimulated by TGF-β signaling includes
the induction of key angiogenic factors, including connective
tissue growth factor, vascular endothelial growth factor and
insulin-like growth factor-binding protein 7, in epithelial cells
and fibroblasts (64,65). In addition, TGF-β can induce the
expression, secretion and activation of matrix metalloproteinase 2
(MMP2) and MMP9 and down regulate the expression of tissue
inhibitor of metalloproteinase in tumor and endothelial cells
(66–68).
However, TGF-β also has angiostatic functions. For
example, in pancreatic cancer and diffuse-type gastric cancer,
TGF-β induces the production of thrombospondin1, a potent
angiogenic inhibitor, whereas perturbations of TGF-β signaling
resulted in accelerated angiogenesis and growth of tumors (69). Whether TGF-β is angiogenic or
angiostaticis is dependent on the cellular context of tumor cells,
epithelial cells and the tumor microenvironment.
Several lines of evidence suggest that the TGF-β
family is involved in tumor initiation and progression, including
cell proliferation, angiogenesis, cancer cell stemness, EMT,
invasion and inflammation. TGF-β signaling is complex and mediates
pro- and anti-tumoral activities in cancer cells depending on their
context in space and time and their microenvironment (52). It is generally accepted that TGF-β
is primarily a tumor suppressor in premalignant cells but functions
as a promoter of metastasis in cancer cells (70,71).
However, the mechanisms underlying the contextual changes in the
role of TGF-β remain to be elucidated. Xu et al provided
molecular insight into how TGF-β converts from a tumor suppressor
to a tumor promoter. It was found that 14-3-3ζ turned off the tumor
suppression function of TGF-β by destabilizing p53, a Smad partner
in premalignant cells. By contrast, 14-3-3ζ promoted TGF-β-induced
distant metastasis by stabilizing Gli family zinc finger 2 (Gli2)
and inducing the partnering of Gli2 with Smads in malignant cells
(70). Therefore, the
14-3-3ζ-driven contextual changes of Smad partners from p53 to Gli2
may provide therapeutic targets in TGF-β-mediated cancer
progression.
This review was supported by grants from the
National Natural Science Foundation of China (grant no. 81502131),
The Natural Science Foundation of Chongqing (grant no.
cstc2016jcyjA0405) and the Science and Technology Project of
Chongqing Municipal Education Commission (grant no. KJ1500332).
|
1
|
Morris SM, Carter KT, Baek JY, Koszarek A,
Yeh MM, Knoblaugh SE and Grady WM: TGF-β signaling alters the
pattern of liver tumorigenesis induced by Pten inactivation.
Oncogene. 34:3273–3282. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Horie Y, Suzuki A, Kataoka E, Sasaki T,
Hamada K, Sasaki J, Mizuno K, Hasegawa G, Kishimoto H, Iizuka M, et
al: Hepatocyte-specific Pten deficiency results in steatohepatitis
and hepatocellular carcinomas. J Clin Invest. 113:1774–1783. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Attisano L and Wrana JL: Signal
integration in TGF-β, WNT, and Hippo pathways. F1000Prime Rep.
5:172013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ikushima H and Miyazono K: TGFbeta
signalling: A complex web in cancer progression. Nat Rev Cancer.
10:415–424. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Derynck R, Akhurst RJ and Balmain A:
TGF-beta signaling in tumor suppression and cancer progression. Nat
Genet. 29:117–129. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kiyono K, Suzuki HI, Morishita Y, Komuro
A, Iwata C, Yashiro M, Hirakawa K, Kano MR and Miyazono K: c-Ski
overexpression promotes tumor growth and angiogenesis through
inhibition of transforming growth factor-beta signaling in
diffuse-type gastric carcinoma. Cancer Sci. 100:1809–1816. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Komuro A, Yashiro M, Iwata C, Morishita Y,
Johansson E, Matsumoto Y, Watanabe A, Aburatani H, Miyoshi H,
Kiyono K, et al: Diffuse-type gastric carcinoma: Progression,
angiogenesis, and transforming growth factor beta signaling. J Natl
Cancer Inst. 101:592–604. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roberts AB and Wakefield LM: The two faces
of transforming growth factor beta in carcinogenesis. Proc Natl
Acad Sci USA. 100:8621–8623. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Massagué J: TGF-beta signal transduction.
Annu Rev Biochem. 67:753–791. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Massagué J, Blain SW and Lo RS: TGFbeta
signaling in growth control, cancer, and heritable disorders. Cell.
103:295–309. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang Y, Liu T, Tang W, Deng B, Chen Y, Zhu
J and Shen X: Hepatocellular carcinoma cells induce regulatory T
cells and lead to poor prognosis via production of transforming
growth factor-β1. Cell Physiol Biochem. 38:306–318. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shen H, Guan D, Shen J, Wang M, Chen X, Xu
T, Liu L and Shu Y: TGF-β1 induces erlotinib resistance in
non-small cell lung cancer by down-regulating PTEN. Biomed
Pharmacother. 77:1–6. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yoshimoto T, Fujita T, Kajiya M, Matsuda
S, Ouhara K, Shiba H and Kurihara H: Involvement of smad2 and
Erk/Akt cascade in TGF-β1-induced apoptosis in human gingival
epithelial cells. Cytokine. 75:165–173. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dai C, Yang J and Liu Y: Transforming
growth factor-beta1 potentiates renal tubular epithelial cell death
by a mechanism independent of Smad signaling. J Biol Chem.
278:12537–12545. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lyons RM, Gentry LE, Purchio AF and Moses
HL: Mechanism of activation of latent recombinant transforming
growth factor beta 1 by plasmin. J Cell Biol. 110:1361–1367. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Andreasen PA, Kjøller L, Christensen L and
Duffy MJ: The urokinase-type plasminogen activator system in cancer
metastasis: A review. Int J Cancer. 72:1–22. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yu Q and Stamenkovic I: Cell
surface-localized matrix metalloproteinase-9 proteolytically
activates TGF-beta and promotes tumor invasion and angiogenesis.
Genes Dev. 14:163–176. 2000.PubMed/NCBI
|
|
18
|
Mir FA, Contreras-Ruiz L and Masli S:
Thrombospondin-1-dependent immune regulation by transforming growth
factor-β2-exposed antigen-presenting cells. Immunology.
146:547–556. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Murphy-Ullrich JE and Poczatek M:
Activation of latent TGF-beta by thrombospondin-1: Mechanisms and
physiology. Cytokine Growth Factor Rev. 11:59–69. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dutta A, Li J, Fedele C, Sayeed A, Singh
A, Violette SM, Manes TD and Languino LR: αvβ6 integrin is required
for TGFβ1-mediated matrix metalloproteinase2 expression. Biochem J.
466:525–536. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Feng XH and Derynck R: Specificity and
versatility in tgf-beta signaling through Smads. Annu Rev Cell Dev
Biol. 21:659–693. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shi Y and Massagué J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Heldin CH, Miyazono K and ten Dijke P:
TGF-beta signalling from cell membrane to nucleus through SMAD
proteins. Nature. 390:465–471. 1997. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Park S, Jung HH, Park YH, Ahn JS and Im
YH: ERK/MAPK pathways play critical roles in EGFR ligands-induced
MMP1 expression. Biochem Biophys Res Commun. 407:680–686. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cheng X, Gao W, Dang Y, Liu X, Li Y, Peng
X and Ye X: Both ERK/MAPK and TGF-Beta/Smad signaling pathways play
a role in the kidney fibrosis of diabetic mice accelerated by blood
glucose fluctuation. J Diabetes Res. 2013:4637402013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yu JS, Ramasamy TS, Murphy N, Holt MK,
Czapiewski R, Wei SK and Cui W: PI3K/mTORC2 regulates TGF-β/Activin
signalling by modulating Smad2/3 activity via linker
phosphorylation. Nat Commun. 6:72122015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Vo BT, Morton D Jr, Komaragiri S, Millena
AC, Leath C and Khan SA: TGF-β effects on prostate cancer cell
migration and invasion are mediated by PGE2 through activation of
PI3K/AKT/mTOR pathway. Endocrinology. 154:1768–1779. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Singha PK, Pandeswara S, Geng H, Lan R,
Venkatachalam MA and Saikumar P: TGF-β induced TMEPAI/PMEPA1
inhibits canonical Smad signaling through R-Smad sequestration and
promotes non-canonical PI3K/Akt signaling by reducing PTEN in
triple negative breast cancer. Genes Cancer. 5:320–336.
2014.PubMed/NCBI
|
|
29
|
Reduced beta 2 glycoprotein I improve
diabetic nephropathy via inhibiting TGF-β1-p38 MAPK pathway
[Retraction]. Int J Clin Exp Med. 8:197922015.PubMed/NCBI
|
|
30
|
Chen IT, Hsu PH, Hsu WC, Chen NJ and Tseng
PH: Polyubiquitination of transforming growth factor β-activated
Kinase 1 (TAK1) at lysine 562 residue regulates TLR4-mediated JNK
and p38 MAPK activation. Sci Rep. 5:123002015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Suzuki T, Dai P, Hatakeyama T, Harada Y,
Tanaka H, Yoshimura N and Takamatsu T: TGF-β signaling regulates
pancreatic β-Cell proliferation through control of cell cycle
regulator p27 expression. Acta Histochem Cytochem. 46:51–58. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Senturk S, Mumcuoglu M, Gursoy-Yuzugullu
O, Cingoz B, Akcali KC and Ozturk M: Transforming growth
factor-beta induces senescence in hepatocellular carcinoma cells
and inhibits tumor growth. Hepatology. 52:966–974. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang Y, Wu J, Lin B, Li X, Zhang H, Ding
H, Chen X, Lan L and Luo H: Galangin suppresses HepG2 cell
proliferation by activating the TGF-β receptor/Smad pathway.
Toxicology. 326:9–17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Guo C, Liu S, Dong P, Zhao D, Wang C, Tao
Z and Sun MZ: Akbu-LAAO exhibits potent anti-tumor activity to
HepG2 cells partially through produced H2O2 via TGF-β signal
pathway. Sci Rep. 5:182152015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Cheng L, Zhang C, Li D, Zou J and Wang J:
Transforming growth factor-β1 (TGF-β1) induces mouse
precartilaginous stem cell proliferation through TGF-β receptor II
(TGFRII)-Akt-β-catenin signaling. Int J Mol Sci. 15:12665–12676.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kudo-Saito C, Shirako H, Takeuchi T and
Kawakami Y: Cancer metastasis is accelerated through
immunosuppression during Snail-induced EMT of cancer cells. Cancer
Cell. 15:195–206. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hay ED: An overview of
epithelio-mesenchymal transformation. Acta Anat (Basel). 154:8–20.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Muthusamy BP, Budi EH, Katsuno Y, Lee MK,
Smith SM, Mirza AM, Akhurst RJ and Derynck R: ShcA protects against
epithelial-mesenchymal transition through compartmentalized
inhibition of TGF-β-Induced Smad activation. PLoS Biol.
13:e10023252015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xu J, Lamouille S and Derynck R:
TGF-beta-induced epithelial to mesenchymal transition. Cell Res.
19:156–172. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lee JY, Chang JW, Yang WS, Kim SB, Park
SK, Park JS and Lee SK: Albumin-induced epithelial-mesenchymal
transition and ER stress are regulated through a common ROS-c-Src
kinase-mTOR pathway: Effect of imatinib mesylate. Am J Physiol
Renal Physiol. 300:F1214–1222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Padua D, Zhang XH, Wang Q, Nadal C, Gerald
WL, Gomis RR and Massagué J: TGFbeta primes breast tumors for lung
metastasis seeding through angiopoietin-like 4. Cell. 133:66–77.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Naka K: TGF-β signaling in cancer stem
cells. Nihon Rinsho. 73:784–789. 2015.(In Japanese). PubMed/NCBI
|
|
43
|
You H, Ding W and Rountree CB: Epigenetic
regulation of cancer stem cell marker CD133 by transforming growth
factor-beta. Hepatology. 51:1635–1644. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Chanmee T, Ontong P, Mochizuki N,
Kongtawelert P, Konno K and Itano N: Excessive hyaluronan
production promotes acquisition of cancer stem cell signatures
through the coordinated regulation of Twist and the transforming
growth factor β (TGF-β)-Snail signaling axis. J Biol Chem.
289:26038–26056. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fan QM, Jing YY, Yu GF, Kou XR, Ye F, Gao
L, Li R, Zhao QD, Yang Y, Lu ZH and Wei LX: Tumor-associated
macrophages promote cancer stem cell-like properties via
transforming growth factor-beta1-induced epithelial-mesenchymal
transition in hepatocellular carcinoma. Cancer Lett. 352:160–168.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yu D, Shin HS, Lee YS and Lee YC: miR-106b
modulates cancer stem cell characteristics through TGF-β/Smad
signaling in CD44-positive gastric cancer cells. Lab Invest.
94:1370–1381. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
El Helou R, Wicinski J, Guille A, Adélaïde
J, Finetti P, Bertucci F, Chaffanet M, Birnbaum D, Charafe-Jauffret
E and Ginestier C: Brief reports: A distinct DNA methylation
signature defines breast cancer stem cells and predicts cancer
outcome. Stem Cells. 32:3031–3036. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Wang J, Shao N, Ding X, Tan B, Song Q,
Wang N, Jia Y, Ling H and Cheng Y: Crosstalk between transforming
growth factor-β signaling pathway and long non-coding RNAs in
cancer. Cancer Lett. 370:296–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Martin M and Herceg Z: From hepatitis to
hepatocellular carcinoma: A proposed model for cross-talk between
inflammation and epigenetic mechanisms. Genome Med. 4:82012.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Hernandez-Gea V, Toffanin S, Friedman SL
and Llovet JM: Role of the microenvironment in the pathogenesis and
treatment of hepatocellular carcinoma. Gastroenterology.
144:512–527. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Nana AW, Yang PM and Lin HY: Overview of
transforming growth factor β superfamily involvement in
glioblastoma initiation and progression. Asian Pac J Cancer Prev.
16:6813–6823. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Neuzillet C, de Gramont A,
Tijeras-Raballand A, de Mestier L, Cros J, Faivre S and Raymond E:
Perspectives of TGF-β inhibition in pancreatic and hepatocellular
carcinomas. Oncotarget. 5:78–94. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Yang L, Pang Y and Moses HL: TGF-beta and
immune cells: An important regulatory axis in the tumor
microenvironment and progression. Trends Immunol. 31:220–227. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Yang L: TGFbeta, a potent regulator of
tumor microenvironment and host immune response, implication for
therapy. Curr Mol Med. 10:374–380. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen J, Yao Y, Gong C, Yu F, Su S, Chen J,
Liu B, Deng H, Wang F, Lin L, et al: CCL18 from tumor-associated
macrophages promotes breast cancer metastasis via PITPNM3. Cancer
Cell. 19:541–555. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pittet MJ: Behavior of immune players in
the tumor microenvironment. Curr Opin Oncol. 21:53–59. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Condeelis J and Pollard JW: Macrophages:
obligate partners for tumor cell migration, invasion and
metastasis. Cell. 124:263–266. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Storz P: Reactive oxygen species in tumor
progression. Front Biosci. 10:1881–1896. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Radisky DC, Levy DD, Littlepage LE, Liu H,
Nelson CM, Fata JE, Leake D, Godden EL, Albertson DG, Nieto MA, et
al: Rac1b and reactive oxygen species mediate MMP-3-induced EMT and
genomic instability. Nature. 436:123–127. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Pelicano H, Carney D and Huang P: ROS
stress in cancer cells and therapeutic implications. Drug Resist
Updat. 7:97–110. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Suh YA, Arnold RS, Lassegue B, Shi J, Xu
X, Sorescu D, Chung AB, Griendling KK and Lambeth JD: Cell
transformation by the superoxide-generating oxidase Mox1. Nature.
401:79–82. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
62
|
Kalluri R: Basement membranes: Structure,
assembly and role in tumour angiogenesis. Nat Rev Cancer.
3:422–433. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hasegawa Y, Takanashi S, Kanehira Y,
Tsushima T, Imai T and Okumura K: Transforming growth factor-beta1
level correlates with angiogenesis, tumor progression, and
prognosis in patients with nonsmall cell lung carcinoma. Cancer.
91:964–971. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Kang Y, Siegel PM, Shu W, Drobnjak M,
Kakonen SM, Cordón-Cardo C, Guise TA and Massagué J: A multigenic
program mediating breast cancer metastasis to bone. Cancer Cell.
3:537–549. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sánchez-Elsner T, Botella LM, Velasco B,
Corbi A, Attisano L and Bernabéu C: Synergistic cooperation between
hypoxia and transforming growth factor-beta pathways on human
vascular endothelial growth factor gene expression. J Biol Chem.
276:38527–38535. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Zhao J, Cheng Q, Ye P, Yang G, Liu S, Ao
Q, Liu Y and Hu Y: Atorvastatin improves pathological changes in
the aged kidney by upregulating peroxisome proliferator-activated
receptor expression and reducing matrix metalloproteinase-9 and
transforming growth factor-β1 levels. Exp Gerontol. 74:37–42. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Hua Y, Zhang W, Xie Z, Xu N and Lu Y:
MMP-2 is mainly expressed in arterioles and contributes to cerebral
vascular remodeling associated with TGF-β1 signaling. J Mol
Neurosci. 59:317–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Şekerci ÇA, Işbilen B, Işman F, Akbal C,
Şimşek F and Tarcan T: Urinary NGF, TGF-β1, TIMP-2 and bladder wall
thickness predict neurourological findings in children with
myelodysplasia. J Urol. 191:199–205. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Schwarte-Waldhoff I, Volpert OV, Bouck NP,
Sipos B, Hahn SA, Klein-Scory S, Lüttges J, Klöppel G, Graeven U,
Eilert-Micus C, et al: Smad4/DPC4-mediated tumor suppression
through suppression of angiogenesis. Proc Natl Acad Sci USA.
97:9624–9629. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Xu J, Acharya S, Sahin O, Zhang Q, Saito
Y, Yao J, Wang H, Li P, Zhang L, Lowery FJ, et al: 14-3-3ζ turns
TGF-β's function from tumor suppressor to metastasis promoter in
breast cancer by contextual changes of Smad partners from p53 to
Gli2. Cancer Cell. 27:177–192. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Akahira J, Sugihashi Y, Suzuki T, Ito K,
Niikura H, Moriya T, Nitta M, Okamura H, Inoue S, Sasano H, et al:
Decreased expression of 14-3-3 sigma is associated with advanced
disease in human epithelial ovarian cancer: Its correlation with
aberrant DNA methylation. Clin Cancer Res. 10:2687–2693. 2004.
View Article : Google Scholar : PubMed/NCBI
|