Introduction
Recent studies indicate that microRNAs (miRNAs) play
an important role in neural growth and development (1,2).
Dicer deficiency may result in the occurrence of neurodegenerative
diseases, and inhibition of miRNA synthesis leads to loss of
dopaminergic neurons (3). Loss of
miRNA activity in Dicer1-deficient mice was found to cause spinal
motor neuron disease, which was attributed to dysregulation of
miR-9 (4). miRNA profiling
revealed differential miRNA expression between healthy subjects and
patients with neurodegenerative diseases. Pre-miR-133b, −218-2,
−15b, −101-1, −107, −335 and −345 were found to be downregulated in
the midbrain of patients with Parkinson's disease (5). miRNA-145 was found to be markedly
increased during neural stem cell differentiation, and inhibition
of miRNA-145 downregulated the expression of neuronal markers
(6). In another study, miRNA-124
was found to increase neuron formation in the subventricular zone
of adult mouse brain (7). In
addition, the role of miR-9 was found to be crucial for neural
progenitor proliferation and differentiation in the developing
telencephalon (8).
Mouse miRNA-153 (miR-153) is located on chromosome
12. Certain disease-related proteins were found to be targets of
miR-153. α-synuclein accumulation was shown to play a central role
in the pathogenesis of Parkinson's disease. Doxakis et al
reported that miRNA-7 and miR-153 can downregulate α-synuclein at
the mRNA and protein levels (9).
It was also reported that overexpression of miR-153 caused SNAP25
downregulation and resulted in near complete paralysis in zebrafish
embryos (10). Another study on
miR-153 demonstrated its inhibitory effect on gliogenesis by
targeting Nfia/b in mouse neural stem/progenitor cells (11).
During neural differentiation, miR-153 may regulate
certain neurogenesis-related genes, including
N-ethylmaleimide-sensitive fusion attachment proteins (SNAPs).
SNAPs and their receptors (SNAREs) constitute the core machinery
for membrane fusion and are essential for intracellular vesicular
trafficking. SNAP23 and SNAP25 are involved in neural regeneration
and differentiation (12).
Peroxiredoxin (PRX)5 was found to protect neural cells from
amyloid-beta oligomer (AβO) damage (13). However, the function of miR-153 in
neural development and differentiation and its relevance to the
mechanism of neurodegenerative diseases has not been fully
elucidated.
The aim of the present study was to investigate the
role of miR-153 in neural development. A series of tests were
performed in vitro to evaluate cell morphology, cell growth,
cell cycle distribution, neural development-related genes and
protein expression in the HT-22 cell line, in order to determine
whether miR-153 induces neural differentiation and elucidate its
effect on the expression of neuron-specific γ-enolase (NSE),
neuronal nuclei (NeuN), PRX5, SNAP23 and SNAP25.
Materials and methods
Construction of the miR-153 plasmid
and lentivirus packaging
Mouse precursor miR-153 (pre-miR153, miRbase
accession no. MI0000175) was synthesized and further constructed
into the pLVX-ZsGreen-miRNA-Puro vector, and packaged into
lentivirus by Wuhan Viraltherapy Technologies Co., Ltd. The final
titers ranged between 107 and 108 transducing
units (TU)/ml.
Cell culture, infection and monoclone
screen
The mouse hippocampal HT-22 cell line was purchased
from Shanghai Xiaoying Biotech Company. HT-22 cells were maintained
at 37°C and 5% CO2 in DMEM/high glucose (Gibco; Thermo
Fisher Scientific, Inc.) supplemented with 10% FBS and 100 U/ml
penicillin/streptomycin (Gibco; Thermo Fisher Scientific, Inc.).
HT-22 cells were infected separately with miR-153 viral particles
and control viral particles at 5х106 TU/106
cells. The green fluorescent protein (GFP) expression in the vector
was used to determine transfection efficiency. After 48 h of
infection, HT-22 cells were passaged and several monoclones were
selected and further cultured into stably infected HT-22 cells. The
stably miR-153-infected and control HT-22 cells were used in all
further analyses.
miRNA extraction, quantitative
polymerase chain reaction (qPCR) and regular PCR analysis (14)
The kits for miRNA isolation (DP501), miRNA
First-Strand cDNA Synthesis (KR211) and miRNA qPCR Detection
(FP411) were purchased from Tiangen Biotech Co., Ltd. For miRNA
isolation, the main procedures were as follows. The cells were
lysed in lysis buffer and maintained at room temperature for 5 min;
200 µl chloroform was added, vortexed for 15 sec and kept at room
temperature for 5 min. The mixture was then centrifuged at 4°C at
10,000 × g for 15 min, and the upper layer of the aqueous phase was
transferred to a new tube. Next, a proper volume of ethanol was
added, and the solution was vortexed and transferred to a spin
column. The solution was centrifuged and the eluate was saved; a
proper volume of ethanol was added, vortexed and transferred to a
spin column, followed by centrifugation at 10,000 g for 30 sec at
room temperature. After rinsing twice, the miRNA in the spin column
was dissolved in RNase-free ddH2O. For miRNA
First-Strand cDNA Synthesis, up to 2 µg of miRNA was mixed with
reaction buffer and enzymes to a final volume of 20 µl for each
test; the mixtures were treated at 42°C for 60 min and then at 95°C
for 5 min to denature the enzymes. For miRNA qPCR, the StepOnePlus
PCR System (Applied Biosystems; Thermo Fisher Scientific, Inc.) was
applied. A total of 20 µl of reaction system was prepared,
containing 10 µl 2Х miRNA Premix (with SYBR & ROX), forward
primers (synthesized by Shanghai Sangon Biotech Company) and
reverse primers (provided in the kit, 10 µm for each) and 2 µl
miRNA First-Strand cDNA. qPCR reactions were carried out in
triplicate, and the data were normalized against the levels of U6
RNA. The sequences of mature mmu-miR-153-5p (accession no.
MIMAT0016992) and mmu-miR-153-3p (accession no. MIMAT0000163) were
used as forward primers in the qPCR analysis. Regular PCR was
performed to detect mouse pre-miR-153 expression. The primers of
regular PCR were as follows: Forward, CGGTGTCATTTTTGTGACGT and
reverse, CAATGATCACTTTTGTGACT. The expression of pre-miR-153 was
analyzed after 38 cycles of regular PCR.
Analysis of cell viability
Cell viability was assessed with the CCK-8 assay.
The CCK-8 kit was purchased from Signalway Antibody Co. (Lot:
8620). HT-22 cells stably expressing rLV-miR and rLV-miR-153 were
cultured in 96-well plates (1х104 cells per well). After
incubation for 24 and 48 h, the cells were then incubated with
CCK-8 (10 µl CCK-8 added to each well) for 1 h at 37°C. Absorbance
was measured at 450 nm by using SpetraMax M3 (Molecular Devices
LLC).
Cell cycle analysis
The Cell Cycle Analysis Kit was purchased from
Beyotime Institute of Biotechnology. Cells were cultured in 6-well
plates and were then collected, washed with cold PBS, fixed in iced
70% ethanol at 4°C for 24 h, washed again with cold PBS and stained
in a propidium iodide (PI)/RNase A mixture at 37°C for 30 min in
the dark. Finally, the cells were analyzed using a
fluorescence-activated cell sorting (FACS) Caliber system
(FACSVerse, BD Biosciences).
RNA extraction, reverse transcription
(RT)-PCR and qPCR
Total RNA was purified using TRIzol reagent
(Takara). RT was performed using 5X Primescript RT Master Mix
(Takara) at 37°C for 15 min and at 85°C for 5 sec to denature the
enzymes. cDNA samples were analyzed by regular PCR (KT211, Tiangen
Biotech Co., Ltd.) or qPCR (SuperReal SYBR Green PreMix Plus,
FP205, Tiangen Biotech Co., Ltd.) with appropriate primers. The PCR
products were analyzed by electrophoresis in agarose gels. For
qPCR, 10 ng RNA was used for each test, and the reactions were
performed in triplicate on a Step One Plus thermocycler (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The data were
normalized against the levels of β-actin mRNA. The primers were as
follows: CDK12: Forward, AACAGCTAATGGAAGGACTGG and reverse,
CAGAGTTATAGAGCCGAGCAAG; PRX5: Forward,
CCCTCAGTGGAGGTATTTGAAG and reverse, CAGAGTTGAGAGAGGATGTTGG;
NSE: Forward, GAACTGGATGGGACTGAGAATAA and reverse,
CTTCAATGGAGACCACAGGATAG; β-actin: Forward,
GAGGTATCCTGACCCTGAAGTA and reverse: GCTCGAAGTCTAGAGCAACATAG.
Western blotting
Whole cell protein extracts were harvested with RIPA
lysis buffer (P0013B; Beyotime Institute of Biotechnology)
consisting of 50 mM Tris (pH 7.4), 150 mM NaCl, 1% Triton X-100, 1%
sodium deoxycholate, 0.1% SDS, sodium orthovanadate, sodium
fluoride, EDTA and leupeptin. For electrophoresis, 20 µg of each
protein extract was loaded onto denatured 12% polyacrylamide gel
and transferred to a PVDF membrane (EMD Millipore). The membrane
was incubated with 5% skimmed milk for 1 h at room temperature
followed by overnight incubation at 4°C with PRX5 (17724-1-AP), NSE
(55235-1-AP), SNAP25 (14093-1-AP) and β-actin (60008-1-lg) primary
antibodies (all from ProteinTech Group, Inc.). The abovementioned
primary antibodies were incubated at a dilution ratio of 1:1,000.
For SNAP23 (ProteinTech Group, Inc., 10825-1-AP), the dilution
ratio was 1:500. For NeuN detection, the primary antibody was
purchased from Cell Signaling Technology, Inc. (no. 12943) and the
dilution ratio was 1:1,000. On the following day, the membranes
were incubated at room temperature for 1 h with horseradish
peroxidase (HRP)-conjugated anti-mouse secondary antibody
(ProteinTech Group, Inc., SA00001-1), or HRP-conjugated anti-rabbit
secondary antibody (ProteinTech Group, Inc., SA00001-2). HRP
signals were detected by enhancing chemical luminol reagent
(ProteinTech Group, Inc.) on Amersham Imager 600 (GE
Healthcare).
Immunofluorescence
Cells were seeded into 6-well plates, fixed in
methanol for 10 min at room temperature, and then permeabilized in
0.5% TritonX-100 for 10 min at room temperature. The samples were
then washed in PBS before blocking overnight at 4°C with primary
antibodies against NSE, SNAP23 and SNAP25, which were the same with
the primary antibodies used in western blotting. The cells were
then incubated with Cy3-conjugated goat anti-rabbit antibody
(ProteinTech Group, Inc., SA00009-2) for 2 h at room temperature.
Prior to visualization, the cells were incubated with DAPI
(Invitrogen; Thermo Fisher Scientific, Inc.) for 5 min, washed with
PBS and then visualized under an EVOS microscope (Thermo Fisher
Scientific, Inc.).
Statistical analysis
Statistical significance was determined by t-test
with GraphPad Prism 7 (GraphPad Software, Inc.). Cell cycle data
analysis was performed by FlowJo.7 software (FlowJo LLC). For cell
cycle and immunofluorescence analysis, statistical significance was
determined by multiple t-tests. For morphology and immunostaining
analysis, cells were quantified in a blinded manner using ImageJ
8.0 software (National Institutes of Health, Bethesda, MD, USA). A
minimum of 5 randomly selected fields were quantified for the
assays. For western blot analysis, the quantification was performed
by ImageJ 8.0 software. In all cases, P<0.05 was considered to
indicate a statistically significant difference.
Results
Generation of a lentivirus expressing
miR-153
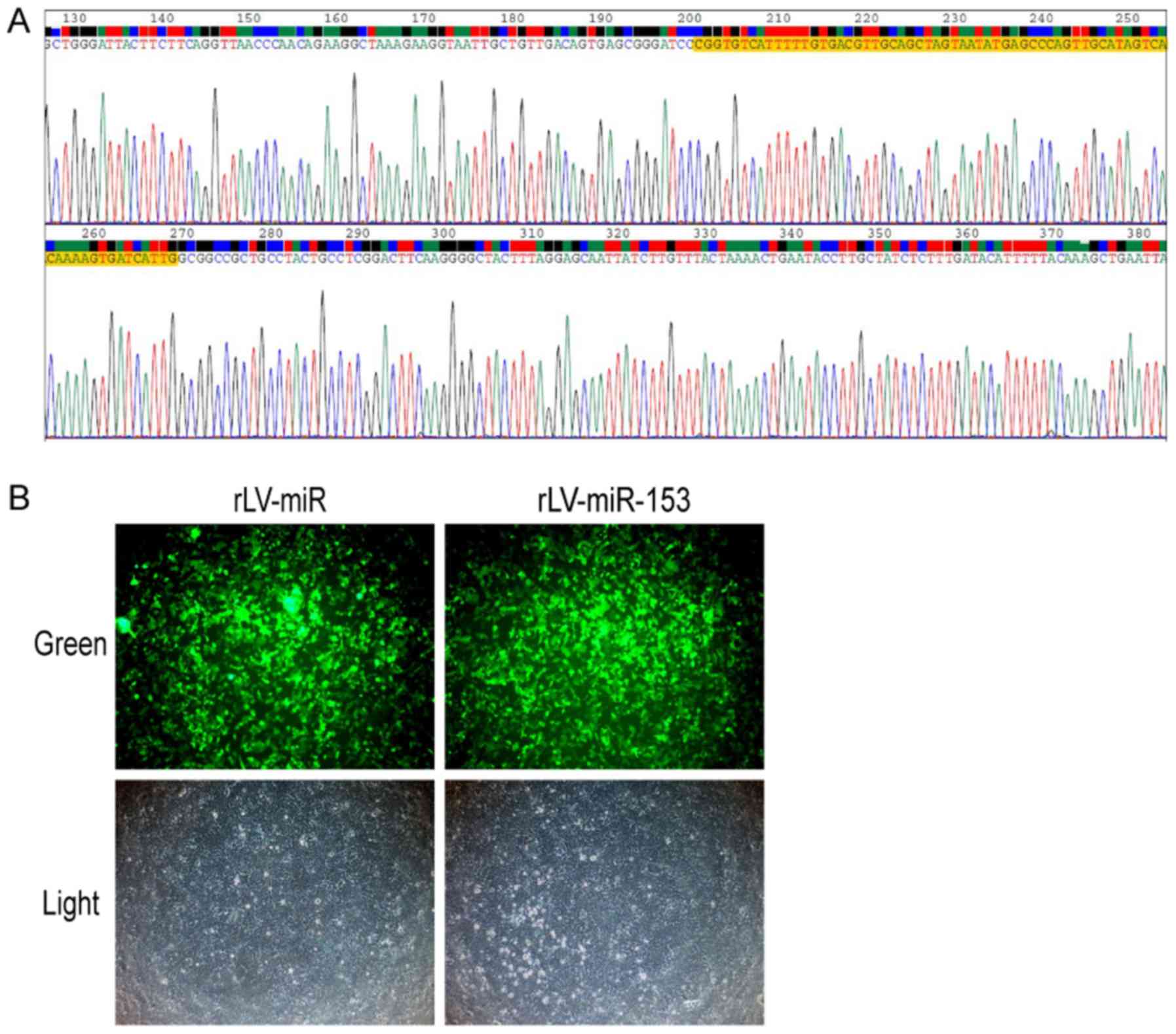
In order to observe the effects of miR-153, we first
generated a lentivirus expressing miR-153. The sequence of miR-153
was obtained from www.mirbase.org. The sequence of mouse stem-loop
precursor miR-153 (pre-miR153, MI0000175) was selected. The results
revealed that the sequence of pre-miR153 inserted in the lentivirus
plasmid was correct (Fig. 1A).
Moreover, a lentivirus expressing miR-153 (rLV-miR-153) was
produced using 293 cells (Fig.
1B). These observations suggested that the lentivirus
expressing rLV-miR-153 was successfully constructed.
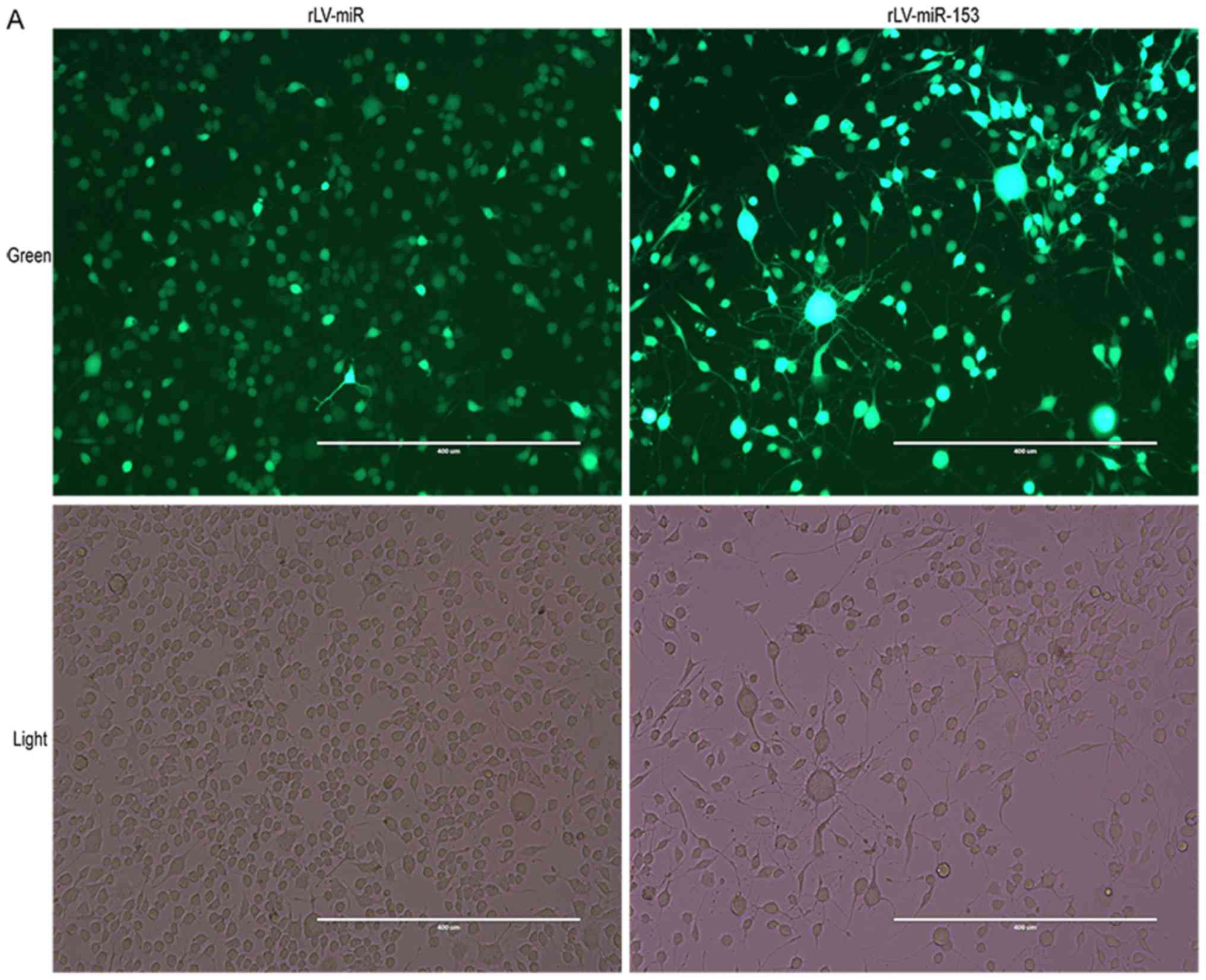
Establishment of
miR-153-overexpressing HT-22 stable cell lines
To assess the effect of miR-153 on neurons, stable
HT-22 cell lines with miR-153 overexpression were established.
First, HT-22 cells were infected by rLV-miR-153 or rLV-miR, and
then the positive monoclonal cells expressing GFP were selected for
culture (Fig. 2A). Next, total RNA
was extracted from the above mentioned two stable cell lines,
RT-PCR was performed, and miRNA was extracted for qPCR analysis. As
shown in Fig. 2B, pre-miR-153 was
significantly increased in the rLV-miR153 group compared with the
rLV-miR group. In particular, the qPCR results demonstrated that
both mature miR-153-3 prime and mature miR-153-5 prime (Fig. 2C) were significantly increased
compared with the rLV-miR group (miR-153-3 prime: 0.0316±0.0019 vs.
0.0015±0.0007, respectively, P=0.0007<0.01; and miR-153-5 prime:
0.0038±0.0007 vs. 0.0004±0.00036, respectively, P=0.0043). These
results suggested that miR-153-overexpressing HT-22 stable cell
lines were established.
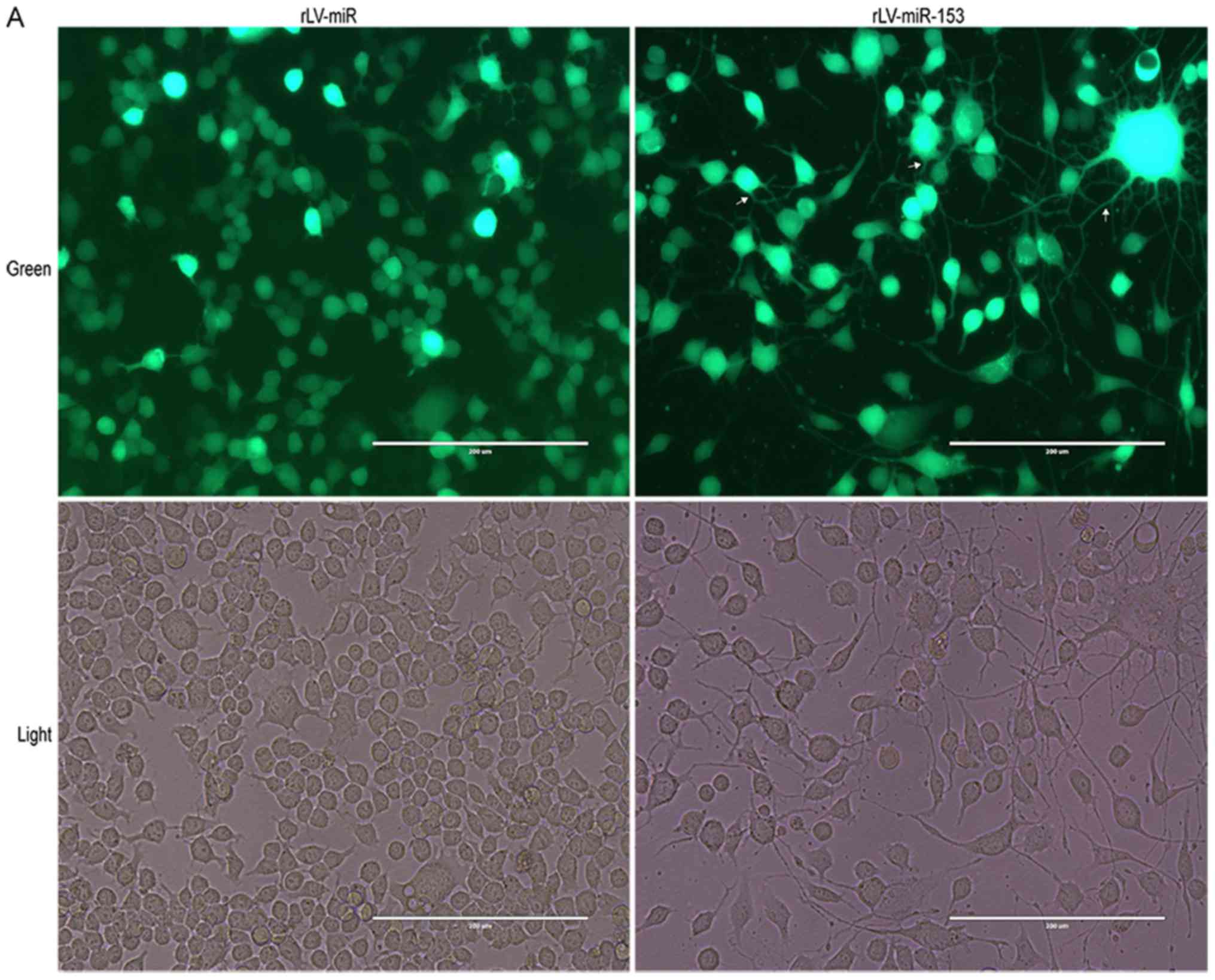
miR-153 induces neural differentiation
of HT-22 cells
To validate whether miR-153 affects the
differentiation of HT-22 cells, in vitro assays were
performed to assess cell differentiation and growth in HT-22 cells
infected with rLV-miR and rLV-miR-153. The results demonstrated
that HT-22 cell differentiation, with protrusions and branches, was
markedly enhanced in the rLV-miR-153 group compared with the
rLV-miR group (Fig. 3A). As shown
in Fig. 3B, the percentage of
differentiated cells was increased by ~2-fold in the rLV-miR-153
group compared with the rLV-miR group (62.80±6.55 vs. 29.26±8.17,
respectively, P=0.0079). The results confirmed that miR-153
promotes cell differentiation.
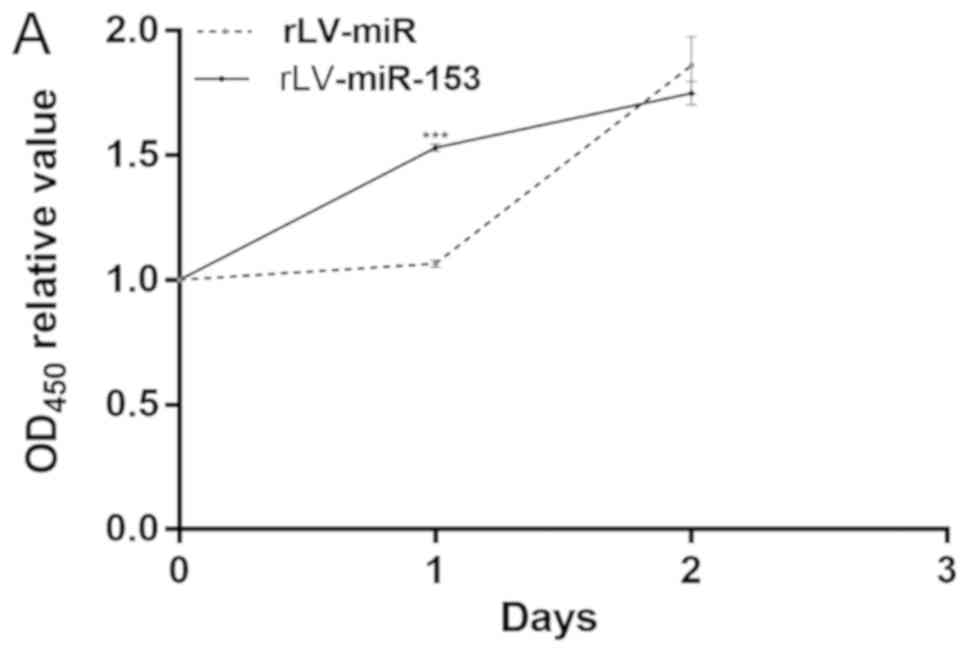
miR-153 attenuates the growth of HT-22
cells due to increased differentiation
Given that miR-153 induces differentiation of HT-22
cells, it was investigated whether the effect of miR-153 on cell
growth is dependent upon the increased differentiation. To support
our observations regarding the differentiation of HT-22 cells, a
cell proliferation assay was performed. First, cell proliferation
was measured with the CCK-8 assay in the two stable HT-22 cell
lines, namely the rLV-miR and rLV-miR-153 groups. As shown in
Fig. 4A, although the number of
HT-22 cells increased rapidly in the rLV-miR-153 group compared
with the rLV-miR group on the first day (1.5308±0.0151 vs.
1.0652±0.0138, P=0.0000245), the proliferation ability of the HT-22
cells was attenuated on the second day in the rLV-miR-153 group
compared with the rLV-miR group (1.7496±0.0466 vs. 1.8604±0.1149,
P=0.0533).
Subsequently, to identify whether miR-153 inhibits
cell growth by altering the cell cycle (G1, S and G2/M phase), FACS
was performed. As shown in Fig. 4B-a
to -c, the percentage of cells in the S phase was significantly
decreased in the rLV-miR-153 group compared with the rLV-miR group
(39.58±0.64 vs. 51.01±2.72, respectively, P=0.0063). The percentage
of cells in the G0/G1 phase was significantly increased in the
rLV-miR-153 group compared with the rLV-miR group (41.03±2.34 vs.
35.70±1.76, respectively, P=0.034). The percentage of cells in the
G2/M phase was significantly increased in the rLV-miR-153 group
compared with the rLV-miR group (13.94±2.32 vs. 7.79±0.31,
respectively, P=0.02).
Furthermore, qPCR analysis of several cell
cycle-related genes revealed that the expression of CDK12
was significantly decreased in the rLV-miR-153 group compared with
the rLV-miR group (Fig. 4C,
0.0288±0.0039 vs. 0.0538±0.0064, respectively, P=0.0294), and
miR-153 may affect the cell cycle by downregulating CDK12.
Taken together, these observations indicated that miR-153
attenuated the growth of HT-22 cells by altering the cell cycle,
which was associated with the process of cell differentiation.
miR-153 regulates neuron-specific
factors in HT-22 cells
To elucidate the mechanism through which miR-153
controls the differentiation of HT-22 cells, several neuron
differentiation-related markers were investigated. The RT-PCR
results demonstrated that the transcriptional level of PRX5
was significantly increased in the rLV-miR-153 group compared with
the rLV-miR group (0.9280±0.0905 vs. 0.3713±0.0940, respectively,
P=0.0018; Fig. 5A-a, upper panel,
and 5A-b). In addition, the
transcriptional level of NSE was significantly increased in
the rLV-miR-153 group compared with the rLV-miR group
(1.1030±0.1198 vs. 0.6333±0.0545, respectively, P=0.0035; Fig. 5A-a, middle panel, and 5A-b).
Moreover, the results of the western blot analysis demonstrated
that the protein expression level of PRX5, NSE, SNAP23, SNAP25 and
NeuN were significantly increased in the rLV-miR-153 group compared
with these levels in the rLV-miR group (Fig. 5B-a and -b).
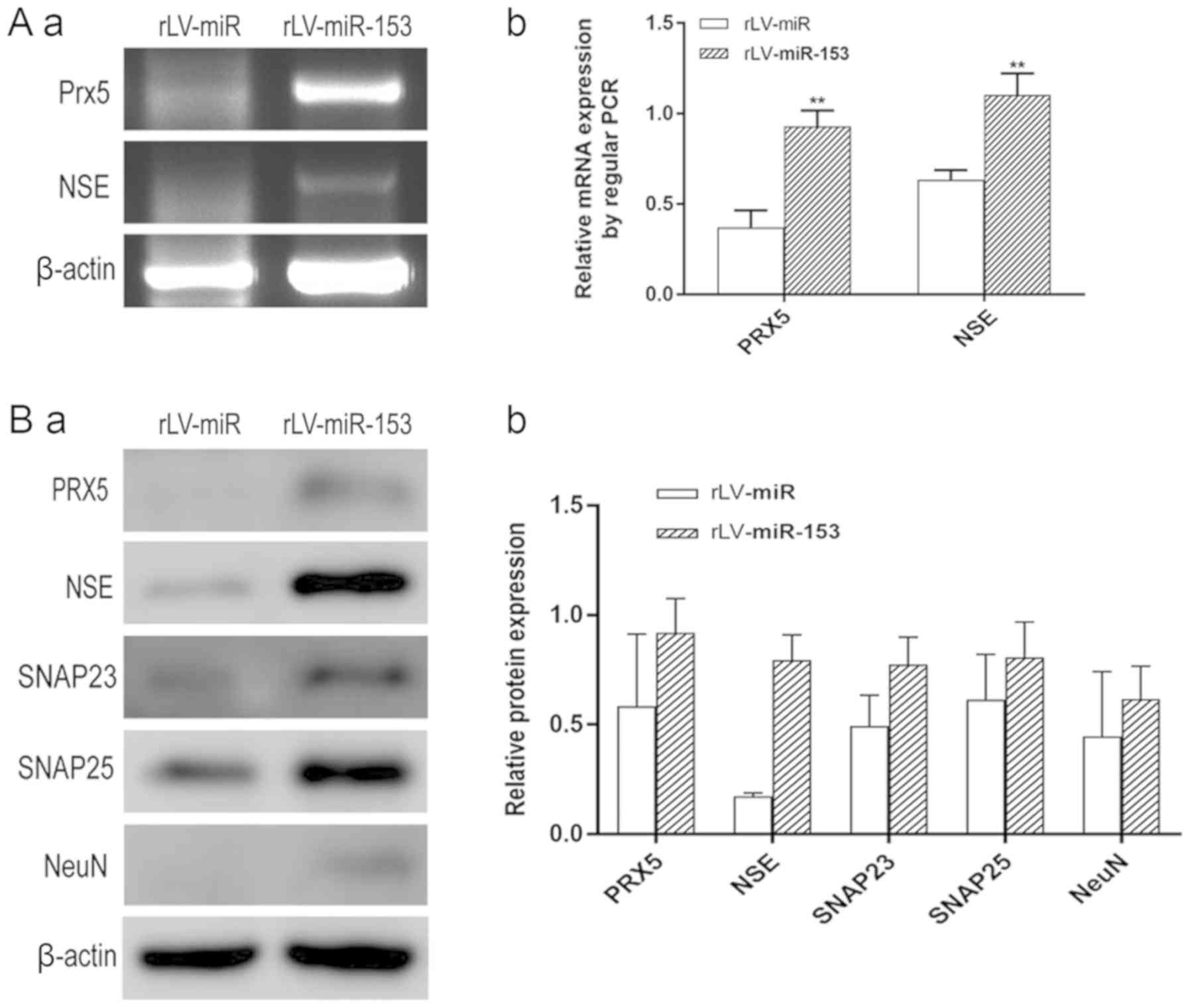 | Figure 5.Neuron-specific markers expressed in
rLV-miR and rLV-miR-153 cells. (A) Relative PRX5 and NSE expression
as detected by regular PCR (a). Significance was determined by
comparing the miR-153 overexpression group to the control with
unpaired t-test; n=3 (b). (B) The protein expression of PRX5, NSE,
SNAP23, SNAP25 and NeuN was analyzed by western blotting (a). Band
density was determined in the miR-153 overexpression group and
control, the error bars represent standard deviation (b). (C) The
protein expression of (a) NSE was analyzed by immunofluorescence
staining. Scale bar 400 µm. (C) The protein expression of (b)
SNAP23 and (c) SNAP25 was analyzed by immunofluorescence staining.
NSE (a), SNAP23 (b) and SNAP25 (c) were found to be significantly
increased in rLV-miR-153-overexpressing cells. Arrows indicate
positively stained cells. Significance was determined by comparing
NSE, SNAP23 and SNAP25 protein expression in the
rLV-miR-153-overexpressing group to control with multiple unpaired
t-tests; n=5. Error bars represent standard deviation. **P<0.01.
Cells exhibiting a strong orange color were identified as positive
cells expressing NSE, SNAP23 and SNAP25 (arrows). (C-d) Five visual
fields of each cell group were examined and the positive percentage
of cells was calculated. NSE, neuron-specific enolase; PRX5,
peroxiredoxin 5; SNAP, N-ethylmaleimide-sensitive fusion attachment
protein; NeuN, neuronal nuclei. |
Furthermore, immunofluorescence staining was also
performed to detect the protein expression of NSE, SNAP23 and
SNAP25. The number of cells expressing NSE was markedly higher in
the rLV-miR-153 group compared with that in the rLV-miR group
(Fig. 5C-a) and the positive rate
of NSE expression was significantly increased in the rLV-miR-153
group compared with the rLV-miR group (0.2816±0.0623 vs.
0.1311±0.0526, respectively, P=0.0075; Fig. 5C-d). Similarly, the positive rate
of SNAP23 expression was significantly increased in the rLV-miR-153
group compared with the rLV-miR group (0.4254±0.0404 vs.
0.1123±0.0142, respectively, P=8х10−6; Fig. 5C-b and -d). In addition, the
positive rate of SNAP25 expression was also significantly increased
in the rLV-miR-153 group compared with the rLV-miR group
(0.5181±0.1571 vs. 0.2392±0.0815, respectively,
P=3х10−5; Fig. 5C-c and
-d). Collectively, these results suggest that miR-153 promoted
the expression of neuron-specific factors, including PRX5, NSE,
SNAP23, SNAP25 and NeuN.
Discussion
A number of studies have investigated the emerging
role of miRNAs in nervous system growth, development and
pathogenesis of neurodegenerative diseases. Typical
neurodegenerative diseases, such as Alzheimer's disease,
Parkinson's disease, amyotrophic lateral sclerosis and Huntington's
disease, are the most extensively investigated and indicate a
potential role of miRNAs in neuronal development and function
(15–17). miRNA profile comparison and
individual miRNA studies have demonstrated that the brain has a
specific miRNA expression profile, and each miRNA may perform a
specific function to maintain brain integrity (18–20).
The results of the present study indicated that
miR-153 promotes the neuronal differentiation of HT-22 cells, which
is reflected by the morphological changes, such as the increasing
number of protrusions and branches, and the upregulation of NSE,
NeuN, PRX5, SNAP23 and SNAP25 expression. Mouse hippocampal HT-22
cells are a sub-line cloned from HT4 cells that has been grown
without establishing synaptic connections (21); thus, there were no obvious
protrusions and branches in HT-22 cells. The change in morphology
was likely due to the effects of miR-153 overexpression on cellular
growth and differentiation. Similarly, another study reported that
miR-153 inhibited astrogliogenesis of neural stem/progenitor cells,
while the neuronal differentiation of tertiary neurospheres was
increased significantly by miR-153 (11). These findings strongly support the
association of miR-153 with neuronal differentiation.
Moreover, we observed that the effect of miR-153 on
cell proliferation was associated with cell cycle changes and cell
differentiation. Our data indicated that the rapid growth on day 1
in the rLV-miR-153 group may have been due to the increasing
percentage of cells in the G2/M phase and the decreasing percentage
of cells in the S phase, which resulted in the slow growth of cells
in the rLV-miR-153 group on day 2. Of note, we found that miR-153
decreased the expression of CDK12. CDK12 can promote
mammalian cell proliferation, including the murine brain, liver and
H1299 cancer cells, by regulating pre-replicative complex assembly
(22). Mouse CDK12 is a
multifunctional protein that was reported to maintain genomic
stability and the pluripotency of murine embryonic stem cells
(23). Chen et al (24), reported that neural progenitor
cells of mice with CDK12 mutation accumulated at the G2/M phase
with neuron misalignment, which demonstrated that CDK12 is crucial
for cell proliferation. Notably, the decreased number of S phase
cells and increased number of G2/M phase cells were associated with
decreased CDK12 mRNA in the miR-153 overexpression group.
This may suggest that miR-153 promoted neuron differentiation and
synchronously delayed cell proliferation. Therefore, it may be
hypothesized that miR-153 has different functions at different
stages. In addition, different cell types may respond differently
to miR-153 modulation. Further studies are required to fully
elucidate the effect of miR-153 on neural cell cycle and related
genes.
In particular, the findings of the present study
indicated that miR-153 regulated the expression of neuron-specific
genes. For example, miR-153 increased the expression of NSE at the
transcriptional and protein level in HT-22 cells. γ-enolase is one
isozyme of three glycolytic enolases, which is neuron-specific and
used as a marker for all types of neurons. The appearance of
γ-enolase is a late event during neural differentiation, making it
a useful index of neural maturation. The remaining two enolases are
α-enolase, which is ubiquitous, and β-enolase, which is
muscle-specific (25). HT-22 cells
are not mature neural cells; therefore, the upregulation of NSE
after miR-153 overexpression indicates that further neural
differentiation and maturation occurred in HT-22 cells. The
upregulation of enolase may be a secondary effect of miR-153. As
this study is a preliminary research, the association between
miR-153 and enolase will be further explored in the following
study. In addition, NeuN is a neuronal nuclear antigen that is
commonly used as a biomarker for neurons, particularly mature
neurons. In the present study, the upregulation of the NeuN
protein, as demonstrated by western blot analysis, further
confirmed the neural differentiation of HT-22 cells induced by
miR-153 overexpression. Importantly, miR-153 also enhanced the
expression of PRX5 in HT-22 cells. PRX5 was previously reported to
protect neural cells from AβO damage (13); PRX5 may also play an important role
during miR-153-induced neural differentiation.
Intriguingly, bioinformatics prediction demonstrated
that miR-153 targets the 3′ untranslated region (UTP) of SNAP25,
Chunyao et al demonstrated that increased miR-153 levels
caused decreased SNAP25 expression resulting in movement defects in
zebrafish embryos, and SNAP25 was identified as the target of
miR-153 (10). However, a unique
aspect of our study was that both SNAP23 and SNAP25 were increased
in miR-153-overexpressing HT-22 cells. SNAP23 and SNAP25 have been
found to modulate exocytosis and neuronal development (12). Indeed, other miRNAs also exhibit
such inconsistencies. For example, Nr2e1 mRNA has a miR-9
responsive element in its 3′ UTR. Nr2e1 protein expression was
found to be reduced in mouse embryos with miR-9 mutation, and miR-9
did not suppress luciferase expression from a reporter conjugated
to the 3′ UTR of Nr2e1 in P19 cells. Moreover, miR-9-2 upregulated
the expression of endogenous Nr2e1 in P19 cells (8). This discrepancy between an miRNA and
the expression of its target genes may have some possible
explanations. On one hand, this discrepancy may be due to
differences in cell type, differentiation stage, animal species or
RNA-binding protein repertoires. Some RNA-binding proteins that
facilitate binding of miR-153 to its target may be missing or
insufficient in HT-22 cells. On the other hand, miR-153 may
increase SNAP25 expression indirectly by suppressing another gene
that inhibits SNAP25 expression. Further studies are required to
elucidate the precise mechanism underlying the regulation of SNAP25
and SNAP23 by miR-153. In addition, SNAP25 misregulation was found
to play a key role in some human diseases including attention
deficit-hyperactivity disorder, schizophrenia, bipolar I disorder,
Huntington's disease and Alzheimer's disease (26–29).
Therefore, the upregulation of SNAP25 by miR-153 in the present
study may be a potential therapy target for some neurodegenerative
diseases.
Collectively, these results on miRNA expression in
neural cells indicate that miR-153 may function differently in
different regions of the central nervous system, or during
different developmental stages. The HT-22 cell line was used to
analyze the effects of miR-153 in the present study, and the
results may differ in embryonic or animal models in response to
miR-153 overexpression.
Although our study demonstrated that miR-153 is
implicated in neuronal differentiation and normal function by
regulating multiple proteins, a series of issues regarding miR-153
controlling neurogenesis should be further explored. First, the
exact role of miR-153 in neural development must be confirmed in
vivo. Second, the detailed molecular mechanism underlying the
function of miR-153 in neurogenesis requires further investigation.
Third, the clinical applicability of miR-153 must be investigated,
particularly in neurodegenerative diseases.
Acknowledgements
Not applicable.
Funding
The present study received funding from ‘Outstanding
Leaders Training Program of Pudong Health Bureau of Shanghai’
(grant no. PWRI2016-02).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YZ and JL designed the study. SJ and JX prepared the
reagents and analyzed data. CW and QM cultured the cells and
drafted the manuscript. CX and JL performed the molecular biology
experiments. CX, YG, QW, WX, YQ and YH analyzed and interpreted the
data, and revised the manuscript for important intellectual
content. CX and JL wrote the manuscript. YZ and JL reviewed and
revised the document. All authors reviewed and approved the final
version of the manuscript. All authors read and approved the
manuscript and agree to be accountable for all aspects of the
research in ensuring that the accuracy or integrity of any part of
the work are appropriately investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Choi PS, Zakhary L, Choi WY, Caron S,
Alvarez-Saavedra E, Miska EA, McManus M, Harfe B, Giraldez AJ,
Horvitz RH, et al: Members of the miRNA-200 family regulate
olfactory neurogenesis. Neuron. 57:41–55. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Makeyev EV, Zhang J, Carrasco MA and
Maniatis T: The MicroRNA miR-124 promotes neuronal differentiation
by triggering brain-specific alternative pre-mRNA splicing. Mol
Cell. 27:435–448. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim J, Inoue K, Ishii J, Vanti WB, Voronov
SV, Murchison E, Hannon G and Abeliovich A: A MicroRNA feedback
circuit in midbrain dopamine neurons. Science. 317:1220–1224. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Haramati S, Chapnik E, Sztainberg Y, Eilam
R, Zwang R, Gershoni N, McGlinn E, Heiser PW, Wills AM, Wirguin I,
et al: miRNA malfunction causes spinal motor neuron disease. Proc
Natl Acad Sci USA. 107:13111–13116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Harraz MM, Dawson TM and Dawson VL:
MicroRNAs in Parkinson's disease. J Chem Neuroanat. 42:127–130.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Morgado AL, Rodrigues CM and Sola S:
MicroRNA-145 regulates neural stem cell differentiation through the
Sox2-Lin28/let-7 signaling pathway. Stem Cells. 34:1386–1395. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Cheng LC, Pastrana E, Tavazoie M and
Doetsch F: miR-124 regulates adult neurogenesis in the
subventricular zone stem cell niche. Nat Neurosci. 12:399–408.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shibata M, Nakao H, Kiyonari H, Abe T and
Aizawa S: MicroRNA-9 regulates neurogenesis in mouse telencephalon
by targeting multiple transcription factors. J Neurosci.
31:3407–3422. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Doxakis E: Post-transcriptional regulation
of alpha-synuclein expression by mir-7 and mir-153. J Biol Chem.
285:12726–12734. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wei C, Thatcher EJ, Olena AF, Cha DJ,
Perdigoto AL, Marshall AF, Carter BD, Broadie K and Patton JG:
miR-153 regulates SNAP-25, synaptic transmission, and neuronal
development. PLoS One. 8:e570802013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tsuyama J, Bunt J, Richards LJ, Iwanari H,
Mochizuki Y, Hamakubo T, Shimazaki T and Okano H: MicroRNA-153
regulates the acquisition of gliogenic competence by neural stem
cells. Stem Cell Reports. 5:365–377. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zylbersztejn K and Galli T: Vesicular
traffic in cell navigation. FEBS J. 278:4497–4505. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim B, Park J, Chang KT and Lee DS:
Peroxiredoxin 5 prevents amyloid-beta oligomer-induced neuronal
cell death by inhibiting ERK-Drp1-mediated mitochondrial
fragmentation. Free Radic Biol Med. 90:184–194. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O'Brien RJ and Wong PC: Amyloid precursor
protein processing and Alzheimer's disease. Annu Rev Neurosci.
34:185–204. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Qiu L, Tan EK and Zeng L: microRNAs and
neurodegenerative diseases. Adv Exp Med Biol. 888:85–105. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leggio L, Vivarelli S, L'Episcopo F,
Tirolo C, Caniglia S, Testa N, Marchetti B and Iraci N: microRNAs
in Parkinson's disease: From pathogenesis to novel diagnostic and
therapeutic approaches. Int J Mol Sci. 18:E26982017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Sempere LF, Freemantle S, Pitha-Rowe I,
Moss E, Dmitrovsky E and Ambros V: Expression profiling of
mammalian microRNAs uncovers a subset of brain-expressed microRNAs
with possible roles in murine and human neuronal differentiation.
Genome Biol. 5:R132004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bak M, Silahtaroglu A, Moller M,
Christensen M, Rath MF, Skryabin B, Tommerup N and Kauppinen S:
MicroRNA expression in the adult mouse central nervous system. RNA.
14:432–444. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
et al: A mammalian microRNA expression atlas based on small RNA
library sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu J, Li L and Suo WZ: HT22 hippocampal
neuronal cell line possesses functional cholinergic properties.
Life Sci. 84:267–271. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lei T, Zhang P, Zhang X, Xiao X, Zhang J,
Qiu T, Dai Q, Zhang Y, Min L, Li Q, et al: Cyclin K regulates
prereplicative complex assembly to promote mammalian cell
proliferation. Nat Commun. 9:18762018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Juan HC, Lin Y, Chen HR and Fann MJ: Cdk12
is essential for embryonic development and the maintenance of
genomic stability. Cell Death Differ. 23:1038–1048. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chen HR, Juan HC, Wong YH, Tsai JW and
Fann MJ: Cdk12 regulates neurogenesis and late-arising neuronal
migration in the developing cerebral cortex. Cereb Cortex.
27:2289–2302. 2017.PubMed/NCBI
|
|
25
|
Isgro MA, Bottoni P and Scatena R:
Neuron-specific enolase as a biomarker: Biochemical and clinical
aspects. Adv Exp Med Biol. 867:125–143. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Corradini I, Verderio C, Sala M, Wilson MC
and Matteoli M: SNAP-25 in neuropsychiatric disorders. Ann N Y Acad
Sci. 1152:93–99. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scarr E, Gray L, Keriakous D, Robinson PJ
and Dean B: Increased levels of SNAP-25 and synaptophysin in the
dorsolateral prefrontal cortex in bipolar I disorder. Bipolar
Disord. 8:133–143. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Smith R, Klein P, Koc-Schmitz Y, Waldvogel
HJ, Faull RL, Brundin P, Plomann M and Li JY: Loss of SNAP-25 and
rabphilin 3a in sensory-motor cortex in Huntington's disease. J
Neurochem. 103:115–123. 2007.PubMed/NCBI
|
|
29
|
Bereczki E, Francis PT, Howlett D, Pereira
JB, Hoglund K, Bogstedt A, Cedazo-Minguez A, Baek JH, Hortobagyi T,
Attems J, et al: Synaptic proteins predict cognitive decline in
Alzheimer's disease and Lewy body dementia. Alzheimers Dement.
12:1149–1158. 2016. View Article : Google Scholar : PubMed/NCBI
|