Introduction
Ischemic stroke (IS), which is a major cause of
disability and mortality worldwide, is a cerebrovascular disease
caused by cerebral blood supply disorder or hemorrhage and results
in the formation of an infarction zone with tissue necrosis and
softening (1,2). It can lead to the severe impairment
of sensory and motor dysfunction, neurodegenerative diseases and
sudden mortality (3). Cerebral
infarction not only seriously affects the life quality of patients,
but also imposes a huge burden and pressure on public health care
and the social economy (4). A
previous study reported that the impact caused by IS will become
more apparent in the next two decades as the world's population
ages and lifestyles change, especially in developing countries
(5). An epidemiological study has
showed that >70% of patients with IS currently have mental and
physical functional disorders (6).
Tissue plasminogen activator is the only thrombolytic drug approved
by the US Food and Drug Administration for the treatment of acute
cerebral infarction, but its clinical application is limited by the
timing of stenosis (onset) and side effects such as bleeding
(7,8). A number of other previous studies
have reported that transient cerebral ischemia/hypoxia can lead to
neuronal death, but the mechanism is not fully understood (9,10).
Therefore, further understanding of the pathogenesis of cerebral
infarction, finding biomarkers that can be used to diagnose and
predict prognosis, and eliciting new therapeutic targets have
become a significant and urgent task.
MicroRNAs (miRNAs) are highly conserved, endogenous
non-coding single-stranded small RNAs that are ~22–24 nucleotides
long and can bind to target mRNAs with a complementary nucleic acid
sequence. In general, miRNAs act post-transcriptionally to inhibit
the translation of the target mRNAs or degrade the target mRNA
(11). Each miRNA can regulate the
expression of multiple target genes, and the same gene can also be
regulated by several different miRNAs (12). Currently, >2,000 human miRNAs
have been identified, and it is hypothesized that these small
molecules regulate about one-third of human genes (13). Therefore, miRNA changes can lead to
various diseases. A number of studies have demonstrated that miRNAs
serve important biological roles in cell growth, proliferation,
differentiation and apoptosis, as well as reconstruction of
hematopoietic and damaged tissues (14,15).
Therefore, the role of miRNAs in neurological diseases has
attracted increasing attention. In the nervous system, miRNAs are
closely related to the growth and differentiation of neurons,
memory and learning functions, as well as neurological degenerative
diseases and mental disorders (16,17).
In addition, abnormal miRNA expression is detected in brain tumors,
Parkinson's disease, Alzheimer's disease and IS (18–20).
Studies on the identification of target genes for
miRNAs associated with cerebral infarction suggest that miRNAs play
a key role in the development and progression of cerebral
infarction (21,22). Among them, apoptosis is the central
link of this process, and related studies have indicated that
apoptosis is a highly regulated and controlled death process
induced by miRNAs (23). In the
present study, bioinformatics analysis was performed to investigate
the putative genes and miRNAs involved in the pathogenesis and
disease process of IS, and the mechanism was studied at the
molecular level, which provided a reference for molecular diagnosis
and individualized treatment of IS.
Materials and methods
Microarray data
Two mRNA microarray datasets, GSE22255 and GSE16561,
and one miRNA profile, GSE110993, of IS were obtained from the Gene
Expression Omnibus (GEO) database (https://www.ncbi.nlm.nih.gov/geo). GSE22255 is based
on the GPL570 (HG-U133_Plus_2) Affymetrix Human Genome U133 Plus
2.0 Array. Gene expression profiling was performed on peripheral
blood mononuclear cells (PBMCs) of 20 patients with IS (10 male; 10
female) and 20 sex- and age-matched healthy control (CTL) patients
using Affymetrix microarrays. GSE16561 contained a total of 63
samples based on the platform of GPL6883 Illumina HumanRef-8 v3.0
expression beadchip, in which peripheral blood samples were
collected in Paxgene Blood RNA tubes from patients who were >18
years of age with MRI diagnosed IS (n=39; 22 females and 17 males)
and CTL (n=24; 14 females and 10 males) who were non-stroke
neurologically healthy. As for GSE110993, RNA sequencing results
were analyzed to study expression changes of circulating miRNAs in
a discovery sample of 20 patients (10 males and 10 females) with IS
and 20 sex- and age-matched healthy control (CTLs) patients was
further applied in independent samples for validation (40 patients
with IS and 40 CTL patients) and replication (200 patients with IS,
100 CTLs).
Identification of differentially
expressed genes (DEGs)
Background correction and quartile data
normalization of the downloaded data were performed using the
robust multi-array average algorithm (24). Principal component analysis was
subsequently conducted to reduce the dimension of multivariate data
to two or three principal components, which can be visualized
graphically with the least information loss. We used R software to
calculate and visualize the principal component analysis (25). Probes without a corresponding gene
symbol were then filtered and the average value of gene symbols
with multiple probes was calculated using R software (26). DEGs between two groups were
screened by Student's t-test and fold-change (FC) filtering with R
software limma package (27). With
a threshold of P<0.05 and |FC|>2, volcano plots were
constructed to identify the DEGs with statistical significance
between two groups by R software ggplot2 package (data not shown)
(28). In addition, hierarchical
clustering and combined analyses were performed for the DEGs. The
GSE16561 dataset was used to validate top 20 of DEGs in the
GSE22255 dataset to identify overlapping DEGs in two
microarrays.
Protein-Protein Interaction (PPI)
network construction and analysis
The PPI network was created by integrating
Biological General Repository for Interaction Datasets (BioGRID)
and IntAct databases at EMBL-EBI. First, the PPI pairs were used to
construct the PPI network. Second, the regulatory relationship
between genes was visualized by using Cytoscape (version 3.4.0)
(29), and analyzed through a
topological property of computing network including the degree
distribution of network by using the CentiScape app version 2.2
(30).
Functional enrichment analysis
The Database for Annotation, Visualization and
Integrated Discovery (DAVID) Bioinformatics resource (http://david.abcc.ncifcrf.gov) was used to conduct
functional enrichment analysis to understand the underlying
biological processes and pathways of aberrantly expressed
intersecting genes.
Gene Ontology (GO) terms for the biological process
(BP) were enriched. The most enriched term within a cluster was
chosen as the one to represent the cluster. Only terms with
P<0.05 and the concentrated gene numbers ≥1.5 were considered
meaningful (≥1 gene number exists).
Prediction of miRNA targets
The target genes for differentially expressed (DE)
miRNAs in GSE110993 were predicted using three established
programs: TargetScan version 7.2.0 (31), miRTarBase version 7.0 (32) and miRDB version 5.03 (33), which are the most popular databases
of experimentally validated miRNA interactions. The genes that were
predicted by all three programs were chosen as the targets of DE
miRNAs and the miRNA-gene network was constructed using miRWalk 2.0
(34). Cytoscape (https://cytoscape.org; version 3.40) was used to
visualize the miRNA regulatory network.
Results
Identification of differentially
expressed genes in female and male IS patients
A total of 10 female and 10 male patients with IS,
and 20 sex- and age-matched respective CTLs from the data in the
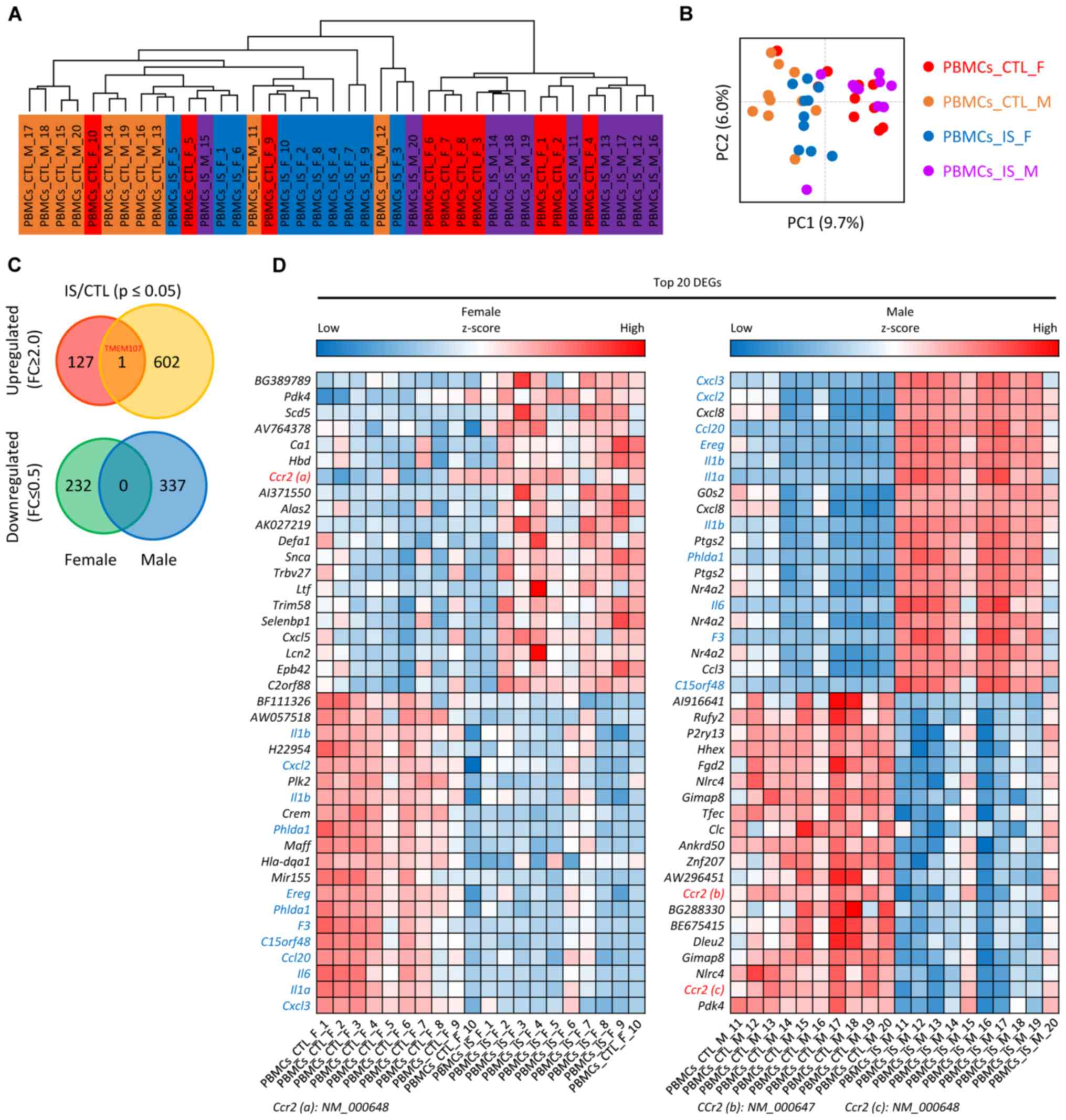
GEO database (GSE22255) were clustered and divided into four groups
(Fig. 1A): i) PBMCs_IS_M group,
ii) PBMCs_IS_F group, iii) PBMCs_CTL_M group and iv) PBMCs_CTL_F
group, as depicted in Fig. 1A.
Principle component analysis of genes in patients with IS and CTL
is presented in Fig. 1B. Following
data preprocessing, the unique genes in the four groups of samples
were analyzed and it was determined that a total of 1,300 genes
were differentially expressed in the IS samples compared with the
CTL samples. Among these DEGs, 128 genes were upregulated in
females, 603 genes were upregulated in males, 232 were genes
downregulated in females and 337 genes were downregulated in males.
For instance, it was found that the chemokines C-X-C motif
chemokine (CXCL)3, CXCL2 and CXCL8 were significantly upregulated
in male IS patients. However, TMEM107 was the only overlapped
upregulated DEG between the female and male group, which is
displayed in a Venn diagram (Fig.
1C). A heat map of the top 20 up- and downregulated DEGs in
male and female groups is shown in Fig. 1D.
Functional and pathway enrichment
analyses
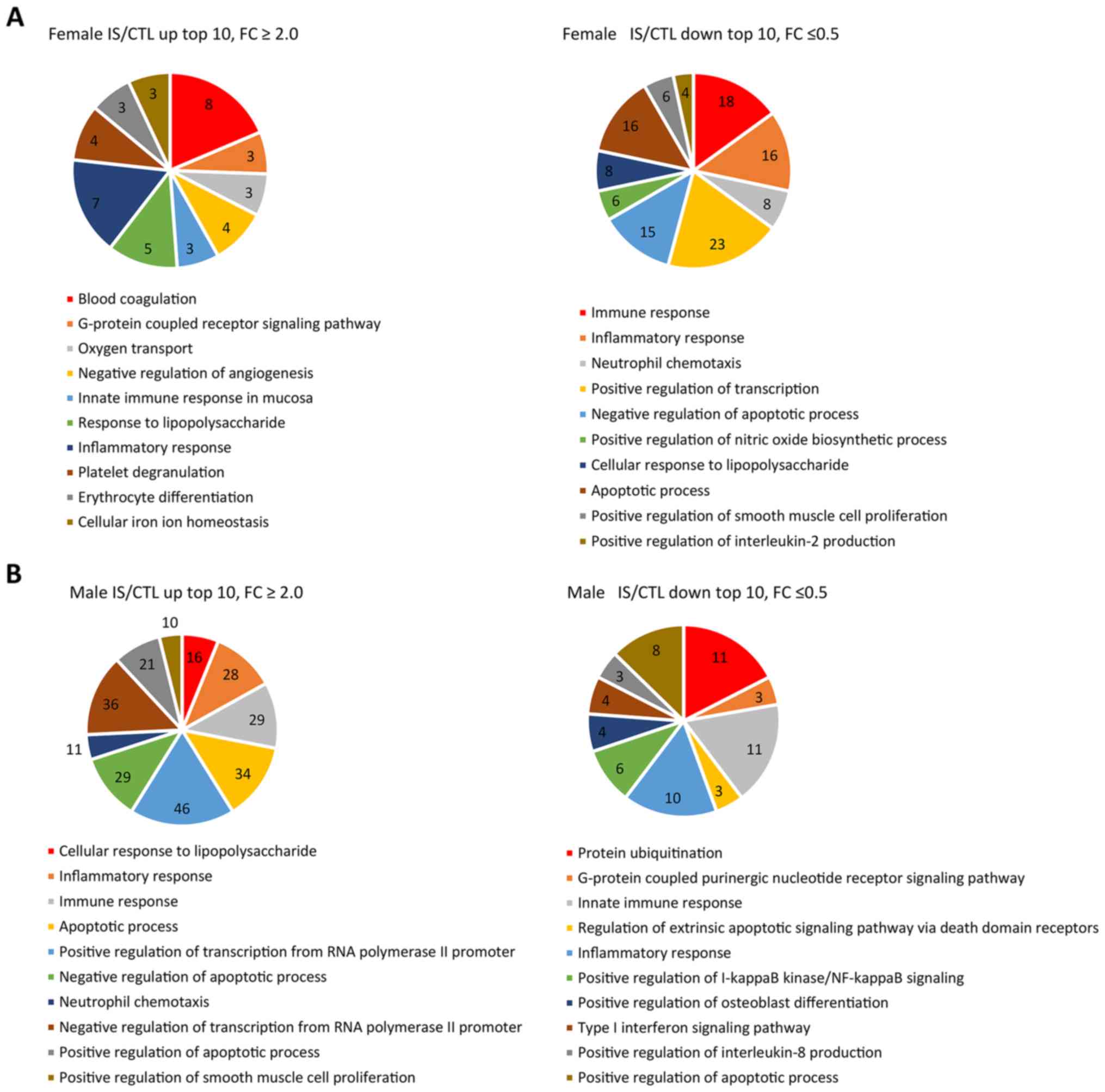
DAVID was used to perform GO term enrichment
analyses to explore the function and pathways of 1,300 identified
DEGs (Fig. 2). GO-BP term
enrichment analysis demonstrated that the upregulated genes were
significantly enriched in ‘blood coagulation’ and ‘inflammatory
response’ in the female IS compared with the CTL group. In
addition, ‘positive regulation of transcription from RNA polymerase
II promoter’ and ‘neutrophil chemotaxis’ were enriched in the male
IS compared with the CTL group (Fig.
2A). Downregulated genes were mainly involved in ‘positive
regulation of transcription’ and ‘immune response’ in the female IS
group (Fig. 2B). It was also
revealed that most of the downregulated genes were involved in
‘inflammatory response’ and ‘protein ubiquitination’ in the male IS
group, respectively. Of note, in both the male and female groups,
the pathways of differential expression were mainly involved in
‘immune response’, ‘inflammatory response’, ‘apoptotic process’ and
‘neutrophil chemotaxis’.
Construction of PPI network and module
analysis
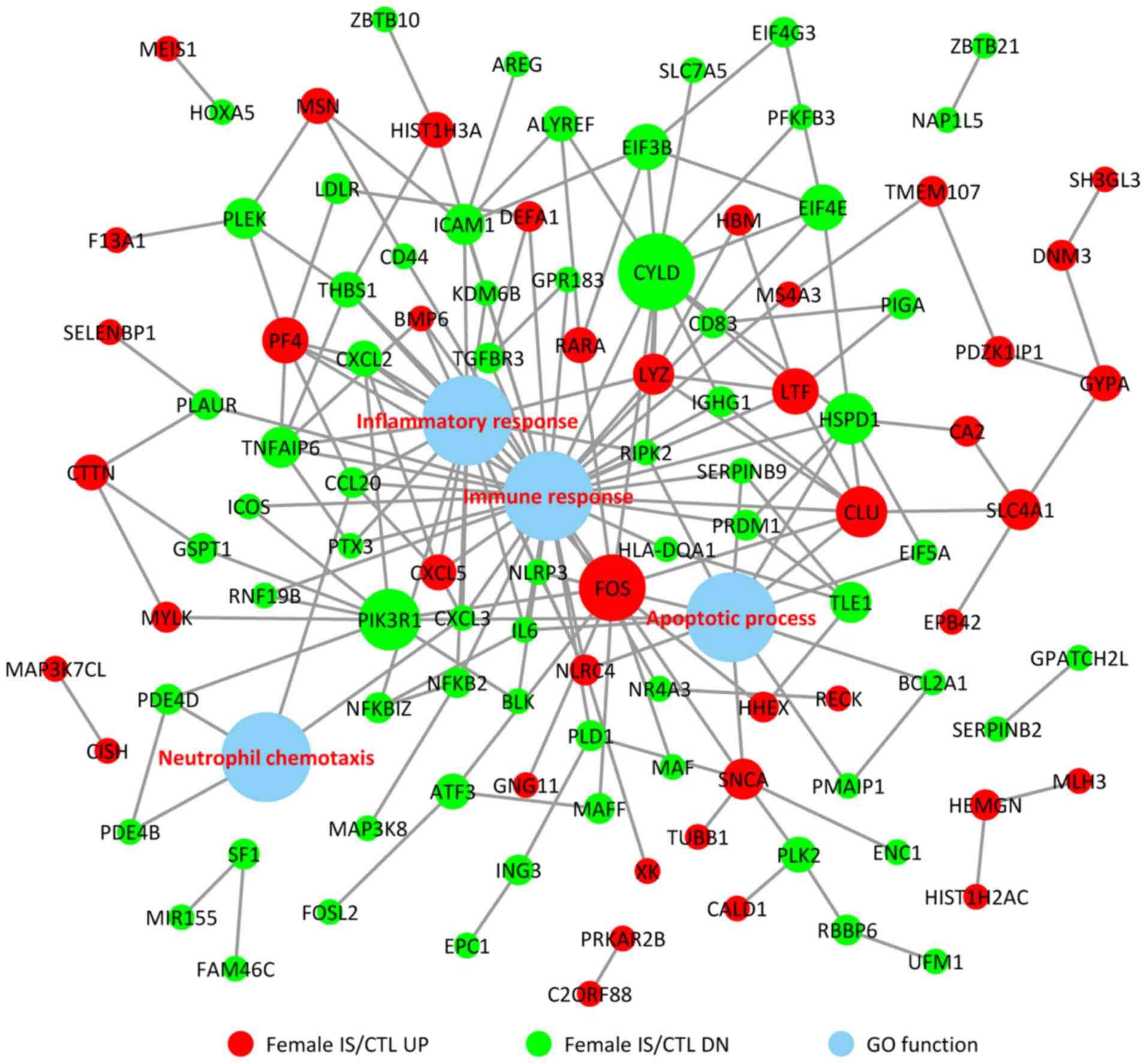
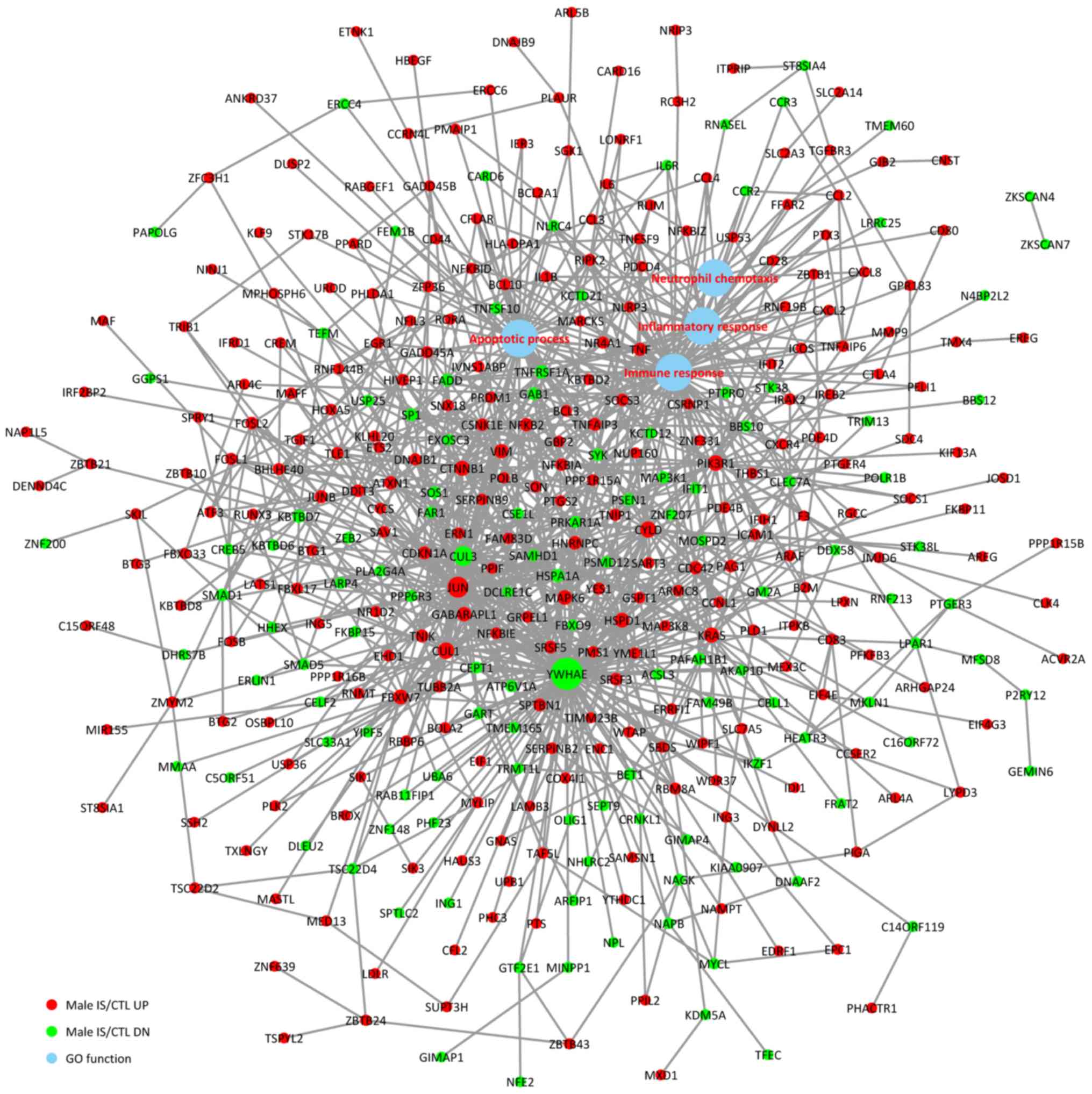
To investigate the associations between the DEGs and
BPs, a comprehensive analysis of protein interactions of the top
up- and downregulated DEGs was conducted using BioGRID, IntAct
database and Cytoscape software (Fig.
3). Direct or indirect interactions were identified, and the
highest-scoring nodes were screened as central genes in the female
IS/CTL group, including FOS, CLU, LTF, PF4, CXCL5 and CYLD
(Fig. 3). In the male IS/CTL
group, the top 3 genes with the most significant expression were
CUL3, YWHAE and JUN (Fig. 4). The
associations between BP and gene expression were categorized into 4
processes, namely inflammatory response, immune response, apoptotic
process and neutrophil chemotaxis.
miRNA target regulatory network
analysis
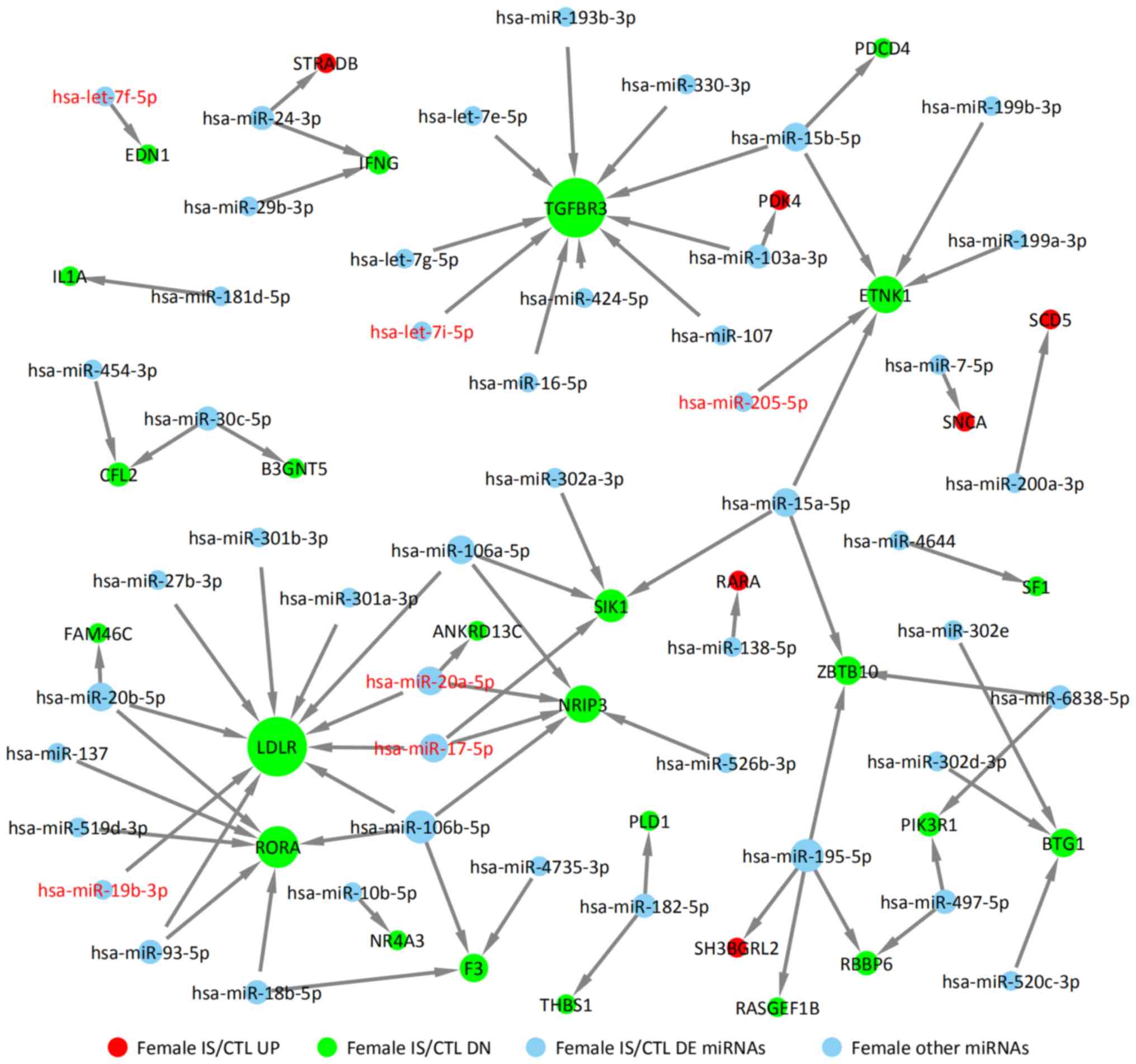
DE miRNAs obtained from GSE110993 database were
analyzed. For the female IS/CTL group, six DE miRNAs were
identified, including let-7f-5p, let-7i-5p, miR-17-5p, miR-20a-5p,
miR-19b-3p and miR-205-5p (Fig.
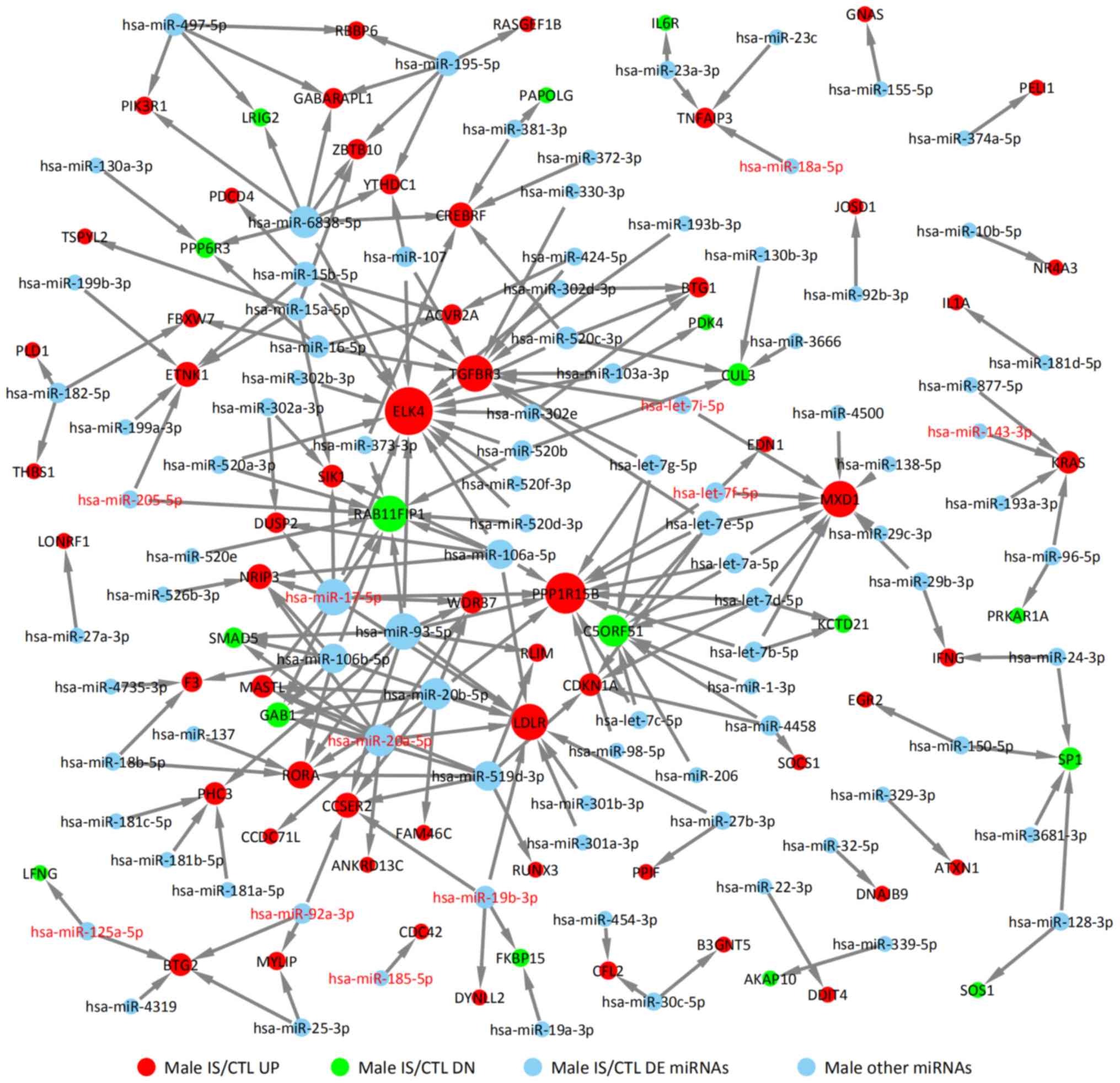
5). In the male IS/CTL group, there were 11 DE miRNA
identified, such as miR-205-5p, miR-17-5p, miR-20a-5p, miR-125a-5p,
miR-92a-3p, miR-18a-5p and miR-143-3p (Fig. 6). Their target genes were predicted
using TargetScan, miRTarBase and miRDB. The upregulated targets
were STRADB, PDK4, SCD5, SNCA, RARA and SHBGRL2 in the female IS
group (solid red circles in Fig.
5); the downregulated target genes are also indicated,
including TGFBR3, ETNK1, LDLR, RORA and RBBP6 (solid green circles
in Fig. 5). The targets of miRNAs
for the male IS/CTL group are shown in Fig. 6 (solid green circles represent
downregulated target genes, such as RAB11FIP1, C5ORF51, GAB1 and
SMAD5, and red circles represented upregulated target genes such as
PPP1R15B, ELK4, MXD1 and LDLR).
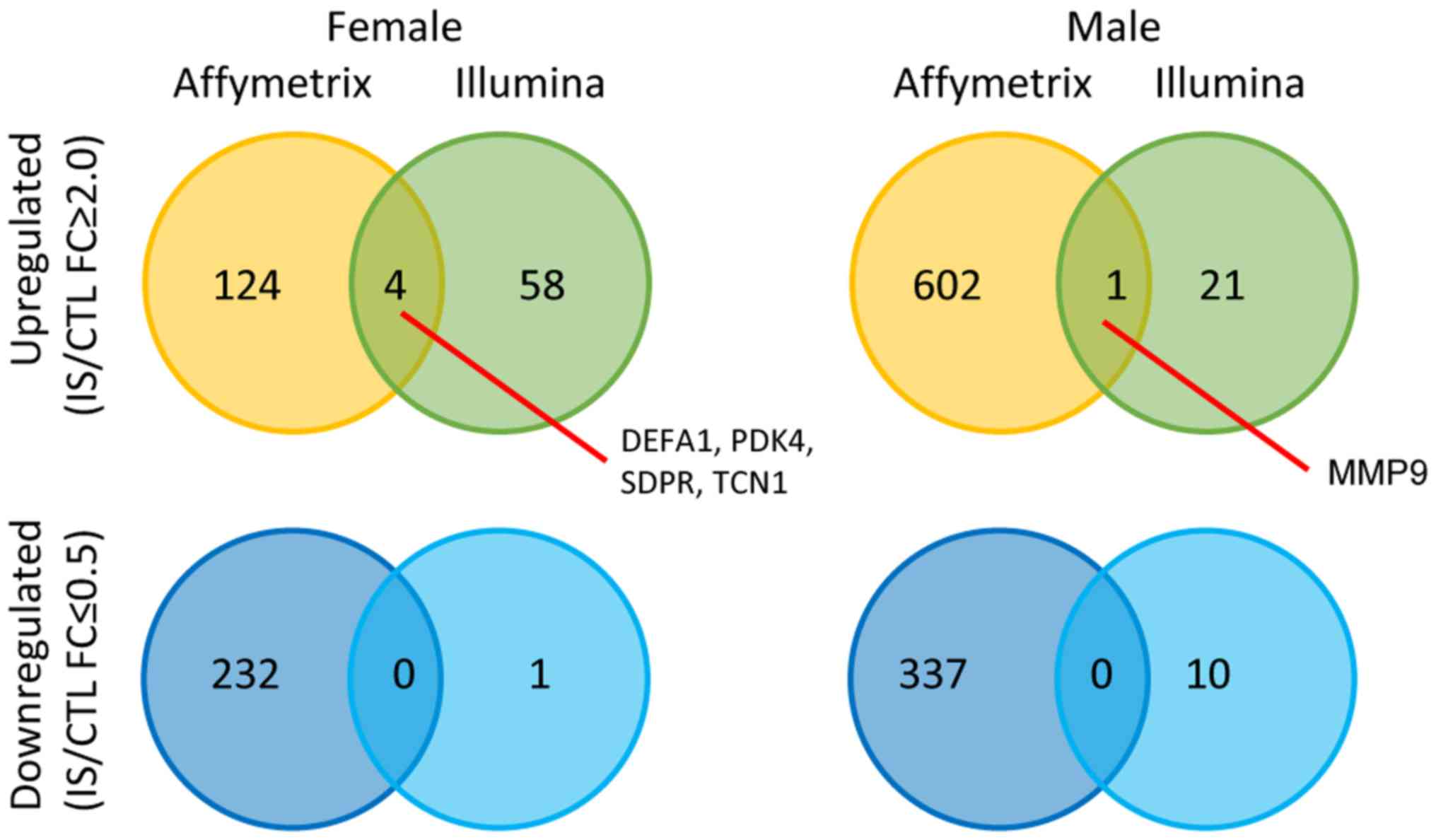
Validation of DEGs in IS
Crosscheck validation analysis of female and male IS
patient DEGs from GSE22255 and another set of Illumina HumanRef-8
v3.0 expression beadchip (GSE16561) are shown in Fig. 7. There were 4 genes that are
upregulated in both datasets in the female group including defensin
α 1 (DEFA1), pyruvate dehydrogenase kinase 4 (PDK4), caveolae
associated protein 2 (SDPR) and transcobalamin 1 (TCN1) and only
one shared upregulated gene in the male group, matrix
metalloproteinase 9 (MMP9). These findings suggested that the
differential expression targets selected had certain repeatability.
Although numerous studies have been published regarding DEGs as
candidate biomarkers for IS (35–37),
the reliability of these findings remains uncertain, as they were
generated from the studies without sufficient evidence of
reproducibility and independent clinical validation.
Discussion
As a high-throughput genomics research technology,
gene chips serve an important role in datamining of potential
biological information and exploring the structure and function of
the genome (38). Gene expression
profiling experiments are mainly used to determine DEGs under
different physiological or pathological conditions (39). Such analysis will facilitate the
exploration of the relationship between health and disease at the
molecular level and the potential mechanism through network
regulation. However, the etiology and physiological pathology of IS
are still not fully understood. Therefore, further clarifying the
pathogenesis of IS, determining its risk factors and developing
effective therapeutic strategies will significantly improve the
clinical outcome of stroke treatment.
The present study used bioinformatics tools and
methods to screen the key DEGs and their functions in stroke. A
total of 128 upregulated genes and 231 downregulated genes were
found in female patients with IS, and 604 upregulated genes and 337
downregulated genes in male patients with IS. These DEGs were
mainly associated with inflammatory response and apoptosis, and
other biological pathways. Combining PPI and GO term enrichment
results, it was also found that the genes involved in the GO
analysis pathway were almost identical to the PPI network
nodule-related genes, especially the CXCL family; tumor necrosis
factor and PDK were most closely related to these pathways, which
was consistent with the reported results.
Following a stroke, the damaged brain tissue
releases proinflammatory chemokines, which trigger the inflammatory
pathway in immune cells (40).
Depending on the sequence of N-terminal residues, chemokines can be
divided into several different types, including CXC, CC, XC and
CX3C, where C is an N-terminal cysteine residue and X represents
any amino acid residue (41). The
chemokine ligands and receptors are emerging as potential
therapeutic targets for cardiovascular diseases and have become
increasingly studied in recent years. Brait et al (42) found that the expression of CXCR2
and its ligands CXCL1 and CXCL2 is significantly upregulated and
peaks 1–3 days after injury in a middle cerebral artery occlusion
(MCAO) rat model. Villa et al (43) inhibited CXCR1 with interleukin
(IL)-8/CXCL8 receptor inhibitors in MCAO model rats and
demonstrated that CXCR2, myeloperoxidase and IL-1A expression
levels decreased, which reduced the symptoms of neurological
deficits. In the present study, it was found that the chemokine
receptors CXCL3, CXCL2 and CXCL8 were significantly upregulated in
male patients with IS, and this was consistent with the previous
reports.
PDK4 is a key enzyme that regulates pyruvate
dehydrogenase complex (PDC) activity and is a key regulator of
pyruvate oxidation and glucose maintenance in vivo (44). In calcified vascular smooth muscle
cells and calcified vessels of patients with atherosclerosis, PDK4
expression is found to be upregulated alongside increased PDC
phosphorylation, suggesting that PDK4 serves an important role in
vascular calcification (45). In
phosphate-treated vascular smooth muscle cells, PDK4 expression is
upregulated, inducing mitochondrial dysfunction and, subsequently,
apoptosis. Atherosclerosis is the result of cholesterol and lipid
deposition on the arterial wall, often related to calcification,
which is formed by the deposition of inorganic calcium on the
arterial wall. Thus, the upregulation of PDK4 increased osteogenic
markers and enhanced vascular calcification without significant
effects on bone formation (46).
This indicated that PDK4 may be a new target for the treatment of
vascular calcification (47,48).
In the pathogenesis of various injuries, the immune
inflammatory signaling pathway caused by ischemia serves a major
role, whereas ischemia itself serves a secondary role (49). In IS the key role of the immune
inflammatory response has been widely recognized, but whether it
plays a role in brain damage or brain protection may depend on the
degree and stage of ischemia. Secondary injuries are caused by the
inflammatory cascade following ischemia. Various inflammatory
mediators, including cytokines, chemokines and cell adhesion
molecules, aggravate ischemic brain damage (27,28).
In the early stage of ischemia, the inflammatory response
exacerbates brain damage, and in the late stage of ischemia, it
serves an important role in the recovery of nerve function and
tissue repair. In addition, patients with IS are prone to a
hypercoagulable state and advanced fibrinolysis. Most patients have
coagulation changes in the clinic, which may lead to adverse events
such as thrombosis and infarction, which affects the prognosis
(50).
miRNAs are important post-transcriptional regulators
that coordinate the regulation of target genes with multi-target
mRNA. Risk factors involved in IS include hypertension,
atherosclerosis, atrial fibrillation, diabetes and dyslipidemia.
The role of miRNAs in the pathophysiology of stroke has become a
focus of interest in recent research (51). A previous study reported that
circulating miRNA expression in peripheral blood is relatively
stable (52), suggesting that
differential expression of specific miRNAs following stroke is
expected to be a potential biomarker for diagnosis and prognosis,
providing an effective theoretical basis for elucidating the
pathogenesis of IS (53).
In the present study, the GSE110993 dataset was
analyzed and 17 differentially expressed miRNAs (6 miRNAs in
females and 11 miRNAs in males) were screened. According to
previous studies, there are differences in the expression patterns
of miRNAs in IS and in normal healthy people (54). For example, the abnormal expression
of miR-221 is related to metabolic syndrome, whereas the expression
of miR-221 in healthy people was significantly higher compared with
expression levels in patients with IS (55). Studies have demonstrated that the
abnormal expression level of miR-221 is significant for
distinguishing healthy individual and those with cardiovascular and
cerebrovascular diseases, tumor populations and endocrine
populations (56,57). Other previous studies have shown
that miR-143/145 (57) and
miR-125-5p (58) can also be used
as markers for the risk of IS, which is consistent with the
findings of the present study. miR-143/145 is most abundant in
normal vascular tissues compared with other miRNAs and is selective
in vascular smooth muscle cells expression. miR-125-5p has been
found to be significantly upregulated in atherosclerosis (58). Inhibition of miR-125-5p expression
significantly increases the uptake of lipids and enhances the
clearance of human expression and secretion of inflammatory cell
molecules (59). Some of these
inflammatory cell molecules (60)
may be involved in the inflammatory response of ox-LDL-stimulated
monocytes, whereas ox-LDL may be taken up by macrophages, converted
into foam cells that secrete pro-inflammatory factors and leads to
cell necrosis and plaque formation. This series of chronic
inflammatory reactions is atherosclerosis.
There are several limitations to the present study.
First, it is notable that only one upregulated gene overlapped and
none of downregulated genes overlapped between male and female
groups. This may be due to differences in the effects of different
types of sex hormones on platelet function. Androgens increase
platelet activity, whereas estrogen inhibits its function (61). This may have an effect on the
relative expression levels of mRNA and miRNA in the blood,
suggesting that doctors might consider different medications for
stroke patients of different sexes (62). Secondly, the data in Fig. 7 do not give confidence in the
high-throughput genomics analyses because there are only 4 genes
that are upregulated in both dataset in the female group and only
one shared gene upregulated in the male group. This phenomenon may
be because GSE22255 is based on GPL570 Affymetrix Human Genome U133
Plus 2.0 Array, whereas GSE16561 contained a total of 63 samples
based on the platform of GPL6883 Illumina HumanRef-8 v3.0
expression beadchip. On the one hand, these two different gene
platforms contain different numbers and different types of genes;
on the other hand, there may be detection errors in chips from
Illumina and Affymetrix.
In summary, all of the miRNAs described above have a
certain correlation with the pathogenesis and development of IS,
which is consistent with the findings of stroke researchers
worldwide in recent years. The correlation between some miRNAs
selected in this study have not been reported previously in the
literature to the best of our knowledge. The possibility that these
miRNAs are novel biomarkers for IS and its biological significance
still needs further study. Further studies on the role of miRNAs
will provide a theoretical basis for understanding the molecular
mechanism of the pathogenesis and development of IS, as well as new
research programs for prevention and treatment.
Acknowledgements
Not applicable.
Funding
This work was funded by HeBei Natural Science
Foundations (grant no. H2017307016).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
LL designed the experiments; HW and SS performed the
experiments; LD and JC conducted the statistical analysis of the
data and drafted the manuscript; CZ and CB conceived the study,
participated in its design and coordination, and helped to draft
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ishak B, Abdul-Jabbar A, Singla A, Yilmaz
E, von Glinski A, Ramey WL, Blecher R, Tymchak Z, Oskouian R and
Chapman JR: Intraoperative ischemic stroke in elective spine
surgery: A retrospective study of incidence and risk. Spine (Phila
Pa 1976). 45:109–115. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Weiss HR, Mellender SJ, Kiss GK, Liu X and
Chi OZ: Improvement in microregional oxygen supply/consumption
balance and infarct size after cerebral ischemia-reperfusion with
inhibition of p70 ribosomal S6 kinase (S6K1). J Stroke Cerebrovasc
Dis. 28:1042762019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Weisberg LA: ‘Stroke and struck’:
Protecting the brain from cerebrovascular disease. South Med J.
96:3312003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Singh RB, Suh IL, Singh VP, Chaithiraphan
S, Laothavorn P, Sy RG, Babilonia NA, Rahman AR, Sheikh S,
Tomlinson B and Sarraf-Zadigan N: Hypertension and stroke in Asia:
Prevalence, control and strategies in developing countries for
prevention. J Hum Hypertens. 14:749–763. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
West R, Hill K, Hewison J, Knapp P and
House A: Psychological disorders after stroke are an important
influence on functional outcomes: A prospective cohort study.
Stroke. 41:1723–1727. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ickenstein GW, Horn M, Schenkel J,
Vatankhah B, Bogdahn U, Haberl R and Audebert HJ: The use of
telemedicine in combination with a new stroke-code-box
significantly increases t-PA use in rural communities. Neurocrit
Care. 3:27–32. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Keller L, Hobohm C, Zeynalova S, Classen J
and Baum P: Does treatment with t-PA increase the risk of
developing epilepsy after stroke? J Neurol. 262:2364–2372. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rudnick SI, Swaminathan J, Sumaroka M,
Liebhaber S and Gewirtz AM: Effects of local mRNA structure on
posttranscriptional gene silencing. Proc Natl Acad Sci USA.
105:13787–13792. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Park JH, Ahn JH, Song M, Kim H, Park CW,
Park YE, Lee TK, Lee JC, Kim DW, Lee CH, et al: A 2-min transient
ischemia confers cerebral ischemic tolerance in non-obese gerbils,
but results in neuronal death in obese gerbils by increasing
abnormal mTOR activation-mediated oxidative stress and
neuroinflammation. Cell. 8:11262019. View Article : Google Scholar
|
|
10
|
Liu S, Jin R, Xiao AY, Zhong W and Li G:
Inhibition of CD147 improves oligodendrogenesis and promotes white
matter integrity and functional recovery in mice after ischemic
stroke. Brain Behav Immun. 82:13–24. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feschotte C, Jiang N and Wessler SR: Plant
transposable elements: Where genetics meets genomics. Nat Rev
Genet. 3:329–341. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sunkar R and Zhu JK: Novel and
stress-regulated microRNAs and other small RNAs from Arabidopsis.
Plant Cell. 16:2001–2019. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cheng AM, Byrom MW, Shelton J and Ford LP:
Antisense inhibition of human miRNAs and indications for an
involvement of miRNA in cell growth and apoptosis. Nucleic Acids
Res. 33:1290–1297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Crippa S, Cassano M and Sampaolesi M: Role
of miRNAs in muscle stem cell biology: Proliferation,
differentiation and death. Curr Pharm Des. 18:1718–1729. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kumar P, Wu H, McBride JL, Jung KE, Kim
MH, Davidson BL, Lee SK, Shankar P and Manjunath N: Transvascular
delivery of small interfering RNA to the central nervous system.
Nature. 448:39–43. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mehler MF and Mattick JS: Non-coding RNAs
in the nervous system. J Physiol. 575:333–341. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jung HL, Choi J, Hwang YK, Kim DH, Yoo KH,
Sung KW and Koo HH: Inhibition of survivin expression by small
interfering RNA (siRNA) in brain tumor cell lines. Cancer Res.
65:2005.
|
|
18
|
Pantano L, Friedländer MR, Escaramís G,
Lizano E, Pallarès-Albanell J, Ferrer I, Estivill X and Martí E:
Specific small-RNA signatures in the amygdala at premotor and motor
stages of Parkinson's disease revealed by deep sequencing analysis.
Bioinformatics. 32:673–681. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Roy J, Sarkar A, Parida S, Ghosh Z and
Mallick B: Small RNA sequencing revealed dysregulated piRNAs in
Alzheimer's disease and their probable role in pathogenesis. Mol
Biosyst. 13:565–576. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bam M, Yang X, Sen S, Zumbrun EE, Dennis
L, Zhang J, Nagarkatti PS and Nagarkatti M: Characterization of
dysregulated miRNA in peripheral blood mononuclear cells from
ischemic stroke patients. Mol Neurobiol. 55:1419–1429. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ma Q, Zhao H, Tao Z, Wang R, Liu P, Han Z,
Ma S, Luo Y and Jia J: MicroRNA-181c exacerbates brain injury in
acute ischemic stroke. Aging Dis. 7:705–714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Watson SJ and Akil H: Gene chips and
arrays revealed: A primer on their power and their uses. Biol
Psychiatry. 45:533–543. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Johnston M: Gene chips: Array of hope for
understanding gene regulation. Curr Biol. 8:R171–R174. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hochreiter S, Clevert DA and Obermayer K:
A new summarization method for Affymetrix probe level data.
Bioinformatics. 22:943–949. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Teng L, Wang K, Liu Y, Ma Y, Chen W and Bi
L: Based on integrated bioinformatics analysis identification of
biomarkers in hepatocellular carcinoma patients from different
regions. Biomed Res Int. 2019:17423412019. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Gelaw YA, Williams G, Assefa Y, Asressie M
and Soares Magalhães RJ: Sociodemographic profiling of tuberculosis
hotspots in Ethiopia, 2014–2017. Trans R Soc Trop Med Hyg.
113:379–391. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Thodberg M, Thieffry A, Vitting-Seerup K,
Andersson R and Sandelin A: CAGEfightR: Analysis of 5′-end data
using R/Bioconductor. 20:4872019.PubMed/NCBI
|
|
28
|
Postma M and Goedhart J: PlotsOfData-A web
app for visualizing data together with their summaries. PLoS Biol.
17:e30002022019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Doncheva NT, Morris JH, Gorodkin J and
Jensen LJ: Cytoscape StringApp: Network analysis and visualization
of proteomics data. J Proteome Res. 18:623–632. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scardoni G, Tosadori G, Faizan M, Spoto F,
Fabbri F and Laudanna C: Biological network analysis with
CentiScaPe: Centralities and experimental dataset integration.
F1000Res. 3:1392014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kalhori MR, Arefian E, Fallah Atanaki F,
Kavousi K and Soleimani M: miR-548× and miR-4698 controlled cell
proliferation by affecting the PI3K/AKT signaling pathway in
Glioblastoma cell lines. Sci Rep. 10:15582020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Tang D, Zhao X, Zhang L and Wang C:
Comprehensive analysis of pseudogene HSPB1P1 and its potential
roles in hepatocellular carcinoma. J Cell Physiol. Jan
27–2020.(Epub ahead of print). View Article : Google Scholar
|
|
33
|
Jiang W, Zheng L, Yan Q, Chen L and Wang
X: miR-532-3p inhibits metastasis and proliferation of non-small
cell lung cancer by targeting FOXP3. J BUON. 24:2287–2293.
2019.PubMed/NCBI
|
|
34
|
Sticht C, De La Torre C, Parveen A and
Gretz N: miRWalk: An online resource for prediction of microRNA
binding sites. PLoS One. 13:e02062392018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Monbailliu T, Goossens J and
Hachimi-Idrissi S: Blood protein biomarkers as diagnostic tool for
ischemic stroke: A systematic review. Biomark Med. 11:503–512.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Li P, Teng F, Gao F, Zhang M, Wu J and
Zhang C: Identification of circulating microRNAs as potential
biomarkers for detecting acute ischemic stroke. Cell Mol Neurobiol.
35:433–447. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dolmans LS, Rutten FH, Koenen NCT,
Bartelink MEL, Reitsma JB, Kappelle LJ and Hoes AW: Candidate
biomarkers for the diagnosis of transient ischemic attack: A
systematic review. Cerebrovasc Dis. 47:207–216. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
McColl BW, Rothwell NJ and Allan SM:
Systemic inflammatory stimulus potentiates the acute phase and CXC
chemokine responses to experimental stroke and exacerbates brain
damage via interleukin-1- and neutrophil-dependent mechanisms. J
Neurosci. 27:4403–4412. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Miller MC and Mayo KH: Chemokines from a
structural perspective. Int J Mol Sci. 18(pii): E20882017.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Mirabelli-Badenier M, Braunersreuther V,
Viviani GL, Dallegri F, Quercioli A, Veneselli E, Mach F and
Montecucco F: CC and CXC chemokines are pivotal mediators of
cerebral injury in ischaemic stroke. Thromb Haemost. 105:409–420.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Griffith JW, Sokol CL and Luster AD:
Chemokines and chemokine receptors: Positioning cells for host
defense and immunity. Annu Rev Immunol. 32:659–702. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Brait VH, Rivera J, Broughton BR, Lee S,
Drummond GR and Sobey CG: Chemokine-related gene expression in the
brain following ischemic stroke: No role for CXCR2 in outcome.
Brain Res. 1372:169–179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Villa P, Triulzi S, Cavalieri B, Di
Bitondo R, Bertini R, Barbera S, Bigini P, Mennini T, Gelosa P,
Tremoli E, et al: The interleukin-8 (IL-8/CXCL8) receptor inhibitor
reparixin improves neurological deficits and reduces long-term
inflammation in permanent and transient cerebral ischemia in rats.
Mol Med. 13:125–133. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Harris RA, Bowker-Kinley MM, Huang B and
Wu PJ: Regulation of the activity of the pyruvate dehydrogenase
complex. Adv Enzyme Regul. 42:249–259. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Holness MJ and Sugden MC: Regulation of
pyruvate dehydrogenase complex activity by reversible
phosphorylation. Biochem Soc Trans. 31:1143–1151. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Cohoon KP, Criqui MH, Budoff MJ, Lima JA,
Blaha MJ, Decker PA, Durazo R, Liu K and Kramer H: Relationship of
aortic wall distensibility to mitral and aortic valve
calcification: The multi-ethnic study of atherosclerosis.
Angiology. 69:443–448. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang R, Zhu Y, Wang Y, Ma W, Han X, Wang X
and Liu N: HIF-1α/PDK4/autophagy pathway protects against advanced
glycation end-products induced vascular smooth muscle cell
calcification. Biochem Biophys Res Commun. 517:470–476. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Sugden MC and Holness MJ: Recent advances
in mechanisms regulating glucose oxidation at the level of the
pyruvate dehydrogenase complex by PDKs. Am J Physiol Endocrinol
Metab. 284:E855–E862. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Tobin MK, Bonds JA, Minshall RD,
Pelligrino DA, Testai FD and Lazarov O: Neurogenesis and
inflammation after ischemic stroke: What is known and where we go
from here. J Cereb Blood Flow Metab. 34:1573–1584. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Cao Q, Wu J, Wang X and Song C: Noncoding
RNAs in vascular aging. Oxid Med Cell Longev. 2020:79149572020.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Gupta MK, Halley C, Duan ZH, Lappe J,
Viterna J, Jana S, Augoff K, Mohan ML, Vasudevan NT, Na J, et al:
miRNA-548c: A specific signature in circulating PBMCs from dilated
cardiomyopathy patients. J Mol Cell Cardiol. 62:131–141. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Cho EY, Lee SJ, Nam KW, Shin J, Oh KB, Kim
KH and Mar W: Amelioration of oxygen and glucose
deprivation-induced neuronal death by chloroform fraction of bay
leaves (Laurus nobilis). Biosci Biotechnol Biochem. 74:2029–2035.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
He W, Chen S, Chen X, Li S and Chen W:
Bioinformatic analysis of potential microRNAs in ischemic stroke. J
Stroke Cerebrovasc Dis. 25:1753–1759. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sørensen SS, Nygaard AB, Nielsen MY,
Jensen K and Christensen T: miRNA expression profiles in
cerebrospinal fluid and blood of patients with acute ischemic
stroke. Transl Stroke Res. 5:711–718. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Huang D, Chen Z, Wang J, Chen Y, Liu D and
Lin K: MicroRNA-221 is a potential biomarker of myocardial
hypertrophy and fibrosis in hypertrophic obstructive
cardiomyopathy. Biosci Rep. 40(pii): BSR201912342020. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Di Martino MT, Arbitrio M, Fonsi M,
Erratico CA, Scionti F, Caracciolo D, Tagliaferri P and Tassone P:
Allometric scaling approaches for predicting human pharmacokinetic
of a locked nucleic acid oligonucleotide targeting
cancer-associated miR-221. Cancers (Basel). 12(pii): E272019.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wei YS, Xiang Y, Liao PH, Wang JL and Peng
YF: An rs4705342 T>C polymorphism in the promoter of miR-143/145
is associated with a decreased risk of ischemic stroke. Sci Rep.
6:346202016. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Liang Y, Xu J, Wang Y, Tang JY, Yang SL,
Xiang HG, Wu SX and Li XJ: Inhibition of miRNA-125b decreases
cerebral ischemia/reperfusion injury by targeting CK2α/NADPH
oxidase signaling. Cell Physiol Biochem. 45:1818–1826. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Li JQ, Hu SY, Wang ZY, Lin J, Jian S, Dong
YC, Wu XF, Lan D and Cao LJ: MicroRNA-125-5p targeted CXCL13: A
potential biomarker associated with immune thrombocytopenia. Am J
Transl Res. 7:772–780. 2015.PubMed/NCBI
|
|
60
|
Chen T, Huang Z, Wang L, Wang Y, Wu F,
Meng S and Wang C: MicroRNA-125a-5p partly regulates the
inflammatory response, lipid uptake, and ORP9 expression in
oxLDL-stimulated monocyte/macrophages. Cardiovasc Res. 83:131–139.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Shi C, Na N, Zhu X and Xu J: Estrogenic
effect of ginsenoside Rg1 on APP processing in post-menopausal
platelets. Platelets. 24:51–62. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Giustino G, Overbey J, Taylor D, Ailawadi
G, Kirkwood K, DeRose J, Gillinov MA, Dagenais F, Mayer ML,
Moskowitz A, et al: Sex-based differences in outcomes after mitral
valve surgery for severe ischemic mitral regurgitation: From the
cardiothoracic surgical trials network. JACC Heart Fail. 7:481–490.
2019. View Article : Google Scholar : PubMed/NCBI
|