Introduction
Colorectal cancer (CRC) is a frequent malignant
tumor in developed countries, with high morbidity and a high death
rate. According to statistics published in 2015, CRC is the third
most frequently diagnosed cancer in males and ranks second in
females (1). According to the
American Joint Committee on Cancer, the 5-year overall survival
(OS) rate of CRC is 65.2% (2).
Common risk factors for CRC include age, obesity, smoking and an
unhealthy diet (3). Previous
studies have found that several patients present with unresectable
stage disease at initial diagnosis (4,5).
Standard treatments for CRC include surgical resection, modern
chemotherapy and radiation therapy. Although chemotherapy plays an
important role in CRC treatment, the chemoresistance-reduced
effectiveness of anti-neoplastic agents causes a lower survival
rate of advanced-stage CRC patients.
Oxaliplatin (OxPt) is a third-generation
platinum-based chemotherapeutic drug that inhibits the DNA
replication and transcription of tumor cells (6). When used in combination with
fluorouracil (5-FU), leucovorin and folinic acid, it is recognized
as the first-line chemotherapy strategy for CRC (7). In recent years, the extensive use of
OxPt has caused increased chemoresistance in clinical practice.
Previous studies have found that the nucleotide excision repair
pathway plays a key role in OxPt resistance (8,9).
Moreover, the overexpression of chemoresistance-associated proteins
(e.g., transforming growth factor-β and WBSCR22) and microRNAs, and
increased messenger RNA levels of XPAC and ERCC1, are believed to
predict chemoresistance to OxPt (10–12).
Irinotecan, an analogue of camptothecin isolated
from the Chinese tree Camptotheca acuminata, was first
approved by the Food and Drug Administration for the treatment of
CRC in 1996 and is now used as the second choice for CRC
chemotherapy (13). Irinotecan
selectively inhibits topoisomerase I and induces the degradation of
double-stranded DNA via intracellular modifications. Ultimately,
irinotecan is activated by carboxylesterases to its active
metabolite SN-38, which combines with topoisomerase and induces the
breakage of DNA strands, DNA replication failure and cellular
apoptosis (14,15). The therapeutic efficacy and
toxicity of irinotecan are related to the expression of
tumor-specific genes and the intra-tumor accumulation of SN-38
(16). Irinotecan treatment may
also cause severe chemoresistance via variable levels of metabolic
enzymes, reduced cellular accumulation from active drug efflux and
reduced levels of Topo I expression, as well as different mutations
and/or the activation of nuclear factor κB (NF-κB) (17).
Long non-coding RNAs (lncRNAs), defined as mRNA-like
transcripts with lengths up to 200 nucleotides, are important
members of the non-coding RNA family (18). lncRNA expression is regulated by
both transcriptional and epigenetic factors. Several studies have
revealed that lncRNAs are expressed in various tumor tissues and
are involved in a number of cellular functions and developmental
processes, such as cell growth, development, invasion, and
apoptosis (19–21). Increasing lines of evidence suggest
that lncRNAs are valuable bio-targets in the diagnosis and
treatment of CRC (22). Recent
studies have suggested that lncRNAs play a key role in drug
function regulation and chemoresistance through various mechanisms
in multiple cancers (23,24). At present, at least 70 CRC-related
lncRNAs have been recognized, including HOTAIR, CCAT1, CCAT2,
MALAT-1 and H19 (25). Changes in
the expression of these lncRNAs could lead to chemotherapy and
radiotherapy resistance. Therefore, further research and efforts
are needed to clarify the chemoresistance mechanism of each lncRNA.
Then a simple and reliable screening program based on the
expression levels of lncRNAs can be developed to guide the
selection of chemotherapy drugs.
In the current study, previously published RNA
expression datasets on the chemoresistance of CRC in the Gene
Expression Omnibus (GEO) database were searched. A comprehensive
bioinformatics analysis was performed by defining the
differentially expressed lncRNAs and other RNAs separately
according to their chemoresistance to irinotecan or OxPt. The
differentially expressed lncRNAs were correlated with
differentially expressed genes (DEGs) and potential diseases based
on the RNA-associated interactions database (RAID) and the
mammalian lncRNA-disease repository (MNDR) database. Gene Ontology
(GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway
enrichment analyses were also performed and a protein-protein
interaction (PPI) network was established to screen for crucial
genes and lncRNAs, whose hazard degrees were further demonstrated
by evaluating their expression in tumors and according to tumor,
node and metastasis (TNM) stage, as well as survival.
Materials and methods
Datasets and DEG identification
Gene expression profiles for chemoresistance to two
chemotherapeutic drugs, OxPt and irinotecan (active metabolite
SN-38), in three CRC cell lines (HCT116, HT29, and LoVo) were
obtained from GSE42387 in the GEO database (26). Gene expression data were explored
and visualized in the ggplot2 package of R software (version 3.2.0,
http://ggplot2.tidyverse.org) for each
sample in the nine groups (normal HCT116, OxPt-resistant HCT116,
SN-38-resistant HCT116, normal HT29, OxPt-resistant HT29,
SN-38-resistant HT29, normal LoVo, OxPt-resistant LoVo and
SN-38-resistant LoVo). A detailed workflow of the data analysis is
shown in Fig. S1. The limma
package, which includes lmFit, eBayes and top Table functions, was
used for the pairwise comparison of DEGs among the nine groups
(27). The cutoff criteria were
P<0.05 and abs(log2FC)>1, where FC indicates
fold-change. The screened DEGs were divided into two groups,
differentially expressed mRNAs and lncRNAs, which were further
analyzed.
GO annotation and KEGG pathway
enrichment analyses of DEGs
As mentioned above, two groups of DEGs: mRNAs and
lncRNAs were analyzed. Based on RAID version 2.0 (28), the potential targets (confidence
score >0.5) of the differentially expressed lncRNAs were
obtained and intersected with DEGs found in the same group. Next,
the intersected DEGs were divided into two groups: OxPt resistance
and irinotecan resistance. GO annotation is a classic method used
to identify the biological attributes of DEGs. This analysis
comprises three parts: Biological process (BP), cell component (CC)
and molecular function (MF). KEGG, a collection of genome,
biological pathway, disease, drug and chemical substance databases,
was used to identify the functional attributes of the DEGs. The
Database for Annotation, Visualization and Integrated Discovery
(DAVID; ver. 6.8) (29,30) was also applied for the functional
interpretation of the two large lists of genes derived from
previous analyses. Statistical significance was set at
P<0.05.
PPI network construction and hub gene
identification
A PPI network was constructed for the DEGs using
STRING (31), an online functional
protein association network tool, to detect potential relationships
among the DEGs with confidence scores ≥0.4 and a maximum number of
interactors of 1. The generated PPI network data were downloaded
and imported into Cytoscape software (ver. 3.5.1), a bioinformatics
tool used to create networks of protein interactions and reassess
and integrate gene information from numerous embedded applications.
The CentiScaPe plugin was used to determine the characteristics of
each node in the PPI network, which gave each gene a degree score,
the simplest topological index, allowing for immediate evaluation
of the average number of edges (interactions) incident to the node
(32). Genes with a degree ≥5 in
significantly perturbed networks were defined as hub genes (mRNA).
The differentially expressed lncRNA-mRNA interaction network was
constructed and displayed by Cytoscape (version 3.5, http://www.cytoscape.org/). The lncRNA holding
interactions with the greatest number of hub genes (mRNA) in the
network were defined as hub lncRNAs.
Constructing the differentially
expressed lncRNA-related disease network
Each lncRNA has a disease association profile.
According to MNDR version 2.0 (33), a global view of the lncRNA-mediated
disease network, which contains all of the differentially expressed
lncRNA-related disease data (confidence score >0.5), was
downloaded and integrated by R software. The disease network was
constructed by Cytoscape. The disease label size was normalized to
the number of lncRNAs related to each disease.
Hub lncRNA analysis
The hub lncRNAs were further investigated using Gene
Expression Profiling Interactive Analysis, a newly developed
interactive web server for analyzing the RNA sequencing expression
data of 9,736 tumors and 8,587 normal samples from The Cancer
Genome Atlas (TCGA) and Genotype-Tissue Expression projects
(34). The expression of each hub
lncRNA in colon adenocarcinoma and rectum adenocarcinoma was
compared with in normal tissue and to different tumor stages and
plotted. Survival analysis was performed based on information from
TCGA database. Relapse-free survival and OS were calculated. Hazard
ratios with 95% confidence intervals and P-values calculated based
on the Mantel-Cox log-rank test are displayed in Kaplan-Meier
plots.
Results
Data quality control and DEG
screening
A total of 32,705 genes were detected in each
sample; their expression values and data distributions were similar
between groups (Fig. S2A) and
among samples (Fig. S2B).
Principal component analysis showed that samples were easily
grouped in different cell lines but not into certain drug
resistance patterns (Fig. S2C).
Therefore, further bioinformatics analysis should be performed
separately based on different cell lines and chemoresistance. This
dataset qualified for the following analysis.
Following data processing using limma, 255 DEGs in
HCT116-OxPt vs. HCT116, 672 DEGs in HT29-OxPt vs. HT29, 491 DEGs in
LoVo-OxPt vs. LoVo, 283 DEGs in HCT116-SN-38 vs. HCT116, 779 DEGs
in HT29-SN-38 vs. HT29 and 1,713 DEGs in LoVo-SN-38 vs. LoVo were
identified (Fig. S2D and E). The
detailed DEGs information were shown in Tables SI–SVI.
Intersection between the predicted
targets of lncRNAs and DEGs
Among all of the DEGs identified in each comparison,
the differentially expressed lncRNAs were isolated and are listed
in Table I. The target prediction
of each lncRNA was searched and obtained using RAID version 2.0.
Next, the target genes were intersected with the identified DEGs in
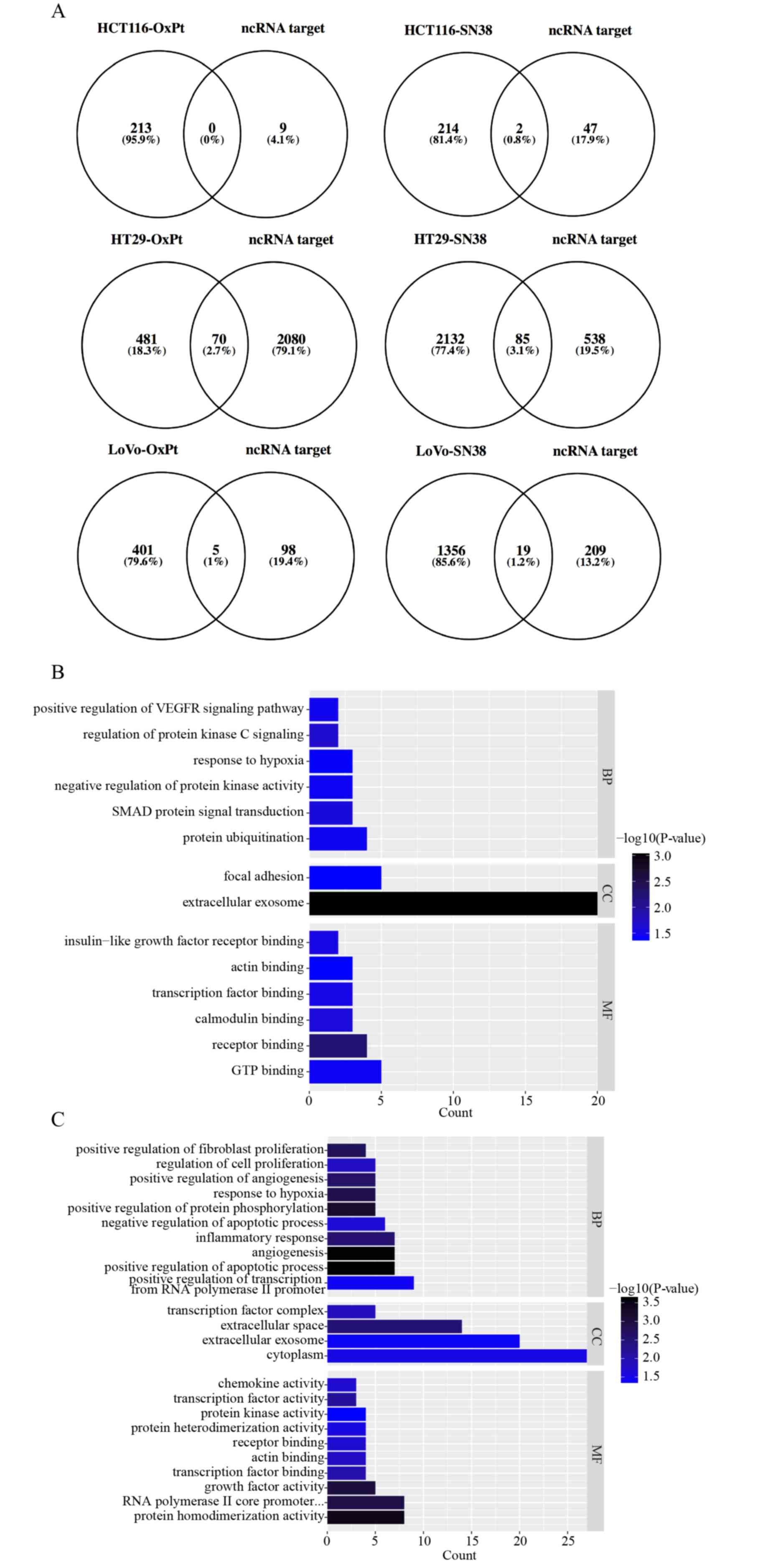
each comparison and are displayed as Venn diagrams (Fig. 1A). The intersected DEGs in the same
drug management were united to perform GO annotation and KEGG
pathway enrichment analyses based on DAVID. All significant
(P<0.05) enriched entries in OxPt resistance are shown in a
histogram (Fig. 1B), with the top
three being protein ubiquitination, SMAD protein signal
transduction and negative regulation of protein kinase activity in
the BP GO annotation category; extracellular exosome and focal
adhesion in the CC category; and GTP binding, receptor binding, and
calmodulin binding in the MF category. All significant (P<0.05)
enriched entries in irinotecan resistance are also shown in a
histogram (Fig. 1C and D), with
the top three being positive regulation of transcription from RNA
polymerase II promoter, positive regulation of apoptotic process,
angiogenesis in the BP GO annotation category; cytoplasm,
extracellular exosome, and extracellular space in the CC category;
protein homodimerization activity, RNA polymerase II core promoter
proximal region sequence-specific DNA binding, and growth factor
activity in the MF category; and pathways in cancer, microRNAs in
cancer, and TNF signaling pathway in the KEGG pathway. Detailed
information on the DEGs involved in the BP GO category and KEGG
pathway are listed in Fig. 1E.
 | Table I.Drug resistance-related
differentially expressed non-coding RNAs. |
Table I.
Drug resistance-related
differentially expressed non-coding RNAs.
| Group | ID | Symbol | logFC | AveExpr | P-value | Change | Gene name |
|---|
| HCT116-OxPt | 285735 | LINC00326 | 1.46 | 7.31 |
2.1×10−05 | UP | Long intergenic
non-protein coding RNA 326 |
|
| 284739 | LINC00176 | 1.11 | 8.93 |
1.3×10−02 | UP | Long intergenic
non-protein coding RNA 176 |
| HT29-OxPt | 283460 | HNF1A-AS1 | −1.72 | 7.65 |
5.0×10−05 | DOWN | HNF1A antisense RNA
1 |
|
| 643911 | CRNDEa | −1.97 | 10.82 |
1.4×10−06 | DOWN | Colorectal
neoplasia differentially expressed |
| LoVo-OxPt | 283120 | H19a | 1.00 | 7.06 |
5.4×10−04 | UP | H19, imprinted
maternally expressed transcript |
|
| 64150 | DIO3OS | −1.79 | 8.51 |
2.5×10−06 | DOWN | DIO3 opposite
strand/antisense RNA |
|
| 196047 | EMX2OS | −1.46 | 7.87 |
2.5×10−03 | DOWN | EMX2 opposite
strand/antisense RNA |
| HCT116-SN38 | 652995 | UCA1a | −1.40 | 12.29 |
4.4×10−06 | DOWN | Urothelial cancer
associated 1 |
| HT29-SN38 | 441951 | ZNFX1-AS1 | 1.35 | 10.22 |
9.2×10−05 | UP | ZNFX1 antisense RNA
1 |
|
| 727701 | LINC00165 | −1.03 | 7.43 |
5.5×10−05 | DOWN | Long intergenic
non-protein coding RNA 165 |
|
| 100169750 | PRINS | −1.50 | 9.81 |
6.8×10−06 | DOWN | Psoriasis
associated RNA induced by stress |
|
| 283460 | HNF1A-AS1 | −1.62 | 7.70 |
6.6×10−05 | DOWN | HNF1A antisense RNA
1 |
|
| 554202 | MIR31HG | −1.49 | 8.07 |
1.3×10−05 | DOWN | MIR31 host
gene |
|
| 283131 | NEAT1 | 1.32 | 11.22 |
9.3×10−04 | UP | Nuclear paraspeckle
assembly transcript 1 |
|
| 100289410 | MCF2L-AS1 | −1.30 | 9.95 |
2.8×10−06 | DOWN | MCF2L antisense RNA
1 |
|
| 643911 | CRNDEa | −1.17 | 11.21 |
2.9×10−05 | DOWN | Colorectal
neoplasia differentially expressed |
| LoVo-SN38 | 100093630 | SNHG8 | −1.49 | 9.80 |
1.3×10−03 | DOWN | Small nucleolar RNA
host gene 8 |
|
| 253264 | ZNF503-AS1 | −2.28 | 7.90 |
8.6×10−07 | DOWN | ZNF503 antisense
RNA 1 |
|
| 60674 | GAS5 | −1.21 | 15.21 |
2.1×10−02 | DOWN | Growth
arrest-specific 5 |
|
| 64150 | DIO3OS | 1.07 | 9.94 |
4.0×10−05 | UP | DIO3 opposite
strand/antisense RNA |
|
| 196047 | EMX2OS | −1.10 | 8.05 |
8.5×10−03 | DOWN | EMX2 opposite
strand/antisense RNA |
|
| 128439 | SNHG11 | −1.39 | 10.12 |
4.7×10−04 | DOWN | Small nucleolar RNA
host gene 11 |
|
| 100124700 | HOTAIRa | 1.58 | 7.21 |
3.1×10−06 | UP | HOX transcript
antisense RNA |
|
| 114915 | EPB41L4A-AS1 | −1.42 | 15.27 |
3.0×10−02 | DOWN | EPB41L4A antisense
RNA 1 |
|
| 652995 | UCA1a | −1.45 | 7.58 |
1.6×10−04 | DOWN | Urothelial cancer
associated 1 |
|
| 401237 | LINC00340 | 1.99 | 7.58 |
1.7×10−06 | UP | Long intergenic
non-protein coding RNA 340 |
|
| 29092 | LINC00339 | −1.55 | 9.20 |
4.0×10−03 | DOWN | Long intergenic
non-protein coding RNA 339 |
|
| 79992 | AGPAT4-IT1 | 1.50 | 8.13 |
2.5×10−04 | UP | AGPAT4 intronic
transcript 1 |
|
| 388815 | LINC00478 | 3.03 | 8.11 |
2.7×10−08 | UP | Long intergenic
non-protein coding RNA 478 |
Identification of hub lncRNAs and
their associated disease network
There was a positive association between mRNA and
protein expression changes; therefore, putative PPI network maps
were constructed for the united datasets of chemoresistance using
the STRING database and Cytoscape. Excluding the DEGs distributed
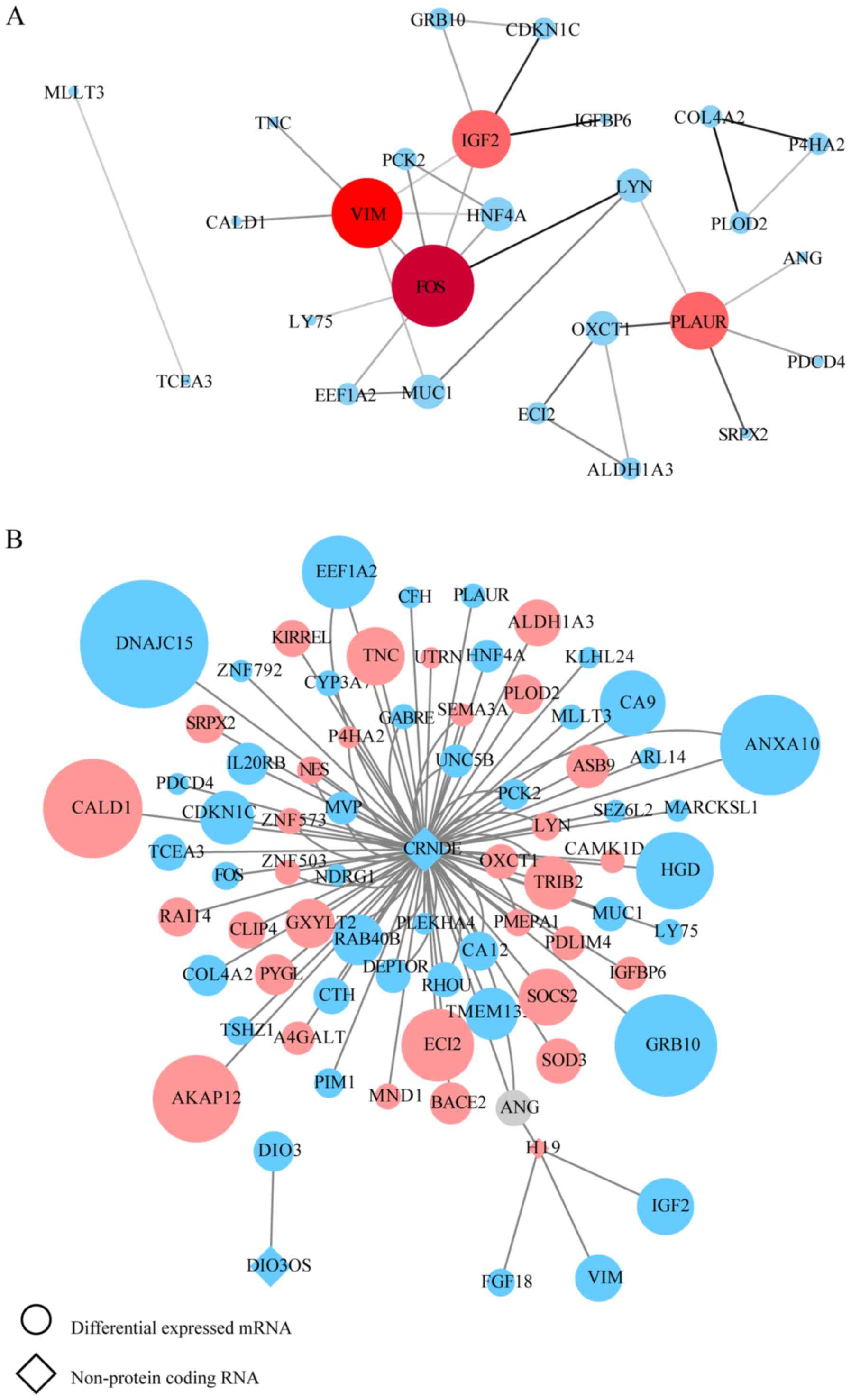
on the edge of the PPI network, the remaining 26 and 57 central
DEGs in the OxPt and SN-38 groups, respectively, were evaluated
using the CentiScaPe plugin. The degree index of each central DEG
was calculated and hub genes were defined using the criterion that
the degree value must be ≥5. In the OxPt group, the hub genes were
FOS, VIM, PLAUR and IGF2. In the SN-38 group, the hub genes were
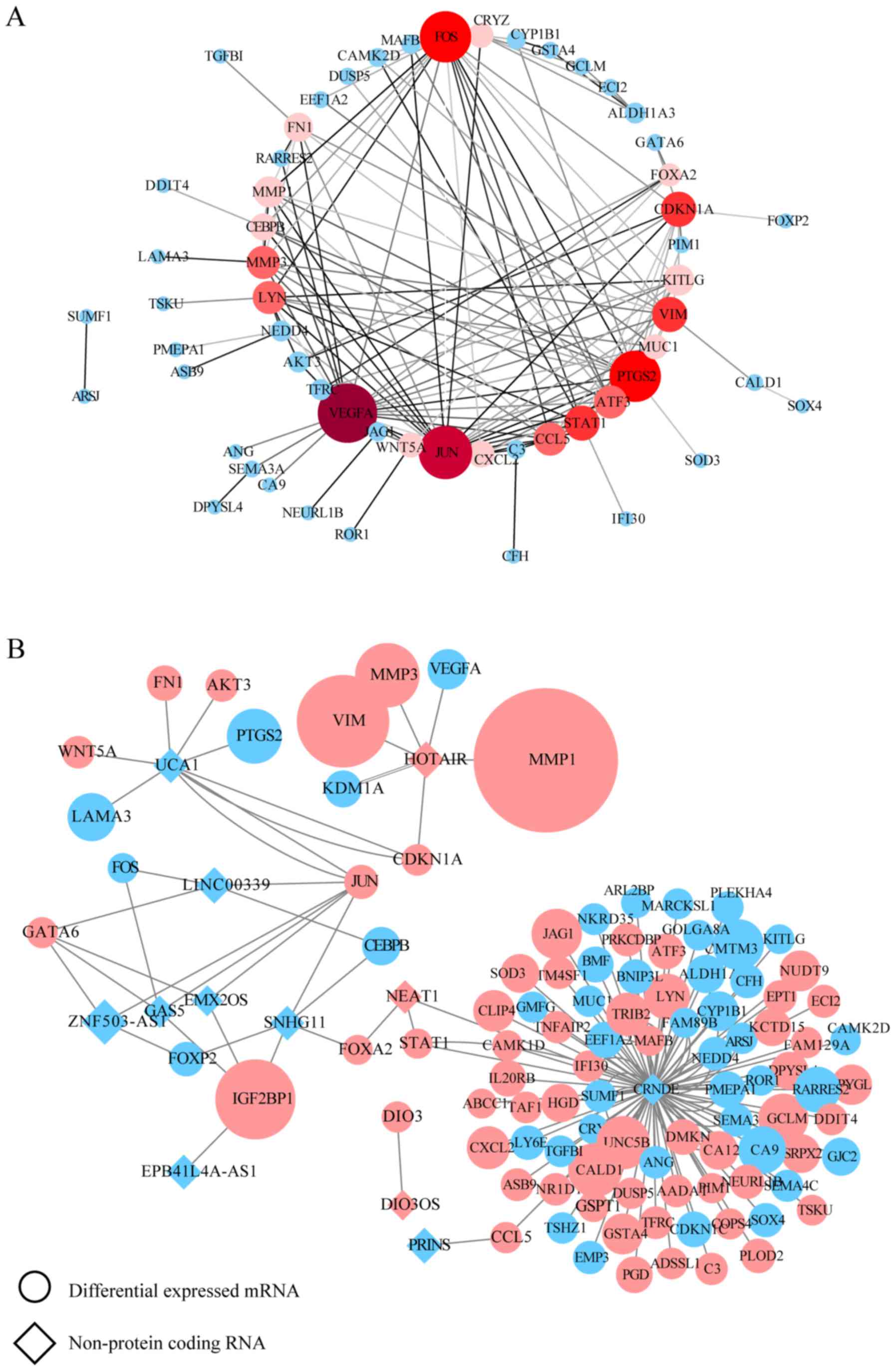
VEGFA, JUN, PTGS2, FOS, VIM, STAT1, CDKN1A, MMP3, CCL5, LYN, ATF3,
KITLG, MMP1, FN1, CEBPB, CXCL2, MUC1, CRYZ, WNT5A and FOXA2
(Tables SVII and SVIII). These hub genes are marked in red
in the PPI network (Figs. 2A and
3A). In a similar manner, the
interaction network between each specific lncRNA and its potential
targets in DEGs were visualized using Cytoscape in Figs. 2B and 3B. Up- and downregulated DEGs are colored
in red and blue, respectively. The log2FC value is also
indicated by the size of the node. Each hub lncRNA was identified
by calculating its downstream hub genes whose count should be
ranked in the top three and larger than 2. During OxPt screening,
the lncRNAs CRNDE (target DEGs: PLAUR and FOS) and H19 (target
DEGs: IGF2 and VIM) stood out. SN-38 screening revealed the
following lncRNAs: CRNDE (target DEGs: STAT1, MUC1, LYN, KITLG,
CXCL2, CRYZ, CCL5 and ATF3), HOTAIR (target DEGs: VEGFA, CDKN1A,
VIM, MMP3, and MMP1), and UCA1 (target DEGs: CDKN1A, JUN, WNT5A,
PTGS2, and FN1).
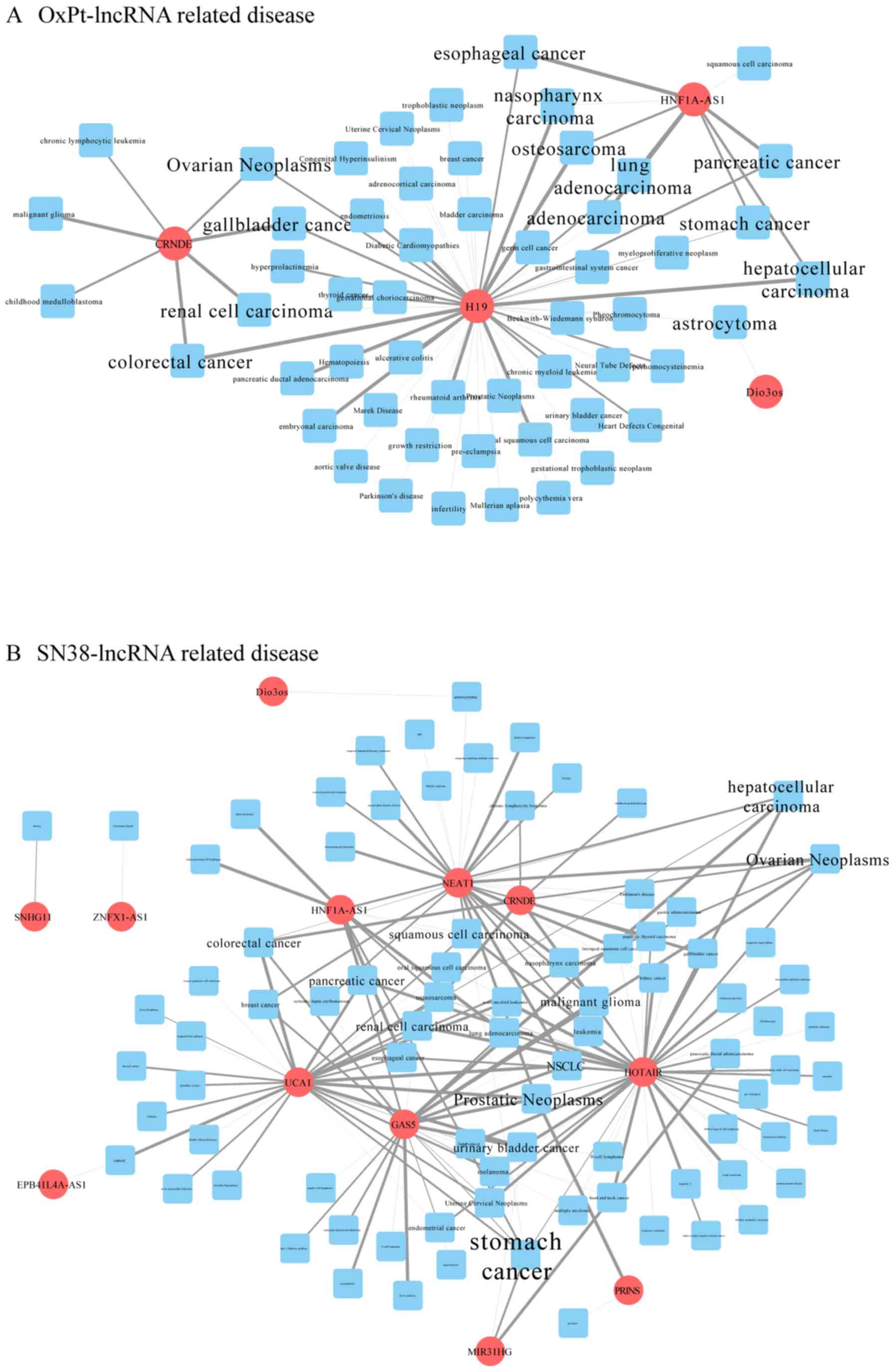
Information on lncRNA-associated diseases was
obtained in full scale according to the MNDR database. By
integrating this information in Cytoscape, the major related
diseases associated with the differentially expressed lncRNAs were
clearly sorted out. The more lncRNAs a disease is connected to, the
larger its label. As shown in Fig.
4, cancer is the word that most frequently appears. Among them,
CRC is obvious in the OxPt-lncRNA-related disease network, which is
strongly connected to CRNDE and H19 (Fig. 4A), whereas the SN-38-lncRNA-related
disease network is strongly connected to GAS5, UCA1, NEAT1, and
CRNDE (Fig. 4B).
lncRNA expression validation and
survival analysis in TCGA
TCGA database was utilized to validate and compare
hub lncRNA expression in tumor and normal tissues. Although the
results cannot relate the expression levels of these lncRNAs to
chemoresistance, it did, to a certain extent, partially confirm the
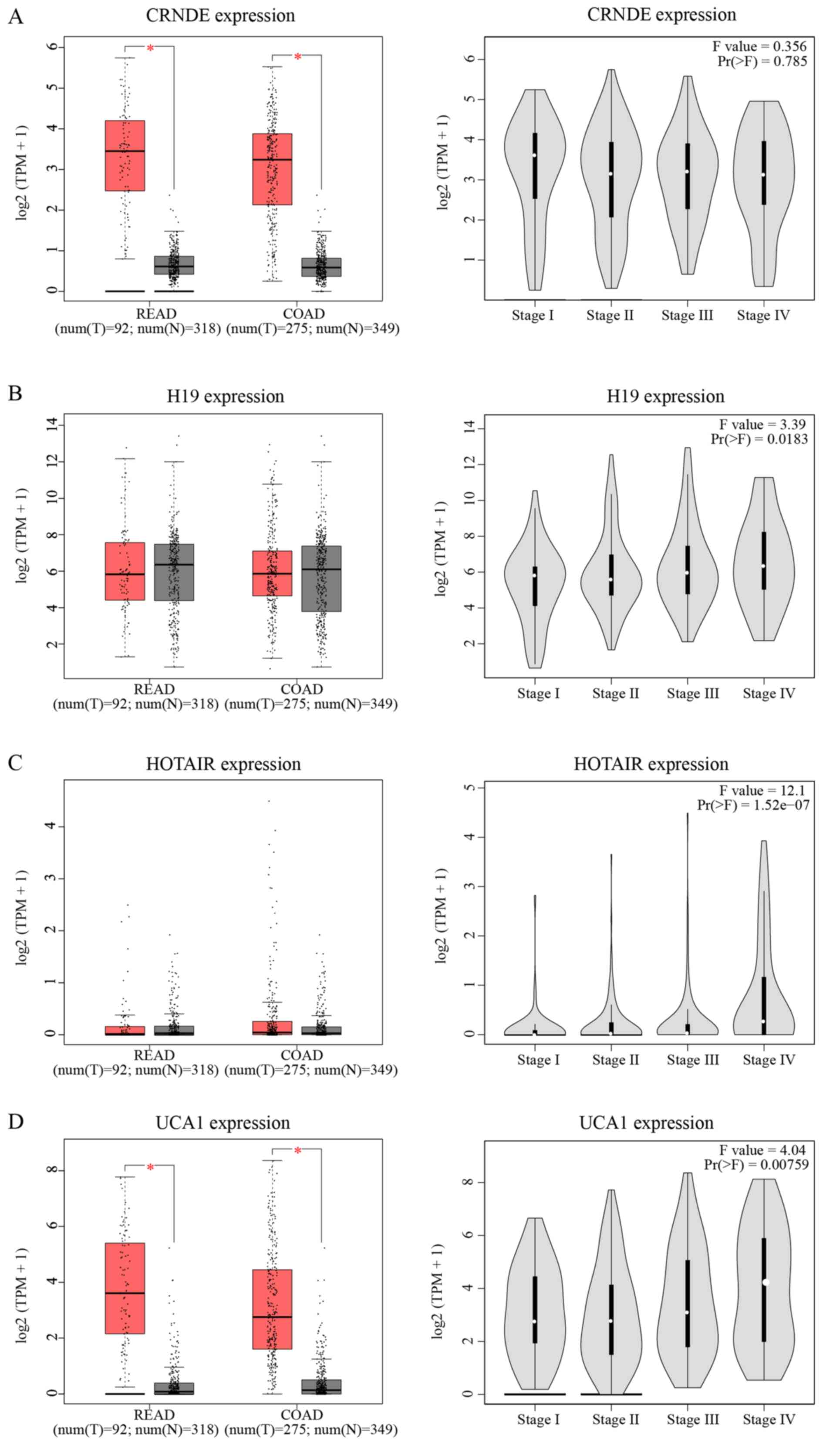
hazard degree of each lncRNA. As indicated in Fig. 5A-D, the expression of CRNDE and
UCA1 was significantly increased in tumor tissue compared with in
normal tissue (P<0.05). Except for CRNDE, the expression of
three other lncRNAs increased as the TNM stage increased. In
Fig. 5E-H, only HOTAIR showed a
significant relationship with OS and disease-free survival (DFS;
P<0.05). Patients with increased expression of HOTAIR suffered a
shorter OS (HR=1.9, P=0.0066) and DFS (HR=1.8, P=0.012) compared to
those with weaker expression.
Discussion
In recent years, the prevalence of CRC has increased
worldwide. Due to the popularity of prophylactic screening, CRC can
be diagnosed and treated increasingly early (3). However, many patients are in late
stages when diagnosed (≥stage III or high-risk stage II) and
require neoadjuvant or adjuvant chemotherapy (7). Some patients must face a tough
reality-chemoresistance-which results in a shorter survival owing
to limited chemotherapy choices. Although a previous bioinformatics
analysis has revealed differences in gene expression patterns in
chemoresistant cells, such as original study of GSE42387 done by
Jensen et al (26), few
have determined pivotal lncRNAs involved in this process.
Therefore, a comprehensive analysis at the whole-genome level is
necessary to identify key lncRNAs and provide detailed gene
signatures for effective chemotherapy. The present study aimed at
exploring the relationship between aberrant expressed lncRNAs and
mRNAs. The GEO dataset was re-analyzed by relaxing DEG screening
condition to gain more information. Only DEGs predicted to be the
targets of differential expressed lncRNAs were involved in the
following analysis.
To gain further insight into the biological pathways
and identify key factors involved in chemoresistance, functional
analyses of lncRNA-targeted DEGs, including GO annotation and KEGG
pathway enrichment analyses were performed. The present study found
that several DEGs are components of the extracellular exosome.
Interestingly, the exosome is believed to confer chemoresistance to
pancreatic cancer (35), breast
cancer (36), gastric cancer
(37) and CRC (38). It is believed that due to their
cell-to-cell communication, exosomes protect tumor cells from the
cytotoxic effects of chemotherapy drugs and transfer
chemoresistance properties to nearby cells (39). The items enriched in GO biological
processes indicate that chemoresistance occurring during both OxPt
and irinotecan therapy is associated with an imbalance between cell
proliferation and apoptosis regulation, cell energetic metabolism
under hypoxic conditions, and angiogenesis. However, it seems that
irinotecan treatment yields more DEGs than OxPt treatment. The
present study hypothesized that this is why OxPt is the first
choice for the clinical therapy of CRC.
In the present study, four hub lncRNAs were
identified: CRNDE, H19, UCA1 and HOTAIR. CRNDE is a 1,910-nt
cancer-secreted lncRNA transcribed by human chromosome 16
(16q12.2). Decades of research have shown that the high expression
of CRNDE is involved in the progression of several cancers, such as
renal cell carcinoma (40),
hepatocellular carcinoma (41) and
glioma tumors (42). Liu et
al (43) found exosomal
CRNDE-h in the sera of CRC patients. They also found a positive
correlation between serum levels of CRNDE-h and regional lymph node
or distant metastasis. Therefore, they believe that exosomal CRNDE
can be used as a serum biochemical marker in the diagnosis and
prognosis of CRC. Another recent study revealed that the
overexpression of CRNDE promoted the development of CRC by
suppressing the expression of dual-specificity phosphatase 5
(DUSP5) and CDKN1A. Moreover, the knockdown of CRNDE significantly
blocked cell proliferation and induced apoptosis (44). In glioma, CRNDE promoted cell
growth and invasion via mTOR signaling (42). The involvement of CRNDE in the
chemoresistance of CRC treatment has also been assessed by a number
of researchers (45,46). After treating CRC cells with
different concentrations of 5-FU, Han et al (45) found that CRNDE-knockdown CRC cells
exhibited increased sensitivity to 5-FU and OxPt treatment.
Together, these results suggest that CRNDE is closely related to
the development and chemoresistance of CRC.
Another hub lncRNA identified in the present study,
lncRNA H19, is a key oncogenic mediator coded by chromosome 11.
According to previous studies, H19 is overexpressed in various
cancers, including breast cancer (47), thyroid carcinoma (48) and lung cancer (49). H19 also plays an oncogenic role in
the development of CRC. The expression of H19 is increased in CRC
tissues from advanced TNM stage patients (50). Enhanced H19 expression was also
found in a CRC cell line and treatment with H19 significantly
induced the proliferation of CRC cells, suggesting an underlying
relationship between H19 and colorectal carcinogenesis (51). H19 also plays an important role in
the migration and invasion of CRC cells (52). Increasing research attention has
been given to the chemoresistance of H19 in the treatment of cancer
(53,54). Emerging evidence also indicates
that H19 knockout in the HT-29 cell line (MTX-resistant colorectal
cells) reduces the risk of drug resistance. Further investigations
showed that this H19-related chemoresistance is mediated by the
Wnt/β-catenin signaling pathway (55).
HOTAIR is a novel oncogenic lncRNA transcribed by
the HoxC gene. As with the abovementioned lncRNAs, HOTAIR has also
been identified in several cancers, including osteosarcoma
(56), cervical cancer (57) and gastric cancer (58). Similarly, HOTAIR exerts its
oncogenic role by regulating CRC cell proliferation, invasion, and
migration by mediating p21 (59).
Elevated HOTAIR expression has also been detected in CRC cells and
tissues relative to adjacent control tissues (60,61).
The precise role of HOTAIR in chemoresistance in the chemotherapy
of CRC was also recently reported. Li et al (62) indicated that HOTAIR promotes the
chemoresistance of CRC cells to 5-FU by suppressing miR-218 and
activating NF-κB signaling. Another study performed by Xiao et
al (63) found that HOTAIR
knockout improved the sensitivity of CRC cells to cisplatin and
paclitaxel and that this mechanism may, at least in part, be
related to the expression of miR-203a-3p and activation of the
Wnt/β-catenin signaling pathway.
The regulatory role of UCA1 in chemotherapy drug
resistance in CRC was also investigated. The upregulated lncRNA
UCA1 is correlated with the progression of lung cancer, esophageal
squamous cell carcinoma and CRC (64–66)
and predicts a poor prognosis. Compared with control tissues, an
increased level of UCA1 was found in CRC tissues and cells and the
level of UCA1 was positively correlated with tumor size and depth
(67). Further in vitro
experiments suggested that UCA1 induces CRC cell proliferation and
inhibits apoptosis. Recent studies have also revealed the potential
role of UCA1 in chemoresistance in CRC. CRC cells with UCA1
knockdown exhibit sensitivity to 5-FU by increasing tumor cell
apoptosis. Conversely, UCA1 overexpression may result in increased
resistance to 5-FU treatment, suggesting that UCA1 may predict the
response to CRC chemotherapy (68). Furthermore, overexpression of UCA1
has a consanguineous connection with radioresistance in CRC
treatment (69).
Collectively, the current study identified four
pivotal lncRNAs, CRNDE, H19, UCA1 and HOTAIR, which are strongly
associated with CRC chemoresistance and can be used as predictive
factors for treatment sensitivity and tumor prognosis. Functional
enrichment analysis indicated that an imbalance between cell
proliferation and apoptosis, cell energetic metabolism under
hypoxic conditions, and angiogenesis were the key biological
processes whereby lncRNAs perturbed gene expression and altered
sensitivity of tumor cells to chemotherapy. However, the results of
the present study are based entirely on bioinformatics analyses and
lack in vivo and in vitro experimental evidence.
Further research is required to delineate their potential roles in
chemoresistance.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used during the present study are
downloaded from TCGA and GEO datasets and analyzed data is
available from the corresponding author on reasonable request.
Authors' contributions
FFS was involved in the conception and design of the
study, project administration and drafted the manuscript; FFS and
WWL collected data from the internet, performed bioinformatics
analyses and organized the figures and tables; WWL, performed the
literature search. JQ was involved in the conception and design of
the study, supervised the study and critically reviewed the
manuscript. All authors have read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
O'Connell JB, Maggard MA and Ko CY: Colon
cancer survival rates with the new American Joint Committee on
cancer sixth edition staging. J Natl Cancer Inst. 96:1420–1425.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ,
Meester RGS, Barzi A and Jemal A: Colorectal cancer statistics,
2017. CA Cancer J Clin. 67:177–193. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Moriarity A, O'Sullivan J, Kennedy J,
Mehigan B and McCormick P: Current targeted therapies in the
treatment of advanced colorectal cancer: A review. Ther Adv Med
Oncol. 8:276–293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Mestier L, Manceau G, Neuzillet C,
Bachet JB, Spano JP, Kianmanesh R, Vaillant JC, Bouché O, Hannoun L
and Karoui M: Primary tumor resection in colorectal cancer with
unresectable synchronous metastases: A review. World J Gastrointest
Oncol. 6:156–169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Raymond E, Faivre S, Woynarowski JM and
Chaney SG: Oxaliplatin: Mechanism of action and antineoplastic
activity. Semin Oncol. 25:4–12. 1998.PubMed/NCBI
|
|
7
|
Benson AB, Venook AP, Al-Hawary MM,
Cederquist L, Chen YJ, Ciombor KK, Cohen S, Cooper HS, Deming D,
Engstrom PF, et al: NCCN guidelines insights: Colon cancer, version
2.2018. J Natl Compr Canc Netw. 16:359–369. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bahrami A, Amerizadeh F, Hassanian SM,
ShahidSales S, Khazaei M, Maftouh M, Ghayour-Mobarhan M, Ferns GA
and Avan A: Genetic variants as potential predictive biomarkers in
advanced colorectal cancer patients treated with oxaliplatin-based
chemotherapy. J Cell Physiol. 233:2193–2201. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hammond WA, Swaika A and Mody K:
Pharmacologic resistance in colorectal cancer: A review. Ther Adv
Med Oncol. 8:57–84. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yan D, Tu L, Yuan H, Fang J, Cheng L,
Zheng X and Wang X: WBSCR22 confers oxaliplatin resistance in human
colorectal cancer. Sci Rep. 7:154432017. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Mao L, Li Y, Zhao J, Li Q, Yang B, Wang Y,
Zhu Z, Sun H and Zhai Z: Transforming growth factor-β1 contributes
to oxaliplatin resistance in colorectal cancer via epithelial to
mesenchymal transition. Oncol Lett. 14:647–654. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gnoni A, Russo A, Silvestris N, Maiello E,
Vacca A, Marech I, Numico G, Paradiso A, Lorusso V and Azzariti A:
Pharmacokinetic and metabolism determinants of fluoropyrimidines
and oxaliplatin activity in treatment of colorectal patients. Curr
Drug Metab. 12:918–931. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Conti JA, Kemeny NE, Saltz LB, Huang Y,
Tong WP, Chou TC, Sun M, Pulliam S and Gonzalez C: Irinotecan is an
active agent in untreated patients with metastatic colorectal
cancer. J Clin Oncol. 14:709–715. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cecchin E, Corona G, Masier S, Biason P,
Cattarossi G, Frustaci S, Buonadonna A, Colussi A and Toffoli G:
Carboxylesterase isoform 2 mRNA expression in peripheral blood
mononuclear cells is a predictive marker of the irinotecan to SN38
activation step in colorectal cancer patients. Clin Cancer Res.
11:6901–6907. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chabot GG, Robert J, Lokiec F and Canal P:
Irinotecan pharmacokinetics. Bull Cancer. 11–20. 1998.(In French).
PubMed/NCBI
|
|
16
|
Hecht JR: Gastrointestinal toxicity of
irinotecan. Oncology (Williston Park). 12:72–78. 1998.PubMed/NCBI
|
|
17
|
Xu Y and Villalona-Calero M: Irinotecan:
Mechanisms of tumor resistance and novel strategies for modulating
its activity. Ann Oncol. 13:1841–1851. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ransohoff JD, Wei Y and Khavari PA: The
functions and unique features of long intergenic non-coding RNA.
Nat Rev Mol Cell Biol. 19:143–157. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huarte M: The emerging role of lncRNAs in
cancer. Nat Med. 21:1253–1261. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kim T and Croce CM: Long noncoding RNAs:
Undeciphered cellular codes encrypting keys of colorectal cancer
pathogenesis. Cancer Lett. 417:89–95. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li H, Ma SQ, Huang J, Chen XP and Zhou HH:
Roles of long noncoding RNAs in colorectal cancer metastasis.
Oncotarget. 8:39859–39876. 2017.PubMed/NCBI
|
|
22
|
Han D, Wang M, Ma N, Xu Y, Jiang Y and Gao
X: Long noncoding RNAs: Novel players in colorectal cancer. Cancer
Lett. 361:13–21. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang M, Han D, Yuan Z, Hu H, Zhao Z, Yang
R, Jin Y, Zou C, Chen Y, Wang G, et al: Long non-coding RNA H19
confers 5-Fu resistance in colorectal cancer by promoting
SIRT1-mediated autophagy. Cell Death Dis. 9:11492018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li L, Shang J, Zhang Y, Liu S, Peng Y,
Zhou Z, Pan H, Wang X, Chen L and Zhao Q: MEG3 is a prognostic
factor for CRC and promotes chemosensitivity by enhancing
oxaliplatin-induced cell apoptosis. Oncol Rep. 38:1383–1392. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Luo J, Qu J, Wu DK, Lu ZL, Sun YS and Qu
Q: Long non-coding RNAs: A rising biotarget in colorectal cancer.
Oncotarget. 8:22187–22202. 2017.PubMed/NCBI
|
|
26
|
Jensen NF, Stenvang J, Beck MK, Hanáková
B, Belling KC, Do KN, Viuff B, Nygård SB, Gupta R, Rasmussen MH, et
al: Establishment and characterization of models of chemotherapy
resistance in colorectal cancer: Towards a predictive signature of
chemoresistance. Mol Oncol. 9:1169–1185. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yi Y, Zhao Y, Li C, Zhang L, Huang H, Li
Y, Liu L, Hou P, Cui T, Tan P, et al: RAID v2. 0: An updated
resource of RNA-associated interactions across organisms. Nucleic
Acids Res. 45:D115–D118. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Szklarczyk D, Gable AL, Lyon D, Junge A,
Wyder S, Huerta-Cepas J, Simonovic M, Doncheva NT, Morris JH, Bork
P, et al: STRING v11: Protein-protein association networks with
increased coverage, supporting functional discovery in genome-wide
experimental datasets. Nucleic Acids Res. 47:D607–D613. 2018.
View Article : Google Scholar :
|
|
32
|
Scardoni G, Tosadori G, Faizaan M, Spoto
F, Fabbri F and Laudanna C: Biological network analysis with
CentiScaPe: Centralities and experimental dataset integration.
F1000Res. 3:e1392014. View Article : Google Scholar
|
|
33
|
Cui T, Zhang L, Huang Y, Yi Y, Tan P, Zhao
Y, Hu Y, Xu L, Li E and Wang D: MNDR v2.0: An updated resource of
ncRNA-disease associations in mammals. Nucleic Acids Res.
46:D371–D374. 2018.PubMed/NCBI
|
|
34
|
Tang Z, Li C, Kang B, Gao G, Li C and
Zhang Z: GEPIA: A web server for cancer and normal gene expression
profiling and interactive analyses. Nucleic Acids Res. 45:W98–W102.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Patel GK, Khan MA, Bhardwaj A, Srivastava
SK, Zubair H, Patton MC, Singh S, Khushman M and Singh AP: Exosomes
confer chemoresistance to pancreatic cancer cells by promoting ROS
detoxification and miR-155-mediated suppression of key
gemcitabine-metabolising enzyme, DCK. Br J Cancer. 116:609–619.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen WX, Liu XM, Lv MM, Chen L, Zhao JH,
Zhong SL, Ji MH, Hu Q, Luo Z, Wu JZ and Tang JH: Exosomes from
drug-resistant breast cancer cells transmit chemoresistance by a
horizontal transfer of microRNAs. PLoS One. 9:e952402014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ji R, Zhang B, Zhang X, Xue J, Yuan X, Yan
Y, Wang M, Zhu W, Qian H and Xu W: Exosomes derived from human
mesenchymal stem cells confer drug resistance in gastric cancer.
Cell Cycle. 14:2473–2483. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hu Y, Yan C, Mu L, Huang K, Li X, Tao D,
Wu Y and Qin J: Fibroblast-derived exosomes contribute to
chemoresistance through priming cancer stem cells in colorectal
cancer. PLoS One. 10:e01256252015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Brinton LT, Sloane HS, Kester M and Kelly
KA: Formation and role of exosomes in cancer. Cell Mol Life Sci.
72:659–671. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ding C, Han F, Xiang H, Xia X, Wang Y, Dou
M, Zheng J, Li Y, Xue W, Ding X and Tian P: LncRNA CRNDE is a
biomarker for clinical progression and poor prognosis in clear cell
renal cell carcinoma. J Cell Biochem. 119:10406–10414. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wang H, Ke J, Guo Q, Barnabo Nampoukime
KP, Yang P and Ma K: Long non-coding RNA CRNDE promotes the
proliferation, migration and invasion of hepatocellular carcinoma
cells through miR-217/MAPK1 axis. J Cell Mol Med. 22:5862–5876.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wang Y, Wang Y, Li J, Zhang Y, Yin H and
Han B: CRNDE, a long-noncoding RNA, promotes glioma cell growth and
invasion through mTOR signaling. Cancer Lett. 367:122–128. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu T, Zhang X, Gao S, Jing F, Yang Y, Du
L, Zheng G, Li P, Li C and Wang C: Exosomal long noncoding RNA
CRNDE-h as a novel serum-based biomarker for diagnosis and
prognosis of colorectal cancer. Oncotarget. 7:85551–85563. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ding J, Li J, Wang H, Tian Y, Xie M, He X,
Ji H, Ma Z, Hui B, Wang K and Ji G: Long noncoding RNA CRNDE
promotes colorectal cancer cell proliferation via epigenetically
silencing DUSP5/CDKN1A expression. Cell Death Dis. 8:e29972017.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Gao H, Song X, Kang T, Yan B, Feng L, Gao
L, Ai L, Liu X, Yu J and Li H: Long noncoding RNA CRNDE functions
as a competing endogenous RNA to promote metastasis and oxaliplatin
resistance by sponging miR-136 in colorectal cancer. Onco Targets
Ther. 10:205–216. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Han J, Han B, Wu X, Hao J, Dong X, Shen Q
and Pang H: Knockdown of lncRNA H19 restores chemo-sensitivity in
paclitaxel-resistant triple-negative breast cancer through
triggering apoptosis and regulating Akt signaling pathway. Toxicol
Appl Pharmacol. 359:55–61. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Li M, Chai HF, Peng F, Meng YT, Zhang LZ,
Zhang L, Zou H, Liang QL, Li MM, Mao KG, et al: Estrogen receptor β
upregulated by lncRNA-H19 to promote cancer stem-like properties in
papillary thyroid carcinoma. Cell Death Dis. 9:11202018. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Huang Z, Lei W, Hu HB, Zhang H and Zhu Y:
H19 promotes non-small-cell lung cancer (NSCLC) development through
STAT3 signaling via sponging miR-17. J Cell Physiol. 233:6768–6776.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Tsang WP, Ng EK, Ng SS, Jin H, Yu J, Sung
JJ and Kwok TT: Oncofetal H19-derived miR-675 regulates tumor
suppressor RB in human colorectal cancer. Carcinogenesis.
31:350–358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Han D, Gao X, Wang M, Qiao Y, Xu Y, Yang
J, Dong N, He J, Sun Q, Lv G, et al: Long noncoding RNA H19
indicates a poor prognosis of colorectal cancer and promotes tumor
growth by recruiting and binding to eIF4A3. Oncotarget.
7:22159–22173. 2016.PubMed/NCBI
|
|
52
|
Yang W, Redpath RE, Zhang C and Ning N:
Long non-coding RNA H19 promotes the migration and invasion of
colon cancer cells via MAPK signaling pathway. Oncol Lett.
16:3365–3372. 2018.PubMed/NCBI
|
|
53
|
Ma H, Yuan L, Li W, Xu K and Yang L: The
LncRNA H19/miR-193a-3p axis modifies the radio-resistance and
chemotherapeutic tolerance of hepatocellular carcinoma cells by
targeting PSEN1. J Cell Biochem. 119:8325–8335. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Si X, Zang R, Zhang E, Liu Y, Shi X, Zhang
E, Shao L, Li A, Yang N, Han X, et al: LncRNA H19 confers
chemoresistance in ERα-positive breast cancer through epigenetic
silencing of the pro-apoptotic gene BIK. Oncotarget. 7:81452–81462.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Wu KF, Liang WC, Feng L, Pang JX, Waye MM,
Zhang JF and Fu WM: H19 mediates methotrexate resistance in
colorectal cancer through activating Wnt/β-catenin pathway. Exp
Cell Res. 350:312–317. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Wang N, Meng X, Liu Y, Chen Y and Liang Q:
LPS promote Osteosarcoma invasion and migration through
TLR4/HOTAIR. Gene. 680:1–8. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Kim HJ, Lee DW, YIm GW, Nam EJ, Kim S, Kim
SW and Kim YT: Long non-coding RNA HOTAIR is associated with human
cervical cancer progression. Int J Oncol. 46:521–530. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Xian HP, Zhuo ZL, Sun YJ, Liang B and Zhao
XT: Circulating long non-coding RNAs HULC and ZNFX1-AS1 are
potential biomarkers in patients with gastric cancer. Oncol Lett.
16:4689–4698. 2018.PubMed/NCBI
|
|
59
|
Lin K, Jiang H, Zhang LL, Jiang Y, Yang
YX, Qiu GD, She YQ, Zheng JT, Chen C, Fang L and Zhang SY:
Down-regulated LncRNA-HOTAIR suppressed colorectal cancer cell
proliferation, invasion, and migration by mediating p21. Dig Dis
Sci. 63:2320–2331. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Lu X, Liu Z, Ning X, Huang L and Jiang B:
The long noncoding RNA HOTAIR promotes colorectal cancer
progression by sponging miR-197. Oncol Res. 26:473–481. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Luo ZF, Zhao D, Li XQ, Cui YX, Ma N, Lu
CX, Liu MY and Zhou Y: Clinical significance of HOTAIR expression
in colon cancer. World J Gastroenterol. 22:5254–5259. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Li P, Zhang X, Wang L, Du L, Yang Y, Liu
T, Li C and Wang C: lncRNA HOTAIR contributes to 5FU resistance
through suppressing miR-218 and activating NF-κB/TS signaling in
colorectal cancer. Mol Ther Nucleic Acids. 8:356–369. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Xiao Z, Qu Z, Chen Z, Fang Z, Zhou K,
Huang Z, Guo X and Zhang Y: LncRNA HOTAIR is a prognostic biomarker
for the proliferation and chemoresistance of colorectal cancer via
MiR-203a-3p-mediated Wnt/ss-catenin signaling pathway. Cell Physiol
Biochem. 46:1275–1285. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Wang HM, Lu JH, Chen WY and Gu AQ:
Upregulated lncRNA-UCA1 contributes to progression of lung cancer
and is closely related to clinical diagnosis as a predictive
biomarker in plasma. Int J Clin Exp Med. 8:11824–11830.
2015.PubMed/NCBI
|
|
65
|
Shalini S, Dorstyn L, Dawar S and Kumar S:
Old, new and emerging functions of caspases. Cell Death Differ.
22:526–539. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Li JY, Ma X and Zhang CB: Overexpression
of long non-coding RNA UCA1 predicts a poor prognosis in patients
with esophageal squamous cell carcinoma. Int J Clin Exp Pathol.
7:7938–7944. 2014.PubMed/NCBI
|
|
67
|
Han Y, Yang YN, Yuan HH, Zhang TT, Sui H,
Wei XL, Liu L, Huang P, Zhang WJ and Bai YX: UCA1, a long
non-coding RNA up-regulated in colorectal cancer influences cell
proliferation, apoptosis and cell cycle distribution. Pathology.
46:396–401. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Bian Z, Jin L, Zhang J, Yin Y, Quan C, Hu
Y, Feng Y, Liu H, Fei B, Mao Y, et al: LncRNA-UCA1 enhances cell
proliferation and 5-fluorouracil resistance in colorectal cancer by
inhibiting miR-204-5p. Sci Rep. 6:238922016. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Yang X, Liu W, Xu X, Zhu J, Wu Y, Zhao K,
He S, Li M, Wu Y, Zhang S, et al: Downregulation of long noncoding
RNA UCA1 enhances the radiosensitivity and inhibits migration via
suppression of epithelialmesenchymal transition in colorectal
cancer cells. Oncol Rep. 40:1554–1564. 2018.PubMed/NCBI
|