Introduction
Lung cancer is the most commonly diagnosed cancer
(11.6% of the total cases) and the leading cause of
cancer-associated mortality (18.4% of the total cancer-associated
mortality cases) globally (1).
Non-small cell lung cancer (NSCLC) accounts for ~83% of lung cancer
cases, and the majority of patients with advanced NSCLC are treated
with chemotherapy (2).
Platinum-based drugs, particularly cisplatin (Cis), are used in the
treatment of numerous cancer types, including NSCLC (3). Cis is the most widely used drug in
cancer therapy and the first Food and Drug Administration-approved
platinum compound for lung cancer treatment (4,5).
However, the use of Cis in lung cancer chemotherapy is limited by
the resistance of cells to the drug (6–8). In
order to enhance the therapeutic efficacy of currently available
cytotoxic drugs and to identify novel anti-tumor drugs, it is
important to identify alternative strategies to overcome Cis
resistance and identify novel therapies.
Natural products are promising sources of anticancer
agents and contribute substantially to cancer therapy (9). Gambogenic acid (GNA), a natural
compound derived from gamboge, has long been used in traditional
Chinese medicine (10). GNA exerts
cytotoxicity in numerous human cancer lines (11–16).
In addition, previous studies have demonstrated that GNA may also
enhance chemosensitivity in certain cancer cell types (17,18).
However, to the best of our knowledge, it is not known whether GNA
serves a function in the treatment of Cis-resistant lung cancer.
The chemical structure of GNA has an active bond (19), which may bind with the target
protein, but the detailed molecular mechanism underlying its
anticancer effects have not yet been fully elucidated. The present
study investigated the anticancer effects and mechanisms of GNA on
Cis-resistant NSCLC cells. The present results suggested that GNA
efficiently inhibited the proliferation of Cis-resistant NSCLC
cells in vitro, highlighting the potential application of
GNA in NSCLC clinical studies.
Materials and methods
Cell culture
The human NSCLC cell line A549 and A549/Cis cells
were obtained from The Cell Resource Center, Institute of Basic
Medicine, Chinese Academy of Medical Sciences. The cells were
cultured in DMEM (Corning Inc.), supplemented with 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) and 1% penicillin/streptomycin
(Gibco; Thermo Fisher Scientific, Inc.). All cells were cultured at
37°C and 5% CO2. A549/Cis cells were cultured in the
presence of 2 µM Cis (Selleck Chemicals), which were collected
subsequent to two generations prior to the experiments being
performed. These cell lines grew in monolayers and when the
confluence reached 70–80%, they were passaged.
MTT assay
Briefly, A549 and A549/Cis cells (3×103
cells/well) were added to 96-well plates in 100 µl cell culture
medium. The cells were treated with various concentrations of Cis
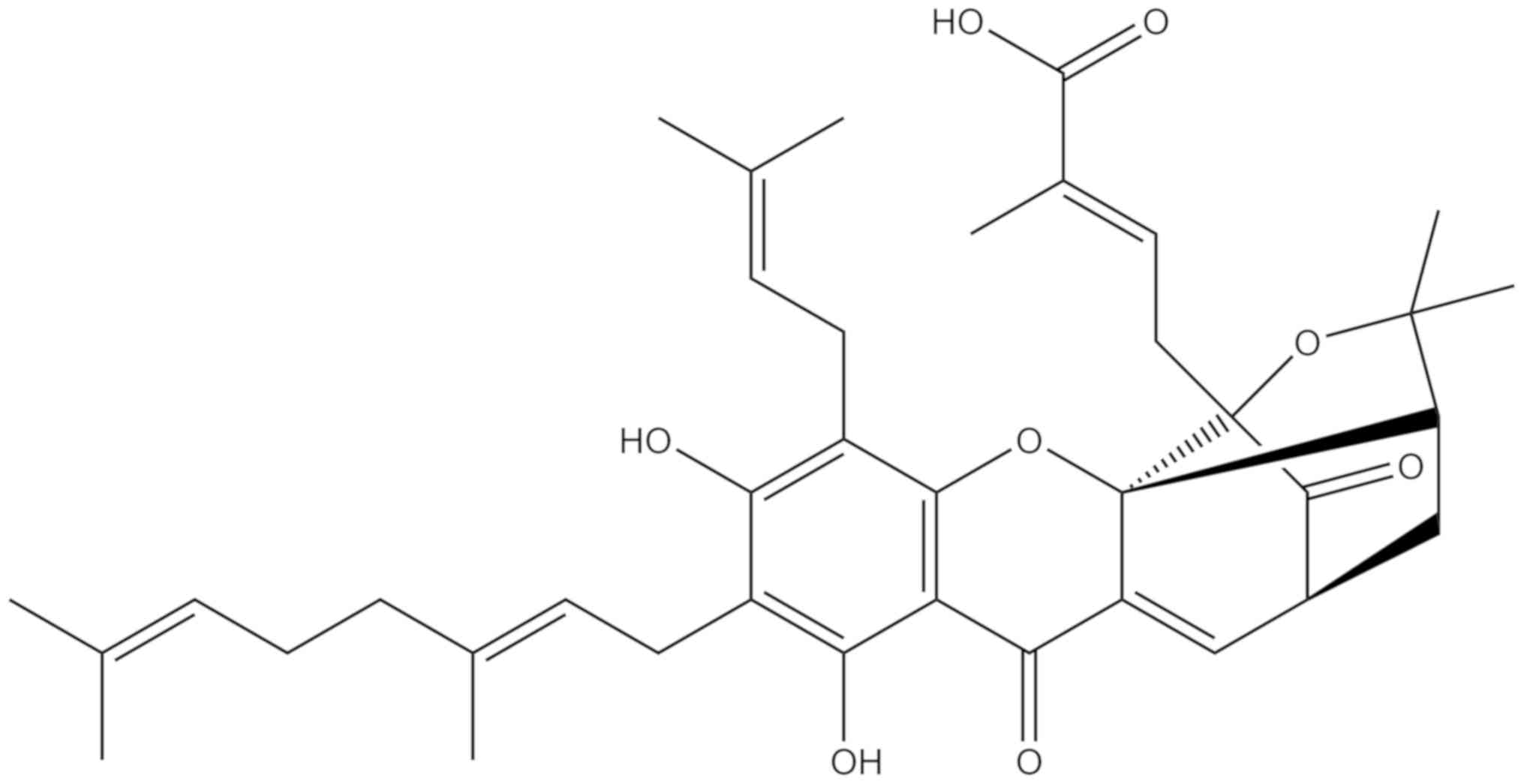
(0, 2.5, 5, 10 or 20 µM) or GNA (0, 0.5, 1, 2, 4 or 6 µM; Fig. 1; purity >98%; Nanjing Dasf
Biotechnology Co., Ltd.). GNA has poor aqueous solubility, and was
dissolved in DMSO as the stock solution at a concentration of 20
mM, and then diluted to working concentration as required in
complete culture medium immediately prior to use. The cells were
then incubated at 37°C and 5% CO2 for 24, 48 or 72 h.
Following the treatments, 20 µl/well MTT (Biosharp; 5 mg/ml) was
added for further incubation at 37°C for 4 h. The supernatant was
then discarded and the formazan product was dissolved in DMSO. The
optical density was measured at 490 nm in a Synergy H1 Hybrid
Multi-Mode Microplate Reader (BioTeke Coporation) and the
experimental results were recorded.
Hoechst 33342 staining analysis
Briefly, A549/Cis cells (5×104
cells/well) were seeded into 12-well plates and cultured at 37°C
for 24 h. Prior to GNA treatment, cells were observed by microscopy
and recorded as the initial point. Then, the cells were treated
with GNA (0, 2 or 4 µM) at 37°C for 24 and 48 h. At the end of the
various treatments, the cells were stained with Hoechst 33342 (10
µg/ml; Beijing Solarbio Science & Technology Co., Ltd.) for 20
min at 37°C and changes in the cell morphology were observed by
fluorescence microscopy (magnification, ×200; Leica Microsystems
GmbH).
Analysis of cell cycle and
apoptosis
The cell cycle and apoptosis were analyzed by flow
cytometry. For the cell cycle analysis, cells were harvested and
washed twice with cold phosphate buffered saline (PBS), then fixed
in precooled 70% alcohol overnight at −20°C. Subsequent to washing
in cold PBS 3 times, cells were incubated with RNase (Tiangen
Biotech Co., Ltd.; 0.5 mg/ml) solution at 37°C for 1 h.
Subsequently, propidium iodide (Beijing Solarbio Science &
Technology Co., Ltd.; 10 µg/ml) was added to the cells and
incubated at room temperature for 15 min. Then, the cell cycle
analysis was performed using a BD FACSCelesta Flow Cytometer (BD
Biosciences). The data were analyzed using Modifit LT 5.0 software
(Verity Software House, Inc.). Cell apoptosis was analyzed using an
Annexin V-APC/7-amino-actinomycin (7-AAD) Apoptosis Detection kit
(BioLegend, Inc.), according to the manufacturer's protocol. The
stained cells were analyzed using a BD FACSCelesta Flow Cytometer
(BD Biosciences) and the data were analyzed using FlowJo 7.6.1
software (FlowJo LLC).
RNA sequencing (RNA-seq)
The total RNA of A549/Cis cells treated with 4 µM
GNA for 24 h was extracted using TRIzol® reagent
(Invitrogen; Thermo Fisher Scientific, Inc.) following the
manufacturer's protocol. The RNA-seq was performed by LC-Bio
Technologies (Hangzhou) Co., Ltd. using an Illumina X10 (LC
Sciences) following the manufacturer's protocol. StringTie
(20) was used to analyze the
expression levels for mRNAs by calculating the fragments per
kilobase of transcript per million mapped reads. The differentially
expressed mRNAs and genes were selected with log2 (fold
change)>1 or log2 (fold change)<-1 and with statistical
significance (P-value <0.05) using the R package - Ballgown, as
described previously (21). For
the Gene Ontology enrichment (22,23)
and Kyoto Encyclopedia of Genes and Genomes (KEGG) (24–26)
pathway analysis, the Database for Annotation, Visualization and
Integrated Discovery web server (http://david.ncifcrf.gov/) was used (27,28).
RNA extraction and reverse
transcription-quantitative PCR (RT-qPCR)
A549/Cis cells were harvested 24 h after 4 µM GNA
treatment and the total RNA was extracted using the GeneJET RNA
Purification kit (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. RT was conducted using the First Strand
cDNA Synthesis kit (Thermo Fisher Scientific, Inc.). The following
RT temperature protocol was used: 65°C for 5 min, 50°C for 50 min
and 85°C for 5 min. qPCR was subsequently performed using the
SYBR® Select Master Mix kit (Thermo Fisher Scientific,
Inc.) and a QuantStudio 3 Real-Time PCR system (Thermo Fisher
Scientific, Inc.). The thermocycling conditions were as follows:
95°C for 10 min, followed by 40 cycles at 95°C for 15 sec and 60°C
for 1 min. The primer sequences used for the qPCR are listed in
Table I. Quantification was
performed using the comparative 2−∆∆Cq method (29). The Cq value for each sample was
normalized to the value of GAPDH. The experiments were performed in
triplicate.
 | Table I.Primer pairs for reverse
transcription-quantitative PCR. |
Table I.
Primer pairs for reverse
transcription-quantitative PCR.
| Gene | Primer sequence
(5′→3′) |
|---|
| GAPDH | F:
ACATCGCTCAGACACCATG |
|
| R:
TGTAGTTGAGGTCAATGAAGGG |
| GADD45A | F:
GGAGAGCAGAAGACCGAAAG |
|
| R:
AGGCACAACACCACGTTATC |
| CCND3 | F:
AACTGTGCATCTACACCGAC |
|
| R:
GCCAGGAAATCATGTGCAATC |
| CCNB1 | F:
GGCTTTCTCTGATGTAATTCTTGC |
|
| R:
GTATTTTGGTCTGACTGCTTGC |
| CDC20 | F:
GATGTAGAGGAAGCCAAGATCC |
|
| R:
AAGGAATGTAACGGCAGGTC |
| CDC25B | F:
CCGAGAGCTGATTGGAGATTAC |
|
| R:
CACGATGTTGCTGAACTTGC |
| PCNA | F:
GTCTCTTTGGTGCAGCTCA |
|
| R:
ATCTTCGGCCCTTAGTGTAATG |
| PLK1 | F:
ACAGTTTCGAGGTGGATGTG |
|
| R:
GGTTGATGTGCTTGGGAATAC |
| MCM2 | F:
ATTTCGTCCTGGGTCCTTTC |
|
| R:
CGCTGGTAGTTCTGATAGATGG |
| MCM3 | F:
AGCGAAGTGAGGATGAATCAG |
|
| R:
CTGTGTCACTGAAGTCATAGGG |
| MCM7 | F:
GATGCCACCTATACTTCTGCC |
|
| R:
TCCTTTGACATCTCCATTAGCC |
| SERPINE1 | F:
GTGGACTTTTCAGAGGTGGAG |
|
| R:
GAAGTAGAGGGCATTCACCAG |
| THBS1 | F:
CTCCCCTATGCTATCACAACG |
|
| R:
AGGAACTGTGGCATTGGAG |
| CASPASE7 | F:
CCTCGATACAAGATCCCAGTG |
|
| R:
GATTTCCAGGTCTTTTCCGTG |
| TNFRSF10B | F:
ACCACGACCAGAAACACAG |
|
| R:
CATTCGATGTCACTCCAGGG |
Western blotting
Total protein was extracted from cells using RIPA
lysis buffer (Beijing Solarbio Science & Technology Co., Ltd.)
which contains 1% (V/V) phenylmethanesulfonyl fluoride (Beijing
Solarbio Science & Technology Co., Ltd.). Nuclear proteins were
prepared using a Nuclear and Cytoplasmic Protein Extraction kit
(Wanleibio Co., Ltd.), according to the manufacturer's protocol. A
BCA protein assay kit (Thermo Fisher Scientific, Inc.) was used to
determine the protein concentration. Samples containing equal
amounts of protein (30 µg) were loaded onto 10% gels (p53, nuclear
p53, CDK6, Lamin B and GAPDH proteins) or 12% gels [p21, CDK4,
growth arrest and DNA damage-inducible α (GADD45A), cyclin D1,
cyclin D3, cyclin B1, caspase 3/7, cleaved-caspase 3/7,
poly(ADP-ribose) polymerase (PARP), cleaved-PARP and GAPDH
proteins], separated by SDS-PAGE and transferred to polyvinylidene
difluoride membranes (EMD Millipore). The membrane was blocked with
TBS with Tween-20 (1:1,000; TBST) containing 1% BSA (Beijing
Solarbio Science & Technology Co., Ltd.) for 1 h at room
temperature and incubated overnight at 4°C with specific primary
antibodies against p21 (1:1,000; cat. no. 2947; Cell Signaling
Technology, Inc.), GADD45A (1:1,000; cat. no. 4632; Cell Signaling
Technology, Inc.), cyclin D3 (1:2,000; cat. no. 2936; Cell
Signaling Technology, Inc.), cyclin D1 (1:500; cat. no. WL01435a;
Wanleibio Co., Ltd.), cyclin B1 (1:500; cat. no. WL01760; Wanleibio
Co., Ltd.), CDK4 (1:500; cat. no. WL02274; Wanleibio Co., Ltd.),
CDK6 (1:500; cat. no. WL01676; Wanleibio Co., Ltd.), caspase 3
(1:500; cat. no. WL01927; Wanleibio Co., Ltd.), cleaved-caspase 3
(1:500; cat. no. WL01992; Wanleibio Co., Ltd.), cleaved-caspase 7
(1:500; cat. no. WL02360; Wanleibio Co., Ltd.), PARP/cleaved-PARP
(1:500; cat. no. WL01932; Wanleibio Co., Ltd.), lamin B (1:500;
cat. no. WL01775; Wanleibio Co., Ltd.), caspase 7 (1:300; cat. no.
sc-56063; Santa Cruz Biotechnology, Inc.), p53 (1:500; cat. no.
bsm-33058M; BIOSS) and GAPDH (1:1,000; cat. no. bs-0755R; BIOSS).
Subsequent to washing three times with TBST, the membranes were
incubated with the following horseradish peroxidase
(HRP)-conjugated secondary antibodies: Goat anti-mouse (1:3,000;
cat. no. bs-0296G-HRP; BIOSS) and goat anti-rabbit (1:3,000; cat.
no. bs-0295G-HRP; BIOSS) for 2 h at room temperature. Following
washing in TBST three times, immunoblots were visualized using an
enhanced chemiluminescent substrate kit (Beijing Solarbio Science
& Technology Co., Ltd.) under an Amersham Imager 600 system (GE
Healthcare Life Sciences). Expression levels were quantified using
ImageJ version 1.52 software (National Institutes of Health).
Statistical analysis
All data are presented as the mean ± SEM, unless
otherwise stated, and represent three independent experimental
repeats. SPSS 20.0 software (IBM Corp.) and GraphPad Prism 5.0
software (GraphPad Software, Inc.) were used for statistical
analysis. One-way ANOVAs were conducted to examine the differences
among the multiple groups, followed by a Tukey's post-hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
GNA inhibits growth and induces the
apoptosis of A549/Cis cells
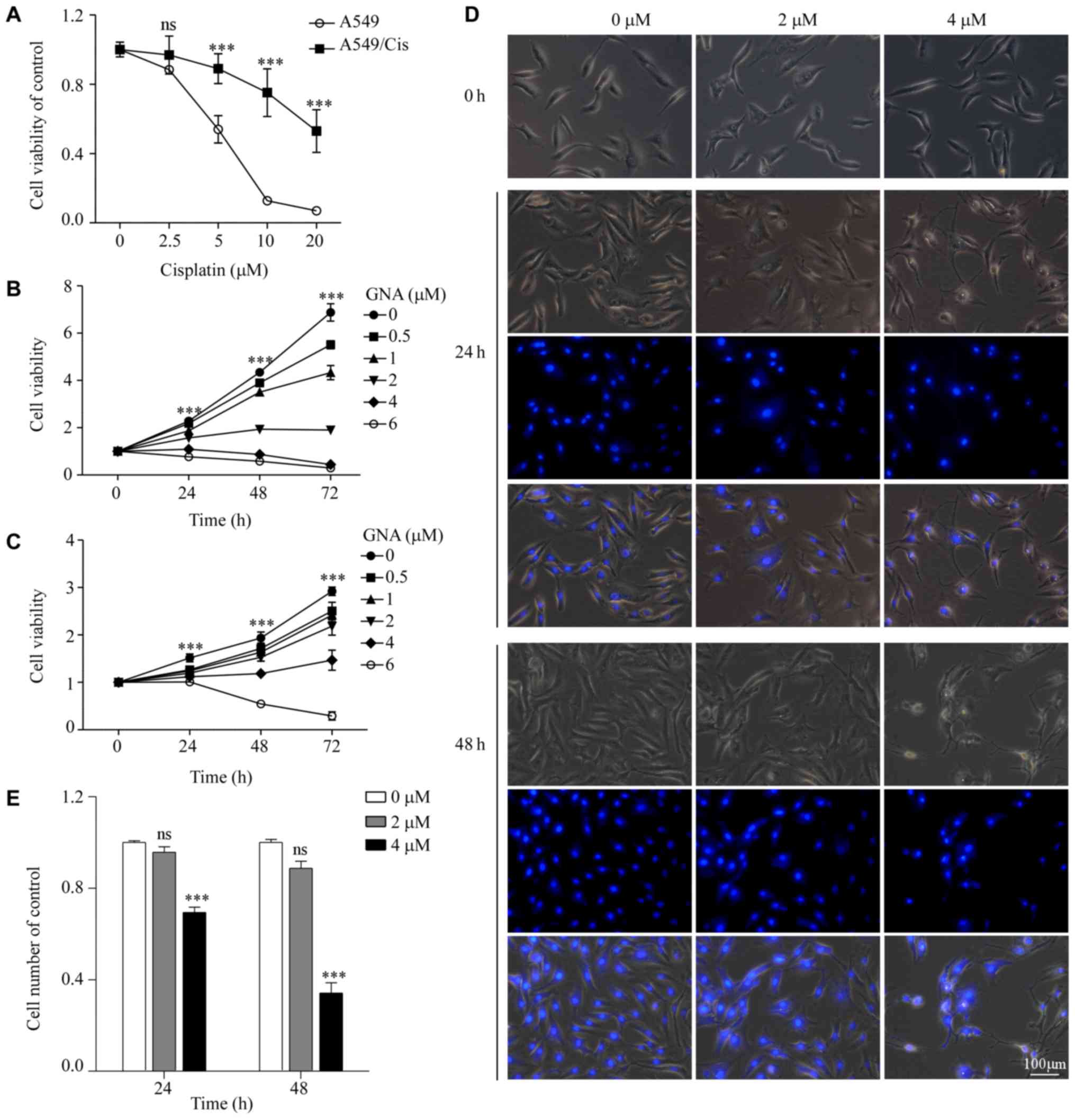
A549/Cis is a Cis-resistant cell line derived from
the A549 lung cancer cell line. To confirm its chemoresistance to
Cis, A549 cells were exposed to Cis in vitro to establish
A549/Cis cells. The MTT assay demonstrated that A549/Cis cells were
significantly more resistant to Cis compared with the parental
cells (P<0.001; Fig. 2A). The
cytotoxic effect of GNA on A549 and A549/Cis cells was determined.
Cells were treated with increasing concentrations of GNA for 24, 48
and 72 h. Cell viability was measured using an MTT assay. As
presented in Fig. 2B and C, GNA
significantly decreased the viability of A549 and A549/Cis cells
compared with the untreated group (P<0.001). GNA induced a high
degree of cell death at a concentration of 6 µM only after 24 h.
Accordingly, 2 and 4 µM GNA was used in the subsequent experiments.
Hoechst 33342 staining further demonstrated the inhibitory effect
of GNA in A549/Cis cells (Fig. 2C and
D). Compared with the untreated cells, the cells treated with
GNA had inhibited proliferation and exhibited morphological
alterations. Furthermore, the nuclear condensation of GNA-treated
cells was also observed.
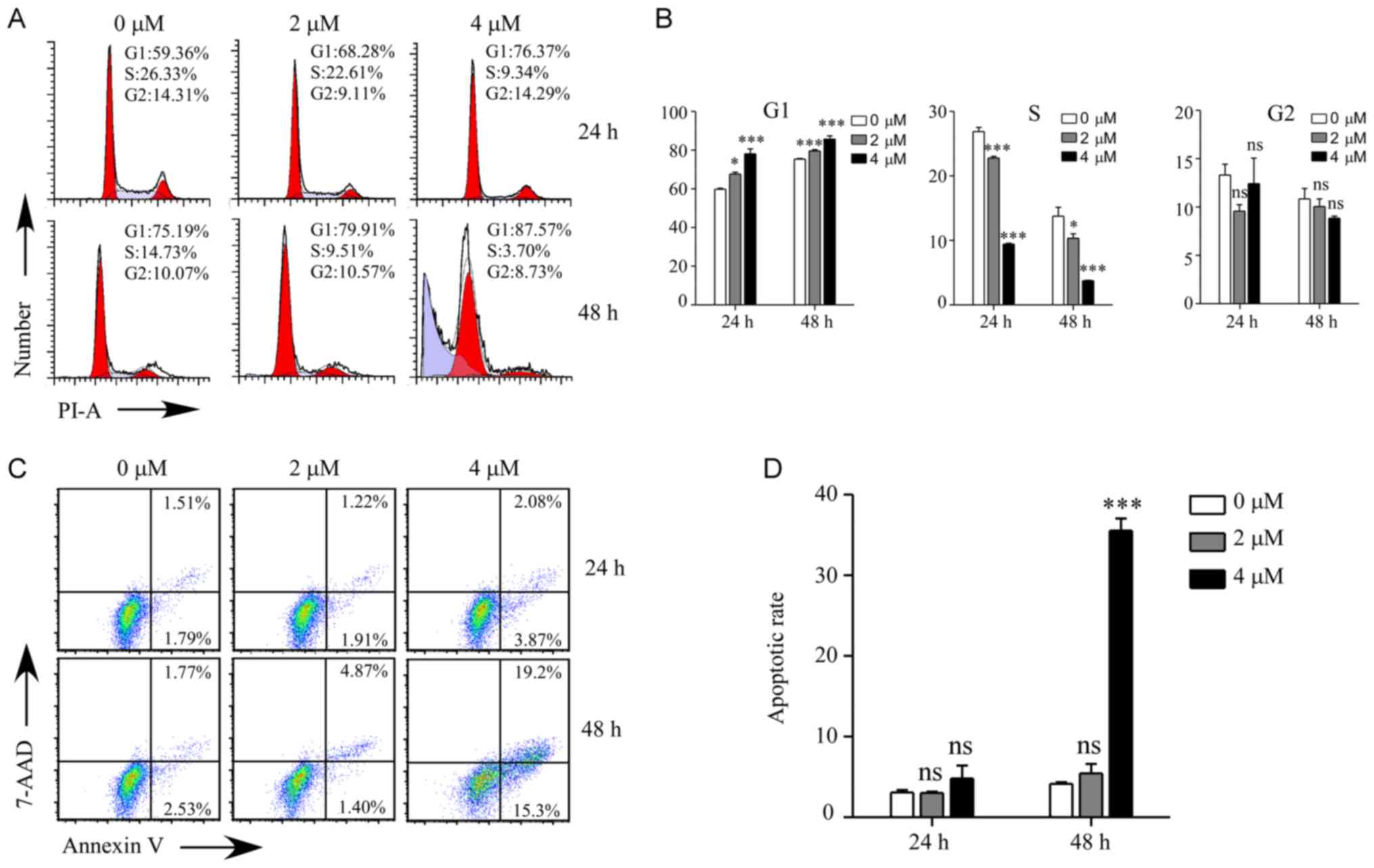
GNA induces cell cycle arrest and
apoptosis in A549/Cis cells
To investigate the cellular process responsible for
the inhibited proliferation by GNA treatment, the cell cycle and
apoptosis were examined by flow cytometry in A549/Cis cells
(Fig. 3). As presented in Fig. 3A and B, the cell cycle of A549/Cis
cells was significantly arrested at the G1 phase
following GNA treatment for 24 and 48 h compared with the untreated
group (P<0.5). There was a significantly higher
sub-G1 population in the cells treated with 4 µM GNA for
48 h compared with the untreated group (P<0.001). Cell cycle
arrest may induce cell death, which was measured using flow
cytometry. The annexin V/7-AAD double staining assay revealed that
the apoptosis rate was significantly increased compared with the
control group when A549/Cis cells were treated with 4 µM GNA for 48
h (P<0.001; Fig. 3C and D).
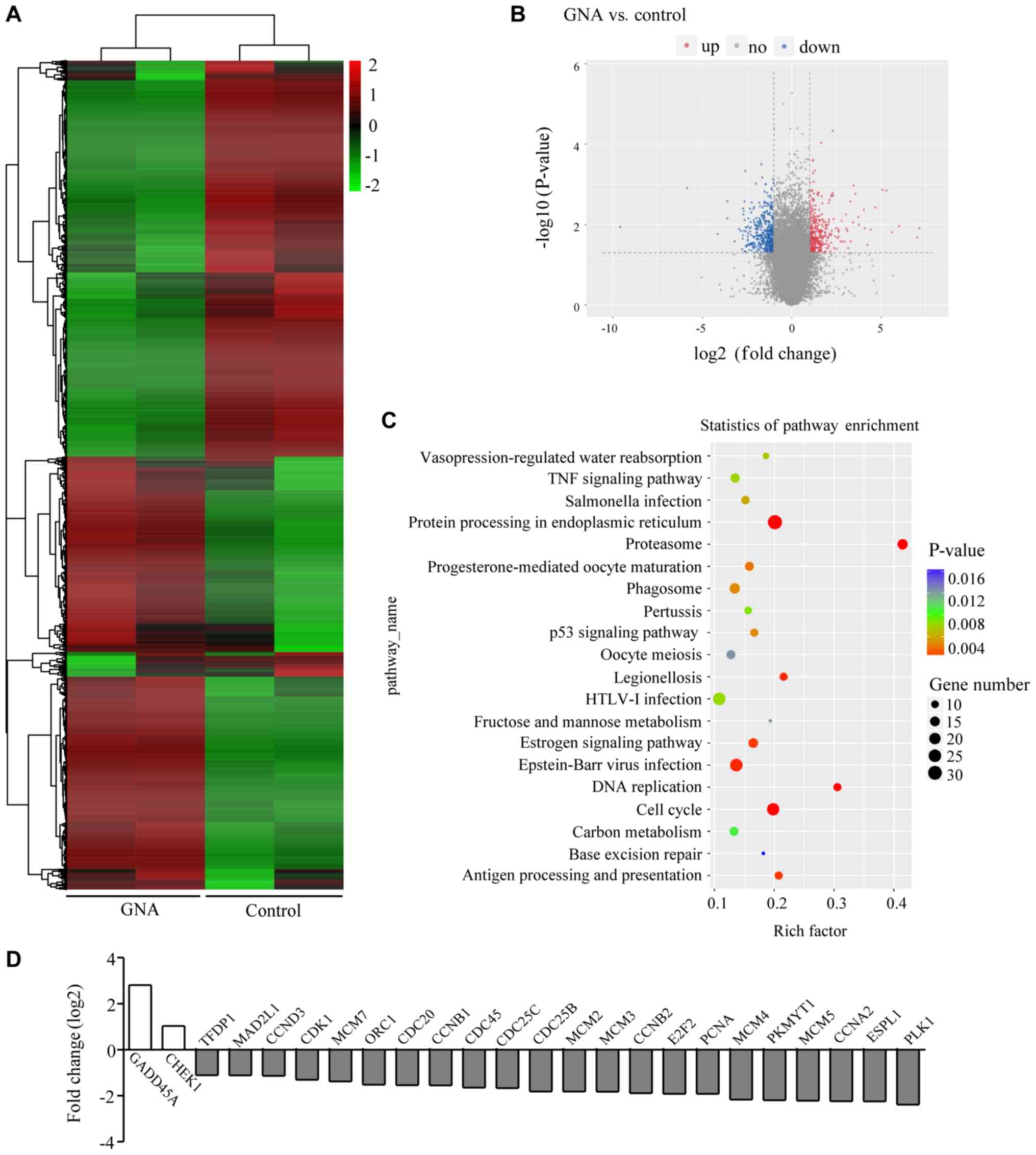
Differential gene expression and
enrichment analysis in A549/Cis cells treated with GNA
To understand how GNA inhibits cell growth and
promotes cell death in A549/Cis cells, an RNA-seq assay was
performed using samples from the control and GNA-treated cells.
Data analysis indicated that GNA treatment induced a global gene
expression change (Fig. 4A). All
genes whose threshold was restricted with a P-value <0.05, fold
change ≥2 or ≤0.5 were identified as differentially expressed genes
(DEGs). There were 353 upregulated DEGs and 425 downregulated DEGs
in the cells treated with GNA; the DEGs are visualized in the
volcano plot (Fig. 4B). To further
investigate the function of DEGs, KEGG pathway analysis was
performed. It was identified that the DEGs influenced by GNA
treatment were mostly enriched in pathways involved in the cell
cycle, DNA replication and p53 signaling pathways associated with
tumor proliferation (Fig. 4C). In
addition, the DEGs associated with the cell cycle were further
enriched (Fig. 4D).
Mechanisms of the anticancer effects
of GNA in A549/Cis cells
To validate the RNA-seq results, 14 DEGs were
selected for RT-qPCR analysis and GAPDH was used to normalize the
gene expression data. Among these genes, 4 were verified to be
upregulated in GNA-treated A549/Cis cells whereas the other 10 were
identified as downregulated. In total, 10 genes [GADD45A, cyclin D3
(CCND3), cyclin B1 (CCNB1), cell division cycle 20, cell division
cycle 25B, proliferating cell nuclear antigen, polo like kinase 1,
minichromosome maintenance complex component (MCM)2, MCM 3 and MCM
7] were involved in the cell cycle, 6 genes [GADD45A, CCND3, CCNB1,
SERPINE1, thrombospondin 1 and TNF receptor superfamily member 10b
(TNFRSF10B)] were associated with the p53 signaling pathway and two
genes (CASPASE7 and TNFRSF10B) were associated with apoptosis. The
results of RT-qPCR were consistent with the RNA-seq analysis
(P<0.05; Fig. 5A). To further
investigate the underlying mechanisms of GNA-induced cell cycle
arrest and apoptosis, a western blotting assay was performed. The
protein expression levels of the hub genes associated with the
G1 phase cell cycle checkpoint and apoptosis are
presented in Fig. 5B, D and F. The
data revealed that the protein levels of cyclin D1, cyclin D3, CDK4
and CDK6 were significantly downregulated (P<0.05), while the
expression of p21, GADD45A, p53 and nuclear p53 were significantly
upregulated in A549/Cis cells following GNA treatment compared with
the untreated group (P<0.05). Furthermore, the regulators of
apoptosis were additionally detected subsequent to GNA treatment.
GNA was able to significantly increase the protein levels of the
precursor forms of caspase 3/7 and their active forms as well
(P<0.05). The hallmark of apoptosis, PARP, which is associated
with DNA repair, was revealed to be significantly upregulated
(P<0.05), and its cleavage was significantly enhanced by GNA
treatment in A549/Cis cells (P<0.05).
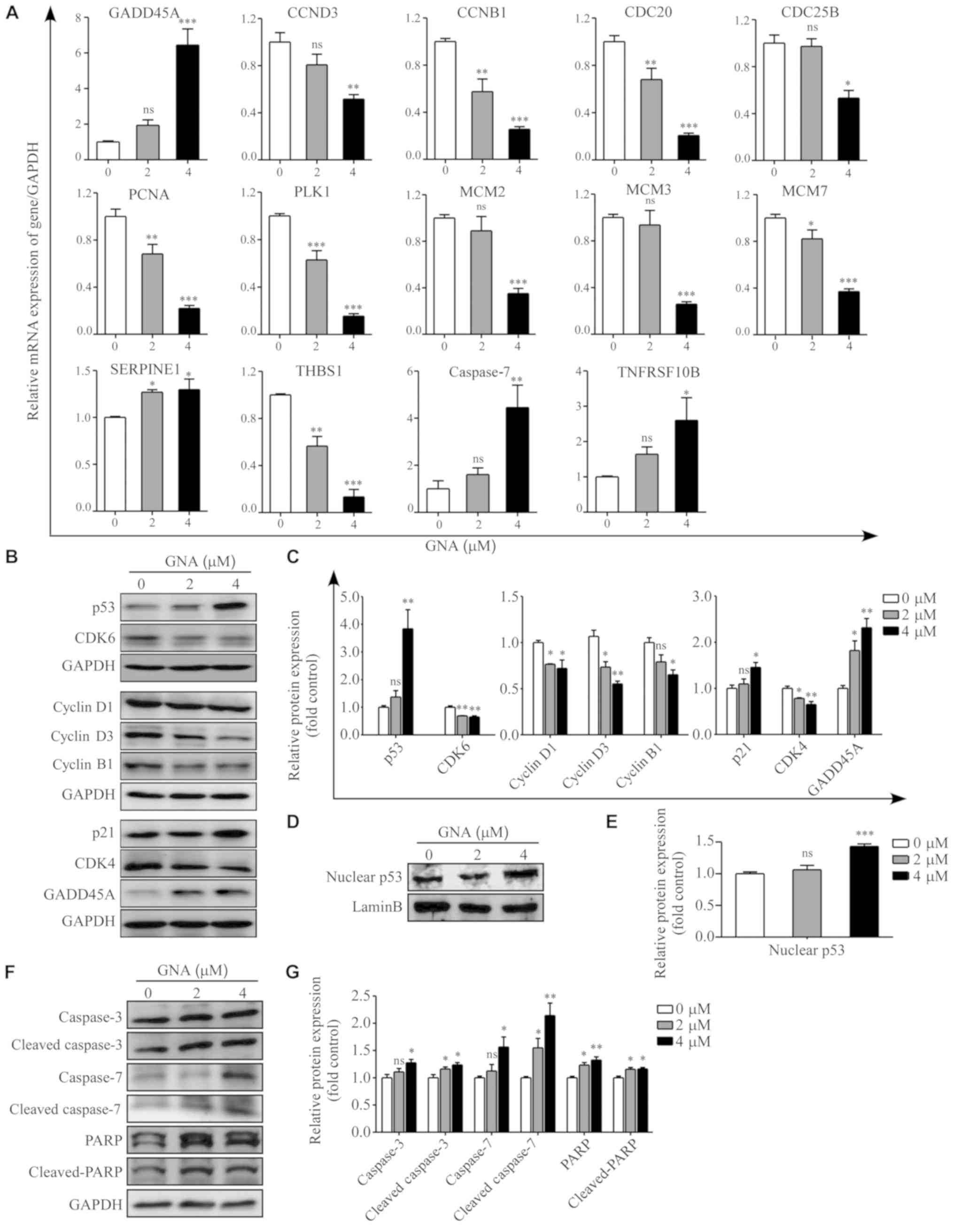 | Figure 5.Mechanisms underlying GNA-induced
cell cycle arrest and apoptosis. (A) Relative expression of
GADD45A, CCND3, CCNB1, CDC20, CDC25B, PCNA, PLK1, MCM2, MCM3, MCM7,
THBS1, SERPINE1, CASPASE7, TNFRSF10B and GAPDH were identified in
A549/Cis cells treated with different concentrations of GNA for 24
h by reverse transcription-quantitative PCR. (B) Western blot
analysis and (C) quantitative results of p53, GADD45A, p21, cyclin
D1, cyclin D3, CDK4, CDK6 and cyclin B1 in whole cell lysates of
A549/Cis cells treated with different doses of GNA for 24 h. The
levels of GAPDH were used as loading controls. (D) Western blot
analysis and (E) quantitative results of p53 in the nucleus of
A549/Cis cells treated with different doses of GNA for 24 h. The
levels of lamin B were used as loading controls. (F) Representative
blots and (G) quantitative results of the expression of caspase 3,
caspase 7 and PARP subsequent to GNA treatments. *P<0.05,
**P<0.01 and ***P<0.001 vs. untreated group. GNA, gambogenic
acid; Cis, cisplatin; ns, not significant; GADD45A, growth arrest
and DNA damage-inducible protein GADD45 α; CCND3, cyclin D3; CCNB1,
cyclin B1; CDC20, cell division cycle 20; CDC25B, cell division
cycle 25B; PCNA, proliferating cell nuclear antigen; PLK1, polo
like kinase 1; MCM, minichromosome maintenance complex component;
THBS1, thrombospondin 1; TNFRSF10B, TNF receptor superfamily member
10b; PARP, poly(ADP-ribose) polymerase. |
Discussion
The present study investigated the anticancer
activity of GNA in A549/Cis cells and investigated the underlying
molecular mechanisms. It was demonstrated that GNA exhibited potent
inhibitory activities in A549/Cis cells. Based on the present data,
GNA induced G0/G1 phase cell cycle arrest
mainly by regulating the expression of the cyclin D-CDK4/CDK6
complex, p21 and p53. The cell cycle arrest subsequently triggers
apoptosis through the activation of caspases in A549/Cis cells.
GNA, an active ingredient isolated from the
traditional Chinese medicine gamboge, was revealed to have
anti-tumor activity in vitro and in vivo, and low
toxicity to normal cells (11,13,14,30).
In the present study, GNA exhibited anti-cancer effects on
Cis-resistant NSCLC cells, indicating that it additionally has the
potential to overcome drug resistance in cancer. However, despite
these advantages, the clinical use of GNA was limited due to its
poor aqueous solubility, short elimination half-life, low
bio-availability and excessive irritation to blood vessels by
intravenous administration (31).
With the aim of overcoming these limitations, researchers have
developed numerous nanocarrier drug delivery systems, including
solid lipid nanoparticles, nanostructure lipid carriers (32), liquid crystal dispersions (33) and mixed polymeric micelles
(34), which are able to enhance
bio-availability and retain antitumor effects. These novel
formulations of GNA may be developed further for its clinical
application.
The cell cycle serves an important function in
maintaining the dynamic balance between proliferation and cell
death. Abnormal regulation of the cell cycle results in cancer
development. Inhibition of the cell cycle is a known target for
cancer therapy (35,36). Cell cycle progression is mainly
regulated by cyclins, cyclin-dependent kinases and cyclin-dependent
protein kinases inhibitors (CDKIs). The fluctuations in the cyclin
levels during the cell cycle may activate CDK (37). The D-type cyclins (D1, D2 and D3)
are the first cyclins sensing the mitogenic signals and activate
CDK4 and CDK6 in the G1 phase. At the G1/S
checkpoint, the cyclin D-CDK4/6 complex is crucial in regulating
the G1/S phase transition. Phosphorylation of Rb by
cyclin D-CDK4/6 and cyclin E-CDK2 releases E2F, allowing the
expression of genes that encode products necessary for S-phase
progression (38–40). The inhibition of the CDK4/6-cyclin
D complex will cause cell cycle arrest (41). The present results were consistent
with these observations. Previous studies also demonstrated that
GNA may induce cell cycle arrest at the G1 phase in
cancer cells (11,13). Consequently, the results of the
present study on the regulatory proteins responsible for
G1/S transition indicated that GNA arrested the cell
cycle at the G1 phase by downregulating cyclin D-CDK4/6
complex proteins. The CDKI family member p21 (also known as
p21WAF1/Cip1) serves an important function in binding to
the cyclin-CDK complex and suppressing its catalytic activity,
causing cell cycle arrest (42,43).
Inactivation of the cyclin D-CDK4/6 complex by p21 may inhibit Rb
phosphorylation and induce G1 phase cell cycle arrest
(44). The present study observed
that p21 was also altered by GNA. In addition, p21 is a direct
transcriptional target of p53 and is required for p53-dependent
cell cycle arrest (45). GADD45A
is a protein that may inactivate cyclin B1 through a
p53-dependent/independent pathway; upregulation of GADD45A may be
regulated by p53. These results suggested that the elevated protein
expression of p53 in response to GNA exposure may be involved in
G1 phase arrest by increasing the p21 protein level.
Cell cycle inhibition would subsequently induce
apoptosis in cancer therapy (46).
In the present study, it was identified that the percentage of
A549/Cis cells in the G1 phase increased following GNA
treatment for 24 and 48 h. However, the apoptotic rate of A549/Cis
cells had no statistical difference following 24 h GNA treatment
and significant differences only occurred 48 h later following 4 µM
GNA treatment. The present results demonstrated that apoptosis may
be a result of cell cycle arrest, which supports the cell cycle
inhibitory effect of GNA. Apoptosis is important as it serves a
pivotal function in growth inhibition, and the agents that induce
apoptosis in tumor cells may be useful for the treatment of
malignancies (39). Apoptosis is
primarily executed by a family of proteases known as the caspases
(47). Caspase 3/7 are the
executors of apoptosis, which are responsible for the definite
cleavage of cellular components (48). Activation of caspase 3/7 may
trigger PARP, which eventually results in apoptosis (49). In the present study, the activation
of caspase 3/7, in addition to cleaved PARP, were observed
following GNA treatment. In summary, it was hypothesized that GNA
induced cell cycle arrest, which results in apoptosis via the
activation of caspases in A549/Cis cells.
In conclusion, the present study demonstrated that
GNA is a natural and effective compound that exerts anticancer
effects against A549/Cis cells by inducing apoptosis via cell cycle
arrest at the G1 phase. GNA induced G1 phase
cell cycle arrest through the regulation of cyclin D1, cyclin D3,
CDK4, CDK6, p21 and p53, which subsequently resulted in apoptosis
via the activation of caspase 3/7 in A549/Cis cells. The present
evidence supported the potential application of GNA as a promising
drug in the treatment of Cis-resistant NSCLC.
Acknowledgements
Not applicable.
Funding
The present study was supported by The National
Natural Science Foundation of China (grant nos. 81473569 and
81973735).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CL and YW conceived and designed the study. DS and
HN performed the experiments. DS wrote the manuscript. YW and DS
analyzed the data. CL, YW, DS and HN reviewed and edited the
manuscript. All authors read and approved the manuscript and agreed
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work is appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bray F, Ferlay J, Soerjomataram I, Siegel
RL, Torre LA and Jemal A: Global cancer statistics 2018: GLOBOCAN
estimates of incidence and mortality worldwide for 36 cancers in
185 countries. CA Cancer J Clin. 68:394–424. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Miller KD, Siegel RL, Lin CC, Mariotto AB,
Kramer JL, Rowland JH, Stein KD, Alteri R and Jemal A: Cancer
treatment and survivorship statistics, 2016. CA Cancer J Clin.
66:271–289. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arriagada R, Bergman B, Dunant A, Le
Chevalier T, Pignon JP and Vansteenkiste J; International Adjuvant
Lung Cancer Trial Collaborative Group, : Cisplatin-based adjuvant
chemotherapy in patients with completely resected non-small-cell
lung cancer. N Engl J Med. 350:351–360. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Mitsudomi T, Morita S, Yatabe Y, Negoro S,
Okamoto I, Tsurutani J, Seto T, Satouchi M, Tada H, Hirashima T, et
al: Gefitinib versus cisplatin plus docetaxel in patients with
non-small-cell lung cancer harbouring mutations of the epidermal
growth factor receptor (WJTOG3405): An open label, randomised phase
3 trial. Lancet Oncol. 11:121–128. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Oliver TG, Mercer KL, Sayles LC, Burke JR,
Mendus D, Lovejoy KS, Cheng MH, Subramanian A, Mu D, Powers S, et
al: Chronic cisplatin treatment promotes enhanced damage repair and
tumor progression in a mouse model of lung cancer. Genes Dev.
24:837–852. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Stewart DJ: Mechanisms of resistance to
cisplatin and carboplatin. Crit Rev Oncol Hematol. 63:12–31. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Maroun JA, Anthony LB, Blais N, Burkes R,
Dowden SD, Dranitsaris G, Samson B, Shah A, Thirlwell MP, Vincent
MD and Wong R: Prevention and management of chemotherapy-induced
diarrhea in patients with colorectal cancer: A consensus statement
by the Canadian working group on chemotherapy-induced diarrhea.
Curr Oncol. 14:13–20. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
de Gramont A, Figer A, Seymour M, Homerin
M, Hmissi A, Cassidy J, Boni C, Cortes-Funes H, Cervantes A, Freyer
G, et al: Leucovorin and fluorouracil with or without oxaliplatin
as first-line treatment in advanced colorectal cancer. J Clin
Oncol. 18:2938–2947. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo F, Gu J, Chen L and Xu X: Systems
pharmacology strategies for anticancer drug discovery based on
natural products. Mol Biosyst. 10:1912–1917. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Asano J, Chiba K, Tada M and Yoshii T:
Cytotoxic xanthones from Garcinia hanburyi. Phytochemistry.
41:815–820. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yan F, Wang M, Chen H, Su J, Wang X, Wang
F, Xia L and Li Q: Gambogenic acid mediated apoptosis through the
mitochondrial oxidative stress and inactivation of Akt signaling
pathway in human nasopharyngeal carcinoma CNE-1 cells. Eur J
Pharmacol. 652:23–32. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen HB, Zhou LZ, Mei L, Shi XJ, Wang XS,
Li QL and Huang L: Gambogenic acid-induced time- and dose-dependent
growth inhibition and apoptosis involving Akt pathway inactivation
in U251 glioblastoma cells. J Nat Med. 66:62–69. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yu XJ, Han QB, Wen ZS, Ma L, Gao J and
Zhou GB: Gambogenic acid induces G1 arrest via GSK3β-dependent
cyclin D1 degradation and triggers autophagy in lung cancer cells.
Cancer Lett. 322:185–194. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yan F, Wang M, Li J, Cheng H, Su J, Wang
X, Wu H, Xia L, Li X, Chang HC and Li Q: Gambogenic acid induced
mitochondrial-dependent apoptosis and referred to phospho-Erk1/2
and phospho-p38 MAPK in human hepatoma HepG2 cells. Environ Toxicol
Pharmacol. 33:181–190. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu P, Wu X, Dai L, Ge Z, Gao C, Zhang H,
Wang F, Zhang X and Chen B: Gambogenic acid exerts antitumor
activity in hypoxic multiple myeloma cells by regulation of miR-21.
J Cancer. 8:3278–3286. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen F, Zhang XH, Hu XD, Zhang W, Lou ZC,
Xie LH, Liu PD and Zhang HQ: Enhancement of radiotherapy by ceria
nanoparticles modified with neogambogic acid in breast cancer
cells. Int J Nanomedicine. 10:4957–4969. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Su J, Cheng H, Zhang D, Wang M, Xie C, Hu
Y, Chang HC and Li Q: Synergistic effects of 5-fluorouracil and
gambogenic acid on A549 cells: Activation of cell death caused by
apoptotic and necroptotic mechanisms via the ROS-mitochondria
pathway. Biol Pharm Bull. 37:1259–1268. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
He Y, Ding J, Lin Y, Li J, Shi Y, Wang J,
Zhu Y, Wang K and Hu X: Gambogenic acid alters chemosensitivity of
breast cancer cells to Adriamycin. BMC Complement Altern Med.
15:1812015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang X, Cao W, Zhang J, Yan M, Xu Q, Wu X,
Wan L, Zhang Z, Zhang C, Qin X, et al: A covalently bound inhibitor
triggers EZH2 degradation through CHIP-mediated ubiquitination.
EMBO J. 36:1243–1260. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pertea M, Pertea GM, Antonescu CM, Chang
TC, Mendell JT and Salzberg SL: StringTie enables improved
reconstruction of a transcriptome from RNA-seq reads. Nat
Biotechnol. 33:290–295. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Frazee AC, Pertea G, Jaffe AE, Langmead B,
Salzberg SL and Leek JT: Ballgown bridges the gap between
transcriptome assembly and expression analysis. Nat Biotechnol.
33:243–246. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The gene
ontology consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
The Gene Ontology Consortium: The gene
ontology resource: 20 years and still GOing strong. Nucleic Acids
Res. 47(D1): D330–D338. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kanehisa M, Sato Y, Furumichi M, Morishima
K and Tanabe M: New approach for understanding genome variations in
KEGG. Nucleic Acids Res. 47(D1): D590–D595. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kanehisa M: Toward understanding the
origin and evolution of cellular organisms. Protein Sci.
28:1947–1951. 2019. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Huang da W, Sherman BT and Lempicki RA:
Bioinformatics enrichment tools: Paths toward the comprehensive
functional analysis of large gene lists. Nucleic Acids Res.
37:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Huang T, Zhang H, Wang X, Xu L, Jia J and
Zhu X: Gambogenic acid inhibits the proliferation of small-cell
lung cancer cells by arresting the cell cycle and inducing
apoptosis. Oncol Rep. 41:1700–1706. 2019.PubMed/NCBI
|
|
31
|
Huang X, Chen YJ, Peng DY, Li QL, Wang XS,
Wang DL and Chen WD: Solid lipid nanoparticles as delivery systems
for gambogenic acid. Colloids Surf B Biointerfaces. 102:391–397.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lin T, Huang X, Wang Y, Zhu T, Luo Q, Wang
X, Zhou K, Cheng H, Peng D and Chen W: Long circulation
nanostructured lipid carriers for gambogenic acid: Formulation
design, characterization, and pharmacokinetic. Xenobiotica.
47:793–799. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Luo Q, Lin T, Zhang CY, Zhu T, Wang L, Ji
Z, Jia B, Ge T, Peng D and Chen W: A novel glyceryl
monoolein-bearing cubosomes for gambogenic acid: Preparation,
cytotoxicity and intracellular uptake. Int J Pharm. 493:30–39.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lin TY, Zhu TT, Xun Y, Tao YS, Yang YQ,
Xie JL, Zhang XM, Chen SX, Ding BJ and Chen WD: A novel drug
delivery system of mixed micelles based on poly(ethylene
glycol)-poly(lactide) and poly(ethylene
glycol)-poly(varepsilon-caprolactone) for gambogenic acid.
Kaohsiung J Med Sci. 35:757–764. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Sherr CJ and Roberts JM: CDK inhibitors:
Positive and negative regulators of G1-phase progression. Genes
Dev. 13:1501–1512. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goyeneche AA, Caron RW and Telleria CM:
Mifepristone inhibits ovarian cancer cell growth in vitro and in
vivo. Clin Cancer Res. 13:3370–3379. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Xu H, Wang Z, Jin S, Hao H, Zheng L, Zhou
B, Zhang W, Lv H and Yuan Y: Dux4 induces cell cycle arrest at G1
phase through upregulation of p21 expression. Biochem Biophys Res
Commun. 446:235–240. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bertoli C, Skotheim JM and de Bruin RA:
Control of cell cycle transcription during G1 and S phases. Nat Rev
Mol Cell Biol. 14:518–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Giacinti C and Giordano A: RB and cell
cycle progression. Oncogene. 25:5220–5227. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Musgrove EA, Caldon CE, Barraclough J,
Stone A and Sutherland RL: Cyclin D as a therapeutic target in
cancer. Nat Rev Cancer. 11:558–572. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rader J, Russell MR, Hart LS, Nakazawa MS,
Belcastro LT, Martinez D, Li Y, Carpenter EL, Attiyeh EF, Diskin
SJ, et al: Dual CDK4/CDK6 inhibition induces cell-cycle arrest and
senescence in neuroblastoma. Clin Cancer Res. 19:6173–6182. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Wu S, Cetinkaya C, Munoz-Alonso MJ, von
der Lehr N, Bahram F, Beuger V, Eilers M, Leon J and Larsson LG:
Myc represses differentiation-induced p21CIP1 expression via
Miz-1-dependent interaction with the p21 core promoter. Oncogene.
22:351–360. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Lee EW, Lee MS, Camus S, Ghim J, Yang MR,
Oh W, Ha NC, Lane DP and Song J: Differential regulation of p53 and
p21 by MKRN1 E3 ligase controls cell cycle arrest and apoptosis.
EMBO J. 28:2100–2113. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Tibbetts RS, Brumbaugh KM, Williams JM,
Sarkaria JN, Cliby WA, Shieh SY, Taya Y, Prives C and Abraham RT: A
role for ATR in the DNA damage-induced phosphorylation of p53.
Genes Dev. 13:152–157. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Liu G and Lozano G: p21 stability: Linking
chaperones to a cell cycle checkpoint. Cancer Cell. 7:113–114.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Evan GI and Vousden KH: Proliferation,
cell cycle and apoptosis in cancer. Nature. 411:342–348. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Li J and Yuan J: Caspases in apoptosis and
beyond. Oncogene. 27:6194–6206. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Thornberry NA and Lazebnik Y: Caspases:
Enemies within. Science. 281:1312–1316. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Saraste A and Pulkki K: Morphologic and
biochemical hallmarks of apoptosis. Cardiovasc Res. 45:528–537.
2000. View Article : Google Scholar : PubMed/NCBI
|