Introduction
Allergic asthma is one of the most common chronic
inflammatory disorders characterized by airway hyperresponsiveness
and increased infiltration of inflammatory cells, especially
eosinophils, into the airways and lungs (1). The T helper cell (Th)2 immune
response and associated cytokines, such as IL-4, IL-5 and IL-13,
are known to play important roles in the pathogenesis of asthma
(2).
A total of seven sirtuin (SIRT) family members have
been identified in mammals, termed SIRT1 to SIRT7 (3). SIRT1 and SIRT2 are found in the
nucleus and cytoplasm, whereas SIRT3, SIRT4 and SIRT5 are
mitochondrial, and SIRT6 and SIRT7 are exclusively nuclear
(4). SIRT1 is a
NAD+-dependent class III histone deacetylase protein,
which performs a wide variety of functions in biological systems,
with implications in metabolic diseases, cancer and inflammation
(5,6). SIRT1 regulates various cellular
processes, including inflammation, chromatin stability and
oxidative stress, by deacetylating nuclear factor-κB, forkhead box
protein O3 and hypoxia-inducible factor 1α (7). Multiple studies have previously
suggested that SIRT1 may be involved in the pathogenesis of
allergic airway inflammatory diseases, such as asthma (8,9). It
was previously reported that SIRT1 inhibition significantly reduced
airway hyperresponsiveness, inflammatory cell infiltration into the
airways, and increased levels of IL-4, IL-5 and IL-13 after OVA
inhalation (8). Furthermore, serum
SIRT1 levels were revealed to be significantly elevated in patients
with asthma and OVA-induced allergic mice, and were positively
correlated with total serum IgE levels and were negatively
correlated with pulmonary function (9). However, the roles of SIRT1 in asthma
remain largely unclear.
Rapamycin represents a macrolide with known
immunosuppressive functions (10).
While rapamycin affects multiple nutrient signaling-associated
pathways, it acts mainly as a mammalian target of rapamycin (mTOR)
inhibitor, by directly suppressing mTOR complex 1 (11). mTOR, an evolutionarily conserved
serine/threonine kinase, is a central regulator of cell
transcription, growth, proliferation and survival (12). Previous studies have reported that
mTOR signaling is involved in inflammatory diseases, including
dermatological ailments (13),
chronic obstructive pulmonary disease (14,15)
and asthma (16). Furthermore, it
was revealed that autophagy may be involved in the regulation of
asthma (17,18), and enhanced autophagy was reported
to be associated with increased asthma severity (18). Autophagy is controlled by the mTOR
signaling pathway (19). In
allergic mice, the lung levels of phosphorylated (p)-mTOR were
decreased, whereas mTOR signaling activation inhibited allergic
airway inflammation by suppressing autophagy (16). Additionally, the mTOR and SIRT1
signaling pathways are closely integrated and play an important
role in the neuroprotective effect of caloric restriction (20). A previous study indicated that
SIRT1 induced the growth and survival of neurons in the central
nervous system though negative modulation of mTOR signaling
(21). Therefore, it is possible
that the mTOR and SIRT1 signaling pathways interact to modulate
autophagy in asthmatic mice.
It has been reported that serum SIRT1 levels are
increased in patients with asthma and OVA-induced allergic mice,
whereas SIRT1 inhibition ameliorates airway inflammation in
allergic mice. However, the mechanism underlying allergic airway
inflammation suppression is unclear. EX-527 is a well-known
specific inhibitor of SIRT1 (22,23),
selective inhibition of SIRT1 by EX-527 could alleviate
endotoxemia-associated acute lung injury (22). Therefore, the aim of this study was
to investigate the effect and mechanism of EX-527 on allergic
airway inflammation in a murine model of asthma.
Materials and methods
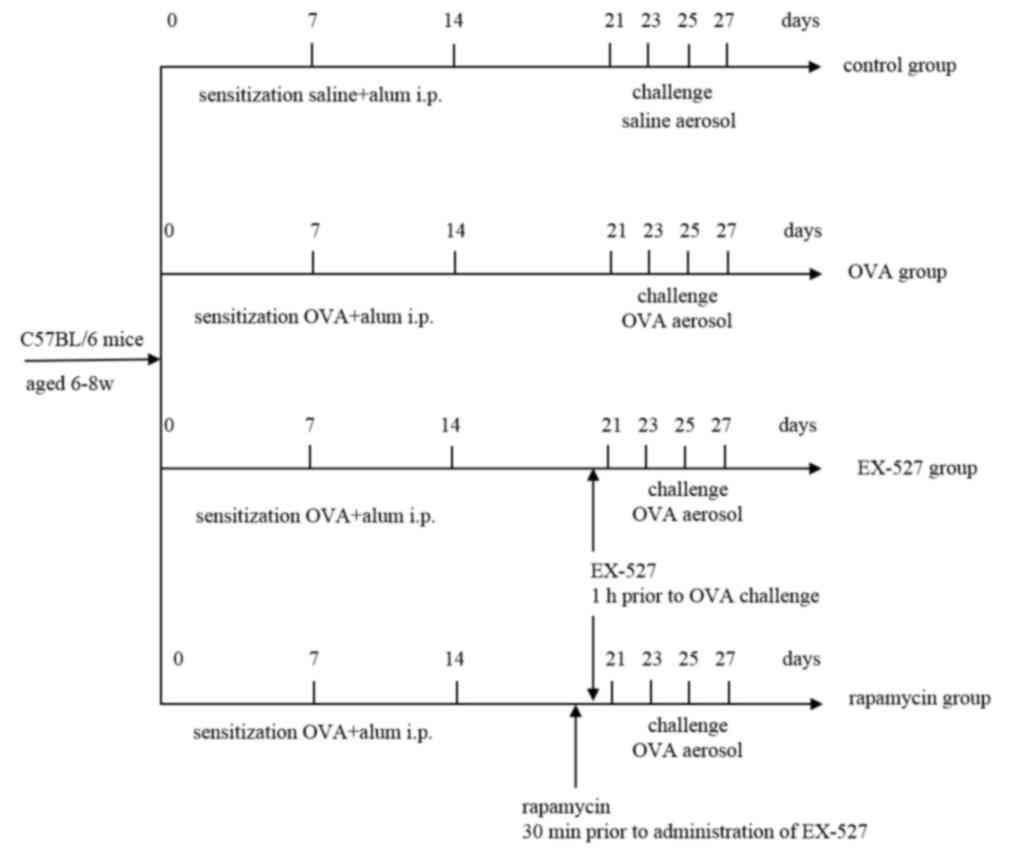
Animals and experimental design
A total of 32 female C57BL/6 mice (age, 6–8 weeks;
weight, 20–22 g) were purchased from the Experimental Animal Center
Medical College of Xi'an Jiaotong University (Xi'an, China). All
animals were maintained in steel cages in a room with controlled
temperature at 23–25°C, under a 12-h light/dark cycle in a specific
pathogen-free facility, with access to food and water ad
libitum. All experimental protocols were approved by the
medical ethics committee of The Second Affiliated Hospital of Xi'an
Jiaotong University. Mice were maintained in an animal facility
under standard laboratory conditions for 1 week prior to
experiments, and were randomly assigned into four groups
(n=8/group): i) Control, ii) OVA, iii) EX-527 and iv) rapamycin
groups. The OVA group was sensitized by subcutaneous injection of
OVA (50 µg/kg) adsorbed to 2 mg aluminum hydroxide in 200 µl normal
saline at 0, 7 and 14 days, then challenged by exposure to an
aerosol containing 1% OVA (w/v) in PBS for 30 min using an
ultrasonic nebulizer (Soniclizer305, ATOM) on days 21, 23, 25 and
27 after the initial sensitization. In the EX-527 group, mice
received EX-527 administration (10 mg/kg) by intraperitoneal
injection 1 h prior to OVA inhalation. The control group mice were
sensitized to PBS and challenged with PBS aerosols.
To further define the role of mTOR activation in the
effect of EX-527 on allergic airway inflammation, asthmatic mice
were intraperitoneally treated with the mTOR inhibitor rapamycin (4
mg/kg) 30 min prior to EX-527 administration. A total of 24 h after
the last OVA challenge, the mice were anesthetized with an
intraperitoneal injection of 1% pentobarbital sodium (50 mg/kg;
Sigma-Aldrich; Merck KGaA) followed by cervical dislocation. Mice
were considered dead when respiratory arrest and cardiac arrest
were observed, their nerve reflexes disappeared, and muscles
relaxed. Mice health and behavior were monitored every day for 28
days, and none of the rats became severely ill or moribund.
Fig. 1 shows the treatment regimen
through the course of the experiment. BALF was collected for
determining cytokine levels and cell counts. The left lungs were
preserved and fixed with 10% formalin for 24 h at room temperature
and then stained with hematoxylin for 5 min and eosin for 3 min at
room temperature for histopathological analysis under a light
microscope (Olympus Corporation). The right lungs were collected
and frozen at −80°C for further experiments.
Reagents
OVA and rapamycin were purchased from Sigma-Aldrich
(Merck KGaA). ELISA kits for detecting IL-4 [cat. no.
KGEHC006(H)-1], IL-13 (cat. no. KGEMC124-1) and interferon (IFN)-γ
(cat. no. KGEBC101g-1) were purchased from Yufeng Technology Co.,
Ltd. Rabbit anti-mouse SIRT1 (1:1,000; cat. no. ab110304),
microtubule-associated protein 1 light chain 3β (LC3B; 1:1,000;
cat. no. ab243506) and Beclin-1(1:1,000; cat. no. ab114071)
antibodies, and EX-527 (cat. no. ab141506) were purchased from
Abcam. Antibodies against total (t)-mTOR (1:1,000; cat. no. 2972)
and p-mTOR (1:1,000; cat. no. 5536) were purchased from Cell
Signaling Technology, Inc. Anti-β-actin antibodies (1:5,000; cat.
no. AM33096PU-S) were from OriGene Technologies, Inc.
Histological analysis
The left lobe of the lung was fixed in formalin and
paraffin embedded. The sections were stained with hematoxylin and
eosin for histopathological evaluation under a light microscope
(Olympus Corporation).
Collection of BALF
A tracheal tube was inserted for a 1-ml ice-cold PBS
lavage, and the obtained BALF was centrifuged at 500 × g at 4°C for
5 min. Next, the supernatant was collected and stored at −80°C for
subsequent cytokine measurements. The levels of IL-4, IL-13 and
IFN-γ were determined by ELISA kits, according to the
manufacturer's instructions. The cell pellets were obtained and
resuspended in PBS for total cell and eosinophil counts. The slides
were fixed and stained with Diff-Quik Stain (cat. no. G1541;
Beijing Solarbio Science & Technology Co., Ltd.) according to
the manufacturer's protocol, and differential cell counts were
obtained using light microscopic evaluation of 300 cells/slide.
Total BALF cells were counted on a haemocytometer.
Western blot analysis
Lung tissue samples were lysed in RIPA Lysis Buffer
(Guangzhou Fansi Biotechnology Co., Ltd.). Lysates were centrifuged
at 6500 × g at 4°C for 15 min, and supernatant was collected and
quantified with a BCA protein assay kit (Pierce; Thermo Fisher
Scientific, Inc.). 40 µg of proteins were separated by 10% SDS-PAGE
and transferred onto a PVDF membrane (cat. no. IPVH00005; EMD
Millipore) and blocked with 5% BSA (Beijing Solarbio Science &
Technology Co., Ltd.) for 1 h at room temperature. The membranes
were probed with polyclonal antibodies against SIRT1, LC3B,
Beclin-1, t-mTOR, p-mTOR, and a monoclonal antibody against β-actin
in Tris-buffered saline containing Tween-20 (TBST; Beijing Solarbio
Science & Technology Co., Ltd.) for 2 h at room temperature.
then incubated with goat anti-rabbit secondary antibody in TBST for
1 h at room temperature. Reactions were developed with SuperSignal
West Pico Chemiluminescent Substrate (Pierce; Thermo Fisher
Scientific, Inc.), followed by exposure to autoradiographic films.
Signals were semi-quantified from scanned films using Quantity One
software (v4.6.6, Bio-Rad Laboratories, Inc.).
Statistical analysis
All data were analyzed with SPSS 19.0 software (IBM
Corp.). Values are presented as the mean ± SD (n=8 mice/group).
Data were assessed by one-way ANOVA followed by Tukey's post hoc
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
EX-527 alleviates OVA-induced mice
airway inflammation, and rapamycin reverses this effect
BALF samples were collected 24 h after the final OVA
challenge. In order to determine the effects of EX-527 on airway
inflammation, total cell and eosinophil counts in BALF samples from
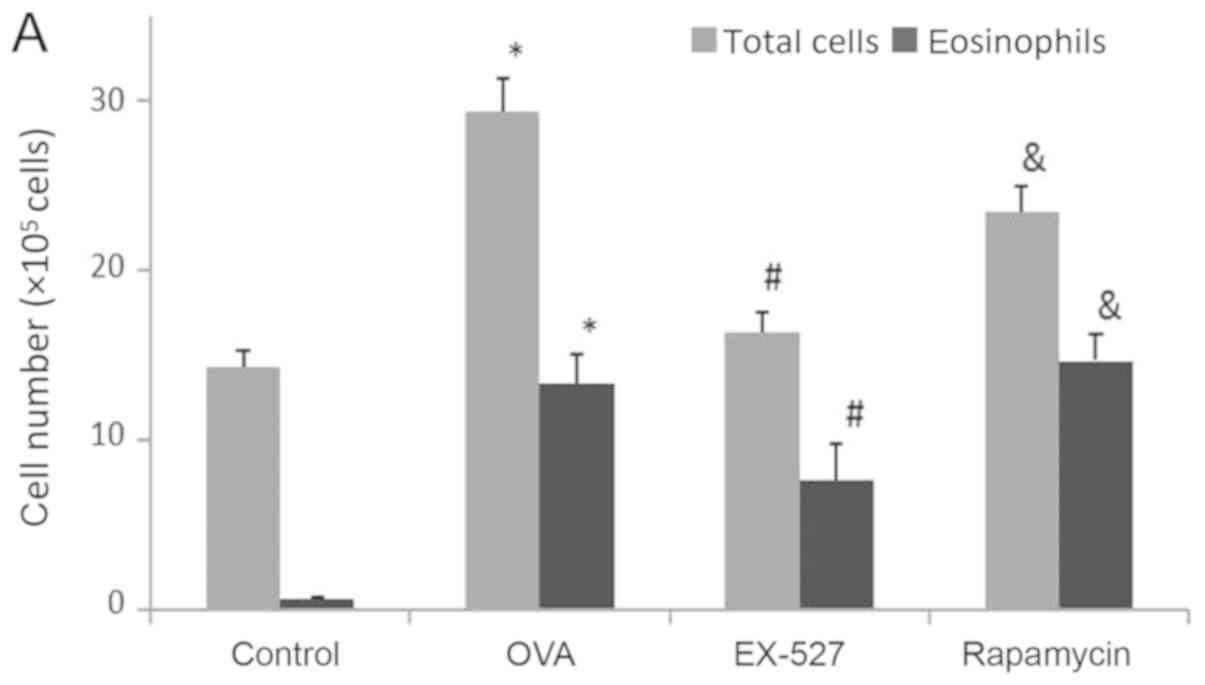
OVA-induced asthmatic mice were examined. As shown in Fig. 2A, OVA administration significantly
increased total cell counts in the BALF compared with the control
group (P<0.05). After EX-527 administration, total cell counts
were significantly reduced compared with the OVA group (P<0.05;
Fig. 2A). BALF eosinophil levels
were consistent with total cell counts. Specifically, OVA treatment
resulted in markedly increased eosinophil levels in BALF samples in
comparison with the control group (P<0.05; Fig. 2A). Meanwhile, the EX-527 group
exhibited decreased eosinophil levels compared with the OVA
treatment group (P<0.05; Fig.
2A). In order to evaluate the role of mTOR in the
EX-527-mediated protective effect on asthma, OVA-induced allergic
mice were treated with EX-527 in combination with rapamycin. As
shown in Fig. 2A, rapamycin
restored the decreased amounts of total inflammatory cells and
eosinophils induced by EX-527 (P<0.05).
A Th1/Th2 imbalance is believed to play a vital role
in asthma pathogenesis, including changes in the levels of Th1
cytokines (IFN-γ) and Th2 cytokines (IL-4 and IL-13) (2). To determine the effects of EX-527 on
cytokine release in OVA-induced asthmatic mice, BALF IL-4, IL-13
and IFN-γ levels normalized to total protein amounts in BALF were
evaluated by ELISA 24 h after the final OVA challenge. Asthmatic
mice exhibited marked increases in IL-4 (0.77±0.03 vs. 0.12±0.03)
and IL-13 (0.86±0.06 vs. 0.23±0.02) levels, and a significant
decrease in IFN-γ (0.11±0.01 vs. 0.27±0.03) levels (P<0.01;
Fig. 2B-D). Treatment with EX-527
resulted in markedly reduced IL-4 (0.28±0.03) and IL-13 (0.44±0.01)
levels, and significantly increased IFN-γ levels (0.20±0.02) in
BALF samples in comparison with the OVA group (P<0.05; Fig. 2B-D). Consistent with the
aforementioned results, rapamycin treatment partially reversed the
reductions in IL-4 (P<0.05; Fig.
2B) and IL-13 (P<0.05; Fig.
2C) levels, and the increase in IFN-γ levels (P<0.05;
Fig. 2D).
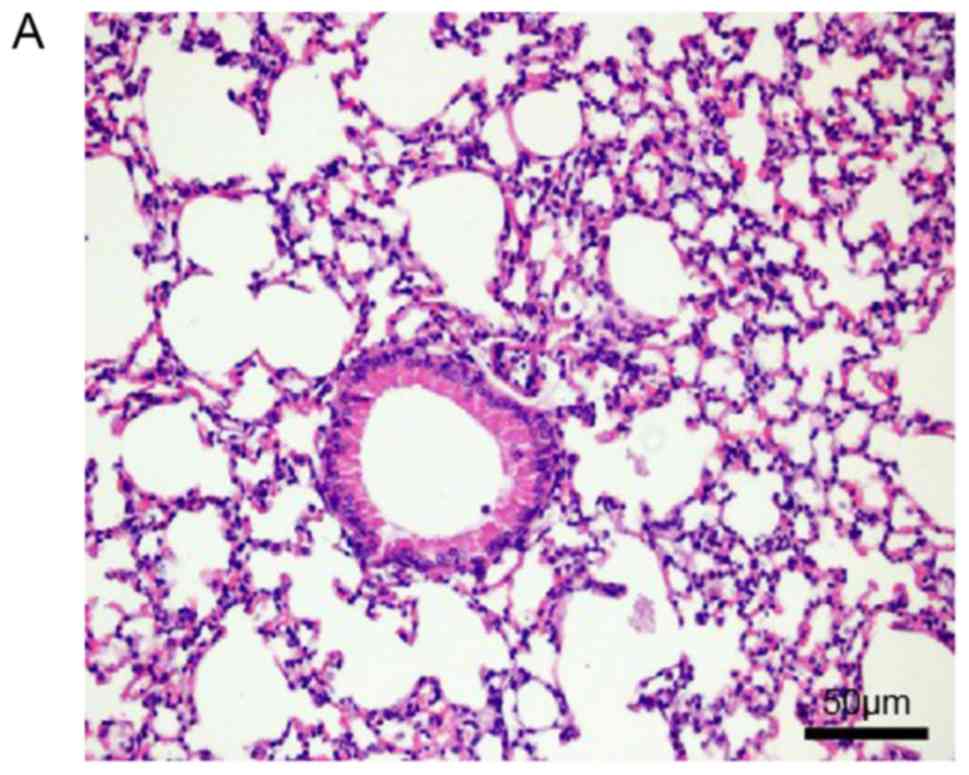
Histopathological examination of mouse lungs was
performed in order to confirm the inhibitory effect of EX-527 on
airway inflammation. The extracted lung tissues exhibited marked
infiltration of inflammatory cells in perivascular and
peribronchiolar connective tissue samples from the OVA group
compared with control mice, and most leukocytes were eosinophils
(Fig. 3A and B). Meanwhile, mice
treated with EX-527 prior to OVA challenge exhibited markedly
decreased inflammatory cell infiltration around the airways and
blood vessels (Fig. 3C). As
expected, co-administration of rapamycin partly abolished the
anti-inflammatory effects of EX-527 (Fig. 3D). These findings indicated that
the beneficial effects of EX-527 on OVA-induced allergic mice may
be inhibited by rapamycin.
Anti-inflammatory effects of EX-527
are associated with suppressed autophagy
Because EX-527 is considered a selective SIRT1
inhibitor, the study next assessed whether its beneficial effects
are associated with SIRT1 inhibition. Western blot analysis
indicated that OVA exposure resulted in increased SIRT1 expression,
which was significantly reversed by treatment with EX-527 (Fig. 4A). These data suggested that EX-527
suppressed airway inflammation in asthmatic mice by inhibiting
SIRT1, since EX-527 is a well-known specific inhibitor of SIRT1-1
(22). LC3B, a mammalian homolog
of yeast Atg8, is comprised of two forms: LC3-I (18 kDa) and LC3-II
(16 kDa). During autophagy the cytoplasmic form LC3-I is processed
and recruited to autophagosomes, where LC3-II is generated by
site-specific proteolysis near the C-terminus (24). The ratio of LC3-II to LC3-I is
commonly used to analyze autophagic activity, of which Beclin-1 is
also considered a hallmark (17).
Furthermore, it has been reported that autophagy suppression may
inhibit airway inflammation in allergic mice (16). Therefore, the present study
assessed whether EX-527 exerted anti-inflammatory effects by
suppressing autophagy. In this study, the LC3-II/LC3-I ratio was
significantly increased in mice in the OVA group compared with the
control group (P<0.01), and this upregulation was alleviated by
EX-527 administration (P<0.05; Fig.
4B). Similarly, Beclin-1 expression was increased by OVA and
decreased by EX-527 (Fig. 4C).
Thus, the beneficial effects of EX-527 might be attributed to
LC3-II and Beclin-1 suppression. These findings indicated that
autophagy was upregulated in OVA-induced allergic asthmatic mice
but alleviated by SIRT1 inhibition.
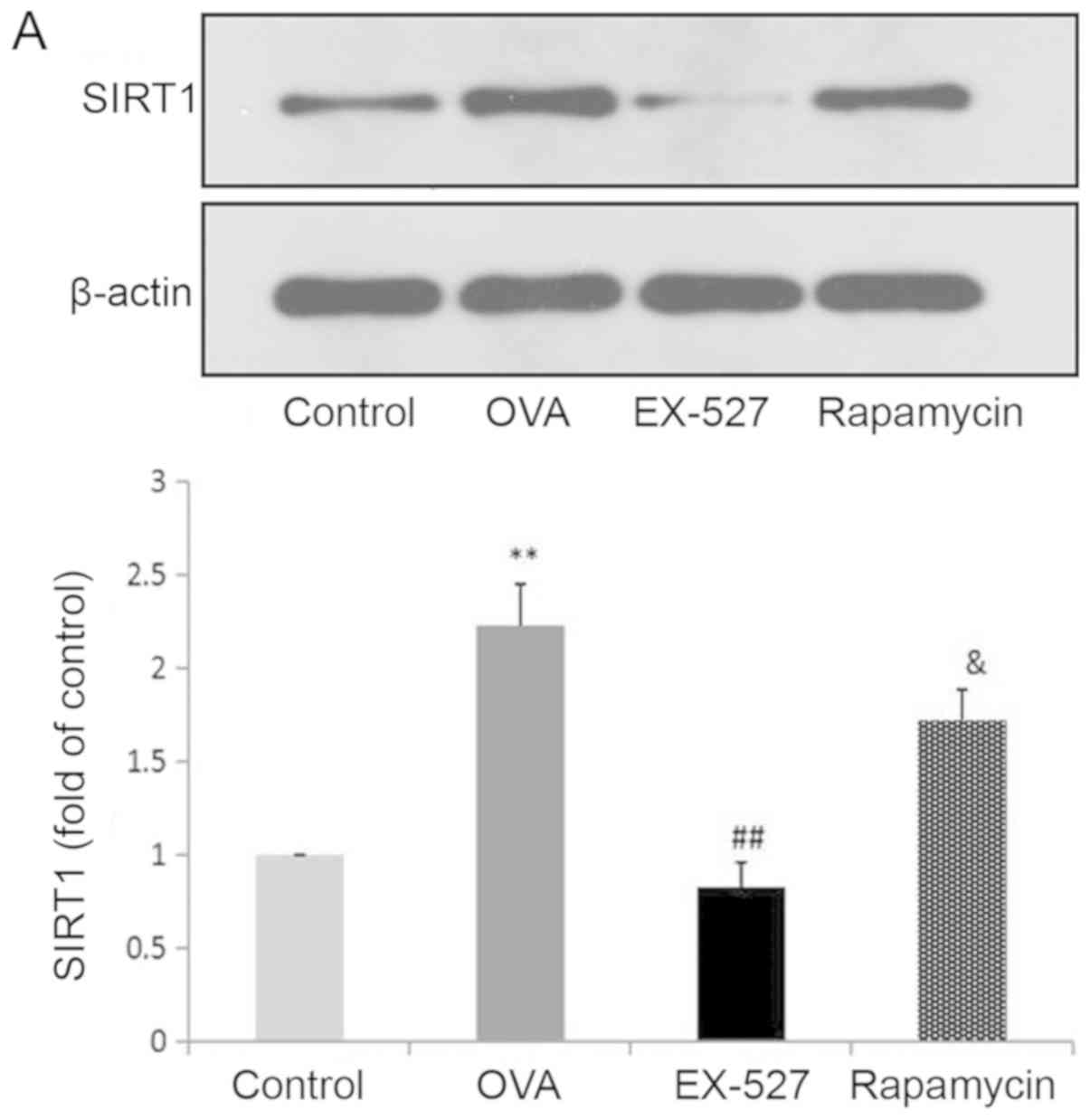 | Figure 4.EX-527 suppresses OVA-induced
autophagy, and this effect is abolished by rapamycin. Western blot
analysis of (A) SIRT1 and (B) LC3-II/LC3-I. EX-527 suppresses
OVA-induced autophagy, and this effect is abolished by rapamycin.
Western blot analysis of (C) Beclin-1 and (D) p-mTOR/t-mTOR.
β-actin was used as a loading control. Control, saline treatment;
OVA, OVA sensitization/challenge; EX-527, EX-527 (10 mg/kg) + OVA
sensitization/challenge; rapamycin, rapamycin (4 mg/kg) + EX-527
(10 mg/kg) + OVA sensitization/challenge. **P<0.01 vs. control
group; #P<0.05 and ##P<0.01 vs. OVA
group; &P<0.05 vs. EX-527 group. OVA, ovalbumin;
SIRT1, sirtuin 1; LC3, microtubule-associated protein 1 light chain
3β; mTOR, mammalian target of rapamycin; p, phosphorylated; t,
total. |
SIRT1 regulates autophagy through the
mTOR signaling pathway
mTOR activation was induced after treatment with
EX-527 in OVA-stimulated mice. To investigate whether mTOR-mediated
autophagy was involved in the protective effect of EX-527 on
OVA-induced allergic airway inflammation, the OVA group was
administered rapamycin 30 min prior to treatment with EX-527.
Subsequently, SIRT1, LC3-II/LC3-I, Beclin-1 and p-mTOR expression
levels were determined by western blotting. Notably, rapamycin
significantly reversed the inhibited autophagic flux induced by
EX-527, as reflected by increased SIRT1 (Fig. 4A; P<0.05), LC3-II/LC3-I
(Fig. 4B; P<0.05) and Beclin-1
expression (Fig. 4C; P<0.05),
and decreased levels of p-mTOR (Fig.
4D; P<0.05). These results suggested that the inhibitory
effect EX-527 on autophagy was significantly suppressed by
rapamycin, which indicated that EX-527 inhibited autophagy through
mTOR signaling suppression.
Discussion
In the present study, SIRT1 inhibition by EX-527
effectively alleviated airway inflammation in OVA-induced asthmatic
mice through suppression of autophagic flux; this effect was
mediated by mTOR pathway activation. It is known that loss of
balance between Th1 and Th2 plays a critical role in asthma
pathophysiology (25). A Th1/Th2
imbalance can be triggered by changes in the levels of IFN-γ and
IL-4 secreted by Th1 and Th2 cells, respectively (26). In the OVA-induced mouse model of
asthma, allergen exposure has been reported to produce
eosinophilia, airway hyperresponsiveness, mucus hypersecretion and
a Th2-dominated response, such as increased IL-4 and IL-13 in
plasma and BALF (27). Previous
findings indicated that suppression of SIRT1 prevented asthma
progression, alleviated airway hyperresponsiveness, decreased
inflammatory cell numbers around the airways, and decreased IL-4,
IL-5 and IL-13 levels in BALF in OVA-inhaled mice (8). The present results indicated that
compared with OVA-induced allergic mice, EX-527-treated mice had
significantly fewer total inflammatory cells and eosinophils, with
decreased levels of the Th2 cytokines IL-4 and IL-13, but increased
IFN-γ in BALF. Administration of rapamycin to
OVA-sensitized/challenged mice significantly reversed the
anti-inflammatory effects of EX-527.
It was previously reported that increased expression
of SIRT1 induced autophagy, and SIRT1 activation could enhance
autophagy, while its inhibition suppressed autophagy in vascular
adventitial fibroblasts (28).
Another study revealed that SIRT1 protected cardiomyocytes from
hypoxic stress by promoting autophagic flux induced by AMPK
activation (29). SIRT1 has also
been reported to be involved in the pathogenesis of OVA-induced
asthma in mice, and airway inflammation and hyperresponsiveness
were attenuated after its inhibition (8). In addition, autophagy in smooth
muscle cells was enhanced in OVA-induced allergic mice and reducing
cell autophagy significantly attenuated OVA-induced airway
inflammation (17). To further
determine whether SIRT1 inhibition suppressed inflammation in
allergic asthma by autophagy suppression, mice were
intraperitoneally injected with EX-527 before challenge with OVA.
It was revealed that LC3-II/LC3-I and Beclin-1 expression levels
were increased in the OVA group but were significantly reduced
after EX-527 administration.
mTOR is an evolutionarily conserved serine/threonine
protein kinase, which acts as a central regulator of cell
proliferation, cell growth, survival, autophagy and transcription
(30,31). mTOR activation has been shown to
reduce inflammation in an OVA-induced allergic mouse model
(16) and autophagy is known to be
controlled by the mTOR signaling pathway (19). Furthermore, mTOR levels were
decreased in the lungs of allergic mice, whereas mTOR activation
inhibited allergic airway inflammation via suppressed autophagy
(16). Therefore, mTOR could
represent a pivotal modulator of asthma development. Meanwhile, the
mTOR and SIRT1 signaling pathways have been shown to be closely
integrated (20). SIRT1 has been
reported to ameliorate systemic sclerosis via inhibition of mTOR
phosphorylation (32). It was
subsequently assessed whether mTOR may be involved in the
anti-inflammatory effects of EX-527 observed in OVA-induced
asthmatic mice. The present results indicated that asthmatic mice
treated with EX-527 exhibited relieved OVA-induced mTOR inhibition
and autophagy enhancement. Co-administration of EX-527 with
rapamycin reversed the suppression of autophagy, and partly
abolished the anti-inflammatory effects of EX-527. In addition,
significantly increased p-mTOR expression, and decreased
LC3-II/LC3-I and Beclin-1 levels after treatment with EX-527 were
reversed by treatment with rapamycin. These findings suggested that
the inhibitory effects of EX-527 on autophagy may be significantly
suppressed by rapamycin and might be achieved by inhibiting mTOR
activation.
The present study revealed that EX-527
administration in asthmatic mice downregulated the expression of
Th2-related cytokines IL-4 and IL-13 and upregulated the Th1-type
cytokine IFN-γ. EX-527 significantly reduced leukocyte and
eosinophil infiltration into the airways and lung tissue of
OVA-induced asthmatic mice. Similarly, LC3-II/LC3-I and Beclin-1
expression were decreased after EX-527 administration. Further
evidence indicated that rapamycin reversed the inhibitory effects
of EX-527 on airway inflammation and autophagy influx in
OVA-induced asthmatic mice.
Taken together, these findings indicated that EX-527
alleviated airway inflammation in asthmatic mice, likely through
modulation of the mTOR pathway. Therefore, mTOR signaling could be
targeted for asthma control. Further related studies are
warranted.
As a notable limitation of the present study, there
wasn't an EX-527-only group. Furthermore, transmission electron
microscopy could be applied to assess autophagy in future studies.
Finally, the present study was performed in a mouse model, which
may not completely recapitulate the clinical situation. Therefore,
clinical trials are also warranted to determine the translational
potential of the present findings.
In conclusion, the present study demonstrated that
EX-527 effectively inhibited allergen-induced airway inflammation
in an asthma model via mTOR-mediated autophagy.
Acknowledgements
Not applicable.
Funding
The present study was supported by from the National
Natural Science Foundation of China (grant no. 81800030), the
Natural Science Foundation of Shanxi Province (grant no.
2018JQ8034) and the Xi'an science and technology project (grant no.
2017113SF/ YX007).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XS and YL contributed to the conception and design
of the study and gave final approval of the final version to be
published. YW performed the majority of the study, data analysis
and drafted the manuscript. YH provided pathological assistance and
was involved in the data analysis. WL contributed to interpretation
of the data and analyses. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
All experimental protocols were approved by the
Medical Ethics Committee of The Second Affiliated Hospital of Xi'an
Jiaotong University (approval no. 2019043).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Tang L, Chen Q, Meng Z, Sun L, Zhu L, Liu
J, Hu J, Ni Z and Wang X: suppression of sirtuin-1 increases IL-6
expression by activation of the Akt pathway during allergic asthma.
Cell Physiol Biochem. 43:1950–1960. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ding F, Fu Z and Liu B: Lipopolysaccharide
exposure alleviates asthma in mice by regulating Th1/Th2 and
Treg/Th17 balance. Med Sci Monit. 24:3220–3229. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Islam S, Abiko Y, Uehara O and Chiba I:
Sirtuin 1 and oral cancer. Oncol Lett. 17:729–738. 2019.PubMed/NCBI
|
|
4
|
Kida Y and Goligorsky MS: Sirtuins, cell
senescence, and vascular aging. Can J Cardiol. 32:634–641. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lin Z and Fang D: The roles of SIRT1 in
cancer. Genes Cancer. 4:97–104. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Rahman S and Islam R: Mammalian Sirt1:
Insights on its biological functions. Cell Commun Signal. 9:112011.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Potteti HR, Rajasekaran S, Rajamohan SB,
Tamatam CR, Reddy NM and Reddy SP: Sirtuin 1 promotes
hyperoxia-induced lung epithelial cell death independent of
NF-E2-related factor 2 activation. Am J Respir Cell Mol Biol.
54:697–706. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kim SR, Lee KS, Park SJ, Min KH, Choe YH,
Moon H, Yoo WH, Chae HJ, Han MK and Lee YC: Involvement of sirtuin
1 in airway inflammation and hyperresponsiveness of allergic airway
disease. J Allergy Clin Immunol. 125:449–460.e4. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang Y, Li D, Ma G, Li W, Wu J, Lai T,
Huang D, Zhao X, Lv Q, Chen M and Wu B: Increases in peripheral
SIRT1: A new biological characteristic of asthma. Respirology.
20:1066–1072. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu Y, Yang F, Zou S and Qu L: Rapamycin:
A bacteria-derived immunosuppressant that has anti-atherosclerotic
effects and its clinical application. Front Pharmacol. 9:15202019.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Saxton RA and Sabatini DM: mTOR signaling
in growth, metabolism, and disease. Cell. 168:960–976. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lin J, Huo X and Liu X: ‘mTOR signaling
pathway’: A potential target of curcumin in the treatment of spinal
cord injury. Biomed Res Int. 2017:16348012017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Salido-Vallejo R, Garnacho-Saucedo G and
Vélez A: Elucidation of the mTOR pathway and therapeutic
applications in dermatology. Actas Dermosifiliogr. 107:379–390.
2016.(In English, Spanish). View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Perl A: Activation of mTOR (mechanistic
target of rapamycin) in rheumatic diseases. Nat Rev Rheumatol.
12:169–182. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wang Y, Liu J, Zhou JS, Huang HQ, Li ZY,
Xu XC, Lai TW, Hu Y, Zhou HB, Chen HP, et al: MTOR suppresses
cigarette smoke-induced epithelial cell death and airway
inflammation in chronic obstructive pulmonary disease. J Immunol.
200:2571–2580. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zou H, Wang LX, Wang M, Cheng C, Li S,
Shen Q, Fang L and Liu R: MTOR-mediated autophagy is involved in
the protective effect of ketamine on allergic airway inflammation.
J Immunol Res. 2019:58797142019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cheng Z, Wang X, Dai L, Jia L, Jing X, Liu
Y, Wang H, Li P, An L and Liu M: Suppression of microRNA-384
enhances autophagy of airway smooth muscle cells in asthmatic
mouse. Oncotarget. 8:67933–67941. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Farooq MB and Walsh GM: Autophagy and
asthma. Clin Exp Allergy. 46:7–9. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Urbanska M, Gozdz A, Swiech LJ and
Jaworski J: Mammalian target of rapamycin complex 1 (mTORC1) and 2
(mTORC2) control the dendritic arbor morphology of hippocampal
neurons. J Biol Chem. 287:30240–30256. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ma L, Dong W, Wang R, Li Y, Xu B, Zhang J,
Zhao Z and Wang Y: Effect of caloric restriction on the SIRT1/mTOR
signaling pathways in senile mice. Brain Res Bull. 116:67–72. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Guo W, Qian L, Zhang J, Zhang W, Morrison
A, Hayes P, Wilson S, Chen T and Zhao J: Sirt1 overexpression in
neurons promotes neurite outgrowth and cell survival through
inhibition of the mTOR signaling. J Neurosci Res. 89:1723–1736.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Huang J, Tian R, Yang Y, Jiang R, Dai J,
Tang L and Zhang L: The SIRT1 inhibitor EX-527 suppresses mTOR
activation and alleviates acute lung injury in mice with
endotoxiemia. Innate Immun. 23:678–686. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhou X, Fan LX, Sweeney WE Jr, Denu JM,
Avner ED and Li X: Sirtuin 1 inhibition delays cyst formation in
autosomal-dominant polycystic kidney disease. J Clin Invest.
123:3084–3098. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kang R, Tang D, Lotze MT and Zeh Iii HJ:
Autophagy is required for IL-2-mediated fibroblast growth. Exp Cell
Res. 319:556–565. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Mushaben EM, Kramer EL, Brandt EB, Khurana
Hershey GK and Le Cras TD: Rapamycin attenuates airway
hyperreactivity, goblet cells, and IgE in experimental allergic
asthma. J Immunol. 187:5756–5763. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Shao YY, Zhou YM, Hu M, Li JZ, Chen CJ,
Wang YJ, Shi XY, Wang WJ and Zhang TT: The anti-allergic rhinitis
effect of traditional Chinese medicine of shenqi by regulating mast
cell degranulation and Th1/Th2 cytokine balance. Molecules.
22:E5042017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee M, Kim S, Kwon OK, Oh SR, Lee HK and
Ahn K: Anti-inflammatory and anti-asthmatic effects of resveratrol,
a polyphenolic stilbene, in a mouse model of allergic asthma. Int
Immunopharmacol. 9:418–424. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang WR, Li TT, Jing T, Li YX, Yang XF, He
YH, Zhang W, Lin R and Zhang JY: SIRT1 regulates the inflammatory
response of vascular adventitial fibroblasts through autophagy and
related signaling pathway. Cell Physiol Biochem. 41:569–582. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Luo G, Jian Z, Zhu Y, Zhu Y, Chen B, Ma R,
Tang F and Xiao Y: Sirt1 promotes autophagy and inhibits apoptosis
to protect cardiomyocytes from hypoxic stress. Int J Mol Med.
43:2033–2043. 2019.PubMed/NCBI
|
|
30
|
Tan FH, Bai Y, Saintigny P and Darido C:
mTOR signalling in head and neck cancer: Heads up. Cells.
8:E3332019. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rabanal-Ruiz Y, Otten EG and Korolchuk VI:
mTORC1 as the main gateway to autophagy. Essays Biochem.
61:565–584. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhu X, Chu H, Jiang S, Liu Q, Liu L, Xue
Y, Zheng S, Wan W, Qiu J, Wang J and Zou H: Sirt1 ameliorates
systemic sclerosis by targeting the mTOR pathway. J Dermatol Sci.
87:149–158. 2017. View Article : Google Scholar : PubMed/NCBI
|