Introduction
The folliculogenesis is a compound process that
leads to the formation of the fully grown ovarian follicle
(1,2). The developed follicle is composed of
several cell layers, including theca cells (TCs) and granulosa
cells (GCs). Our recent study has shown that porcine GCs undergo
permanent morphological modification when kept in the primary in
vitro culture system. After long-term culture, GCs underwent
transformation into spindle-like structures, which was accompanied
by changes in their gene expression profile. Moreover, the GCs
display structural and functional role in the active pre-ovulatory
and ovulatory follicle, forming follicular architecture and
expressing a number of hormone receptors. Some of that properties
seem to prevail ex vivo, as we have observed a
time-dependent expression of LHR, FSHR, CYP19, and
progesterone receptor genes, in porcine GCs during long-term in
vitro culture.
Using a porcine model, we have recently shown that
GCs, isolated from pre-ovulatory ovarian follicles, can be
successfully cultured in vitro for short and/or long time
periods. Moreover, we found that these GCs are characterized by
increased in vitro proliferation capability. Although the
increase in cells' proliferation is accompanied by cellular ageing
and subsequently decreased by a number of passages, the GCs have
also displayed significant differentiational capacity. It was also
presented that these cells may differentiate even into cell types
unassociated with the reproductive process. Kossowska-Tomaszczuk
recently determined that after long-term cultivation and under
influence of several substances, including LIF and dexamethasone,
GCs may differentiate into chondroblasts, osteoblasts and other
adult precursor cells (3,4). These experiments have also shown how
the huge differentiational plasticity of human GCs during primary
in vitro culture can be used in clinical situations. This
may open a new gate in the potential application of human granulosa
cells, which are nowadays mostly treated as remnant IVF material,
mostly in autologous and donor-host grafts, as they exhibit
increased stem-like specificity.
The experiments by Kossowska-Tomaszczuk open the
possibility of finding new stemness markers in human ovarian
granulosa cells. Microarray assays allow for robust and complex
transcriptomic analysis, leading to the discovery of new molecular
markers of cells function. In this work, we have used the
microarrays to describe the transcriptomic profile of human GCs
during their long-term primary in vitro culture. We have
detected the expression of genes belonging to more than five
hundred gene ontology groups, associated with the ‘biology of human
GCs’ during their in vitro proliferation and
differentiation. In this study, we have investigated the ‘heart
development and morphogenesis’ ontological group, as the potential
source of new markers of GCs plasticity.
Materials and methods
A large part of the materials and methods section is
based on other, published work of the author, presenting results
from the same cycle of studies describing human granulosa cells
(5).
Patients and collection of granulosa
cells
The GCs were derived from patients undergoing in
vitro fertilization (IVF) procedures, who had given their
informed, written consent to be included in this protocol. The
study group consisted of 8 patients, aged 18–40 years, with
diagnosed infertility, referred to the Division of Infertility and
Reproductive Endocrinology, Poznan University of Medical Sciences,
Poznan, Poland. Patients underwent IVF procedure, based on
controlled ovarian hyperstimulation protocol, adjusted to the
patient's initial infertility workup and ovarian response.
Stimulation was performed with human recombinant FSH (Gonal-F,
Merck Serono) and highly purified hMG-HP (Menopur, Ferring). The
injections with cetrorelix acetate (Cetrotide, Merck Serono) were
administered in an adequate dose, to suppress pituitary function.
Ovulation triggering was based on the subcutaneous injection of
6,500 U of hCG (Ovitrelle, Merck-Serono). GC containing follicular
fluid was collected during transvaginal, ultrasound-guided oocyte
pick-up, performed 36 h after human chorionic gonadotropin
administration. The content of follicles that were over 16 mm in
diameter was immediately passed to an embryologist, who isolated
the oocyte, and pooled the follicular fluid from each ovary. The
fresh pooled samples were centrifuged for 10 min at 200 g, to
separate and collect GCs. Patients with polycystic ovary syndrome
(PCOS), endometriosis, and diminished ovarian reserve (serum
antimüllerian hormone (AMH) less than 0.7 ng/ml, and/or day 2–3 FSH
serum level higher than 15 mU/ml, and/or antral follicle count less
than 9) were excluded from the study. This research has been
approved by Poznan University of Medical Sciences Bioethical
Committee with resolution 558/17.
Primary cell culture
The collected cells were washed twice by
centrifugation at 200 × g for 10 min at RT in the culture medium.
Medium consisted of Dulbecco's modified Eagle's medium (DMEM,
Sigma; Merck KGaA, Darmstadt, Germany), 2% foetal bovine serum FBS
(FBS; Sigma; Merck KGaA), 4 mM L-glutamine (stock 200 mM,
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA), 10
mg/ml gentamicin (Invitrogen; Thermo Fisher Scientific, Inc.),
10,000 U/ml penicillin, and 10,000 µg/ml streptomycin (Invitrogen;
Thermo Fisher Scientific, Inc.). Cells were cultivated at 37°C
under aerobic conditions (5% CO2). Once adherent cells
were more than 90% confluent, they were detached with 0.05%
trypsin-EDTA (Invitrogen; Thermo Fisher Scientific, Inc.) for 1–2
min and counted using a ‘Neubauer improved’ counting chamber (ISO
LAB Laborgerate GmbH, DIN EN ISO Certified 9001). GCs were
cultivated for 30 days. The medium was changed twice a week.
Finally, total RNA was isolated from GCs after 1, 7, 15 and 30
days.
Total RNA isolation
Total RNA was isolated at 4 time periods, after 1,
7, 15 and 30 days of cultivation. The improved Chomczyński-Sacchi
method was used for the RNA isolation (6). The GCs were suspended in 1 ml of a
monophase solution of guanidine thiocyanate and phenol (TRI
Reagent®, Sigma; Merck KGaA). The chloroform was then
added, with the samples centrifuged to obtain three separate
phases. RNA was located in the upper, aqueous phase. RNA was then
stripped with 2-propanol (Sigma; Merck KGaA, catalogue number
I9516), added in an amount adequate for 1 ml of TRI-reagent, and
washed with 75% ethanol. Samples prepared in that way were used for
further analysis.
Microarray expression analysis and
statistics
Total RNA (100 ng) from each pooled sample was
subjected to two rounds of sense cDNA amplification
(Ambion® WT Expression kit). The obtained cDNA was used
for biotin labelling and fragmentation, with the use of
GeneChip® WT Terminal Labeling and Hybridization
(Affymetrix). Biotin-labelled fragments of cDNA (5.5 µg) were
hybridized to the Affymetrix® Human Genome U219 Array
(48°C/20 h). Microarrays were then washed and stained according to
the technical protocol using the Affymetrix GeneAtlas Fluidics
Station. The array strips were scanned employing Imaging Station of
the GeneAtlas System. Preliminary analysis of the scanned chips was
performed using Affymetrix GeneAtlas™ Operating
Software. The quality of gene expression data was confirmed
according to the quality control criteria provided by the software.
The obtained CEL files were imported into downstream data analysis
software.
All of the presented analyses and graphs were
performed using Bioconductor and R programming languages. Each CEL
file was merged with a description file. The Robust Multiarray
Averaging (RMA) algorithm was used to correct background,
normalize, and summarize the results. To determine the statistical
significance of the analyzed genes, moderated t-statistics from the
empirical Bayes method were performed. The obtained P-value was
corrected for multiple comparisons using Benjamini and Hochberg's
false discovery rate. The selection of significantly altered genes
was based on a P-value beneath 0.05 and expression higher than
two-fold.
Differentially expressed genes were subjected to
selection by examination of genes involved in heart development and
morphogenesis. The differentially expressed gene list (separated
for up- and down-regulated genes) was uploaded to the DAVID
software (Database for Annotation, Visualization and Integrated
Discovery) (7).
Subsequently, the relationship between the genes
belonging to chosen GO terms was analysed with the use of GOplot
package (8). The GoPlot package
had calculated the Z-score: The number of upregulated genes minus
the number of downregulated genes divided by the square root of the
count. This information allowed to estimate the direction of
changes in each gene-ontology term.
Moreover, interactions between differentially
expressed genes/proteins belonging to the chosen GO terms were
investigated by STRING10 software (Search Tool for the Retrieval of
Interacting Genes) (9). The list
of gene names was used as a query for the interaction prediction.
The search criteria were based on co-occurrences of genes/proteins
in scientific texts (text mining), co-expression, and
experimentally observed interactions. The results of these analyses
generated a gene/protein interaction network where the intensity of
the edges reflected the strength of the interaction score.
Finally, the functional interactions between the
genes belonging to the chosen GO BP terms were investigated using
REACTOME FIViz application from the Cytoscape 3.6.0 software. This
app accesses the pathways stored in the Reactome database, allowing
for pathway enrichment analysis of a set of genes, visualization of
hit pathways using manually laid-out pathway diagrams directly in
Cytoscape, and investigation of functional relationships among
genes in hit pathways. The app can also access the Reactome
Functional Interaction (FI) network, a highly reliable, manually
curated pathway-based protein functional interaction network
covering over 60% of human proteins.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
The RT-qPCR method was performed to confirm the
results obtained in the analysis of expression microarrays. Three
genes were selected from each heatmap: The ones showing highest,
lowest, and intermediate-level of expression. Changes in the level
of expression of those genes were then examined. Three biological
samples of each gene were used for the analysis. Each test was
performed in 3 replicates. Reverse transcription was based on the
protocols and reagents of SABiosciences (RT2 First Stand
kit-330401), using a Veritimer 96-well Thermal Cycler. One
microgram of each gene's RNA transcript was used for reverse
transcription. Real-time PCR was performed using the 7900HT Fast
Real-Time PCR System (Applied Biosystems; Thermo Fisher Scientific,
Inc.), RT2 SYBR® Green ROX™ qPCR
Master Mix (Qiagen Sciences, Gaithersburg, MD, USA) and
sequence-specific primers (Table
I). Glyceraldehyde-3-phosphate dehydrogenase (GADPH),
β-actin (ACTB), hypoxanthine phosphoribosyltransferase 1
(HRPT1) were used as reference genes. Gene expression was
analyzed using the relative quantification (RQ) method. The q-PCR
starters were designed using Primer3Plus software (primer3plus.com/cgi-bin/dev/primer3plus.cgi).
 | Table I.Oligonucleotide sequences of primers
used for reverse transcription-quantitative polymerase chain
reaction analysis. |
Table I.
Oligonucleotide sequences of primers
used for reverse transcription-quantitative polymerase chain
reaction analysis.
| Gene | Primer sequences
(5′-3′) |
|---|
| PCNA | F:
GGCGTGAACCTCACCAGTAT |
|
| R:
TCTCGGCATATACGTGCAAA |
| LOX | F:
CAGAGGAGAGTGGCTGAAGG |
|
| R:
CCAGGTAGCTGGGGTTTACA |
| OXTR | F:
TTCTTCGTGCAGATGTGGAG |
|
| R:
GGACGAGTTGCTCTTTTTGC |
| BMP4 | F:
AAGCGTAGCCCTAAGCATCA |
|
| R:
TGGTTGAGTTGAGGTGGTCA |
| TPM1 | F:
CTCTCAAAGATGCCCAGGAG |
|
| R:
TCTCATCTGCTGCCTTCTCA |
| TNNC1 | F:
ACCTCTTCCGCATGTTTGAC |
|
| R:
CCACACCCTTCATGAACTCC |
| TTN | F:
GGCATCCCCAAACCTAAAAT |
|
| R:
TTTTGCCACTGCTGATTCTG |
| TGFBR1 | F:
TGTTGGTACCCAAGGAAAGC |
|
| R:
CACTCTGTGGTTTGGAGCAA |
| SMARCD3 | F:
CTCTGAAGAGGCCCATGAAG |
|
| R:
GAACTTCCGCTTCTGTTTGC |
| NDST1 | F:
TCACCTTCAACCTGGGCTAC |
|
| R:
ACGGACTGGTTGTGGAAAAG |
| GAPDH | F:
TCAGCCGCATCTTCTTTTGC |
|
| R:
ACGACCAAATCCGTTGACTC |
| ACTB | F:
AAAGACCTGTACGCCAACAC |
|
| R:
CTCAGGAGGAGCAATGATCTTG |
| HPRT | F:
TGGCGTCGTGATTAGTGATG |
|
| R:
ACATCTCGAGCAAGACGTTC |
Statistical analysis
The analysis of the presented results was conducted
using Bioconductor (www.bioconductor.org) and R programming languages (R
version 3.5.1; www.r-project.org). To determine the statistical
significance of the analysed genes, moderated t-statistics from the
empirical Bayes method were performed. The P-value was corrected
for multiple comparisons using Benjamini and Hochberg's false
discovery rate. P<0.05 was considered to indicate a
statistically significant difference. The statistical significance
of enriched GO terms and KEGG pathways was performed by DAVID
database software (v.6.8; david.ncifcrf.gov). Statistical significance was
calculated using Benjamini method. Each GO term and KEGG pathway
were considered significantly enriched if they contained at least 5
differently expressed genes and showed P<0.05. Statistical
analysis of RT-qPCR results was conducted using Real Statistics
Resource Pack add-on for MS Excel 2016 (Microsoft Corporation,
Redmond, WA, USA).
Results
Whole transcriptome profiling by Affymetrix
microarray allowed for the analysis of expression of gene
transcripts in human ovarian granulosa cells during long-term in
vitro culture. Using Affymetrix® Human HgU 219 Array
we have examined the expression of 22480 transcripts. Genes with a
fold change higher then abs (2)
and with a corrected P-value lower than 0.05 were considered
differentially expressed. This set of genes consisted of 2278
different transcripts.
DAVID (Database for Annotation, Visualization and
Integrated Discovery) software was used for extraction of gene
ontology biological process terms (GO BP). Up- and down-regulated
gene sets were subjected to DAVID search separately, and only gene
sets where adj. P-value was lower than 0.05 were selected. The
DAVID software analysis showed that differentially expressed genes
belong to 582 Gene ontology groups and 45 KEGG pathways. In this
study, we focused on ‘heart morphogenesis’ and ‘heart development’
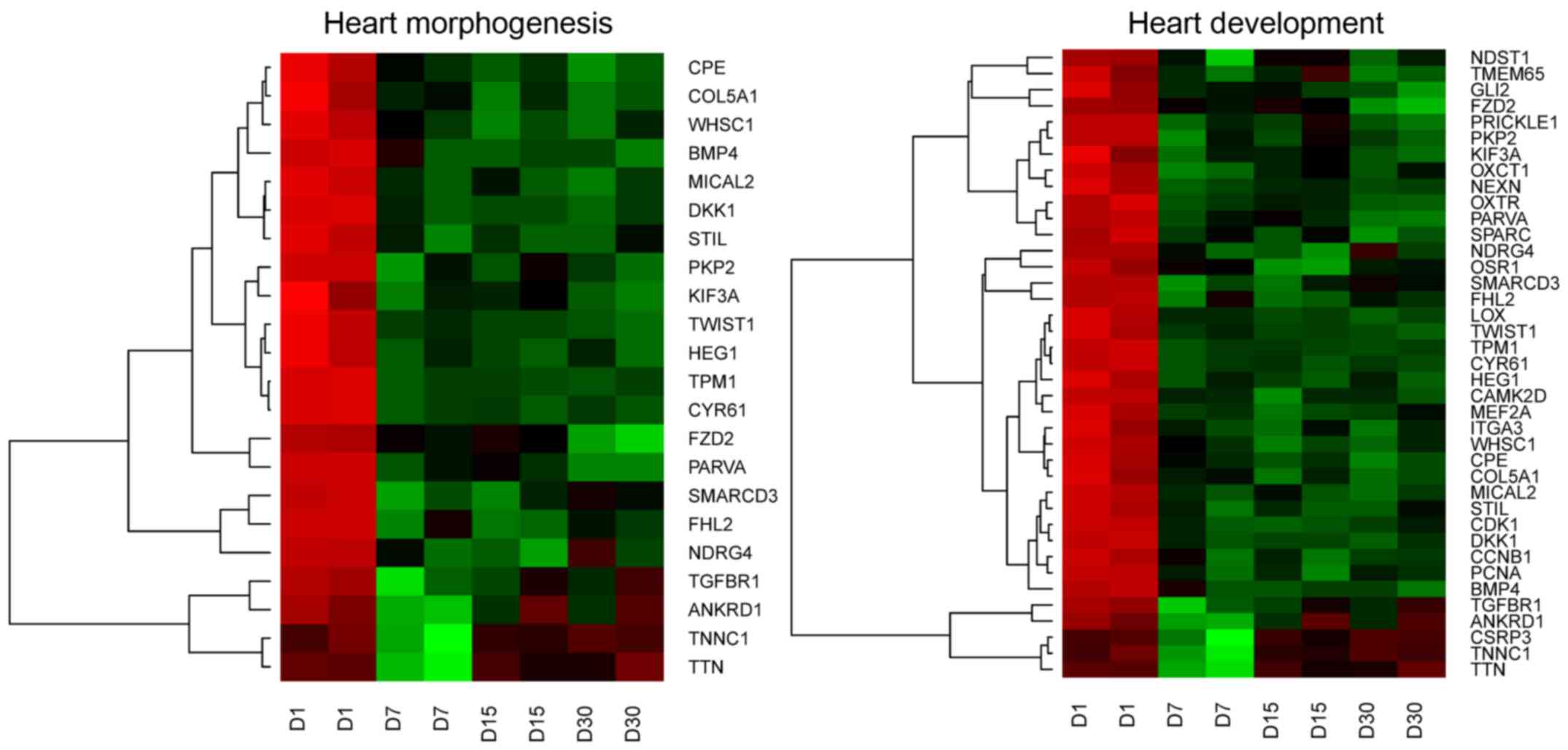
GO BP terms. These sets of genes were subjected to hierarchical
clusterization procedure and presented as heatmaps (Fig. 1). The gene symbols, fold changes in
expression, Entrez gene IDs and corrected P-values of those genes
were shown in Table II.
| Table II.Gene symbols, fold changes in
expression, Entrez gene IDs and corrected P-values of studied
genes. |
Table II.
Gene symbols, fold changes in
expression, Entrez gene IDs and corrected P-values of studied
genes.
| Nazwa | foldD7_D1 | foldD15_D1 | foldD30_D1 |
adj.P.Val.D7_D1 |
adj.P.Val.D15_D1 |
adj.P.Val.D30_D1 | Entrez.Gene ID |
|---|
| LOX | 53,155,736 | 80,241,544 | 102,191,488 | 0.0008508 | 0.0005524 | 0.0004038 | 4,015 |
| OXTR | 38,386,169 | 2,260,881 | 54,854,523 | 0.0006796 | 0.001012 | 0.0003993 | 5,021 |
| NEXN | 35,206,319 | 19,134,152 | 29,664,545 | 0.0012665 | 0.0019093 | 0.0010376 | 91,624 |
| CSRP3 | 27,412,789 | 1,585,293 | 1,057,341 | 0.0191031 | 0.6877273 | 0.9672829 | 8,048 |
| TPM1 | 17,883,412 | 16,942,974 | 18,049,628 | 0.0004891 | 0.0004529 | 0.0003078 | 7,168 |
| ITGA3 | 16,927,809 | 18,842,178 | 22,470.094 | 0.0277094 | 0.0222552 | 0.016508 | 3,675 |
| CDK1 | 13,880,751 | 18,568,254 | 12,196,334 | 0.002233 | 0.0014905 | 0.0018488 | 983 |
| TWIST1 | 13,410,656 | 16,061,334 | 20,741,443 | 0.0013489 | 0.0010638 | 0.000668 | 7,291 |
| ANKRD1 | 11,887,373 | 2,508,097 | 2,617,912 | 0.0247916 | 0.2787596 | 0.2411828 | 27,063 |
| MICAL2 | 1,054,092 | 9,421,967 | 12,712,885 | 0.0068803 | 0.0072788 | 0.0042113 | 9,645 |
| BMP4 | 10,489,539 | 17,221,545 | 21,243,468 | 0.0204669 | 0.0100715 | 0.0070848 | 652 |
| DKK1 | 8,954,961 | 9,635,247 | 10.055,325 | 0.0024355 | 0.0020428 | 0.0015034 | 22,943 |
| COL5A1 | 8,845,619 | 15,695,652 | 18,51,511 | 0.016744 | 0.0071873 | 0.005229 | 1,289 |
| PRICKLE1 | 7,301,909 | 4,726,273 | 9,353,689 | 0.0106664 | 0.0202006 | 0.0056925 | 144,165 |
| GLI2 | 6,93,552 | 7,103,782 | 14,112,758 | 0.0191991 | 0.0163042 | 0.0056787 | 2,736 |
| CCNB1 | 6,641,253 | 8,124,624 | 7,003,387 | 0.0318394 | 0.0211764 | 0.0237061 | 891 |
| STIL | 56,288 | 540,042 | 4,900,424 | 0.0141252 | 0.0134497 | 0.0143661 | 6,491 |
| TTN | 4,908,741 | 1,271,044 | 1,144,088 | 0.0047259 | 0.4647411 | 0.6871576 | 7,273 |
| NDRG4 | 4,817,497 | 7,098,036 | 3,324,764 | 0.0375828 | 0.0171188 | 0.0672424 | 65,009 |
| CPE | 4,531,794 | 6,031,618 | 8,102,031 | 0.0083776 | 0.0043779 | 0.0024281 | 1,363 |
| CYR61 | 4,318,043 | 424,915 | 4,154,475 | 0.0011941 | 0.0011841 | 0.0009317 | 3,491 |
| HEG1 | 4,138,551 | 4,604,733 | 4,311,688 | 0.0083402 | 0.0059934 | 0.0059049 | 57,493 |
| FZD2 | 3,767,679 | 3,278,544 | 13,843,888 | 0.0057158 | 0.0072683 | 0.0006712 | 2,535 |
| TNNC1 | 3,349,751 | 1,217,197 | 1,066,087 | 0.0053652 | 0,4460103 | 0.8215354 | 7,134 |
| FHL2 | 3,073,142 | 3,898,276 | 2,849,228 | 0.0282662 | 0.0139549 | 0.0283799 | 2,274 |
| MEF2A | 3,019,241 | 3,647,594 | 2,798,678 | 0.0041095 | 0.002502 | 0.0039542 | 4,205 |
| WHSC1 | 2,810,461 | 3,935,288 | 3,454,839 | 0.017237 | 0.0063714 | 0.0075821 | 7,468 |
| TMEM65 | 2,790,505 | 19,071 | 3,094,836 | 0.0273694 | 0.0922242 | 0.0162561 | 157,378 |
| SMARCD3 | 2,528,987 | 2,267,934 | 175,636 | 0.0106455 | 0.0135429 | 0.0390055 | 6,604 |
| NDST1 | 2,511,612 | 1,661,929 | 2,158,918 | 0.0456627 | 0.0897835 | 0.0622311 | 3,340 |
| OXCT1 | 2,382,383 | 180,451 | 1,980,136 | 0.0045742 | 0.0135215 | 0.0075821 | 5,019 |
| PARVA | 2,313,442 | 2,094,473 | 3,017,466 | 0.0060141 | 0.0080188 | 0.0021427 | 55,742 |
| PCNA | 2,300,166 | 2,407,186 | 2,057,527 | 0.0170402 | 0.0125668 | 0.0210382 | 5,111 |
| CAMK2D | 2,259,943 | 2,712,418 | 2,455,167 | 0.0133284 | 0.0063681 | 0.0076605 | 817 |
| SPARC | 2,230,562 | 2,335,873 | 2,990,594 | 0.0204267 | 0.0152481 | 0.0060567 | 6,678 |
| KIF3A | 219,478 | 1,850,005 | 2,377,716 | 0.0242452 | 0.04543 | 0.0142647 | 11,127 |
| TGFBR1 | 2,188,996 | 1,585,418 | 146,801 | 0.026412 | 0.0065456 | 0.0556766 | 7,046 |
| PKP2 | 2,108,275 | 183,479 | 2,092,256 | 0.0277094 | 0.0459483 | 0.0231586 | 5,318 |
| OSR1 | 2,071,946 | 4,175,767 | 2,370,344 | 0.0060753 | 0.0009141 | 0.0029158 | 130,497 |
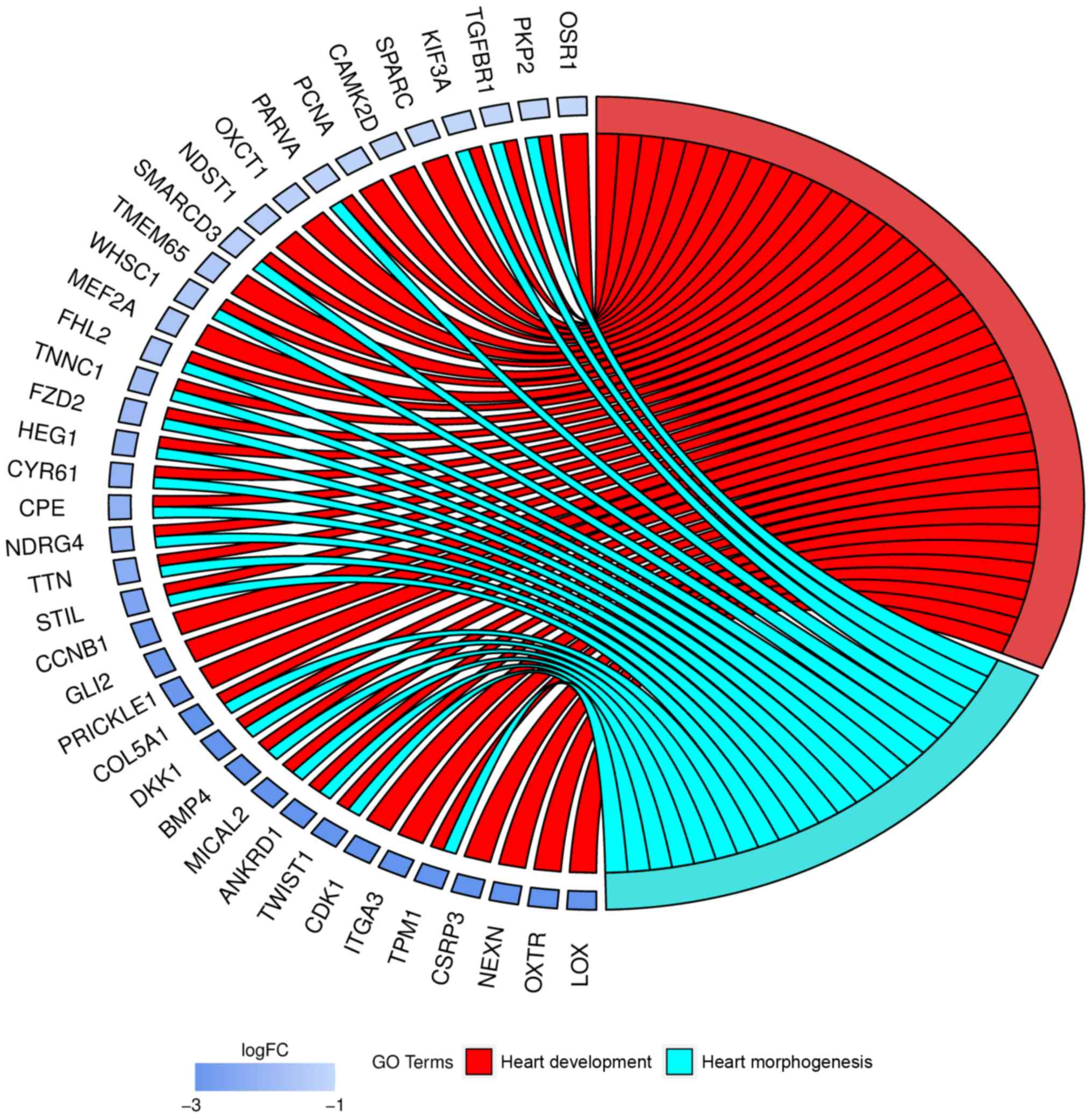
Moreover, genes that formed one particular GO group
can also belong to other different GO term categories. Because of
that, we explored the gene intersections between selected GO BP
terms. The relation between those GO BP terms was presented as
circle plot (Fig. 2).
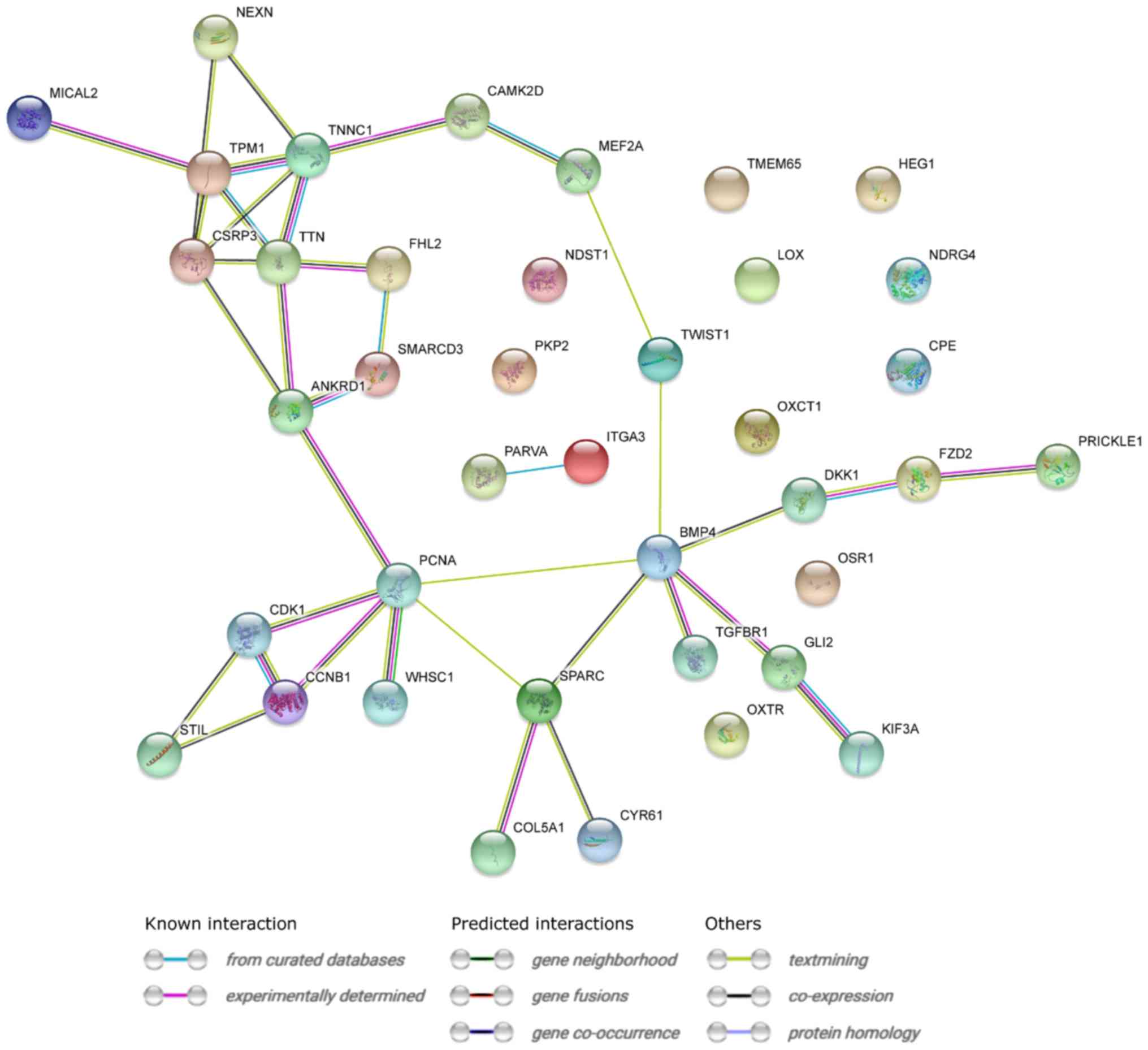
STRING interaction network was generated among
differentially expressed genes belonging to each of selected GO BP
terms. Using this prediction method yielded a molecular interaction
network formed between protein products of studied genes (Fig. 3).
RT-qPCR was conducted to validate the results
obtained during microarray analysis. The outcomes were presented
and compared in a form of a bar graph (Fig. 4).
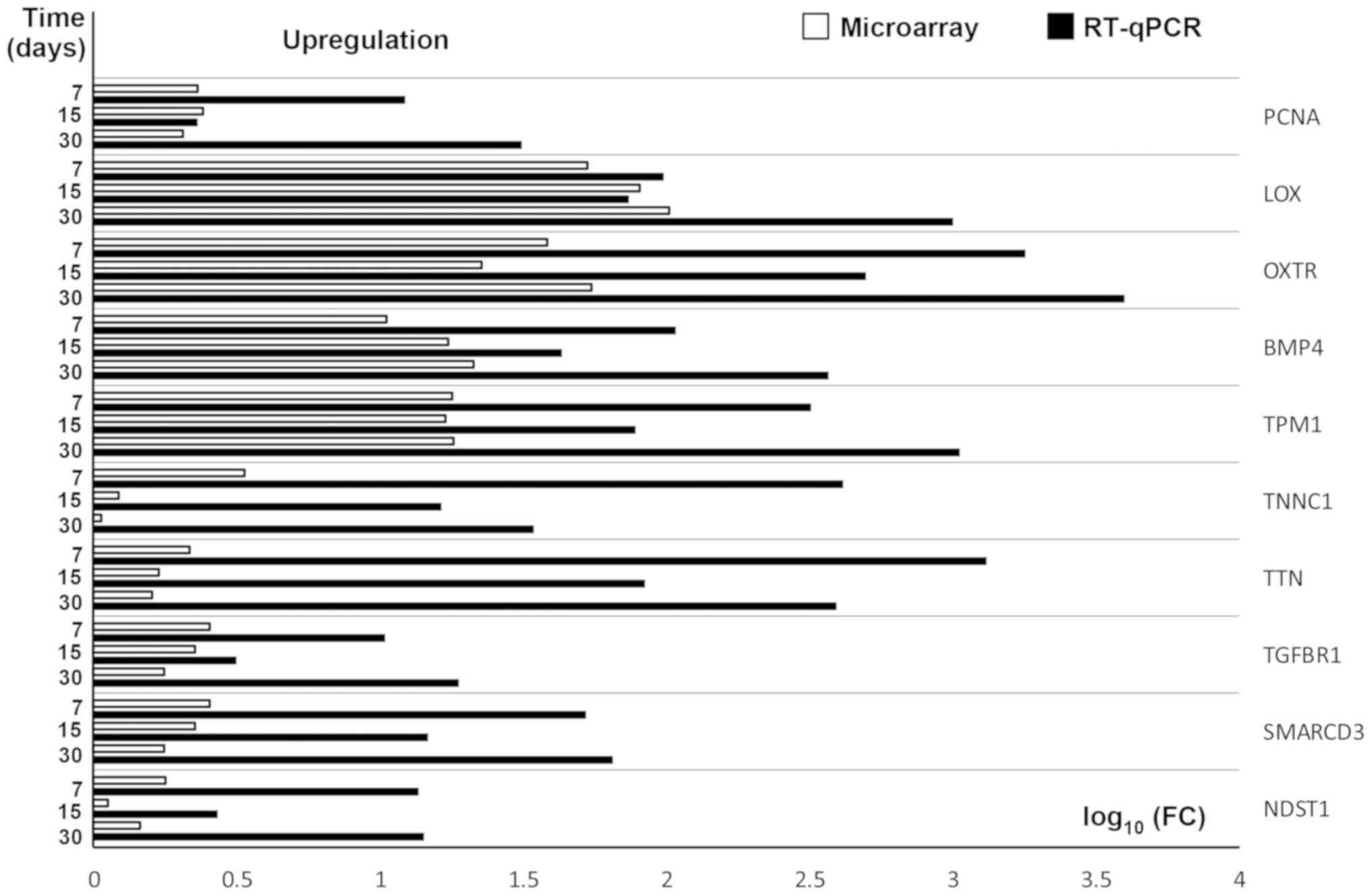 | Figure 4.RT-qPCR validation of microarray
results, presented in a form of a bar graph. FC was presented in
its logarithmic form to provide clear comparability of the results.
PCNA, Proliferating cell nuclear antigen; LOX, Lysyl Oxidase; OXTR,
oxytocin receptor; BMP4, Bone morphogenetic protein 4; TPM1,
Tropomyosin 1; TNNC1, Troponin 1; TTN, Titin; TGFBR1, Transcription
growth factor-β receptor 1; SMARCD3, SWI/SNF-related
matrix-associated actin-dependent regulator of chromatin subfamily
D member 3; NDST1, N-deacetylase and N-sulfotransferase 1; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction; FC,
fold change. |
As can be seen, the direction of changes in
expression was confirmed in all examples. However, the scale of
differences in transcript levels varied between both of the methods
analysed.
The GCs have undergone extensive morphological
modifications through the time of the culture. They changed their
shape from star-like structures to broader, fibroblast-like form.
These observable changes were presented as photographs, taken under
an inverted microscope, using a relief contrast (Fig. 5).
Discussion
Ovarian follicles are formed during the process of
folliculogenesis and oogenesis. The follicle consists of the
oocyte, follicular fluid and oocyte-associated cells-granulosa
cells (GCs) (10). In
physiological conditions, GCs largely influence the growth and
development of the ovarian follicle. In addition, they produce gap
junctions that allow for their bi-directional communication with
the oocyte (11,12). During this dialogue, nutrients and
metabolites are delivered to the oocyte (13). In addition, the key role of GCs in
physiological conditions is the production of estradiol during
follicular growth and the secretion of progesterone after ovulation
(14).
The process of folliculogenesis begins with the
change of GCs' shape from flattened to cuboidal through numerous
mitotic divisions of cells. A characteristic feature is the
production of follicle stimulating hormone receptor (FSHR) by the
GCs. In preovulatory follicle, four types of GCs can be
distinguished: The outermost layer is the granulosa membrane, the
periantral granulosa is located inside the follicle, the
intermediate layer forms the cumulus oophorus, with
corona radiata being the final layer. Granulosa cells
forming the corona radiata directly surround the oocyte
(15). The individual types of GCs
forming the four layers are functionally different, secreting
different molecules and expressing different receptors (16).
Recent studies indicate that there is a pool of stem
cells in mature mouse and human ovaries that support oogenesis and
folliculogenesis. Thanks to these stem cells, the primary
follicular resource can be produced during the reproductive period.
Previously, it was thought that the number of primary follicles is
determined and established shortly before the birth of the female
(17,18). In recent years, however, the
mentioned stem cells have become the major focus of scientists. It
is proven that every tissue or organ (including the ovary) contains
a population of stem cells capable of differentiating (19–22).
Many authors point to new possibilities of using GCs
in regenerative medicine and broadly understood transplantology. GC
containing follicular fluid, transferred for utilization during
routine in vitro fertilization procedures, may be the source
of stem cells. Several authors clearly indicate these previously
unnoticed properties of GCs. Many articles follow a different lead,
suggesting that GCs are subject to the transdifferentiation
process. Transdifferentiation is the transformation of already
differentiated cells into another cell type, which consists of two
stages: cells dedifferentiation, followed by the natural
developmental path towards a specific cell lineage. During this
process, the mature cell is reprogrammed and changes into another
cell type under the influence of the appropriate factors (23–25).
Kossowska-Tomaszczuk et al (3,4)
conducted experiments in which they indicated that GCs have
stem-like properties. Luteinized GC subpopulations express the
OCT-4 gene, escalation of which is characteristic for
mesenchymal stem cells. In addition, the research team carried out
the differentiation of GCs cells towards osteoblasts, neuroblasts
and chondroblasts. They succeeded to obtain populations of
differentiated cells (3,4). The results of our research indicate
that GCs have stem cell properties and can differentiate, in the
long-term primary in vitro culture, towards e.g.:
osteoblasts or muscle cells (unpublished results). The results of
our research have been confirmed by the literature. Oki et
al (26) stated that
pig-derived GCs may transdifferentiate towards osteoblasts both
in vitro and in vivo. They conducted a microarray
analysis clearly indicating that GCs show the GC-specific marker
genes, together with osteogenic markers, during
transdifferentiation (26).
Brevini et al (27) proved
that GCs, under the influence of external factors (Vascular
endothelial growth factor-VEGF and 5-azacytidine-5-aza-CR), can
differentiate towards muscle cells. Their research confirms the
change in the phenotype of GCs. In addition, the differentiated GCs
expressed the genes characteristic for muscle cells (including
Desmin and Myosin heavy chain).
Following up on Brevini et al (27), we have analyzed the results of
microarrays in terms of the possibility of GC differentiation into
cardiomyocytes. It turned out that, in long-term in vitro
culture, GCs express some genes involved in the process of heart
morphogenesis and development.
In our research, we identified 39 genes responsible
for heart development and morphogenesis processes. All of these
genes are presented on heatmaps (Fig.
1). Among all genes, 22 are involved in both: Heart development
and heart morphogenesis ontology groups. The remaining 17 genes are
characteristic for the heart development process. The expression of
these genes could confirm the theory that GCs can differentiate
into cells proprietary to the heart tissue, during in vitro
cell culture.
It is known that the heart is formed from the
mesoderm that arises during the process of embryo gastrulation. In
humans, the mesoderm begins to form at the third week of embryo
development (28). At this stage,
the mesodermal cells are still precardiac cells. Mesodermal cells
differentiate into cardiac cells in response to endodermal
induction signals-primarily bone morphogenetic proteins (BMPs)
(29). Five transcription factors
that play a primary role in cardiac development in various
organisms have been identified as NKX2.5, Mef2, GATA, Tbx and Hand
(30). Precardiac cells are
multipotent and differentiate in myocardial, endothelial and smooth
muscle cells through a process called progressive lineage
restriction (31). The last stage
of cardiac development is the differentiation of the atrial and
ventricular specific myocytes and heart conduction cells (32). In our studies, we showed the
expression of the Mef2A gene, and therefore suggest that
GCs, in long-term in vitro culture, show features of
mesenchymal stem cells that can differentiate into heart cells.
In addition, this is confirmed by the number of
studies that describe the transition of multipotent mesodermal
precursor cells into myoblasts, and then into mature myocytes. The
myocyte-specific enhancer factor 2 (MEF2) (33) is responsible for the above process.
Xu et al (34), in their
studies, proved that overexpression of Mef2A causes
cardiomyopathy and hypertension. Liu et al (35) demonstrated that Mef2 becomes
active during heart formation in the early stages of embryogenesis
and is responsible for the differentiation of atria and ventricles
of the heart. Other studies demonstrate the key role of
Mef2A in maintaining the appropriate number of mitochondria
and the proper architecture of the heart's cells. Mef2a-deficient
mice showed high mortality within one week after birth and marked
dilation of the ventricle. These mice have been proven to be
susceptible to sudden death (36).
The above studies indicate the key role played by Mef2a in
the formation of the heart. This, together with the findings of our
research that shows that GCs can give rise to heart cells, opens up
a way for further research into the application of human granulosa
in heart regeneration medicine. As it has already been mentioned,
inducers of the endoderm, such as BMPs, are an important group of
factors involved in the development of the heart. In long-term
in vitro culture, GCs express the BMP4 gene.
According to the literature, this protein is involved in the
development of endocardial epithelium and the correct formation of
particular chambers of the heart. Deficiency of BMP4 in mice
causes an abnormal number of cardiac cells, defective valve
structure, defects in the septum of the heart, and abnormal heart
valve formation (37,38).
Another gene characteristic for both morphogenesis
and heart development is LOX (Lysyl oxidase). This
gene shows the biggest change in expression in relation to the
initial condition during long-term in vitro culture. Li
et al (39) demonstrated
that lysyl oxidase is present in the smooth muscle cells of
the blood vessels of rats. In addition, Kaneda et al
(40) noticed that LOX may
be a suppressor of ovarian or stomach tumours. Our research
indicates that this gene is expressed in granulosa cells and thus
they could possibly differentiate into the blood vessels of the
heart and be a suppressor of pathological changes in the ovary.
The gene responsible for the formation of the
oxytocin receptor (oxytocin receptor-OXTR) showed
prominently increased expression. To date, the expression of this
gene has been associated with fertility, lactation and reproductive
behaviour. Recently, mice research has suggested that OXTR
plays a large role in the regulation and development of social
behaviour (41). In addition, in
recent years, the role of the above gene in the formation of the
cardiovascular system has been indicated (42).
Another gene characteristic for the development and
morphogenesis of the heart is NEXN (Nexilin F-actin
binding protein) and Cysteine-rich protein 3
(CSRP3) also known as Muscle LIM protein
(MLP). These genes show activity in the fetal and adult
heart, as well as skeletal muscle adhesion and migration during
embryogenesis, and are responsible for hyperthyroid cardiomyopathy
(43–45).
During the long-term in vitro cultivation of
GCs, it was noted that these cells show expression of
cyclin-dependent kinases (CDKs), specifically CDK1. These
compounds are responsible for the regulation of the cell cycle of
cardiomyocytes (46). This serves
as further evidence that in vitro, GCs may be differentiated
into cardiomyocytes. The above genes showed clearly increased
expression during long-term in vitro GC culture.
Among the genes characteristic for the development
and morphogenesis of the heart, none showed a drastic decline
during long-term in vitro culture. Some of these genes
showed only a smaller change in expression. These include OSR1,
PKP2, TGFBR1, KIF3A.
The OSR1 gene is expressed in GCs under
physiological conditions but also participates in the formation of
cardiac septa (47,48). We can assume that even a small
change in the expression of this gene may indicate the possibility
of GC differentiation. Similar observations were found in the case
of the PKP2 gene (plakophilin). Plakophilin is a
component of desmosomes and is necessary for the proper functioning
and distribution of connexins, a type gap junction protein. It was
noted that, during the silencing of the PKP2 gene in rat
cardiac myocytes, there was a reduction in the number of gap
junctions between the heart cells. This led to changes within the
right ventricle (49). Other
studies indicate that connexin is also present in GCs and is
necessary for bi-directional communication between the oocyte and
granulosa cells (50).
Another gene that seems to be characteristic for the
processes related to the development of heart is TNNC1
(troponin c). This gene is responsible for the proper
functioning of the muscle fibres of the heart, with all of its
known mutations leading to cardiomyopathy (51). Another very important gene,
demonstrating that GCs in long-term in vitro culture can
undergo differentiation/transdifferentiation into cardiac muscle,
is SMARCD3 (actin-dependent regulator of chromatin,
subfamily D). Lickert et al (52) noticed that this gene is expressed
in the hearts of early mouse embryos. Deficiency of this gene
caused defects in heart morphogenesis, as well as abnormal
differentiation of myocardium and skeletal muscles. In humans, the
impairment of this gene is associated with the occurrence of many
congenital heart defects (52).
Other studies show that, in general, mesodermal cells may undergo
transdifferentiation towards cardiac myocytes. SMARCD3,
among others, is deeply involved in this process. It is noted that
the presence of this gene can cause the transdifferentiation of
mesoderm cells into cardiomyocytes (53). It can, therefore, be suggested that
GCs, in long-term in vitro culture, show stem-like potential
and may undergo differentiation or transdifferentiation towards
cardiomyocytes.
It may be concluded that GCs have many
characteristics of stem cells in long-term in vitro culture.
The presented results suggest that it is possible to differentiate
GCs into cells that build the heart and, at the same time, it may
become a new possibility in the treatment of heart diseases and
regenerative medicine of blood vessels. However, the conclusions
were based only on large-scale transcriptomic analysis. While
validated by more focused RT-qPCR tests in terms of the direction
of changes, the scale of differences in transcript levels varied
between the methods. This may be caused by many factors, mainly
associated with the huge array of processes occurring from the time
of transcription to expression of protein products. Therefore, the
conclusions of this study require further analyses on the protein
level, followed by practical trials of obtaining a stable,
differentiated population of cardiac cells coming from GC cultures,
which could later be applied in possible clinical situations.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Polish
National Science Centre (grant nos. 2014/15/B/NZ7/00999 and
UMO-2016/21/B/NZ9/03535) and from Poznan University of Medical
Sciences (grant no. 502-01-02227367-08414).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WK conducted the experiments, chose the models and
wrote the manuscript. MBrą designed the experiments and wrote the
manuscript. PC performed data analysis and figure preparation, and
wrote the manuscript. ArB conducted the experiments, performed data
analysis and prepared the figures. MJN wrote the manuscript and
performed data analysis. KO conducted the experiments, revised the
medical methodology, and performed material extraction and
preparation. MaJ conducted the experiments, and performed data
analysis and language corrections. MiJ designed the experiments and
performed models analysis. LP designed the study and revised the
medical methodology (particularly that of in vitro
fertilization procedures). AnB designed the study, and revised the
methodology and the manuscript. DR designed the study. MTS designed
the study, revised the methodology and critically revised the
manuscript for important intellectual content. MBru designed the
experiments, supervised the project, was involved in the medical
procedure design and approved the final draft of the manuscript. MZ
revised the methodology, performed data analysis, provided writing
assistance and approved the final draft of the manuscript. MN
supervised the project, designed the experiments (particularly the
primary cell culture procedure), provided editorial supervision and
approved the final draft of the manuscript. BK supervised the
project, designed the study, revised the methodology, provided
editorial supervision, and wrote the manuscript. All authors read
and approved the final manuscript
Ethics approval and consent to
participate
The present study has been approved by Poznan
University of Medical Sciences Bioethical Committee with resolution
558/17. All patients provided written informed consent.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Rybska M, Knap S, Jankowski M, Jeseta M,
Bukowska D, Antosik P, Nowicki M, Zabel M, Kempisty B and Jaśkowski
JM: Cytoplasmic and nuclear maturation of oocytes in mammals-living
in the shadow of cells developmental capability. Med J Cell Biol.
6:13–17. 2018. View Article : Google Scholar
|
|
2
|
Rybska M, Knap S, Jankowski M, Jeseta M,
Bukowska D, Antosik P, Nowicki M, Zabel M, Kempisty B and Jaśkowski
JM: Characteristic of factors influencing the proper course of
folliculogenesis in mammals. Med J Cell Biol. 6:33–38. 2018.
View Article : Google Scholar
|
|
3
|
Kossowska-Tomaszczuk K, De Geyter C, De
Geyter M, Martin I, Holzgreve W, Scherberich A and Zhang H: The
multipotency of luteinizing granulosa cells collected from mature
ovarian follicles. Stem Cells. 27:210–219. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kossowska-Tomaszczuk K and De Geyter C:
Cells with stem cell characteristics in somatic compartments of the
ovary. Biomed Res Int 2013. 3108592013.
|
|
5
|
Kranc W, Brązert M, Ożegowska K, Nawrocki
MJ, Budna J, Celichowski P, Dyszkiewicz-Konwińska M, Jankowski M,
Jeseta M, Pawelczyk L, et al: Expression profile of genes
regulating steroid biosynthesis and metabolism in human ovarian
granulosa cells-a primary culture approach. Int J Mol Sci. 18(pii):
E26732017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chomczynski P and Sacchi N: Single-step
method of RNA isolation by acid guanidinium
thiocyanate-phenol-chloroform extraction. Anal Biochem.
162:156–159. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang DW, Sherman BT, Tan Q, Kir J, Liu D,
Bryant D, Guo Y, Stephens R, Baseler MW, Lane HC and Lempicki RA:
DAVID bioinformatics resources: Expanded annotation database and
novel algorithms to better extract biology from large gene lists.
Nucleic Acids Res. 35:W169–W175. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Walter W, Sánchez-Cabo F and Ricote M:
GOplot: An R package for visually combining expression data with
functional analysis. Bioinformatics. 31:2912–2914. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
von Mering C, Jensen LJ, Snel B, Hooper
SD, Krupp M, Foglierini M, Jouffre N, Huynen MA and Bork P: STRING:
Known and predicted protein-protein associations, integrated and
transferred across organisms. Nucleic Acids Res. 33:D433–D437.
2004. View Article : Google Scholar :
|
|
10
|
Kranc W, Budna J, Kahan R, Chachuła A,
Bryja A, Ciesiółka S, Borys S, Antosik MP, Bukowska D, Brussow KP,
et al: Molecular basis of growth, proliferation, and
differentiation of mammalian follicular granulosa cells. J Biol
Regul Homeost Agents. 31:1–8. 2017.PubMed/NCBI
|
|
11
|
Mora JM, Fenwick MA, Castle L, Baithun M,
Ryder TA, Mobberley M, Carzaniga R, Franks S and Hardy K:
Characterization and significance of adhesion and junction-related
proteins in mouse ovarian follicles. Biol Reprod. 86(153): 1–14.
2012.
|
|
12
|
Kempisty B, Ziółkowska A, Piotrowska H,
Zawierucha P, Antosik P, Bukowska D, Ciesiółka S, Jaśkowski JM,
Brüssow KP, Nowicki M and Zabel M: Real-time proliferation of
porcine cumulus cells is related to the protein levels and cellular
distribution of Cdk4 and Cx43. Theriogenology. 80:411–420. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Su YQ, Sugiura K and Eppig JJ: Mouse
oocyte control of granulosa cell development and function:
Paracrine regulation of cumulus cell metabolism. Semin Reprod Med.
27:32–42. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Jeppesen JV, Kristensen SG, Nielsen ME,
Humaidan P, Dal Canto M, Fadini R, Schmidt KT, Ernst E and Yding
Andersen C: LH-receptor gene expression in human granulosa and
cumulus cells from antral and preovulatory follicles. J Clin
Endocrinol Metab. 97:E1524–E1531. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Findlay JK, Kerr JB, Britt K, Liew SH,
Simpson ER, Rosairo D and Drummond AE: Ovarian physiology: Follicle
development, oocyte and hormone relationships. Anim Reprod.
6:16–19. 2009.
|
|
16
|
Nguyen T, Lee S, Hatzirodos N, Hummitzsch
K, Sullivan TR, Rodgers RJ and Irving-Rodgers HF: Spatial
differences within the membrana granulosa in the expression of
focimatrix and steroidogenic capacity. Mol Cell Endocrinol.
363:62–73. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guigon CJ and Magre S: Contribution of
germ cells to the differentiation and maturation of the ovary:
Insights from models of germ cell depletion. Biol Reprod.
74:450–458. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Johnson J, Canning J, Kaneko T, Pru JK and
Tilly JL: Germline stem cells and follicular renewal in the
postnatal mammalian ovary. Nature. 428:145–150. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Parte S, Bhartiya D, Telang J, Daithankar
V, Salvi V, Zaveri K and Hinduja I: Detection, characterization,
and spontaneous differentiation in vitro of very small
embryonic-like putative stem cells in adult mammalian ovary. Stem
Cells Dev. 20:1451–1464. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Virant-Klun I, Rozman P, Cvjeticanin B,
Vrtacnik-Bokal E, Novakovic S, Rülicke T, Dovc P and Meden-Vrtovec
H: Parthenogenetic embryo-like structures in the human ovarian
surface epithelium cell culture in postmenopausal women with no
naturally present follicles and oocytes. Stem Cells Dev.
18:137–149. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Virant-Klun I, Zech N, Rozman P, Vogler A,
Cvjeticanin B, Klemenc P, Malicev E and Meden-Vrtovec H: Putative
stem cells with an embryonic character isolated from the ovarian
surface epithelium of women with no naturally present follicles and
oocytes. Differentiation. 76:843–856. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Virant-Klun I: Postnatal oogenesis in
humans: A review of recent findings. Stem Cells Cloning. 8:49–60.
2015.PubMed/NCBI
|
|
23
|
Buganim Y and Jaenisch R:
Transdifferentiation by defined factors as a powerful research tool
to address basic biological questions. Cell Cycle. 11:4485–4486.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shen CN, Burke ZD and Tosh D:
Transdifferentiation, metaplasia and tissue regeneration.
Organogenesis. 1:36–44. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dzafic E, Stimpfel M and Virant-Klun I:
Plasticity of granulosa cells: On the crossroad of stemness and
transdifferentiation potential. J Assist Reprod Genet.
30:1255–1261. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oki Y, Ono H, Motohashi T, Sugiura N,
Nobusue H and Kano K: Dedifferentiated follicular granulosa cells
derived from pig ovary can transdifferentiate into osteoblasts.
Biochem J. 447:239–248. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Brevini TAL, Pennarossa G, Rahman MM,
Paffoni A, Antonini S, Ragni G, deEguileor M, Tettamanti G and
Gandolfi F: Morphological and molecular changes of human granulosa
cells exposed to 5-azacytidine and addressed toward muscular
differentiation. Stem Cell Rev. 10:633–642. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moorman A, Webb S, Brown NA, Lamers W and
Anderson RH: Development of the heart: (1) formation of the cardiac
chambers and arterial trunks. Heart. 89:806–814. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Meilhac SM, Lescroart F, Blanpain C and
Buckingham ME: Cardiac cell lineages that form the heart. Cold
Spring Harb Perspect Med. 4:a0138882014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Olson EN: Gene regulatory networks in the
evolution and development of the heart. Science. 313:1922–1927.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Epstein JA: Franklin H. Epstein Lecture.
Cardiac development and implications for heart disease. N Engl J
Med. 363:1638–1647. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wu SM, Fujiwara Y, Cibulsky SM, Clapham
DE, Lien CL, Schultheiss TM and Orkin SH: Developmental origin of a
bipotential myocardial and smooth muscle cell precursor in the
mammalian heart. Cell. 127:1137–1150. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Breitbart RE, Liang CS, Smoot LB, Laheru
DA, Mahdavi V and Nadal-Ginard B: A fourth human MEF2 transcription
factor, hMEF2D, is an early marker of the myogenic lineage.
Development. 118:1095–1106. 1993.PubMed/NCBI
|
|
34
|
Xu J, Gong NL, Bodi I, Aronow BJ, Backx PH
and Molkentin JD: Myocyte enhancer factors 2A and 2C induce dilated
cardiomyopathy in transgenic mice. J Biol Chem. 281:9152–9162.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu N, Williams AH, Kim Y, McAnally J,
Bezprozvannaya S, Sutherland LB, Richardson JA, Bassel-Duby R and
Olson EN: An intragenic MEF2-dependent enhancer directs
muscle-specific expression of microRNAs 1 and 133. Proc Natl Acad
Sci USA. 104:20844–20849. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Naya FJ, Black BL, Wu H, Bassel-Duby R,
Richardson JA, Hill JA and Olson EN: Mitochondrial deficiency and
cardiac sudden death in mice lacking the MEF2A transcription
factor. Nat Med. 8:1303–1309. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
37
|
McCulley DJ, Kang JO, Martin JF and Black
BL: BMP4 is required in the anterior heart field and its
derivatives for endocardial cushion remodeling, outflow tract
septation, and semilunar valve development. Dev Dyn. 237:3200–3209.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Abdelwahid E, Rice D, Pelliniemi LJ and
Jokinen E: Overlapping and differential localization of Bmp-2,
Bmp-4, Msx-2 and apoptosis in the endocardial cushion and adjacent
tissues of the developing mouse heart. Cell Tissue Res. 305:67–78.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li W, Nellaiappan K, Strassmaier T, Graham
L, Thomas KM and Kagan HM: Localization and activity of lysyl
oxidase within nuclei of fibrogenic cells. Proc Natl Acad Sci USA.
94:12817–12822. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kaneda A, Wakazono K, Tsukamoto T,
Watanabe N, Yagi Y, Tatematsu M, Kaminishi M, Sugimura T and
Ushijima T: Lysyl oxidase is a tumor suppressor gene inactivated by
methylation and loss of heterozygosity in human gastric cancers.
Cancer Res. 64:6410–6415. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Takayanagi Y, Yoshida M, Bielsky IF, Ross
HE, Kawamata M, Onaka T, Yanagisawa T, Kimura T, Matzuk MM, Young
LJ and Nishimori K: Pervasive social deficits, but normal
parturition, in oxytocin receptor-deficient mice. Proc Natl Acad
Sci USA. 102:16096–16101. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gutkowska J and Jankowski M: Oxytocin
revisited: Its role in cardiovascular regulation. J
Neuroendocrinol. 24:599–608. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Hassel D, Dahme T, Erdmann J, Meder B,
Huge A, Stoll M, Just S, Hess A, Ehlermann P, Weichenhan D, et al:
Nexilin mutations destabilize cardiac Z-disks and lead to dilated
cardiomyopathy. Nat Med. 15:1281–1288. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Zhao Y, Wei YJ, Cao HQ and Ding JF:
Molecular cloning of NELIN, a putative human cytoskeleton
regulation gene. Sheng Wu Hua Xue Yu Sheng Wu Wu Li Xue Bao
(Shanghai). 33:19–24. 2001.PubMed/NCBI
|
|
45
|
Geier C, Gehmlich K, Ehler E, Hassfeld S,
Perrot A, Hayess K, Cardim N, Wenzel K, Erdmann B, Krackhardt F, et
al: Beyond the sarcomere: CSRP3 mutations cause hypertrophic
cardiomyopathy. Hum Mol Genet. 17:2753–2765. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ikenishi A, Okayama H, Iwamoto N,
Yoshitome S, Tane S, Nakamura K, Obayashi T, Hayashi T and Takeuchi
T: Cell cycle regulation in mouse heart during embryonic and
postnatal stages. Dev Growth Differ. 54:731–738. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lan CW, Chen MJ, Jan PS, Chen HF and Ho
HN: Differentiation of human embryonic stem cells into functional
ovarian granulosa-like cells. J Clin Endocrinol Metab.
98:3713–3723. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Zhou L, Liu J, Olson P, Zhang K, Wynne J
and Xie L: Tbx5 and Osr1 interact to regulate posterior second
heart field cell cycle progression for cardiac septation. J Mol
Cell Cardiol. 85:1–12. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Oxford EM, Musa H, Maass K, Coombs W,
Taffet SM and Delmar M: Connexin43 remodeling caused by inhibition
of plakophilin-2 expression in cardiac cells. Circ Res.
101:703–711. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Kempisty B, Ziółkowska A, Ciesiółka S,
Piotrowska H, Antosik P, Bukowska D, Nowicki M, Brüssow KP and
Zabel M: Study on connexin gene and protein expression and cellular
distribution in relation to real-time proliferation of porcine
granulosa cells. J Biol Regul Homeost Agents. 28:625–635.
2014.PubMed/NCBI
|
|
51
|
Parvatiyar MS, Landstrom AP,
Figueiredo-Freitas C, Potter JD, Ackerman MJ and Pinto JR: A
mutation in TNNC1-encoded cardiac troponin C, TNNC1-A31S,
predisposes to hypertrophic cardiomyopathy and ventricular
fibrillation. J Biol Chem. 287:31845–31855. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Lickert H, Takeuchi JK, Von Both I, Walls
JR, McAuliffe F, Adamson SL, Henkelman RM, Wrana JL, Rossant J and
Bruneau BG: Baf60c is essential for function of BAF chromatin
remodelling complexes in heart development. Nature. 432:107–112.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Takeuchi JK and Bruneau BG: Directed
transdifferentiation of mouse mesoderm to heart tissue by defined
factors. Nature. 459:708–711. 2009. View Article : Google Scholar : PubMed/NCBI
|