Introduction
The incidence of metabolic disorders and obesity has
increased globally, and this has become a major health problem.
Although <5% of the global population live in America, 13% of
the obese population globally reside there (1). Two-thirds of the obese population
reside in developing countries, including China and India (2,3).
Obesity and nonalcoholic fatty liver disease (NAFLD) are not
superficial diseases; they have become major risk factors for
morbidity and mortality (4). For
example, they increase the risk of type 2 diabetes and high blood
pressure, and cause dyslipidemia, cardiovascular disease,
hyperuricemia and obstructive sleep apnea-hypopnea syndrome
(5).
Leptin, a type of protein hormone secreted by
adipose tissue and skeletal muscle, may participate in the
regulation of sugar, adipose and energy metabolism (6). Free leptin, its active form, binds to
specific transport proteins, which are transmitted via signal
transduction and STAT3 pathways (7,8). The
phosphorylation of STAT3 results in a reduction in ingestion,
increased energy release and inhibition of the synthesis of adipose
cells, which results in an overall reduction in weight (9). Leptin is then cleared by the kidneys
(10).
In the endoplasmic reticulum stress (ERS) of
eukaryotic cells, unfolded or misfolded proteins accumulate and
trigger the unfolded protein response (UPR) (11). The UPR is an indicator of ERS and
is mediated by three important ERS receptor proteins: Double
stranded RNA-dependent protein kinase-like endoplasmic reticulum
kinases (PERKs), inositol requiring enzyme l (IRE) and activating
transcription factor 6c (ATF6) (12). In the resting state, the three
types of transmembrane protein and glucose regulation protein 78
(GRP78) binding immunoglobulin protein are in the inactive state
(13). Numerous studies have
demonstrated that chronic overeating may result in tissue ERS,
insulin resistance and inflammation in adipose tissue in ob/ob mice
and high-fat diet (HFD)-fed mice (14,15).
NAFLD refers to the clinicopathologic syndrome
characterized by excessive fat deposition in hepatocytes caused by
alcohol exclusion and other definite hepatogenic factors (16). A large quantity of free fatty acids
(FFAs) accumulate in hepatocytes, inducing lipid peroxidation in
hepatocytes and eventually the apoptosis and necrosis of cells. In
this process, strong lipid peroxidation is the inducing factor of
ERS (17). ERS is conducive to the
homeostasis of hepatocytes (18).
However, due to the excessive accumulation of lipids and persistent
lipid peroxidation reaction, the ERS response lasts too long and is
too strong, which causes cellular damage (19). A previous study revealed that mice
without ATF6-α, eukaryotic initiation factor 2α (eIF2-α) and IRE1-α
in the ERS pathway, exhibited ERS disorder and hepatic steatosis,
suggesting that ERS may be involved in the cause of fatty liver
disease (20).
Ginsenosides (GS) [Chemical Abstracts Service (CAS)
no. 90045-38-8] are an extract from the roots, stems and leaves of
ginseng (21). It is a
yellow-white or light yellow amorphous powder that has a slight
smelly odor and is bitter and hygroscopic (22). GS have various pharmacological
effects on the human body (23).
In the cardiovascular system, GS reduce arrhythmia, cardiac
hypertrophy, myocardial ischemia and myocardial cell apoptosis
(24). In terms of a tumor, GS may
induce apoptosis, inhibit tumor cell proliferation and regulate
signal pathways and immune functions (25). With regards to metabolic
regulation, GS inhibit feeding behavior through the ventromedial
hypothalamic nucleus, and inhibit the absorption of fatty foods in
the intestinal tract by reducing the activity of pancreatic lipase
(PL) or through regulation of the lipogenic transcription
factor-peroxidase proliferator activation receptor (PPAR) (26,27).
Glucolipid metabolism regulates the expression of adenosine
activated protein kinase (AMPK) and its target genes (28). However, the mechanism by which GS
reduces dietary intake is unknown. In a previous study, GS were
revealed to exert a significant effect on reducing the food intake
and weight of HFD-induced obese mice, significantly reducing serum
triglycerides (TGs; P=0.03), increasing glucose tolerance,
improving insulin resistance and reducing the degree of obesity in
mice, compared with HFD mice (29). A further study demonstrated that GS
significantly inhibited the transcriptional activity of C/EBP
homologous protein (CHOP; P=0.028) and GRP7 (P=0.04) compared with
HFD mice, indicating that GS reduce ERS (30). Therefore, the present study
hypothesized that GS may alleviate ERS and thus increase the
sensitivity of the leptin receptor in order to inhibit obesity.
Materials and methods
Compounds
GS (CAS no. 90045-38-8; cat. no. PS010497) were
purchased from Chengdu Push Bio-Technology Co., Ltd. According to
the product specification, the extract is mainly composed of
saponins (total content, >80%). The GS were dissolved in
dimethylsulfoxide (DMSO; Sigma-Aldrich; Merck KGaA) for in
vitro studies. Tunicamycin (TM; CAS no. 11089-65-9; cat. no.
T7765) was obtained from Sigma-Aldrich (Merck KGaA) in order to
induce ERS.
Animals and diet
The present study was conducted in accordance with
the ethical standards and according to the Ethical Committee of
Shanghai University of Traditional Chinese Medicine (Shanghai,
China). The protocols were ethnically approved by the Institutional
Animal Care and Use Committee of Shanghai University of Traditional
Chinese Medicine (approval no. SZY201708002).
Male C57BL/6 mice (n=15; weight, 15–20 g) were
purchased from Shanghai Laboratory Animal Center, certificate no.
20080016722050; Shanghai, China) at 4 weeks of age. The mice were
individually housed under a 12 h light-dark cycle at 22–23°C, with
access to a standard chow diet and distilled water ad
libitum during the adaptation week. Subsequently, the mice were
placed on a HFD (60% of calories derived from fat, 5.24 Kcal/gm;
cat. no. D12492; Research Diets, Inc.) for 3 months to induce
obesity (31). The HFD-fed mice
were distributed into two groups of five mice and housed in cages
to permit control of their food intake and body weight. All mice in
these two groups continued to receive a HFD. A separate group of
mice (n=5) were fed a standard chow diet (10% of calories derived
from fat; cat. no. D12450B; Research Diets, Inc.) as a control
group.
The C57BL/6 were fed either a standard chow diet
(CHOW group; n=5) or a HFD (HFD group; n=5) for 3 months. The
HFD-fed mice were treated with either GS at 120 mg/kg/day (HFD+GS
group; n=5) or with the vehicle (HFD group; n=5) for the final 28
days of the study period.
Cell culture
HepG2 and GT1-7 cell lines (American Type Culture
Collection) were cultured in DMEM (Biological Industries)
supplemented with 10% FBS (Thermo Fisher Scientific, Inc.) and 1%
penicillin/streptomycin solution (Thermo Fisher Scientific, Inc.).
The GT1-7 cells were incubated in a fully humidified 5%
CO2 incubator at 37°C. The medium was changed every 2–3
days, and the cells were routinely passaged every 6–8 days.
Briefly, for the ERS group, the cells were seeded at a density of
2×105 cells per well in 6-well plates. The first well
contained no treatment (control). From the second to the sixth
wells, 5 µg/ml TM, 5 µg/ml TM + 25 µg GS, 5 µg/ml TM + 50 µg GS, 5
µg/ml TM + 100 µg GS and 5 µg/ml TM + 200 µg GS were added,
respectively, subsequent to incubation at 37°C for 24 h to analyze
the presence of ERS through the expression levels of GRP78, CHOP,
phosphorylated (p)-PERK, PERK and activating transcription factor 4
(ATF4). For the phosphorylated p-STAT3/STAT3 group, the cells were
seeded at a density of 2×105 cells per well in 6-well
plates. The first well contained no treatment (control). From the
second to the sixth wells, GS (12.5, 25, 50, 100 and 200 µg) were
added, respectively, and the cells were incubated at 37°C for 24 h
to extract the protein to measure the p-STAT3/STAT3 ratio (32).
Preparation of FFAs
A hepatocyte steatosis model was established using
complete medium containing oleic acid (OA; Sigma-Aldrich; Merck
KGaA) and palmitic acid (PA; Sigma-Aldrich; Merck KGaA) and the
efficacy was evaluated using oil red O staining. DMEM medium with
10% FBS containing 200 M OA and 100 M PA was used. Initially, the
palmitic acid was saponified with sodium hydroxide solution and
turned into sodium palmitate to prepare the solution. At a normal
temperature water-soluble OA and PA sodium was low, so they were
dissolved in a 75°C water bath for 10 min. Then, 20 and 40%
degreased bovine serum albumin (BSA; Sigma-Aldrich; Merck KGaA)
solutions were prepared, and the BSA was completely dissolved using
centrifugation at 5,255 × g for 15 min at room temperature.
Completely dissolved OA or PA sodium solution, respectively, was
mixed with 20% BSA or 40% BSA solution, and then run through a 0.22
µm filter subsequent to eliminating bacteria, and stored at 4°C in
a refrigerator.
Western blotting
Following the treatments listed above,
2×105 cells/well were lysed using RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing a protease
inhibitor cocktail (Beyotime Institute of Biotechnology). A BCA
assay kit was used for protein concentration determination
(Beyotime Institute of Biotechnology) and either 20 or 80 µg
protein, to measure ERS or the p-STAT/STAT3 ratio, respectively,
was mixed with 2X SDS-PAGE sample buffer (25% SDS, 62.5 mM
Tris-HCl, pH 6.8, 25% glycerol, 0.5 M DTT and 0.1% Bromophenol
Blue) and then boiled for 7 min at 99°C. Proteins were separated by
12% SDS-PAGE and then transferred to a PVDF membrane (EMD
Millipore). The membranes were blocked with 5% BSA (Gibco; Thermo
Fisher Scientific, Inc.) for 2 h at room temperature. Subsequently,
the membranes were incubated with primary antibodies against GRP78,
CHOP, p-PERK, PERK, ATF4 and β-actin for 4°C for 12 h to assess
ERS, whereas primary antibodies against p-STAT3, STAT3 and β-actin
were incubated at 4°C for 24 h to obtain the p-STAT3/STAT3 ratio.
Following the primary antibody incubation, membranes were incubated
with an IR-Dye™−700/800-labeled fluorescent secondary antibody
(LI-COR Biosciences) for 1 h at room temperature in the dark.
Finally, proteins were visualized using the LI-COR Odyssey infrared
imaging system (LI-COR Biosciences) and expression levels were
quantified using Image-Pro Plus version 6.0 software (Media
Cybernetics, Inc.).
For the tissue-based assays, at the end of the
treatment, overnight fasting blood samples were withdrawn by
cardiac puncture following anesthesia (urethane, 2 g/kg,
intraperitoneal). Whether the mice were in a coma was determined
using the following three criteria: i) The depth of anesthesia in
mice was determined by pinching the toes with forceps; ii) the
normal respiratory rate of the mice should be 180 times per min,
and when anesthetized, the respiratory rate should decrease
proportional to the depth of the anesthesia, but the mice should
remain breathing; and iii) the mucous membranes of the mice must
remain pink, not blue or gray. Subsequent to extracting the tissue
and blood, the mice were placed into a sealed box and euthanized
(filled with CO2, 30% per min, 1–3 min). When all the
mice had succumbed to mortality (determined by observing the
respiratory rate of the mice and the color change of the mucous
membrane), they were registered (species, sex, number, etc.) and
handed over to a professional for disposal. The liver and
hypothalamus tissues of the mice in each group were homogenized
using RIPA buffer containing protease inhibitor cocktail and
phosphatase inhibitor cocktails (Beyotime Institute of
Biotechnology) using a hand-held motor and maintained on ice for 1
h to lyse the cells completely. The homogenates were then
centrifuged at 12,000 × g for 30 min at 4°C. The supernatants were
collected, and protein concentration was determined using BCA
protein assay kit, and then boiled for 10 min at 99°C. The
remaining steps are consistent with the cell western blotting
protocol.
Antibodies against p-STAT3 (1:500; cat. no. CST9134)
and STAT3 (1:1,000; cat. no. CST9139) were obtained from Cell
Signaling Technology, Inc. Antibodies against CHOP (1:1,000; cat.
no. 15204-1-AP), ATF4 (1:1,000; cat. no. 24169-1-AP), PERK
(1:1,000; cat. no. 20582-1-AP) and β-actin (1:1,000; cat. no.
66009-1-lg) were purchased from ProteinTech Group, Inc. Antibodies
against GRP78 (1:500; cat. no. 35394) and p-PERK (1:500; cat. no.
12379) were purchased from Signalway Antibody LLC. The
IR-Dye™−700/800-labeled fluorescent secondary antibody (1:10,000;
cat. no. 042-07-18-06) was purchased from LI-COR Biosciences.
Administration of GS
Intraperitoneal injections were performed in the
daytime. The mice were acclimatized for 28 days by daily
intraperitoneal injections of vehicle (DMSO+ normal saline; NS)
prior to the start of GS injections. GS (120 mg/kg/mice) were
dissolved in DMSO (10 µl) + NS (40 µl) and administered to the mice
intraperitoneally once a day. The corresponding vehicle groups were
injected intraperitoneally with a total volume of 50 µl DMSO+NS
once a day. The daily food intake was calculated as the mean food
intake per day during the four weeks of treatment.
Intraperitoneal glucose and insulin
tolerance tests
For the glucose tolerance test, the C57BL/6 mice
were fasted for 12 h and the basal blood glucose levels (0 min)
were measured from the tail vein. Subsequently the mice were
intraperitoneally injected with glucose (1 g/kg body weight) and
the blood glucose levels were measured at 15, 30, 60 and 90 min.
For the insulin tolerance test, the mice were intraperitoneally
injected with 0.75 U/kg insulin (cat. no. I-9278; Sigma-Aldrich;
Merck KGaA) without fasting. Prior to the injection of insulin, the
blood samples were collected from the tail vein for measurement of
basal blood glucose levels (0 min), and the blood glucose levels
were also measured at 15, 30, 60 and 90 min, as previously
described (33).
Metabolic chamber measurements
Metabolic parameters of the mouse models (lean and
diet-induced obesity) were determined using a Comprehensive Lab
Animal Monitoring System (Columbus Instruments International). The
following four parameters were tested: Oxygen consumption
(VO2), carbon dioxide consumption (VCO2),
respiratory exchange ratio (RER) and ambulatory activity (X-Amb)
(34).
Magnetic resonance imaging (MRI)
The mice were examined in an MRI scanner
(EchoMRI™-500; EchoMRI LLC) in a prostrate position, using a
whole-body coil and a two-point Dixon sequence. The mice were fixed
in plastic cylindrical barrels to reduce their movement. The
remaining operations were conducted according to the manufacturer's
protocol.
Hematoxylin and eosin (H&E) and
oil red O staining
For H&E staining, the liver tissue was fixed
with 10% formaldehyde for 24 h at 4°C, embedded in paraffin and cut
into 10 mm sections according to a standard protocol. The sections
were stained with H&E (hematoxylin, 10 min; eosin, 30 sec; room
temperature). The adipose tissue or frozen sections of liver were
washed with phosphate buffered saline twice, fixed with 10%
formalin at room temperature for 10 min and then stained with oil
red O (Sigma-Aldrich; Merck KGaA) at 60°C for 10 min. Subsequently,
images were captured using a light microscope (magnification, ×200
or ×400; Nikon Corporation). Oil red O stained sections were
evaluated using Image-ProPlus 6.0 software (Media Cybernetics,
Inc.) and H&E staining sections were evaluated according to The
Pathology Committee of the NASH Clinical Research Network's
histological feature scoring system (35).
Serum biochemistry analysis
Subsequent to overnight fasting for 12 h, all mice
were anesthetized with urethane (2 g/kg, intraperitoneally) and
then blood samples were collected for analysis. The blood samples
were drawn from the apex cordis into an epoxy resin tube, and serum
samples were separated from the blood. Serum TG, total cholesterol
(TC), high-density lipoprotein cholesterol (HDL-c), low-density
lipoprotein cholesterol (LDL-c) levels, alanine aminotransferase
(ALT) and aspartate aminotransferase (AST) were measured using a
Hitachi 7020 Automatic Analyzer (Hitachi, Ltd.).
Liver lipid content analysis
The liver, abdominal adipose, perirenal adipose and
epididymis adipose were collected rapidly at the end of treatment,
frozen in liquid nitrogen and stored at −80°C for the following
experiments. To assess the liver lipid content, 50 mg frozen liver
tissues were homogenized in 1 ml lysis buffer (20 mM Tris-HCl, pH
7.5, 150 mM NaCl and 1% Triton) and mixed with an equal volume of
chloroform. The chloroform layer was separated, dried and
resuspended in isopropyl alcohol to measure the lipid levels using
a Triglycerid Reagent kit (cat. no. A0-10017) and a Total
Cholesterol Reagent kit (cat. no. A010027; both Nanjing JianCheng
Bioengineering Institute) according to the manufacturer's
protocol.
Feces lipid content analysis
The feces were collected rapidly within three days
prior to the end of the treatment, frozen in liquid nitrogen, and
stored at −80°C for the following experiments. To assess the feces
lipid content, 100 mg frozen feces were homogenized in 1 ml lysis
buffer (20 mM Tris-HCl, pH 7.5, 150 mM NaCl and 1% Triton) and
mixed with an equal volume of chloroform. The chloroform layer was
separated, dried and resuspended in isopropyl alcohol to measure
the lipid levels using a using a Triglycerid Reagent kit (cat. no.
A0-10017) and a Total Cholesterol Reagent kit (cat. no. A010027;
both Nanjing JianCheng Bioengineering Institute) according to the
manufacturer's protocol.
ELISA
The serum insulin and leptin levels were measured
using an ELISA. The mouse serum was stored at −80°C and incubated
at 37°C for thawing prior to analysis. The kits for insulin (cat.
no. 80-INSMSU-E01) and leptin (cat. no. 80-LEPMS-E01) were obtained
from ALPCO. The enzyme levels in the supernatants were measured
according the manufacturer's protocol.
Statistical analysis
All values are expressed as the mean ± SEM. Data
were analyzed using SPSS software (version 21.0; IBM Corp., Armonk,
NY, USA). One-way ANOVA and Duncan's multiple range tests were used
to determine the significance of differences between groups.
P<0.05 was considered to indicate a statistically significant
difference.
Results
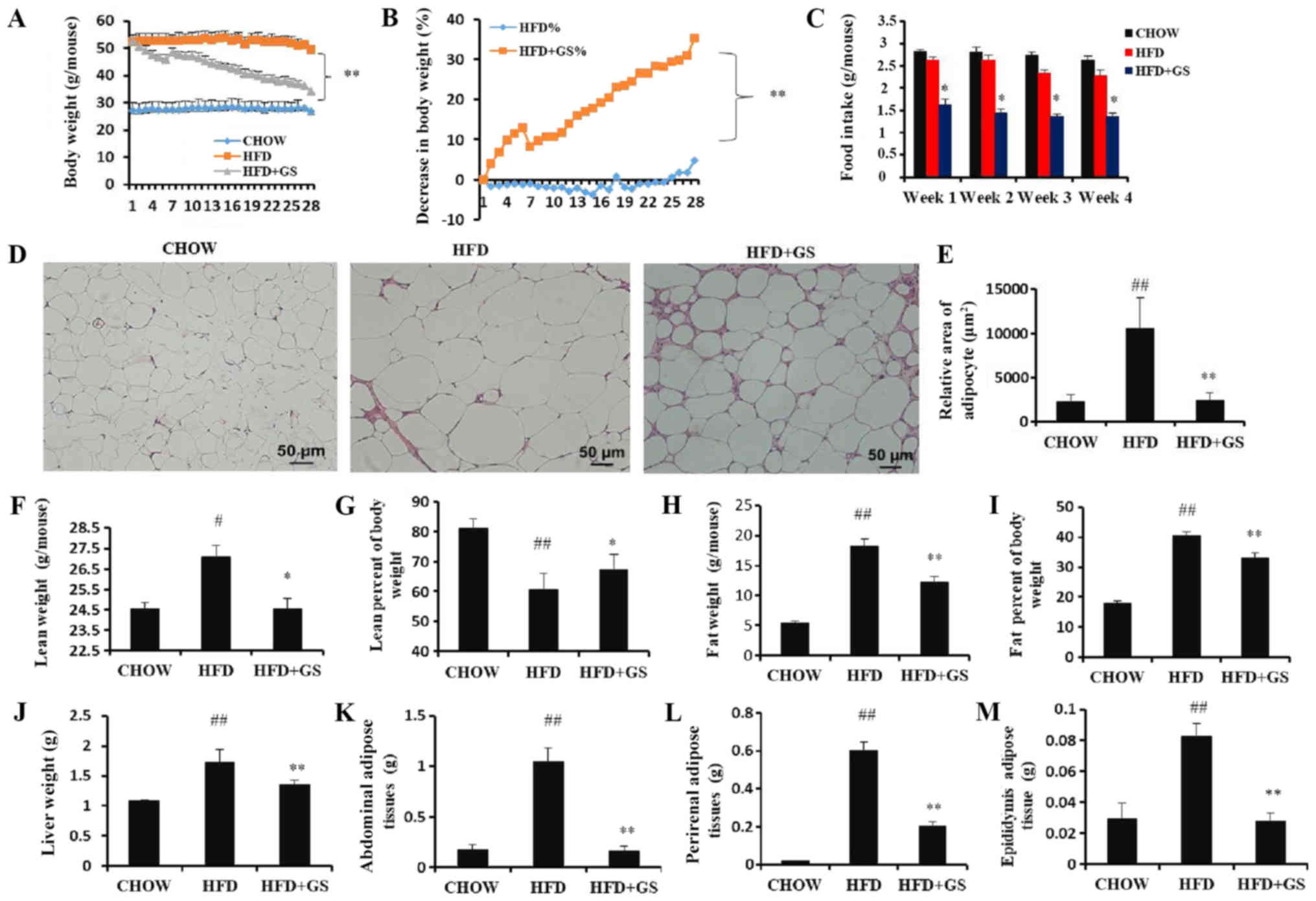
GS administration reduces food intake
and reverses the obesity and hyperleptinemia induced by HFD
To assess whether GS reduce diet-induced obesity,
C5BL/6 mice were fed a HFD for 4 weeks and administered GS via an
intraperitoneal injection every day (120 mg/kg/day). The mice fed
the HFD had an above mean body weight compared with the standard
chow diet-fed mice subsequent to 4 weeks of treatment (Fig. 1A). The HFD+GS group, however, had a
significantly reduced body weight in the mice compared with the HFD
group (Fig. 1A and B; P<0.01).
Similarly, the food intake amount was significantly different
between the HFD and HFD+GS group (P<0.05; Fig. 1C), implying that the lower body
weight in the HFD+GS group resulted from a lower calorie intake.
The diameter of fat cells may indirectly reflect lipid metabolism
(36). Histological analysis
demonstrated that the GS group significantly reduced the size of
the white adipose tissue compared with the HFD group (P<0.01),
implying that GS reduces adipocyte mass in HFD-fed mice (Fig. 1D and E). GS administration altered
the lean mass of HFD-fed mice relative to the standard chow group
(Fig. 1F and G; P<0.05). Also,
the percentage fat of the bodyweight in the HFD+GS group were
significantly less compared with that of standard chow diet-fed
groups (Fig. 1H and I; P<0.01).
When the experiment was completed, the weight of visceral fat in
the mice was measured. The data revealed that the liver weight,
abdominal fat, epididymis fat and perirenal fat were significantly
reduced in the HFD+GS group compared with the HFD group (Fig. 1J-M; P<0.01). These results
suggested that GS may induce a suppressed appetite and lose the
weight caused by a HFD in mice.
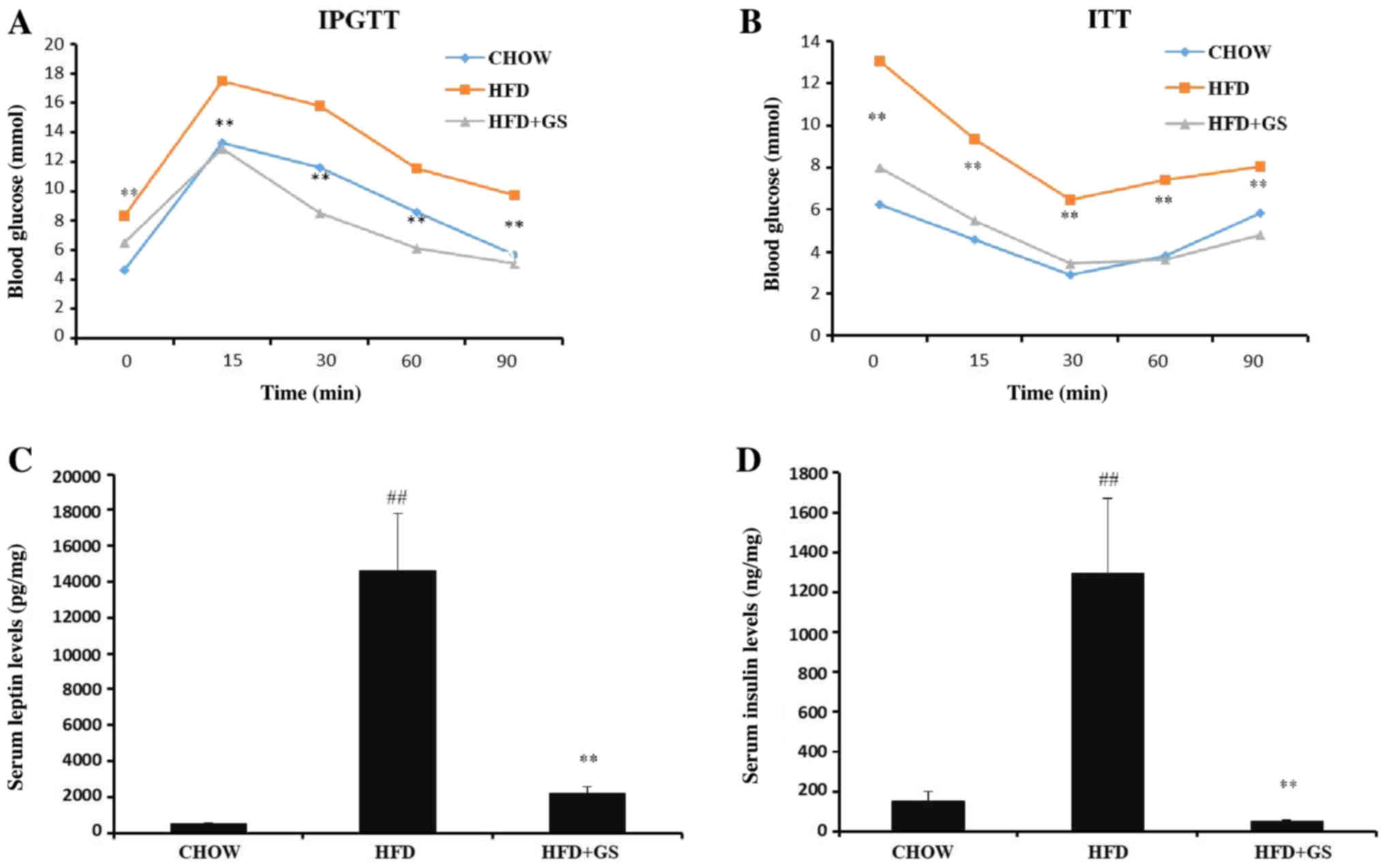
GS increase glucose tolerance, reduce
insulin resistance and reduce blood leptin and insulin levels in
HFD mice
A large number of studies have reported that HFD-fed
mice demonstrate impaired glucose and insulin intolerance compared
with standard chow-fed mice (37–40).
In the present study, the HFD+GS group had a significantly improved
fasting glucose level, glucose tolerance and insulin tolerance
compared with the HFD group. (Fig. 2A
and B; P<0.01). Similarly, leptin, an endocrine hormone
secreted by adipose tissue, is present at significantly higher
levels in HFD-fed mice compared with those in standard chow
diet-fed mice (41). In order to
examine whether the leptin levels were increased in HFD-fed mice in
the present study, leptin contents were assessed using an ELISA. GS
significantly reduced the leptin levels of the HFD-fed mice
compared with the HFD group. (Fig.
2C; P<0.01). Obesity is able to increase blood sugar, reduce
glucose tolerance, raise insulin resistance and cause
hyperinsulinemia and hyperleptinemia (42–44).
In order to examine whether the insulin levels were increased in
the HFD-fed mice, the insulin parameters were assessed using an
ELISA. The results demonstrated that the insulin levels were
increased significantly in the HFD-fed mice compared with the CHOW
group (P<0.01). Furthermore, GS treatment significantly
suppressed the insulin level, indicating that GS improved the
hyperinsulinemia in the HFD-induce mice (Fig. 2D; P<0.01).
GS improved the lipid metabolism of
HFD mice
In order to determine the toxic effect of GS, the
levels of ALT and AST in the blood were tested. The levels in the
obese mice induced by a HFD-diet were significantly increased
compared with standard chow-fed mice, while the AST and ALT levels
were significantly decreased following GS treatment (Fig. 3A and B; P<0.01). The results
revealed that GS exerted no toxicity to the liver at this dose. As
expected for the HFD-fed mice, the serum TC, TG, LDL-c and HDL-c
levels were significantly increased compared with the CHOW group
(Fig. 3C-F; P<0.05 or
P<0.01). By contrast, the GS group exhibited significantly
reduced serum TG levels compared with the HFD group (Fig. 3C; P<0.01). GS treatment also
significantly reduced serum TC and HDL-c levels compared with the
HFD group (Fig. 3D and E;
P<0.05). However, no difference in the serum LDL-c levels were
observed between the HFD + GS group and the HFD group (Fig. 3F). Subsequently, the TG levels in
the liver and TC in the feces were tested. The levels of TG in the
liver were significantly reduced in the HFD + GS group compared
with the HFD group, whereas TC levels in the feces were
significantly increased compared with the HFD group (Fig. 3G and H; P<0.05). In summary, the
results suggest that GS are not only non-toxic but may also
alleviate lipid metabolic disorders.
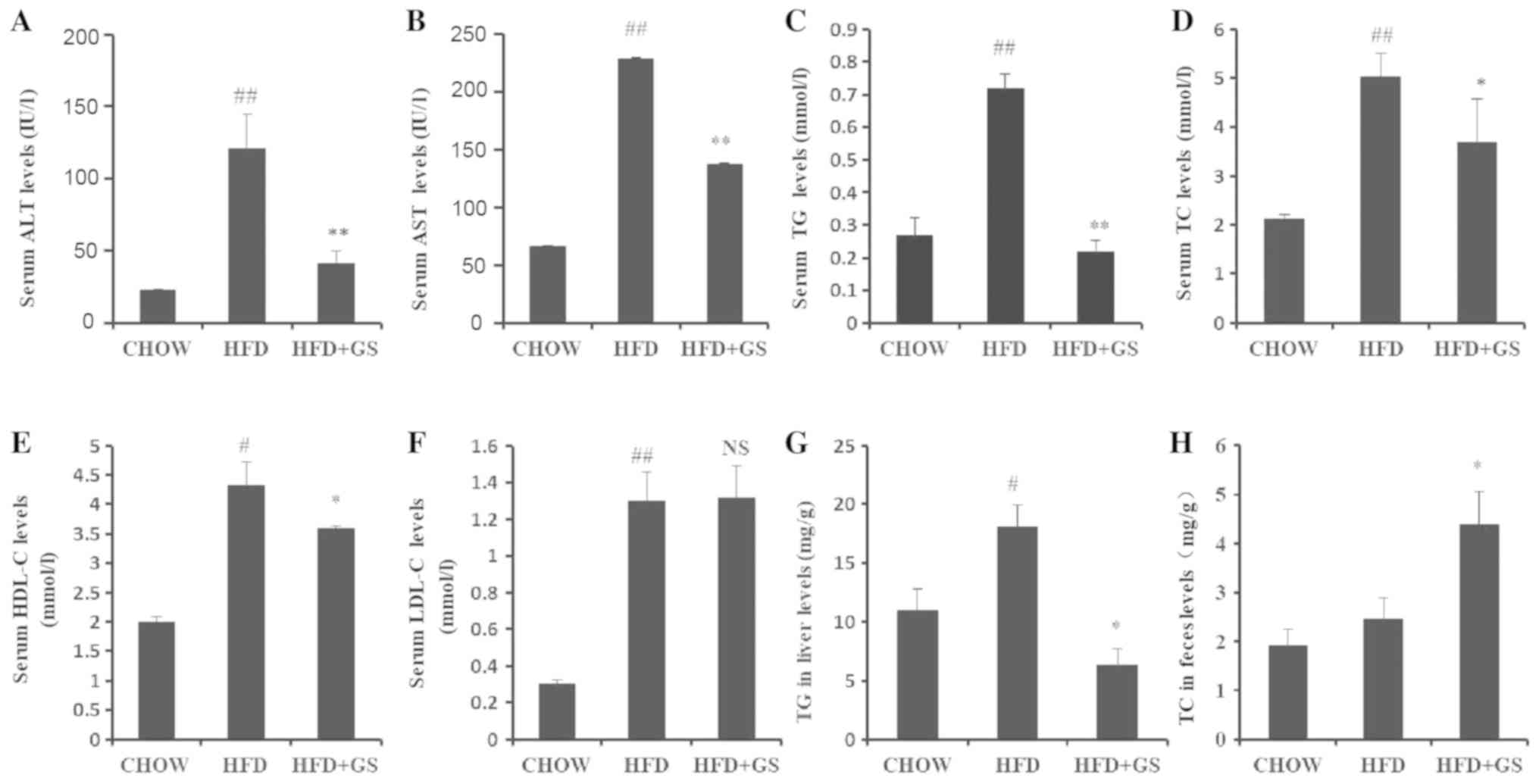 | Figure 3.GS improve the lipid metabolism of
HFD-fed mice. (A) Serum ALT, (B) serum AST, (C) serum TG, (D) serum
TC, (E) serum HDL-c, (F) serum LDL-c, (G) liver TG levels and (H)
feces TG levels. Error bars present the mean ± standard error of
the mean. P-values were determined by one-way ANOVA. *P<0.05 and
**P<0.01. vs. HFD group; #P<0.05 and
##P<0.01 vs. CHOW group; n=5 for all groups. GS,
ginsenosides; HFD, high-fat diet; CHOW, mice administered a
standard chow diet; ALT, alanine aminotransferase; AST, aspartate
aminotransferase; TG, triglyceride; TC, total cholesterol; HDL-c,
high-density lipoprotein cholesterol; LDL-c, low-density
lipoprotein cholesterol; NS, no significance. |
GS alleviates lipid droplet
accumulation in vivo and vitro
Excessive accumulation of TGs in cells is also a
manifestation of the disorder of lipid metabolism in hepatocytes
(45). A hepatocyte steatosis
model was established using complete medium of OA and PA. The
efficacy was evaluated using oil red O staining subsequent to drug
intervention. In the present study, 24 h following the
administration of GS, it was revealed that the oil red O inside the
cells became lighter and the area decreased compared with the OA+PA
well. This suggests that the mixture of OA and PA increased the
lipid accumulation in HepG2 cells, while GS inhibited the lipid
accumulation (Fig. 4A and G;
P<0.05).
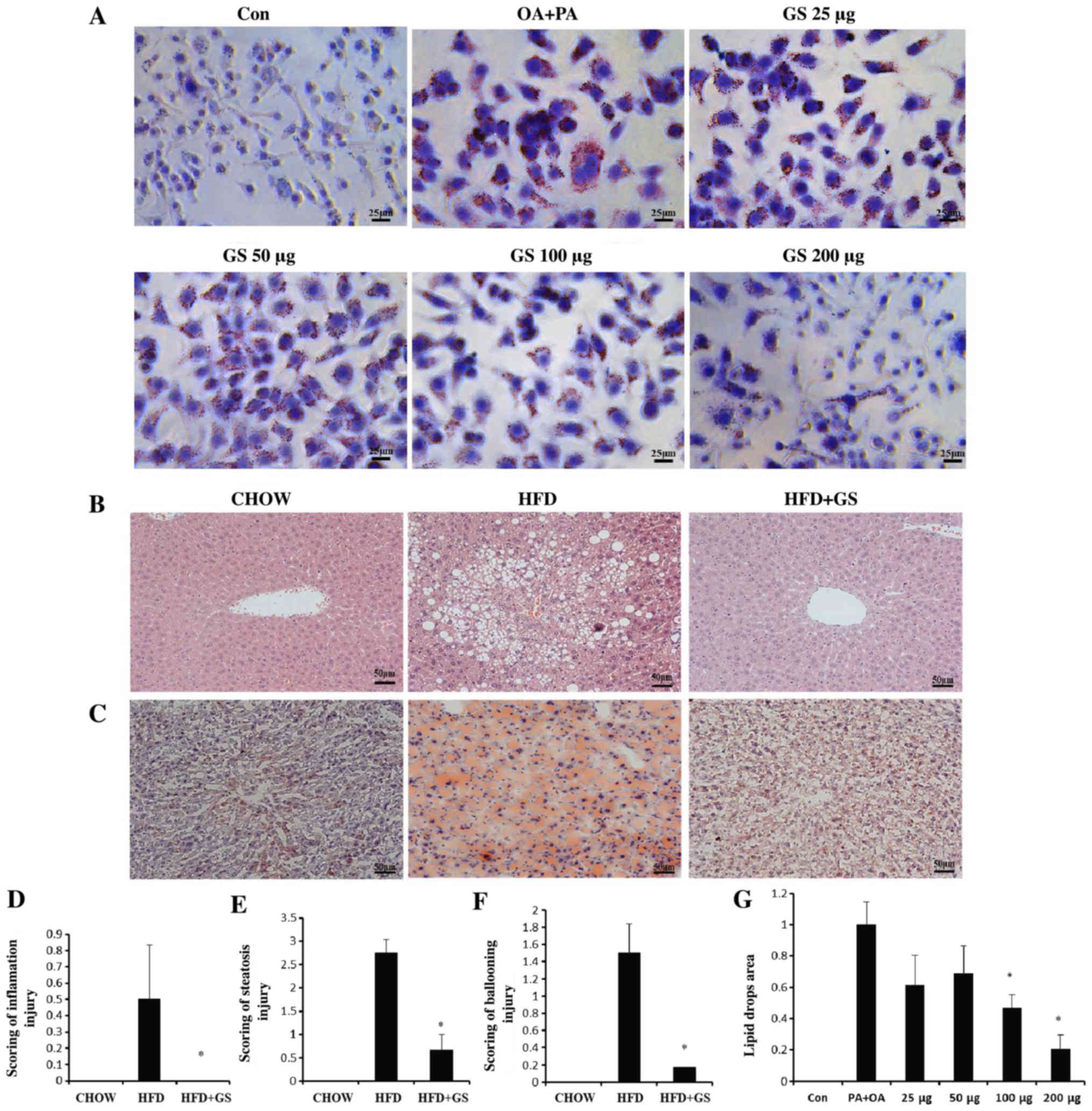 | Figure 4.GS alleviates lipid droplet
accumulation in vivo and in vitro. (A) Oil red O
staining of HepG2 cells (magnification, ×400). (B) Hemoxylin and
eosin and (C) oil red O staining of liver sections (magnification,
×200). Scoring of (D) inflammation injury, (E) steatosis injury and
(F) ballooning injury. (G) Lipid accumulation in HepG2 cells. Error
bars present the mean ± standard error of the mean. P-values were
determined by a one-way ANOVA. Scale bars, 25 or 50 µm. *P<0.05,
vs. HFD group (D-F); *P<0.05 vs. PA + OA group (G); n=5 for all
groups. GS, ginsenosides; HFD, high-fat diet; CHOW, mice
administered a standard chow diet; PA, palmitic acid; OA, oleic
acid. |
Obesity develops into hepatic steatosis, primarily
due to the destruction of the balance between fatty acid storage
and mobilization (46,47). In the present study,
H&E-stained liver sections revealed microvesicular and
macrovesicular steatosis of the liver in the HFD-fed mice compared
with the normal liver structure of the standard chow diet-fed mice.
By contrast, GS treatment efficiently reversed the structural
changes and improved the hepatic steatosis (Fig. 4B). In addition, oil red O staining
is an important method to observe fat accumulation. The liver
sections confirmed that there were few lipids in the livers of the
standard chow diet-fed mice, but a massive lipid accumulation in
the livers of the HFD mice. GS treatment notably mitigated the
lipid accumulation in the livers of the HFD-fed mice (Fig. 4C). Altogether, GS attenuate fatty
liver and reduce the diameter of fat cells in HFD-fed mice
(Fig. 4D-F; P<0.05).
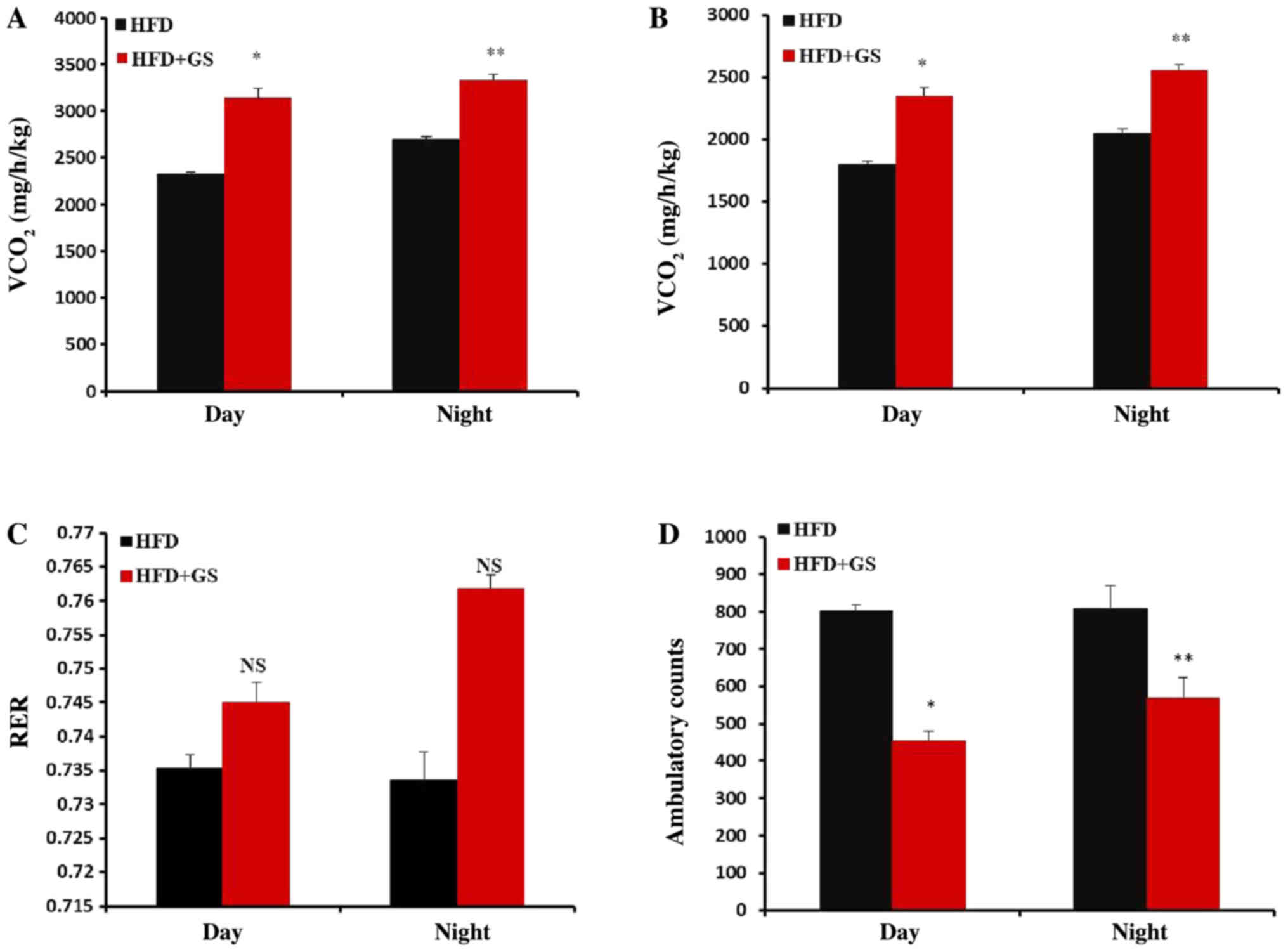
Effect of GS on metabolic
measures
Based on the aforementioned data, it was
hypothesized that GS, despite their robust effects on food intake
and body weight, also enhance oxygen consumption, carbon dioxide
exhalation and result in the utilization of fat as the main energy
source. Therefore following metabolic parameters were measured in
the HFD-fed mice and HFD+GS group: VO2, VCO2,
RER and X-Amb. The results revealed that the HFD+GS group had
significantly increased VO2 and VCO2 compared
with the HFD-fed mice (Fig. 5A and
B; P<0.05 and P<0.01). However, as the two parameters
increased (VO2 and VCO2), their ratio
remained relatively constant (Fig.
5C). Notably, the levels of activity were significantly lower
in the HFD+GS group compared with the HFD group. (Fig. 5D; P<0.05 and P<0.01). There
are two potential reasons for this effect. First, previous studies
have reported that large doses of GS inhibit the activity of mice
(48,49). Second, aside from their appetite,
mice exhibit reduced consumption and exercise levels naturally
(50,51). In conclusion, GS increase the basal
metabolic rate of mice to enhance fat mobilization and thereby
reduce weight.
GS improve liver ERS and regulate
STAT3 phosphorylation in GT1-7 cells and hypothalamus
Specific markers for intracellular STAT3 signaling,
and ERS in the GT1-7 GnRH neuron cell line and the liver tissues
were analyzed using immunoblotting as previously described
(32). As presented in Fig. 6, GS not only reduced the expression
of ERS-associated proteins in GT1-7 cells and the liver (Fig. 6A and B; P<0.05), but also
regulated the p-STAT3/STAT3 ratio in GT1-7 and hypothalamus tissue
(Fig. 6C and D).
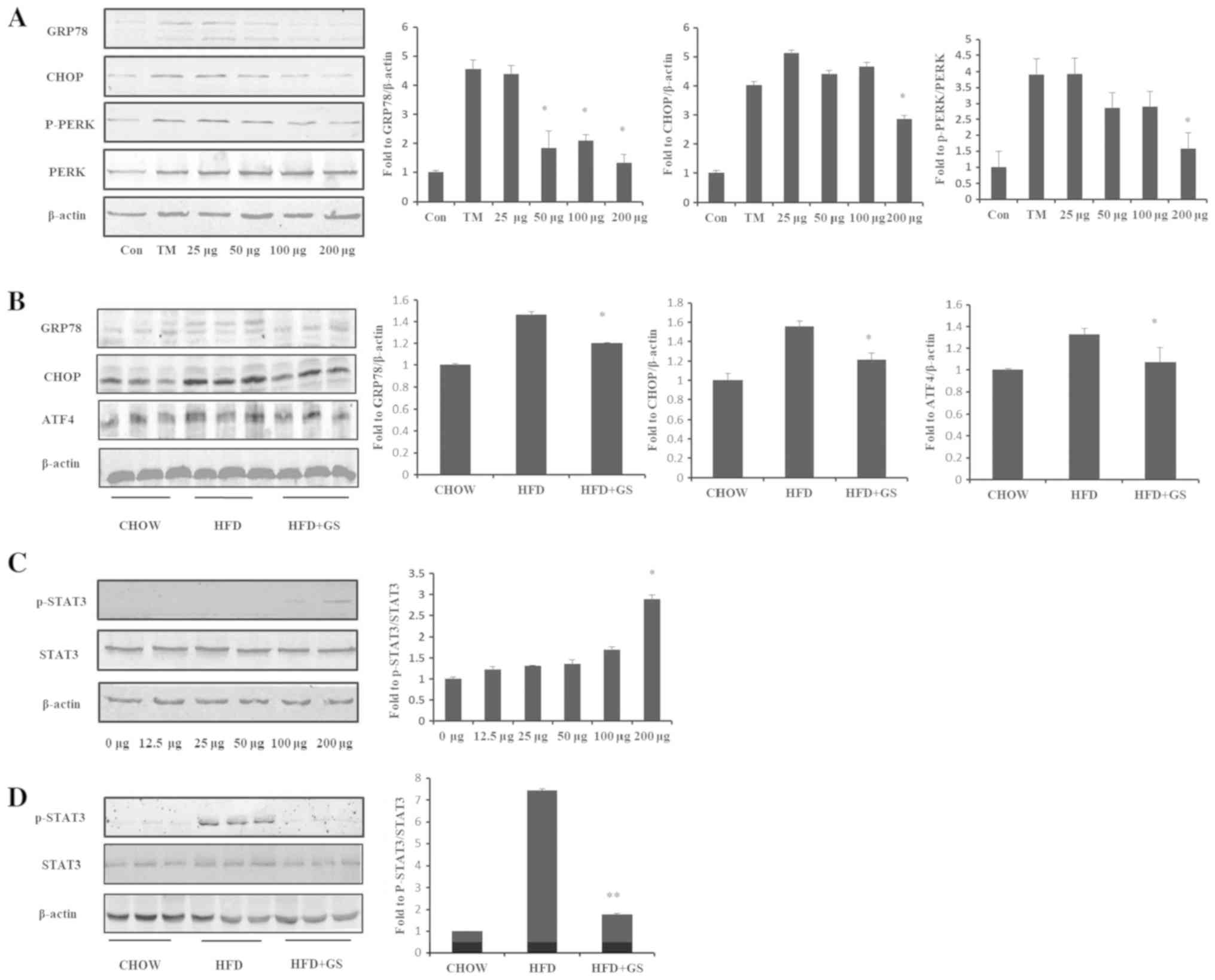 | Figure 6.Effect of GS on p-STAT3/STAT3 and
ERS. Following treatment for 24 h (ERS)/12 h (p-STAT3/STAT3),
western blotting was performed to determine the relative expression
of ERS-associated proteins. (A) Expression of GRP78, CHOP, total
PERK and p-PERK in GT1-7 cells treated with 5 µg/ml TM, 5 µg/ml TM
+ 25 µg GS, 5 µg/ml TM + 50 µg GS, 5 µg/ml TM + 100 µg GS or 5
µg/ml TM + 200 µg GS. *P<0.05 vs. TM. (B) Expression of GRP78,
CHOP and ATF4 in the liver. *P<0.05 vs. HFD group. (C)
Expression of p-STAT3 and total STAT3 in GT1-7 cells treated with
GS (12.5, 25, 50, 100 and 200 µg). *P<0.05 vs. 0 µg. (D)
Expression of p-STAT3 and total STAT3 in the hypothalamus.
**P<0.01 vs. HFD group. GS, ginsenosides; HFD, high-fat diet;
CHOW, mice administered a standard chow diet; STAT3, signal
transducer and activator of transcription 3; p-, phosphorylated;
ERS, endoplasmic reticulum stress; GRP78, glucose regulation
protein 78; CHOP, C/EBP homologous protein; PERK, protein
kinase-like endoplasmic reticulum kinase; ATF4, activating
transcription factor 4; TM, tunicamycin. |
Discussion
The present study identifies a novel function for GS
as a potential weight-reducing medicine based on its function in
reducing obesity, nonalcoholic fatty liver disease, hyperleptinemia
and insulin resistance. The results of the present study
demonstrate that GS alleviate metabolic disorders, including
obesity, fatty liver, dyslipidemia and hyperglycemia in mice.
Furthermore, GS regulate the expression of p-STAT3 and reduce ERS
in GT1-7 cells and the liver. These results reveal that GS reduce
weight and ameliorate nonalcoholic fatty liver disease partly by
increasing the expression of p-STAT3 and reducing the ERS.
China was the first country globally to use and
record ginseng in medical literature. Ginseng has been recorded in
Chinese medical literature for >2,000 years (51). GS are regarded as the main active
ingredient in ginseng, thus becoming the point of focus in research
(52). GS have a wide range of
pharmacological effects and are extensively used to treat numerous
different diseases (53). Examples
include in the treatment of neurodegenerative diseases in the
nervous system, improvement of memory function and protection of
brain tissue (54). In the
cardiovascular system, GS reduce arrhythmia, cardiac hypertrophy,
myocardial ischemia and myocardial apoptosis (24). In terms of tumor types, they may
induce apoptosis, inhibit tumor cell proliferation, regulate signal
pathways and regulate immune functions (55). With regards to metabolism, they
inhibit the absorption of fatty food in the intestinal tract by
reducing the activity of PL or through the regulation of the
lipogenesis transcription factor PPAR and the glucolipid metabolism
regulation molecule AMPK and its target gene expression (56). The compound used in the present
study contained up to 80% GS. Thus, it is rational to propose that
GS may be one of the ingredients responsible for the activities of
ginseng.
In the present study, the GS treatment reduced the
body weight gain induced by HFD and food intake was markedly
reduced, indicating that the effect was caused by a reduction in
calorie intake (21). The results
also revealed that the treatment lowered blood glucose, serum AST,
serum ALT, serum TG and liver TG contents and increased the fecal
excretion of TG in the mice fed a HFD, suggesting that GS
effectively lower HFD-induced hyperglycemia and hyperlipidemia
(57).
Decreased food intake may be associated with
drug-induced liver injury (58,59).
Thus, in the present study, the effect of drug toxicity on the
feeding of the HFD-fed mice was investigated extensively.
Therefore, H&E and Oil Red O staining were performed on the
liver tissues to observe the effect of the drug, and the results
demonstrated that the reduction of dietary intake was not caused by
drug-induced liver injury (60).
This was supported by further evidence, including low levels of ALT
and AST in the serum (61). In
addition, GS reduced lipid accumulation in vivo and in
vitro (21). The present
results are consistent with previous reports.
Obesity usually causes hyperinsulinemia and
hyperleptinemia in the blood circulation, which mainly manifest as
reduced glucose tolerance, leptin resistance and dyslipidemia
(62,63). Leptin and insulin resistance were
induced in mice following 12 weeks on a HFD (64). In the present study, GS
administration significantly reduced the levels of circulating
insulin and leptin in the HFD-fed mice (P<0.05). The combination
of the results of the glucose tolerance and insulin tolerance tests
indicate that GS improve leptin and insulin resistance in HFD-fed
mice (21).
Previously, an increasing number of studies have
demonstrated that leptin resistance was a major cause of obesity
(65–67). The important reasons are the
regulation of p-STAT3 and increased ERS in the hypothalamus
(68,69).
STAT3 is a member of the STAT protein family
(70). Leptin receptor is
activated by leptin and p-STAT3 (71). Thus, STAT3 is a classical
up-regulator of leptin signaling. For example, Liu et al
(72) reported that celastrol
suppresses appetite, reduces body weight and ameliorates disorders
of glucose and lipid metabolism through p-STAT3. Ibars et al
(68) reported that
proanthocyanidins reduce appetite and weight by increasing
phosphorylated STAT3. Interestingly, the present study produced
opposing results in vivo and in vitro. The potential
reason for this is that there is no ligand of leptin in cells to
bind to the leptin receptor, and the effect of increased
phosphorylation of STAT3 is all derived from GS. However, in
vivo, the excess ligand binding of hyperleptinemia in the serum
of obese mice ameliorated the decrease in the receptor ligand
binding rate caused by receptor passivation, so the results were
inconsistent. In addition, age and other factors would also cause
different results (8,73–76).
The endoplasmic reticulum is an extremely active and
important organelle in the cell (76,77).
It is involved in protein synthesis, storage, processing and
modification, and folding (78).
Dysfunction of the endoplasmic reticulum results in the
accumulation of unfolded or misfolded proteins in the endoplasmic
reticulum, which results in ERS (79). There are three mechanisms to
survive a stress response in the endoplasmic reticulum: The ATF6
mechanism, the IRE1-splice X-box binding protein 1 mechanism and
the PERK-eIF2 mechanism (80). The
results of the present study indicate that GS reduce ERS via the
PERK-eIF2-α mechanism. For example, Joe et al (81) reported that fibroblast growth
factor 21 induced by carbon monoxide mediated metabolic homeostasis
via the PERK/ATF4 pathway. Cai et al (82) reported that the alleviation of ERS
by aerobic exercise was able to suppress appetite and reduce weight
in HFD-fed mice. Wang et al (83) revealed that the alleviation of ERS
was able to reduce nonalcoholic fatty liver disease. Furthermore,
Fujii et al (84)
demonstrated that reducing ERS does reduce nonalcoholic fatty liver
disease. Notably, the present study reports that GS significantly
reduce ERS through the p-PERK/PERK/ATF4 pathway.
The results of the present study provide evidence
that GS serve an important function in weight loss and ameliorating
nonalcoholic fatty liver disease in HFD-fed mice. GS also
upregulated the phosphorylated protein expression of STAT3 and
reduced ERS in vitro and ameliorated nonalcoholic fatty
liver disease in vivo. GS may be a novel choice as a highly
efficient non-toxic strategy for the treatment of metabolic
disorders. The present study still has numerous limitations. Future
studies ought to: i) Detect the constituents of total ginsenosides
with high-performance liquid chromatography, ii) Observe the feed
intake of the obese mice following the injection of GS into the
lateral ventricle and iii) Analyze the hypothalamic samples (the
dorsomedial hypothalamus, the ventromedial hypothalamus and the
arcuate nucleus) using immunostaining with p-STAT3
(Tyr705)-specific antibodies. In conclusion, results from the
present study demonstrated that GS reduced weight and improved
non-alcoholic fatty liver, and identified possible pathways
regulating p-STAT3 and alleviating endoplasmic reticulum stress
pathway.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
Experiment design, operation and writing the
manuscript was completed by YY.
Ethics approval and consent to
participate
The present study was conducted in accordance with
the ethical standards and according to the Ethical Committee of
Shanghai University of Traditional Chinese Medicine (Shanghai,
China). The protocols were ethically approved by the Institutional
Animal Care and Use Committee of Shanghai University of Traditional
Chinese Medicine (approval no. SZY201708002).
Patient consent for publication
Not applicable.
Competing interests
The author declares that they have no competing
interests.
References
|
1
|
Kinlen D, Cody D and O'Shea D:
Complications of obesity. QJM. 111:437–443. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ahirwar R and Mondal PR: Prevalence of
obesity in India: A systematic review. Diabetes Metab Syndr.
13:318–321. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu L, Huang X, You C, Li J, Hong K, Li P,
Wu Y, Wu Q, Wang Z, Gao R, et al: Prevalence of overweight,
obesity, abdominal obesity and obesity-related risk factors in
southern China. PLoS One. 12:e01839342017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Yilmaz Y and Younossi ZM:
Obesity-associated nonalcoholic fatty liver disease. Clin Liver
Dis. 18:19–31. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fruh SM: Obesity: Risk factors,
complications, and strategies for sustainable long-term weight
management. J Am Assoc Nurse Pract. 29 (Suppl 1):S3–S14.
2017.PubMed/NCBI
|
|
6
|
Sáinz N, Barrenetxe J, Moreno-Aliaga MJ
and Martínez JA: Leptin resistance and diet-induced obesity:
Central and peripheral actions of leptin. Metabolism. 64:35–46.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Eseberri I, Lasa A, Miranda J, Gracia A
and Portillo MP: Potential miRNA involvement in the anti-adipogenic
effect of resveratrol and its metabolites. PLoS One.
12:e01848752017. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Xu L, Li H, Zhou G, Lu W, Yang R, Liu H
and Yang G: DNA-binding activity of STAT3 increased in hypothalamus
of DIO mice; the reduction of STAT3 phosphorylation may facilitate
leptin signaling. Biochem Biophys Res Commun. 505:229–235. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Y, Lu W, Jin Z, Yu J and Shi B:
Carbenoxolone ameliorates hepatic lipid metabolism and inflammation
in obese mice induced by high fat diet via regulating the
JAK2/STAT3 signaling pathway. Int Immunopharmacol. 74:1054982019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Habib DF, Fahmi AA, Kholousy NM, Amin AI,
Shalaby M, Ahmed MM and Shanab AMA: The role of liver in leptin
metabolism in experimental nephrotic syndrome. EXCLI J. 10:322–331.
2011.PubMed/NCBI
|
|
11
|
Bergmann TJ and Molinari M: Three branches
to rule them all? UPR signaling in response to chemicallyversus
misfolded proteins-induced ER stress. Biol Cell. 110:197–204. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saito A: Physiological functions of
endoplasmic reticulum stress transducer OASIS in central nervous
system. Anat Sci Int. 89:11–20. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Yilmaz E: Endoplasmic reticulum stress and
obesity. Adv Exp Med Biol. 960:261–276. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ding S, Jiang J, Zhang G, Bu Y, Zhang G
and Zhao X: Resveratrol and caloric restriction prevent hepatic
steatosis by regulating SIRT1-autophagy pathway and alleviating
endoplasmic reticulum stress in high-fat diet-fed rats. PLoS One.
12:e01835412017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kozuka C, Shimizu-Okabe C, Takayama C,
Nakano K, Morinaga H, Kinjo A, Fukuda K, Kamei A, Yasuoka A, Kondo
T, et al: Marked augmentation of PLGA nanoparticle-induced
metabolically beneficial impact of ү-oryzanol on fuel
dyshomeostasis in genetically obese-diabetic ob/ob mice. Drug
Deliv. 24:558–568. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Abenavoli L, Di Renzo L, Boccuto L,
Alwardat N, Gratteri S and De Lorenzo A: Health benefits of
Mediterranean diet in nonalcoholic fatty liver disease. Expert Rev
Gastroenterol Hepatol. 12:873–881. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Shin J, He M, Liu Y, Paredes S, Villanova
L, Brown K, Qiu X, Nabavi N, Mohrin M, Wojnoonski K, et al: SIRT7
represses Myc activity to suppress ER stress and prevent fatty
liver disease. Cell Rep. 5:654–665. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Henkel A and Green RM: The unfolded
protein response in fatty liver disease. Semin Liver Dis.
33:321–329. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sozio MS, Liangpunsakul S and Crabb D: The
role of lipid metabolism in the pathogenesis of alcoholic and
nonalcoholic hepatic steatosis. Semin Liver Dis. 30:378–390. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Y, Tang C, Tian Y, Yuan H, Gao R and
Cao J: Effects of electroacupuncture combined with dietary control
on liver endoplasmic reticulum stress in rats with non-alcoholic
fatty liver disease. Zhongguo Zhen Jiu. 36:951–956. 2016.(In
Chinese). PubMed/NCBI
|
|
21
|
Li Z and Ji GE: Ginseng and obesity. J
Ginseng Res. 42:1–8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Shin BK, Kwon SW and Park JH: Chemical
diversity of ginseng saponins from Panax ginseng. J Ginseng Res.
39:287–298. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Christensen LP: Chapter 1 Ginsenosides:
Chemistry, biosynthesis, analysis, and potential health effects.
Adv Food Nutr Res. 55:1–99. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim JH: Pharmacological and medical
applications of Panax ginseng and ginsenosides: A review for use in
cardiovascular diseases. J Ginseng Res. 42:264–269. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Cui ZY, Jo E, Jang HJ, Hwang HI, Lee KB,
Yoo HS, Park SJ, Jung MK, Lee YW and Jang IS: Modified ginseng
extract induces apoptosis in HepG2 cancer cells by blocking the
CXCL8-mediated Akt/Nuclear Factor-κB signaling pathway. Am J Chin
Med. 46:1645–1662. 2018. View Article : Google Scholar
|
|
26
|
Lee JB, Yoon SJ, Lee SH, Lee MS, Jung H,
Kim TD, Yoon SR, Choi I, Kim IS, Chung SW, et al: Ginsenoside Rg3
ameliorated HFD-induced hepatic steatosis through downregulation of
STAT5-PPARү. J Endocrinol. 235:223–235. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Etou H, Sakata T, Fujimoto K, Terada K,
Yoshimatsu H, Ookuma K, Hayashi T and Arichi S: Ginsenoside-Rb1 as
a suppressor in central modulation of feeding in the rat. Nihon
Yakurigaku Zasshi. 91:9–15. 1988.(In Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hurtado-Carneiro V, Sanz C, Roncero I,
Vazquez P, Blazquez E and Alvarez E: Glucagon-Like Peptide 1
(GLP-1) can reverse AMP-activated protein kinase (AMPK) and S6
Kinase (P70S6K) activities induced by fluctuations in glucose
levels in hypothalamic areas involved in feeding behaviour. Mol
Neurobiol. 45:348–361. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Z and Ji GE: Ginseng and obesity. J
Ginseng Res. 42:1–8. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wu LX, Xu YY, Yang ZJ and Feng Q:
Hydroxytyrosol and olive leaf extract exert cardioprotective
effects by inhibiting GRP78 and CHOP expression. J Biomed Res.
32:371–379. 2018.PubMed/NCBI
|
|
31
|
Fan S, Zhang Y, Sun Q, Yu L, Li M, Zheng
B, Wu X, Yang B, Li Y and Huang C: Extract of okra lowers blood
glucose and serum lipids in high-fat diet-induced obese C57BL/6
mice. J Nutr Biochem. 25:702–709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gao J, Bai H, Li Q, Li J, Wan F, Tian M,
Li Y, Song Y, Zhang J and Si Y: In vitro investigation of
the mechanism underlying the effect of ginsenoside on the
proliferation and differentiation of neural stem cells subjected to
oxygen-glucose deprivation/reperfusion. Int J Mol Med. 41:353–363.
2018.PubMed/NCBI
|
|
33
|
Luo L, Yin L, Wang D, Zhao Y, Wang Y, Li
F, Fang J, Chen H, Fan S and Huang C: Ginkgolide B lowers body
weight and ameliorates hepatic steatosis in high-fat diet-induced
obese mice correlated with pregnane X receptor activation. RSC Adv.
7:37858–37866. 2017. View Article : Google Scholar
|
|
34
|
Arif A, Terenzi F, Potdar AA, Jia J, Sacks
J, China A, Halawani D, Vasu K, Li X, Brown JM, et al: EPRS is a
critical mTORC1-S6K1 effector that influences adiposity in mice.
Nature. 542:357–361. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kleiner DE, Brunt EM, Van Natta M, Behling
C, Contos MJ, Cummings OW, Ferrell LD, Liu YC, Torbenson MS,
Unalp-Arida A, et al: Design and validation of a histological
scoring system for nonalcoholic fatty liver disease. Hepatology.
41:1313–1321. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
la Fuente FP, Quezada L, Sepúlveda C,
Monsalves-Alvarez M, Rodríguez JM, Sacristán C, Chiong M, Llanos M,
Espinosa A and Troncoso R: Exercise regulates lipid droplet
dynamics in normal and fatty liver. Biochim Biophys Acta Mol Cell
Biol Lipids. 1864:1585192019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gerlini R, Berti L, Darr J, Lassi M,
Brandmaier S, Fritsche L, Scheid F, Böhm A, Königsrainer A,
Grallert H, et al: Glucose tolerance and insulin sensitivity define
adipocyte transcriptional programs in human obesity. Mol Metab.
18:42–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Coccia F, Testa M, Guarisco G, Di
Cristofano C, Silecchia G, Leonetti F, Gastaldelli A and Capoccia
D: Insulin resistance, but not insulin response, during oral
glucose tolerance test (OGTT) is associated to worse histological
outcome in obese NAFLD. Nutr Metab Cardiovasc Dis. Aug 10–2019.doi:
10.1016/j.numecd.2019.08.001 (Epub ahead of print). PubMed/NCBI
|
|
39
|
Rosenzweig T, Skalka N, Rozenberg K,
Elyasiyana U, Pinkusb A, Greenb B, Stanevskyb M and Drori E: Red
wine and wine pomace reduced the development of insulin resistance
and liver steatosis in HFD-fed mice. J Funct Foods. 34:379–389.
2017. View Article : Google Scholar
|
|
40
|
Garg N, Thakur S, McMahan CA and Adamo ML:
High fat diet induced insulin resistance and glucose intolerance
are gender-specific in IGF-1R heterozygous mice. Biochem Biophys
Res Commun. 413:476–480. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Loftus TM, Maggs DG and Lane MD: The
adipose tissue/central nervous system axis. Diabetologia. 40 (Suppl
3):B16–B20. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Svensson AM, Hellerstrom C and Jansson L:
Diet-induced obesity and pancreatic islet blood flow in the rat: A
preferential increase in islet blood perfusion persists after
withdrawal of the diet and normalization of body weight. J
Endocrinol. 151:507–511. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zakrzewska KE, Cusin I, Stricker-Krongrad
A, Boss O, Ricquier D, Jeanrenaud B and Rohner-Jeanrenaud F:
Induction of obesity and hyperleptinemia by central glucocorticoid
infusion in the rat. Diabetes. 48:365–370. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Strader AD, Reizes O, Woods SC, Benoit SC
and Seeley RJ: Mice lacking the syndecan-3 gene are resistant to
diet-induced obesity. J Clin Invest. 114:1354–1360. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Müller C, Gardemann A, Keilhoff G, Peter
D, Wiswedel I and Schild L: Prevention of free fatty acid-induced
lipid accumulation, oxidative stress, and cell death in primary
hepatocyte cultures by a Gynostemma pentaphyllum extract.
Phytomedicine. 19:395–401. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Sheng L, Cho KW, Zhou Y, Shen H and Rui L:
Lipocalin 13 protein protects against hepatic steatosis by both
inhibiting lipogenesis and stimulating fatty acid β-oxidation. J
Biol Chem. 286:38128–38135. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Gao Q, Jia Y, Yang G, Zhang X, Boddu PC,
Petersen B, Narsingam S, Zhu YJ, Thimmapaya B, Kanwar YS and Reddy
JK: PPARα-deficient ob/ob obese mice become more obese and manifest
severe hepatic steatosis due to decreased fatty acid oxidation. Am
J Pathol. 185:1396–1408. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Takagi K, Saito H and Tsuchiya M: Effect
of panax ginseng root on spontaneous movement and exercise in mice.
Jpn J Pharmacol. 24:41–48. 1974. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Watanabe H, Ohta H, Imamura L, Asakura W,
Matoba Y and Matsumoto K: Effect of Panax ginseng on age-related
changes in the spontaneous motor activity and dopaminergic nervous
system in the rat. Jpn J Pharmacol. 55:51–56. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lee J, Liu J, Feng X, Salazar Hernández
MA, Mucka P, Ibi D, Choi JW and Ozcan U: Withaferin A is a leptin
sensitizer with strong antidiabetic properties in mice. Nat Med.
22:1023–1032. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Xu Y, Ding J, An JN, Qu YK, Li X, Ma XP,
Zhang YM, Dai GJ and Lin N: Effect of the Interaction of Veratrum
Nigrum with Panax ginseng on estrogenic activity in vivo and in
vitro. Sci Rep. 6:269242016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Zheng M, Xin Y, Li Y, Xu F, Xi X, Guo H,
Cui X, Cao H, Zhang X and Han C: Ginsenosides: A potential
neuroprotective agent. Biomed Res Int. 2018:81743452018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Riaz M, Rahman NU, Zia-Ul-Haq M, Jaffar
HZE and Manea R: Ginseng: A dietary supplement as immune-modulator
in various diseases. Trends Food Sci Technol. 83:12–30. 2019.
View Article : Google Scholar
|
|
54
|
Kim KH, Lee D, Lee HL, Kim CE, Jung K and
Kang KS: Beneficial effects of Panax ginseng for the treatment and
prevention of neurodegenerative diseases: Past findings and future
directions. J Ginseng Res. 42:239–247. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Chen T, Li B, Qiu Y, Qiu Z and Qu P:
Functional mechanism of Ginsenosides on tumor growth and
metastasis. Saudi J Biol Sci. 25:917–922. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kim JH, Yi YS, Kim MY and Cho JY: Role of
ginsenosides, the main active components of Panax ginseng, in
inflammatory responses and diseases. J Ginseng Res. 41:435–443.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Zhang L, Virgous C and Si H: Ginseng and
obesity: Observations and understanding in cultured cells, animals
and humans. J Nutr Biochem. 44:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Majumdar SK, Shaw GK, O'Gorman P and
Thomson AD: The effect of naftidrofuryl on ethanol-induced liver
damage in chronic alcoholic patients. Drug Alcohol Depend.
10:135–142. 1982. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Musialik J, Suchecka W, Klimacka-Nawrot E,
Petelenz M, Hartman M and Błońska-Fajfrowska B: Taste and appetite
disorders of chronic hepatitis C patients. Eur J Gastroenterol
Hepatol. 24:1400–1405. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Kim HG, Jang SS, Lee JS, Kim HS and Son
CG: Panax ginseng Meyer prevents radiation-induced liver injury via
modulation of oxidative stress and apoptosis. J Ginseng Res.
41:159–168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Niranjana Murthy H, Dandin VS and Yoeup
Paek K: Hepatoprotective activity of ginsenosides from Panax
ginseng adventitious roots against carbon tetrachloride treated
hepatic injury in rats. J Ethnopharmacol. 158:442–446. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Chen W, Balland E and Cowley MA:
Hypothalamic insulin resistance in obesity: Effects on glucose
homeostasis. Neuroendocrinology. 104:364–381. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Hofmann A, Peitzsch M, Brunssen C, Mittag
J, Jannasch A, Frenzel A, Brown N, Weldon SM, Eisenhofer G,
Bornstein SR and Morawietz H: Elevated steroid hormone production
in the db/db mouse model of obesity and type 2 diabetes. Horm Metab
Res. 49:43–49. 2017.PubMed/NCBI
|
|
64
|
Chen HJ and Liu J: Actein ameliorates
hepatic steatosis and fibrosis in high fat diet-induced NAFLD by
regulation of insulin and leptin resistant. Biomed Pharmacother.
97:1386–1396. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Crujeiras AB, Carreira MC, Cabia B,
Andrade S, Amil M and Casanueva FF: Leptin resistance in obesity:
An epigenetic landscape. Life Sci. 140:57–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Adam CL and Findlay PA: Decreased
blood-brain leptin transfer in an ovine model of obesity and weight
loss: Resolving the cause of leptin resistance. Int J Obes (Lond).
34:980–988. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Myers MG Jr, Leibel RL, Seeley RJ and
Schwartz MW: Obesity and leptin resistance: Distinguishing cause
from effect. Trends Endocrinol Metab. 21:643–651. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ibars M, Ardid-Ruiz A, Suárez M, Muguerza
B, Bladé C and Aragonès G: Proanthocyanidins potentiate
hypothalamic leptin/STAT3 signalling and Pomc gene expression in
rats with diet-induced obesity signalling. Int J Obes (Lond).
41:129–136. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Contreras C, Fondevila MF and López M:
Hypothalamic GRP78, a new target against obesity? Adipocyte.
7:63–66. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Raptis L, Arulanandam R, Geletu M and
Turkson J: The R(h)oads to Stat3: Stat3 activation by the Rho
GTPases. Exp Cell Res. 317:1787–1795. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Koga S, Kojima A, Ishikawa C, Kuwabara S,
Arai K and Yoshiyama Y: Effects of diet-induced obesity and
voluntary exercise in a tauopathy mouse model: Implications of
persistent hyperleptinemia and enhanced astrocytic leptin receptor
expression. Neurobiol Dis. 71:180–192. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Liu J, Lee J, Salazar Hernandez MA,
Mazitschek R and Ozcan U: Treatment of Obesity with Celastrol.
Cell. 161:999–1011. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Sahin-Efe A, Polyzos SA, Dincer F,
Zaichenko L, McGovern R, Schneider B and Mantzoros CS:
Intracellular leptin signaling following effective weight loss.
Metabolism. 64:888–895. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Scarpace PJ, Matheny M and Shek EW:
Impaired leptin signal transduction with age-related obesity.
Neuropharmacology. 39:1872–1879. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Scarpace PJ and Tümer N: Peripheral and
hypothalamic leptin resistance with age-related obesity. Physiol
Behav. 74:721–727. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Scarpace PJ, Matheny M and Tümer N:
Hypothalamic leptin resistance is associated with impaired leptin
signal transduction in aged obese rats. Neuroscience.
104:1111–1117. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Oakes SA and Papa FR: The role of
endoplasmic reticulum stress in human pathology. Annu Rev Pathol.
10:173–194. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Schwarz DS and Blower MD: The endoplasmic
reticulum: Structure, function and response to cellular signaling.
Cell Mol Life Sci. 73:79–94. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Ye Z, Liu G, Guo J and Su Z: Hypothalamic
endoplasmic reticulum stress as a key mediator of obesity-induced
leptin resistance. Obes Rev. 19:770–785. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Tsuchiya Y, Saito M and Kohno K:
Pathogenic mechanism of diabetes development due to dysfunction of
unfolded protein response. Yakugaku Zasshi. 136:817–825. 2016.(In
Japanese). View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Joe Y, Kim S, Kim HJ, Park J, Chen Y, Park
HJ, Jekal SJ, Ryter SW, Kim UH and Chung HT: FGF21 induced by
carbon monoxide mediates metabolic homeostasisviathe PERK/ATF4
pathway. FASEB J. 32:2630–2643. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Cai M, Wang H, Li JJ, Zhang YL, Xin L, Li
F and Lou SJ: The signaling mechanisms of hippocampal endoplasmic
reticulum stress affecting neuronal plasticity-related protein
levels in high fat diet-induced obese rats and the regulation of
aerobic exercise. Brain Behav Immun. 57:347–359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Wang D, Lao L, Pang X, Qiao Q, Pang L,
Feng Z, Bai F, Sun X, Lin X and Wei J: Asiatic acid from Potentilla
chinensis alleviates non-alcoholic fatty liver by regulating
endoplasmic reticulum stress and lipid metabolism. Int
Immunopharmacol. 65:256–267. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Fujii J, Homma T, Kobayashi S and Seo HG:
Mutual interaction between oxidative stress and endoplasmic
reticulum stress in the pathogenesis of diseases specifically
focusing on non-alcoholic fatty liver disease. World J Biol Chem.
9:1–15. 2018. View Article : Google Scholar : PubMed/NCBI
|