Introduction
Non-alcoholic fatty liver disease (NAFLD) is caused
by metabolic stress and genetic susceptibility. NAFLD includes
simple steatosis and NA steatohepatitis (NASH) and may include
hepatocellular carcinoma (HCC) (1,2). The
global prevalence of NAFLD has been estimated to be 25% in recent
years (3). Therefore, NAFLD has
emerged as an important global public health issue, particularly in
the Chinese population (4,5). Increasing evidence has indicated that
NAFLD also contributes to a number of cardiovascular complications,
in addition to liver-associated complications (6,7). The
prevalence of coronary, cerebrovascular and peripheral vascular
disease is remarkably higher among patients with NAFLD than among
those in the general population (8). Currently, liver biopsy is the standard
method to diagnose NAFLD. Computed tomography scans and ultrasound
have also been used for the detection of NAFLD; however, these
methods have a number of disadvantages, including financial cost,
invasiveness and sampling variability (9). Therefore, a simpler, non-invasive
strategy for the evaluation of NAFLD is required.
Recently, microRNAs (miRNAs/miRs), a type of small
(19–25 nucleotides in length) non-coding RNA that regulates
post-transcription gene expression levels, have been considered as
a promising molecular marker for clinical application (10). In the liver, miRNAs are abundantly
expressed and regulate various functions, including cell death,
cell proliferation and developmental timing (11). Several studies have identified miRNA
profiles that are altered in NAFLD. Dysregulated circulating
miRNAs, including miR-27b, miR-122 and miR-21, were found to
differentiate patients with NAFLD from healthy controls (12–14).
However, miRNAs expressed by circulating dysfunctional cells can
affect circulating miRNAs (15).
Extracellular vesicles (EVs), membrane-derived
vesicles 100–1,000 nm in size, are secreted by numerous types of
cell, including tumor and hepatic cells (16). EVs contain nucleic acids and
proteins and are abundant in various types of bodily fluid, such as
blood, ejaculates, urine, cerebrospinal fluid, saliva and breast
milk (17). Accumulating evidence
(18) has demonstrated that miRNAs
in EVs in serum are more stable than serum circulating miRNAs, and
are protected from endogenous RNases degradation (19). This indicates that miRNAs derived
from serum EVs are preferable as diagnostic biological markers,
compared with serum circulating miRNAs. Numerous studies have
suggested the importance of EV-derived miRNAs in the diagnosis of
cancer (20–22); however, further investigation is
still required.
Our previous study identified dysregulated miRNA
expression levels in EVs secreted by hepatic cells in mice fed
high-fat diet (HFD) compared with mice fed chow diet (CD) using
microarray analysis (23). It is
still not known whether liver EV-derived miRNAs can be used to
discriminate between patients with NAFLD and healthy controls. In
the present study, the expression levels of four selected miRNAs
were investigated based on our previous study. The aim of the
present study was to identify the diagnostic potential of
EV-derived miRNAs as a non-invasive method for the diagnosis of
NAFLD.
Materials and methods
Study subjects
Serum samples were collected from 50 patients with
NAFLD and 50 healthy individuals at The First Affiliated Hospital
of Nanjing Medical University (Nanjing, China) between January and
August 2018. The presence of FLD was assessed using
ultrasonography. Patients were included in the study if they had
confirmed fatty liver disease with no history of significant
alcohol consumption, viral hepatitis, autoimmune hepatitis,
metabolic diseases or hepatotoxic medication (e.g. amiodarone).
Clinical information, including sex, age, body mass index and
biochemical measurements were obtained from each patient. The
Ethics Committees of Nanjing Medical University approved the
present study and all participants provided written informed
consent.
Animal experiments
The Model Animal Research Center of Nanjing
University provided male C57BL/6J mice, (age, 4–5 weeks; weight,
18–22 g; n=40) which were kept in a pathogen-free barrier facility
at 22°C, with 50% relative humidity and under a 12-h light/dark
cycle. The animals were randomly assigned either into the normal
control (chow diet; CD) or NAFLD group (HFD; 5.56 kcal/g; fat, 58
kcal%; hydrogenated coconut oil, 54%; carbohydrate, 25.5 kcal%) for
12 weeks, as described previously (24). The mice were weighed and euthanized
by cervical dislocation at the end of week 12. The intact livers
were rapidly excised from the abdominal cavity and washed with PBS
for primary hepatic cell culture, RNA extraction or histopathology.
The Animal Experiment Ethics Committee of Nanjing Medical
University approved the study (approval no. IACUC-1601176).
Histopathology
Fresh liver tissue samples were washed with PBS and
placed in 10% buffered formalin at 4°C for 24 h. After gradient
dehydration in ethanol and infiltration in xylene for 30 min, liver
samples were sectioned into 5-µm slices and stained using
hematoxylin and eosin (hematoxylin for 10 min; and eosin for 15 sec
at room temperature) to assess organization and architecture. In
addition, Masson's trichrome and Sirius red staining was performed
for extracellular matrix and fibrosis evaluation using commercially
available kits (Beijing Solarbio Science & Technology Co.,
Ltd.), according to the manufacturer's protocols. Cell apoptosis
was determined using the TUNEL method, as previously described
(25). Neovascularization within
the liver parenchyma and nodules was also determined using CD31
staining (Abcam; cat. no. ab182981; 1:2,000; 4°C overnight). All
stained slides were observed using a Zeiss Axioskop 40®
upright research light microscope with an original magnification,
×40 (Zeiss AG). Histological evaluation was performed in a blinded
manner by two independent pathologists. In order to define the
stage of steatosis, fibrosis, ballooning and parenchymal
inflammation, NASH Activity Score (NAS) (26) was calculated.
Primary hepatic cell isolation
Hepatic cells were isolated using collagenase (type
IV; Sigma-Aldrich; Merck KGaA) perfusion, as described previously
(27). In brief, after the portal
vein was cannulated, the liver was perfused for 5 min with
calcium-free HEPES buffer (0.33 mM; pH, 7.6), followed by perfusion
for 8 min using collagenase (0.025%) and calcium chloride (0.075%)
at 37°C. Following enzymatic digestion, the solution was
centrifuged (50 × g for 2 min at 4°C) to obtain hepatic cells.
Freshly isolated hepatic cells were maintained in DMEM/F12 (Gibco;
Thermo Fisher Scientific, Inc.) containing 10% exosome-depleted FBS
and 1% penicillin/streptomycin. The cells were incubated at 37°C in
a humidified chamber with 5% CO2 for 24 h.
Biochemical assays
Blood was collected from animals via cardiac
puncture following sacrifice. Serum from animals and human subjects
was obtained via centrifugation of whole blood (2,000 × g for 15
min at room temperature) and stored at −80°C. Serum biochemical
markers [alanine aminotransferase (ALT), alkaline phosphatase
(ALP), aspartate aminotransferase (AST), albumin (ALB), globulin,
γ-glutamyl transpeptidase, lactate dehydrogenase (LDH), creatine
kinase (CK), hydroxybutyrate dehydrogenase, total bilirubin,
cholesterol (CHOL), triglyceride (TG), high-density lipoprotein
(HDL), low-density lipoprotein (LDL), lipoprotein a (Lpa), fasting
blood-glucose (GLU0), creatinine (Cr) and uric acid] characterizing
liver disease metabolic features were measured using a biochemical
analyzer (Roche Diagnostics).
Bioinformatics analysis
Putative target genes of the selected miRNAs
(miR-135a-3p and miR-504-3p) were predicted using two databases,
with a pure algorithm (miRDB: mirdb.org and TargetScan: www.targetscan.org/mamm_31) and overlapping
target genes were selected for further functional analysis. In
order to investigate the biological function of the predicted
genes, Gene Ontology (GO) classification and Kyoto Encyclopedia
Genes and Genomes (KEGG) enrichment were performed according to
gene annotation using Database for Annotation, Visualization and
Integrated Discovery (www.kegg.jp).
Fisher's exact test, modified by the false discovery rate, was used
to evaluate the statistical significance of enrichment.
EVs isolation
EVs from in vitro hepatocellular culture
medium were separated using differential centrifugation and
filtration. Briefly, cell culture medium was centrifuged at 10,000
× g for 30 min at 4°C to deplete the cells and debris.
Subsequently, the supernatant was filtered using a 0.22-µm pore
membrane and centrifuged for 70 min at 100,000 × g at 4°C to pellet
the EVs. The pellets were then suspended in 100 µl PBS. Using
ExoQuick reagent (System Biosciences, LLC), EVs from mouse or human
serum were extracted according to the manufacturer's protocol. In
brief, the EVs were isolated by adding EV extraction reagent and
centrifuging for 20 min at 10,000 × g at 4°C. The resulting EV
pellets were stored at −80°C for subsequent experiments.
Western blot analysis
Using RIPA buffer (Beyotime Institute of
Biotechnology), whole mouse primary hepatic cell extracts and EV
proteins were prepared and measured using a BCA protein assay kit
(Beyotime Institute of Biotechnology) according to the
manufacturer's protocol. The extracted proteins (20 µg per lane)
were separated using 10% SDS-PAGE, transferred onto PVDF membranes
and blocked with 5% non-fat milk in TBS-0.05% Tween 20 for 2 h at
room temperature then incubated with primary antibodies against
tumor susceptibility 101 (TSG101; 1:1,000; cat. no. ab125011;
Abcam), CD9 (1:1,000; cat. no. ab92726; Abcam), heat shock protein
70 (HSP70; 1:1,000; cat. no. 33-3800; Invitrogen; Thermo Fisher
Scientific, Inc.), calreticulin (1:1,000; cat. no. ab92516; Abcam)
and GAPDH (1:3,000; cat. no. ab8245; Abcam) overnight at 4°C.
Subsequently, the membrane was incubated with secondary antibody
[goat anti-rabbit IgG (HRP), cat. no. sc-2004; goat anti-mouse IgG
(HRP), cat. no. sc-2005; 1:10,000; Santa Cruz Biotechnology, Inc.]
for 1 h, then a SuperSignal West Pico Chemiluminescent Substrate
kit (Pierce; Thermo Fisher Scientific, Inc.) was used to visualize
the proteins according to the manufacturer's instructions.
BandScan5.0 software (Glyko, Inc.) was used to analyze protein
expression level.
Transmission electron microscopy
(TEM)
In total, 5 µl EV preparation was fixed with 5 µl 4%
paraformaldehyde at 4°C for 30 min and added onto formvar-coated
copper grids to settle for 30 min. Subsequently, the samples were
fixed with 1% glutaraldehyde for 5 min at 25°C, then stained with
2% uranyl oxalate for 5 min at 25°C and 1.8% methyl cellulose
uranyl acetate for 10 min on ice in the dark, embedded in epoxy
resin and polymerized at 35°C for 12 h, 45°C for 12 h and 60°C for
24 h. Residual liquid was removed from the grid with filter paper
and the samples were visualized at 120 kV using the FEI Tecnai G2
Spirit Bio TWIN TEM (FEI; Thermo Fisher Scientific, Inc.). Images
were acquired using an AMT 2k CCD camera (Advanced Microscopy
Techniques, Corp.).
Nanoparticle tracking analysis
(NTA)
In order to identify the diameter and concentration
of EVs, Nanosight NS300 system (Malvern Instruments, Ltd.) fitted
with a fast video capture was used. The images of sample movement
were captured for 60 sec at room temperature. In order to analyze
the videos and measure particle concentrations and size
distribution, the NanoSight LM10 system (NanoSight Ltd.) was used.
Each sample was evaluated ≥3 times.
RNA extraction and reverse
transcription-quantitative (RT-q)PCR
Total RNA was extracted from EVs and cells using
TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's instructions and quantitated using
the NanoDrop 2000 (NanoDrop Technologies; Thermo Fisher Scientific,
Inc.). TRIzol® LS (Invitrogen; Thermo Fisher Scientific,
Inc.) was also used to extract serum RNA. A total of 1 pM
cel-miR-39 RNA oligonucleotide was added to each sample as a
spike-in control when extracting RNA from EVs or serum. A
Taqman® miRNA RT kit and miRNA-specific stem-loop
primers (Applied Biosystems; Thermo Fisher Scientific, Inc.) were
used to detect the expression levels of miRs, according to the
manufacturer's instructions. In order to detect the expression
levels of miRNA, the miR-X miRNA RT-qPCR SYBR kit (Clontech
Laboratories, Inc.) was used for RT-qPCR with the Step One Plus
Real-Time PCR system (Applied Biosystems; Thermo Fisher Scientific,
Inc.) with the following program: Initial denaturation at 95°C for
3 min, followed by 40 cycles of 95°C for 10 sec and annealing and
extension at each annealing temperature at 60°C for 30 sec.
Relative expression level of each miRNA were normalized to
cel-miR-39 and via the 2−ΔΔCq method (28). The following primer sequences were
used for RT-qPCR: cel-miR-39 forward (F),
5′-ACACTCCAGCTGGGGTCACCGGGTGTAAATC-3′ and reverse (R),
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCAAGCTGA−3′; miR-122-5p F,
5′-ACACTCCAGCTGGGTGGAGTGTGACAATGG-3′ and R,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCAAACACC−3′; miR-129-5p F,
5′-ACACTCCAGCTGGGCTTTTTGCGGTCTGG-3′ and R,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGCAAGCCC-3′; miR-135a-3p F,
5′-ACACTCCAGCTGGGTATAGGGATTGGAGCC-3′ and R,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGCCACGG-3′; and miR-504-3p
F, 5′-ACACTCCAGCTGGGGGGAGTGCAGGGCAG-3′ and R,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGAAACCCT-3′. Universal
reverse primer: 5′-TGGTGTCGTGGAGTCG-3′.
Statistical analysis
The data are presented as the mean ± SEM (n≥3). The
differences between two groups were analyzed using unpaired
Student's t-test. Receiver operating characteristic (ROC)
curves were generated based on the expression level of each miRNA
to determine the utility of each miRNA for diagnosing NAFLD.
GraphPad Prism 7 (GraphPad Software, Inc.) and SPSS v22.0 software
(IBM Corp) were used for statistical analysis. P<0.05 was
considered to indicate a statistically significant difference.
Results
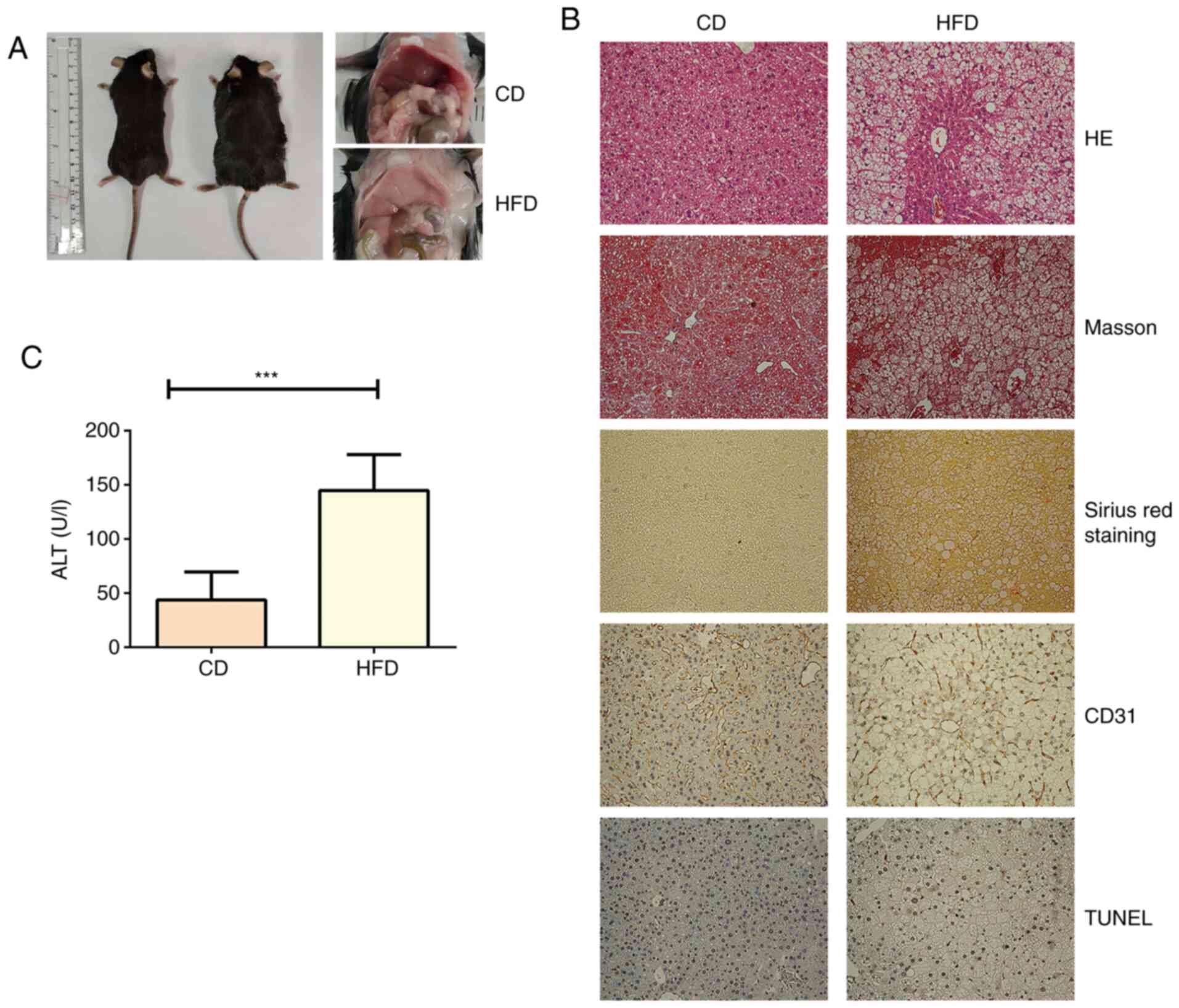
Liver steatosis is induced by HFD
After feeding for 12 weeks, the liver from mice in
the control group showed normal morphology with red, shiny and
smooth appearance, whereas the liver from mice fed HFD exhibited a
slightly yellow color, smooth tight membranes and soft texture
(Fig. 1A). The biochemical analyses
showed that mice fed HFD exhibited significantly higher AST,
albumin, ALP, CHOL, HDL and LDL levels compared with in mice fed CD
(Table I). In addition, the mice
fed HFD exhibited notable hepatic steatosis, liver injury,
hepatocellular ballooning, fibrosis, angiogenesis and cell death
(Fig. 1B and Table SI). Serum ALT levels were also
significantly increased in mice fed HFD compared with in mice fed
CD, suggesting that a HFD induced hepatocyte damage (Fig. 1C).
 | Table I.Biochemical characteristics of HFD
mice and controls. |
Table I.
Biochemical characteristics of HFD
mice and controls.
| Parameter | CD (n=5) | HFD (n=5) | P-value |
|---|
| AST, U/l | 144.80±10.06 | 205.00±11.93 | 0.0048 |
| ALB, g/l | 28.22±0.37 | 26.22±0.42 | 0.0070 |
| ALP, U/l | 77.00±4.65 | 52.60±2.66 | 0.0019 |
| CHOL, mg/dl | 2.56±0.08 | 4.31±0.18 | <0.0001 |
| TG, mmol/l | 0.95±0.06 | 0.76±0.08 | 0.1011 |
| HD, mmol/l | 1.50±0.06 | 1.96±0.06 | 0.0005 |
| LDL, mmol/l | 0.32±0.02 | 0.68±0.05 | 0.0001 |
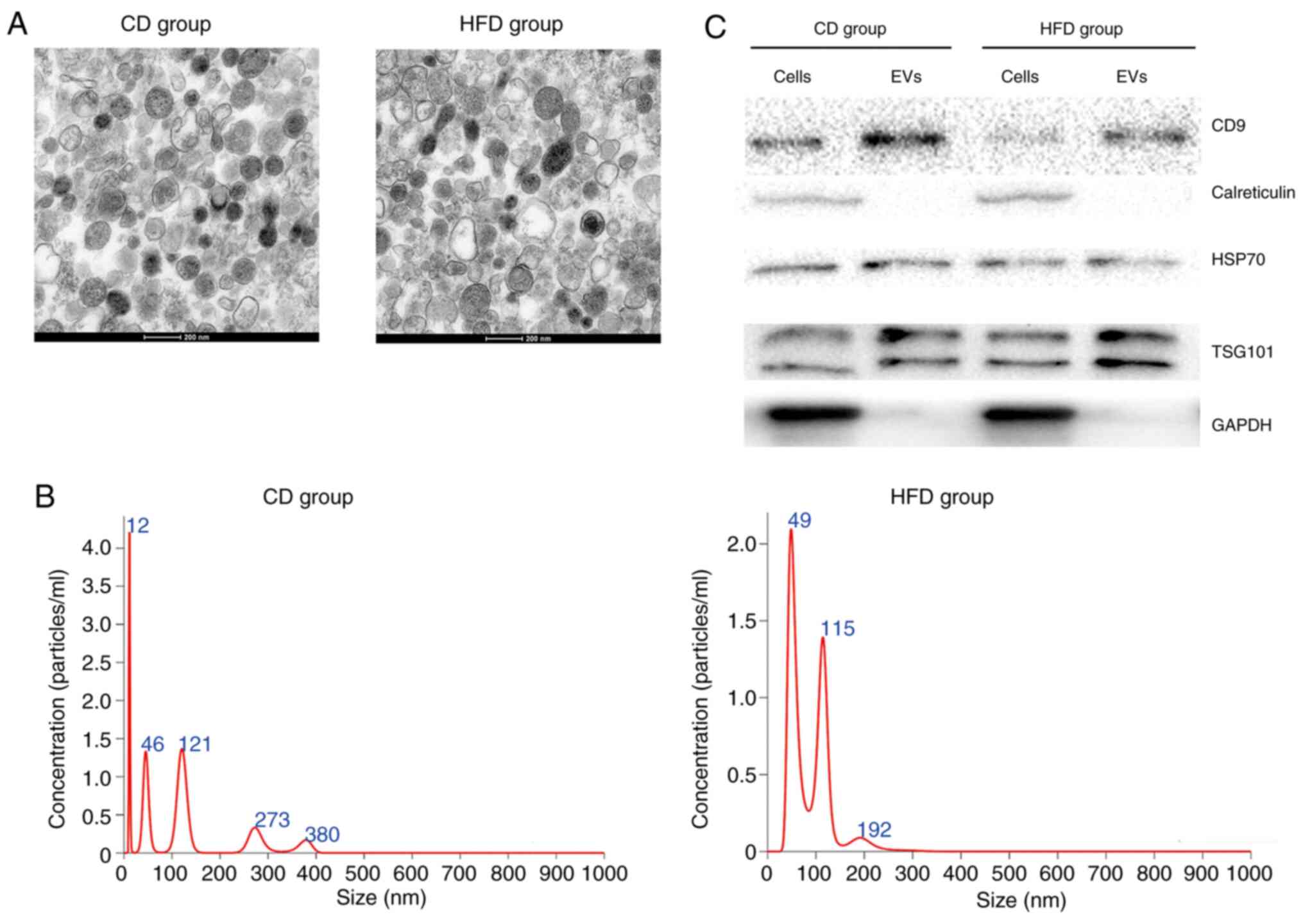
Isolation and characterization of
hepatocyte-released EVs
TEM images showed that isolated EVs demonstrated
typical morphology with an exosomal characteristic bilayer membrane
(Fig. 2A). The size distribution of
EVs was measured using NTA (Fig.
2B), which was within the normal range of EV sizes. Equal
amounts of protein from EVs and primary hepatocytes were separated;
expression levels of exosomal markers CD9, HSP70 and TSG101 were
higher in EVs compared with in primary hepatocytes (Fig. 2C). In addition, calreticulin and
GAPDH were expressed in primary hepatocytes, but not expressed in
EVs (Fig. 2C).
miR-129-5p, miR-135a-3p and miR-504-3p
expression levels decrease in serum EVs in mice fed HFD
According to our previous study (23), C57BL/6J mice were fed HFD for 12
weeks for induction of an NAFLD model. EVs were collected from the
culture medium of hepatocytes from mice fed HFD or CD. There was a
marked difference in the expression levels of several miRNAs in the
hepatocyte-derived EVs from mice fed HFD compared with mice fed CD
(Table II). From these miRNAs,
three human and mouse homologous miRNAs were selected, with fold
change >2 and P<0.05 (miR-129-5p, miR-135a-3p and miR-504-3p)
for further investigation. miR-122-5p, which served as a good
indicator of NAFLD in previous studies (12,15),
was also included for verification.
| Table II.Differential miR signature of
extracellular vesicles derived from hepatocytes of high-fat diet
mice compared with chow diet mice. |
Table II.
Differential miR signature of
extracellular vesicles derived from hepatocytes of high-fat diet
mice compared with chow diet mice.
| miR | P-value | Fold change |
|---|
|
mmu-miR-129b-5p | 0.041 | 0.133 |
|
mmu-miR-135a-3p | 0.050 | 0.431 |
| mmu-miR-1927 | 0.033 | 0.144 |
| mmu-miR-2137 | 0.021 | 0.498 |
|
mmu-miR-3070-2-3p | 0.045 | 0.114 |
|
mmu-miR-3102-3p.2-3p | 0.019 | 2.197 |
| mmu-miR-504-3p | 0.048 | 0.184 |
| mmu-miR-5131 | 0.038 | 0.584 |
|
mmu-miR-5622-3p | 0.026 | 0.433 |
|
mmu-miR-6904-5p | 0.044 | 0.170 |
|
mmu-miR-7018-5p | 0.030 | 0.569 |
|
mmu-miR-7218-5p | 0.040 | 0.196 |
|
mmu-miR-7219-5p | 0.034 | 0.422 |
|
mmu-miR-7221-5p | 0.037 | 0.634 |
|
mmu-miR-7654-3p | 0.032 | 0.469 |
|
mmu-miR-7654-5p | 0.010 | 0.481 |
In order to verify our previous microarray results
(23), the expression levels of
miR-129-5p, miR-135a-3p and miR-504-3p was measured in
hepatocellular EVs using RT-qPCR. The results showed that all three
miRNAs exhibited lower expression levels in hepatocellular EVs from
mice fed HFD compared with in mice fed CD, showing consistency with
our previous microarray results (Fig.
3B-D). The expression levels of miR-122-5p were slightly
decreased in hepatocellular EVs derived from mice fed HFD (Fig. 3A). In order to investigate whether
these miRNAs were present in the liver, the expression levels of
candidate miRNAs in hepatocytes was investigated. miR-122-5p and
miR-504-3p were decreased in primary hepatocytes from mice fed HFD,
while differences in miR-129-5p and miR-135a-3p were not
significant (Fig. S1). Then, it
was investigated whether the selected miRNAs were differentially
expressed in serum or hepatocyte-derived EVs. Expression levels of
miR-504-3p, miR-135a-3p and miR-129-5p were significantly decreased
in serum from mice fed HFD compared with in mice fed CD (Fig. 3F-H). However, there was increased
miR-122-5p expression in serum EVs from HFD mice compared with CD
mice (Fig. 3E).
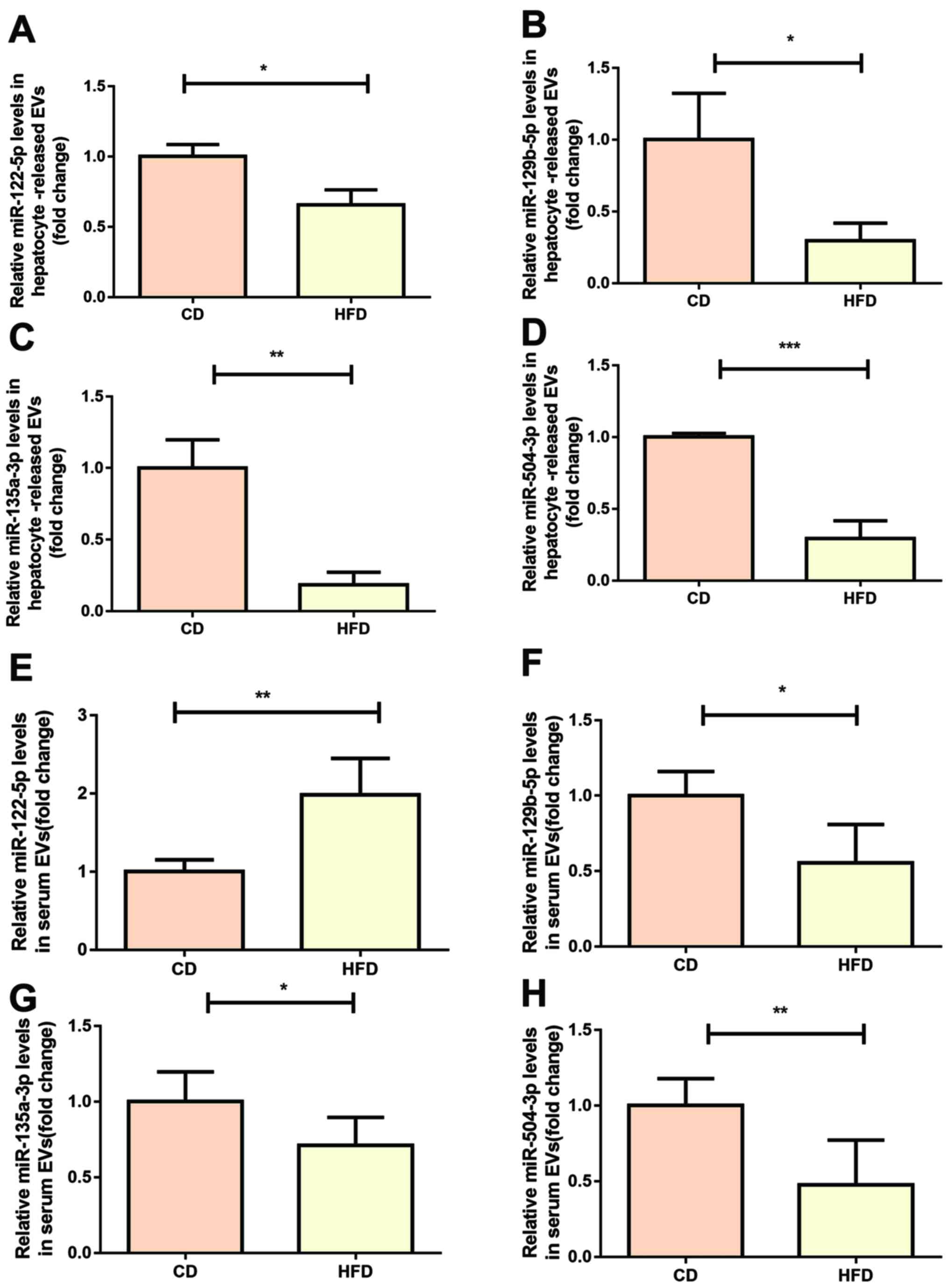 | Figure 3.miR signatures of hepatocyte-released
and serum-derived EVs in mice fed HFD or CD. (A) miR-122-5p, (B)
miR-129-5p, (C) miR-135a-1-3p and (D) miR-504-3p were verified
using RT-qPCR in hepatocyte-released EVs. Expression levels of (E)
miR-122-5p, (F) miR-129-5p, (G) miR-135a-1-3p and (H) miR-504-3p
were detected in serum-derived EVs isolated from mice fed HFD or CD
mice using RT-qPCR (n=5/group). Data are presented as the mean ±
SD. *P<0.05, **P<0.01, ***P<0.001. RT-q, reverse
transcription-quantitative; miR, microRNA; HFD, high-fat diet; CD,
chow diet; EVs, extracellular vesicles. |
miR-135a-3p and miR-504-3p expression
levels decrease in serum EVs from patients with NAFLD
In order to investigate the expression levels of
selected miRNAs, 50 healthy individuals with no signs of liver
disease and 50 patients with NAFLD were recruited. The clinical
features of subjects are shown in Table III. The patients were sex- and
age-matched. The patients with NAFLD and healthy controls did not
exhibit significantly different levels of ALP, CK, hydroxybutyrate
dehydrogenase, total bilirubin, total CHOL, LDL, Lpa, total
protein, ALB, globulin, ALB/globulin, GLU0 and Cr. However,
patients with NAFLD had notably higher AST, γ-glutamyl
transpeptidase, LDH, TG and urea levels and lower HDL levels
compared with healthy controls.
| Table III.Clinical and biochemical
characteristics. |
Table III.
Clinical and biochemical
characteristics.
| Parameter | Healthy controls,
n=50 | Patients with
NAFLD, n=50 | P-value |
|---|
| Sex |
|
|
|
|
Male | 36 (72%) | 39 (78%) |
|
|
Female | 14 (28%) | 11 (22%) |
|
| Age, years | 42.60±1.67 | 42.24±1.50 | 0.8727 |
| AST, U/l | 22.95±0.84 | 26.63±1.23 | 0.0153 |
| ALP, U/l | 79.55±2.56 | 78.59±2.66 | 0.7958 |
| GGT, U/l | 25.60±1.94 | 50.38±6.10 | 0.0002 |
| LDH, U/l | 164.50±2.98 | 175.00±3.47 | 0.0231 |
| CK, U/l | 115.70±13.72 | 118.20±14.64 | 0.9060 |
| HBDH, U/l | 106.30±3.79 | 103.00±2.37 | 0.5102 |
| TBIL, µmol/l | 13.44±0.60 | 12.64±0.62 | 0.3536 |
| TC, µmol/l | 4.89±0.12 | 5.09±0.15 | 0.2961 |
| TG, µmol/l | 1.40±0.11 | 2.47±0.37 | 0.0061 |
| HDL, µmol/l | 1.27±0.03 | 1.15±0.04 | 0.0272 |
| LDL, µmol/l | 3.19±0.10 | 3.27±0.10 | 0.5662 |
| Lpa, mg/l | 210.10±41.60 | 126.00±23.25 | 0.1154 |
| TP, g/l | 73.13±0.75 | 71.71±0.52 | 0.1211 |
| ALB, g/l | 45.73±0.42 | 45.01±0.34 | 0.1871 |
| GLB g/l | 27.90±0.45 | 26.70±0.45 | 0.0638 |
| A/G | 1.66±0.03 | 1.71±0.04 | 0.2861 |
| GLU0, mmol/l | 5.55±0.19 | 5.69±0.32 | 0.7048 |
| Urea, mmol/l | 5.36±0.16 | 5.16±0.17 | 0.4004 |
| Cr, µmol/l | 69.32±1.54 | 68.17±1.63 | 0.6099 |
| UA, µmol/l | 328.60±9.25 | 373.30±13.46 | 0.0074 |
As EVs protect circulating miRNA from degradation by
RNase, it was hypothesized that miRNA expression levels in EVs from
serum would detect NAFLD progression more effectively compared with
those in total serum. Using RT-qPCR and cel-miR-39 as an exogenous
normalizer, it was found that miR-135a-3p, miR-129b-5p and
miR-504-3p expression levels in EVs isolated from the serum of
patients with NAFLD were markedly decreased compared with in the
healthy controls, confirming this hypothesis. In addition,
miR-122-5p in circulating EVs and serum ALT expression levels were
significantly increased in patients with NAFLD (Fig. 4A-E). In order to evaluate the
diagnostic potential of candidate miRNAs, ROC curve analysis was
performed. miR-135a-3p exhibited the highest area under the curve
(AUC; 0.849; 95% CI, 0.777–0.921), while miR-122-5p and miR-504-3p
had AUC values of 0.790 (95% CI, 0.700–880) and 0.708 (95% CI,
0.606–0.810), respectively. The AUC of the traditional diagnostic
indicator ALT was 0.672 (95% CI, 0.567–0.778; Fig. 4F-J). These data indicated that
miR-135a-3p in EVs may be a promising diagnostic indicator and were
more sensitive than ALT.
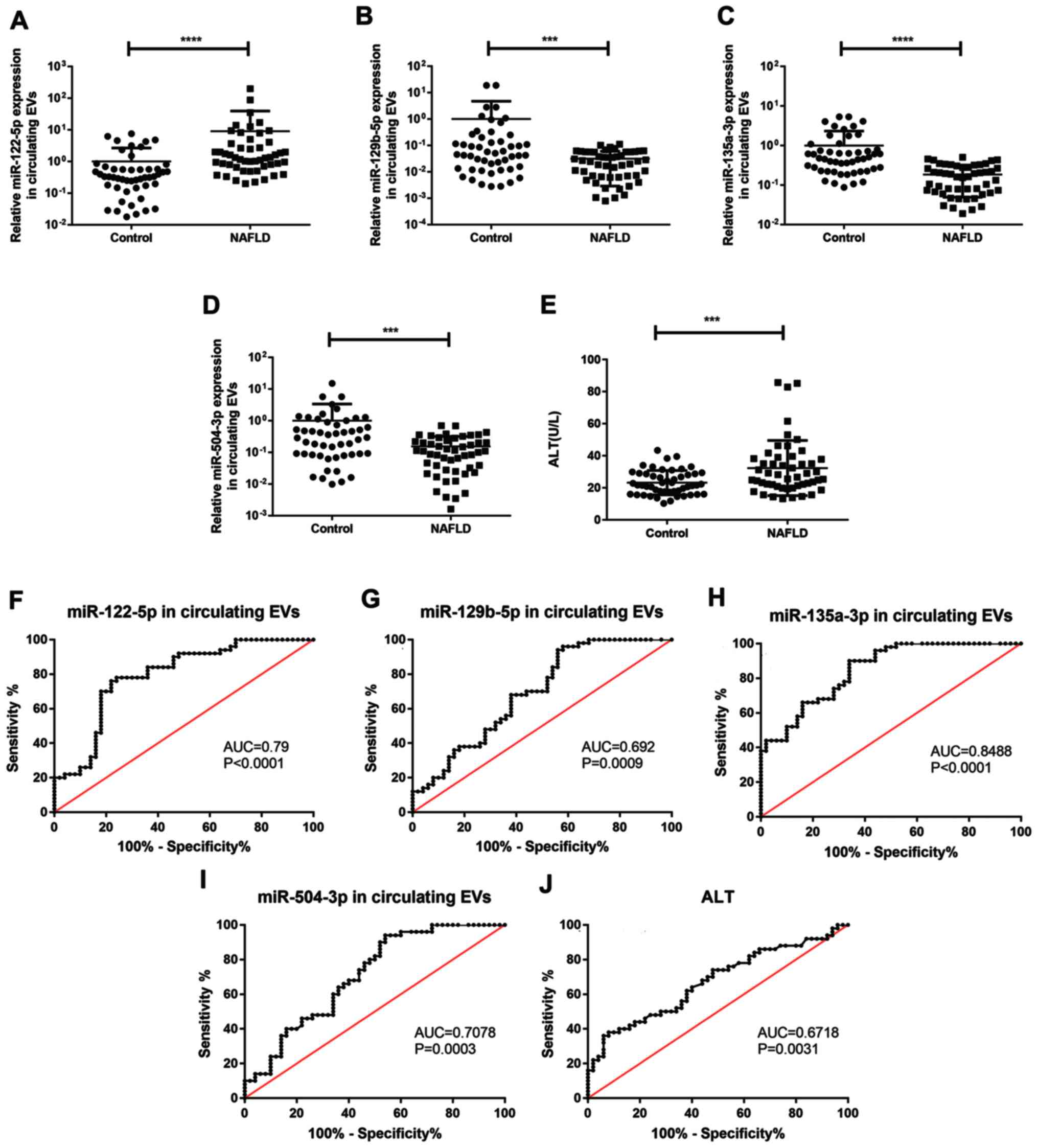 | Figure 4.Expression levels and ROC curves of
screened miRs in serum EVs. Reverse transcription-quantitative PCR
was used to determine mRNA expression levels of (A) miR-122-5p, (B)
miR-129-5p, (C) miR-135a-3p and (D) miR-504-3p in serum EVs and (E)
serum ALT levels in patients with NAFLD (n=50) and control subjects
(n=50). ROC curves and AUC for serum EV-derived (F) miR-122-5p, (G)
miR-135a-3p, (H) miR-129-5p, (I) miR-504-3p and (J) ALT levels in
patients with NAFLD. Data are presented as the mean ± SD.
***P<0.001, ****P<0.0001. ROC, receiver operator
characteristic; miR, microRNA, EVs, extracellular vesicles; AUC,
area under the curve; ALT, alanine aminotransferase; NAFLD,
non-alcoholic fatty liver disease. |
Subsequently, the presence of these miRNAs in serum
from patients with NAFLD was investigated. miR-135a-3p expression
levels were markedly lower, whereas those of miR-122-5p were
notably increased in patients with NAFLD compared with healthy
controls. However, there was no notable difference in expression
levels of miR-129-5p and miR-504-3p in serum between patients with
NAFLD and healthy controls (Fig.
S2A-D). Thus, in NAFLD, some miRNAs were specifically enriched
in EVs, and detection of serum exosomes was a more favorable
diagnostic method than direct detection of serum miRNA for
NAFLD.
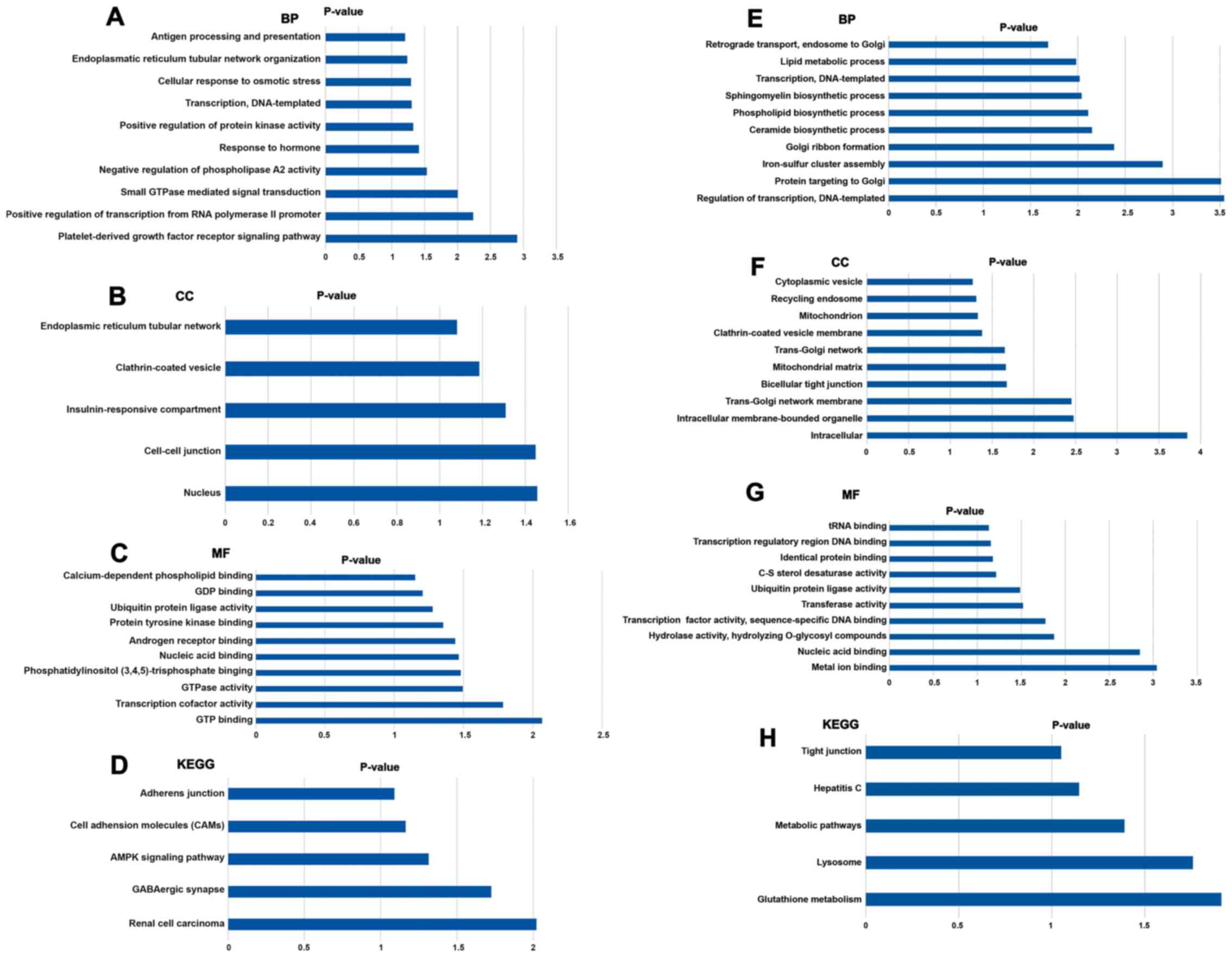
Biological function analysis
KEGG and GO analysis were used to investigate the
key biological associations, signaling pathways and molecular
functions of target genes of miR-135a-3p and miR-504-3p. For
miR-135a-3p, the most significantly enriched GO terms were
associated with ‘platelet-derived growth factor receptor signaling
pathway’, ‘cell-cell junction’ and ‘GTP binding’ (Fig. 5A-C). The significantly enriched KEGG
pathways had functions associated with ‘AMP-activated protein
kinase (AMPK) signaling pathway’ (Fig.
5D). The target genes of miR-504-3p were significantly enriched
in the GO terms ‘nucleotide-binding, oligomerization domain-like
receptor signaling pathway’, ‘glutamatergic synapse’, ‘inflammatory
responses’ and ‘ion transmembrane transport’ (Fig. 5E-G). According to the KEGG
enrichment analysis, the most significantly enriched pathway for
the target genes of miR-504-3p was ‘glutathione metabolism’
(Fig. 5H). Furthermore, ‘lysosome’
and ‘metabolic pathways’ were also highly enriched.
Discussion
NAFLD is a chronic liver disease and widely occurs
in adult and children. During the past 20 years, the prevalence of
NAFLD in China has doubled (3). The
overall prevalence rate of NAFLD for adults in China has been ~15%,
although there are regional variations (3). Moreover, the prevalence of NAFLD is
2.1% in children and 68.2% in obese children (3,5).
Measurement of the serum AST/ALT ratio and AFP have been suggested
as indicators of NAFLD; however, normal ALT or AFP levels can be
detected in patients with steatosis and early-stage HCC (29–31).
There are limitations to the existing diagnostic approaches (such
as unfeasible serum markers and imaging techniques); therefore,
tissue biopsy continues to be the primary NASH diagnostic standard
(32–35), although biopsy is invasive and
sampling errors can occur (36). In
addition, biopsy cannot be used to predict disease progression
(37). Thus, potential stable and
non-invasive markers for liver disease progression are increasingly
investigated.
During lipotoxicity, hepatocytes release a high
number of EVs that affect various target cells (including
macrophages, Kuffer cells and hepatic stellate cells) and improve
key processes associated with NAFLD pathogenesis (such as fibrosis,
angiogenesis and immune regulation) (38,39).
EVs generated and released during NAFLD progression exhibit a
special antigenic composition (such as C-X-C motif chemokine ligand
10, C16:0 ceramide and inositol requiring enzyme 1α), which
reflects characteristic pathological changes in its progression,
the number of miRNAs and proteins released from the liver increases
(40). In addition, the level of
EVs is dynamic and associated with variations in liver
histopathology. Thus, in the present study well-defined and stable
miRNAs were investigated to demonstrate whether Evs-derived miRNAs
may be effective non-invasive biomarkers to determine NAFLD
progression.
Previous studies have revealed that serum miR-16,
miR-34a, miR-21 and miR-122 expression levels increase in patients
with NAFLD (15,41). Furthermore, circulating EVs exhibit
increased expression levels of asialoglycoprotein receptor 1,
carboxylesterase 1 and liver miRNAs (miR-192 and miR-122) in
patients with NAFLD, while miR-192 and miR-122 expression levels
decrease in the liver (42–44). EVs protect miRNAs from RNase
degradation (18); therefore, both
miRNA biological marker sensitivity and specificity may be enhanced
by enriching EVs.
In the present study, EV-derived miR-504-3p and
miR-135a-3p expression levels were notably lower in patients with
NAFLD compared with healthy controls. ROC analysis indicated that
EV-derived miR-135a-3p and miR-504-3p may be sensitive biological
markers for diagnosis of NAFLD, particularly miR-135a-3p, which
exhibited the highest AUC and was more sensitive than ALT. However,
there was no significant difference in serum circulating miR-504-3p
expression levels between patients with NAFLD and healthy
individuals. Total miRNAs from circulating serum may contain
endogenous cellular miRNAs originating from debris from various
cells; cell lysis and artifacts may influence expression levels of
serum circulating miRNA (15).
Recent studies have shown that exosomal miRNA is stable in serum
because exosomes serve a protective function against degradation
from enzymes, such as RNAse (15,18).
Therefore, most studies use EVs to enhance the sensitivity and
specificity of miRNA biological markers (45,46).
Regarding the difference between miRNAs from hepatocytes and
hepatocyte-derived EVs, a more notable change was observed between
patients with NAFLD and healthy individuals in hepatocyte-derived
EVs, suggesting that EVs derived from hepatocytes may selectively
increase the expression of certain miRNAs to reflect the condition
of the body (47). Hence, miRNAs
from hepatocyte-derived EVs may be a more sensitive biological
marker than miRNAs from hepatocytes. When comparing the expression
level patterns of miRNAs from serum EVs and hepatocyte-derived EVs,
consistency was found between these two indicators. As serum EVs
are more accessible, miRNAs derived from serum EVs may be used to
determine NAFLD progression. Consistent with previous studies
(48,49), miR-122-5p was increased in serum EVs
and decreased in the liver of mice fed HFD. Hepatocytes may serve
as a key source of circulating EVs and miR-122 is predominantly
encapsulated in EVs in animals with NAFLD (42). Following stress or damage,
hepatocytes release these miRNAs in EVs during NAFLD progression,
which may explain the decrease in the expression levels of miR-122
in liver from patients with NAFLD (49). In renal fibrosis, the expression
levels of exosomal miR-29c are negatively associated with
tubulointerstitial fibrosis score (50). Severity of liver injury also
dictates EV-derived miRNA compartmentalization (51). Thus, it is important to investigate
the use of EVs to reflect the activity and severity of NAFLD in
future.
Hepatic cells may simultaneously serve as an
EV-mediated intercellular source of communication and as a target
(52). The effect of EV-derived
miR-135a-3p and miR-504-3p is important during the development of
NAFLD. Dysregulated expression of miR-504 is found in numerous
types of tumor (such as HCC). Overexpression of miR-504 attenuates
the invasion and proliferation of HCC cells by targeting frizzled
class receptor 7, thus inhibiting the Wnt/β-catenin signaling
pathway, which is key to the metabolism and pathology of the liver
(53,54). A tumor suppressing effect of miR-504
in HCC, mediated by negatively regulating the expression levels of
CDK16, has also been reported by Li et al (55). In addition to miR-504, miR-135a also
suppresses the development of tumors by suppressing C-C motif
chemokine receptor 2 expression levels in ovarian cancer (56). Furthermore, miR-135a can predict and
prognose patients with urothelial carcinoma of the bladder
(57). The results of the present
study confirmed the diagnostic potential of miR-135a-3p and
miR-504-3p in NAFLD. In addition, KEGG pathway analysis predicted
the potential roles of miR-135a-3p and miR-504-3p in regulating the
AMPK and metabolic pathways, which are involved in NAFLD (58).
However, there are certain limitations to the
present study. NAFLD is not an isolated disease but can occur as a
complication of insulin resistance and other metabolic disorders
(59). Thus, a variety of tissues
(such as pancreas, muscle and adipose tissue) are impacted by the
associated oxidative stress and fatty acid flux (59), thereby promoting immune cell
activation and migration to these sites. Therefore, extrahepatic
release of EVs may hide NAFLD pathogenesis. In addition, there is a
lack of specific liver molecular markers; further studies are
required to investigate the association between circulating EVs and
its source from the liver. Furthermore, there was a small number of
subjects enrolled into the present study; a prospective cohort
study is required to verify the diagnostic value of miR-135a-3p and
miR-504-3p. In addition, the target genes of the selected miRNAs
and their roles in the development of NAFLD were not investigated.
The functions of miR-135a-3p and miR-504-3p should be determined in
future studies.
In summary, the present study suggested that
downregulated expression levels of circulating EV-derived
miR-135a-3p was a potential novel and non-invasive marker to
diagnose NAFLD; however further investigation is required.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Gongqian Jiao
(The First Affiliated Hospital of Nanjing Medical University) for
providing serum samples from patients with NAFLD.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81830023 and
81600640).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
TY and QF worked on the conception and design of the
study, HMJ wrote the manuscript. HMJ, YQ, ZYS, YQH and SC performed
the experiments. HMJ, YWL, RG and MS collected and analyzed data.
HMJ, SC and MS confirmed the authenticity of all the raw data. All
authors contributed to the preparation of the manuscript, and read
and approved the final manuscript.
Ethics approval and consent to
participate
The Ethics Committees of Nanjing Medical University
approved the present study and all participants provided written
informed consent. All animal experiments in this study were
approved by The Animal Experiment Ethics Committee of Nanjing
Medical University (approval no. IACUC-1601176).
Patient consent for publication
Not applicable.
Competing interests
The authors confirm that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
RT-q
|
reverse transcription-quantitative
|
|
TEM
|
transmission electron microscopy
|
|
NTA
|
nanoparticle tracking analysis
|
|
miR
|
microRNA
|
|
CD
|
chow diet
|
|
HFD
|
high fat diet
|
|
EVs
|
extracellular vesicles
|
|
AMPK
|
AMP-activated protein kinase
|
|
TSG101
|
tumor susceptibility 101
|
|
HSP70
|
heat shock protein 70
|
References
|
1
|
Albhaisi S and Sanyal A: Recent advances
in understanding and managing non-alcoholic fatty liver disease.
F1000Res. 7:F1000 Faculty Rev-720. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lucas C, Lucas G, Lucas N, Krzowska-Firych
J and Tomasiewicz K: A systematic review of the present and future
of non-alcoholic fatty liver disease. Clin Exp Hepatol. 4:165–174.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Younossi Z: Non-alcoholic fatty liver
disease-A global public health perspective. J Hepatol. 70:531–544.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fan JG and Farrell GC: Epidemiology of
non-alcoholic fatty liver disease in China. J Hepatol. 50:204–210.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan JG: Epidemiology of alcoholic and
nonalcoholic fatty liver disease in China. J Gastroenterol Hepatol.
28 (Suppl 1):S11–S17. 2013. View Article : Google Scholar
|
|
6
|
Adams LA, Harmsen S, St Sauver JL,
Charatcharoenwitthaya P, Enders FB, Therneau T and Angulo P:
Nonalcoholic fatty liver disease increases risk of death among
patients with diabetes: A community-based cohort study. Am J
Gastroenterol. 105:1567–1573. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Katsiki N, Athyros VG, Karagiannis A and
Mikhailidis DP: Managing the combination of non-alcoholic fatty
liver disease and metabolic syndrome. Expert Opin Pharmacother.
13:287–290. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Byrne CD and Targher G: NAFLD: A
multisystem disease. J Hepatol. 62 (1 Suppl):S47–S64. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mazhar K: The future of nonalcoholic fatty
liver disease treatment. Med Clin North Am. 103:57–69. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
He Z, Yang JJ, Zhang R, Li HT, Wu L, Jiang
F, Jia WP and Hu C: Circulating miR-29b positively correlates with
non-alcoholic fatty liver disease in a Chinese population. J Dig
Dis. 20:189–195. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
DiStefano JK and Gerhard GS: Circulating
microRNAs in nonalcoholic fatty liver disease. Expert Rev
Gastroenterol Hepatol. 10:161–163. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yamada H, Suzuki K, Ichino N, Ando Y,
Sawada A, Osakabe K, Sugimoto K, Ohashi K, Teradaira R, Inoue T, et
al: Associations between circulating microRNAs (miR-21, miR-34a,
miR-122 and miR-451) and non-alcoholic fatty liver. Clin Chim Acta.
424:99–103. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu CH, Ampuero J, Gil-Gomez A,
Montero-Vallejo R, Rojas Á, Muñoz-Hernández R, Gallego-Durán R and
Romero-Gómez M: miRNAs in patients with non-alcoholic fatty liver
disease: A systematic review and meta-analysis. J Hepatol.
69:1335–1348. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Tan Y, Ge G, Pan T, Wen D and Gan J: A
pilot study of serum microRNAs panel as potential biomarkers for
diagnosis of nonalcoholic fatty liver disease. PLoS One.
9:e1051922014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Afonso MB, Rodrigues PM, Simao AL and
Castro RE: Circulating microRNAs as potential biomarkers in
non-alcoholic fatty liver disease and hepatocellular carcinoma. J
Clin Med. 5:302016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
He Y, Deng F, Yang S, Wang D, Chen X,
Zhong S, Zhao J and Tang J: Exosomal microRNA: A novel biomarker
for breast cancer. Biomark Med. 12:177–188. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang G, Lin G, Zhu Y, Duan W and Jin D:
Emerging technologies for profiling extracellular vesicle
heterogeneity. Lab Chip. 20:2423–2437. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ibáñez F, Ureña-Peralta JR, Costa-Alba P,
Torres JL, Laso FJ, Marcos M, Guerri C and Pascual M: Circulating
MicroRNAs in extracellular vesicles as potential biomarkers of
alcohol-induced neuroinflammation in adolescence: Gender
differences. Int J Mol Sci. 21:67302020. View Article : Google Scholar
|
|
19
|
Nedaeinia R, Manian M, Jazayeri MH,
Ranjbar M, Salehi R, Sharifi M, Mohaghegh F, Goli M, Jahednia SH,
Avan A and Ghayour-Mobarhan M: Circulating exosomes and exosomal
microRNAs as biomarkers in gastrointestinal cancer. Cancer Gene
Ther. 24:48–56. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Takahasi K, Iinuma H, Wada K, Minezaki S,
Kawamura S, Kainuma M, Ikeda Y, Shibuya M, Miura F and Sano K:
Usefulness of exosome-encapsulated microRNA-451a as a minimally
invasive biomarker for prediction of recurrence and prognosis in
pancreatic ductal adenocarcinoma. J Hepatobiliary Pancreat Sci.
25:155–161. 2018. View
Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yoshikawa M, Iinuma H, Umemoto Y,
Yanagisawa T, Matsumoto A and Jinno H: Exosome-encapsulated
microRNA-223-3p as a minimally invasive biomarker for the early
detection of invasive breast cancer. Oncol Lett. 15:9584–9592.
2018.PubMed/NCBI
|
|
22
|
Zhang W, Ni M, Su Y, Wang H, Zhu S, Zhao A
and Li G: MicroRNAs in serum exosomes as potential biomarkers in
clear-cell renal cell carcinoma. Eur Urol Focus. 4:412–419. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Fu Q, Li Y, Jiang H, Shen Z, Gao R, He Y,
Liu Y, Xu K and Yang T: Hepatocytes derived extracellular vesicles
from high-fat diet induced obese mice modulate genes expression and
proliferation of islet β cells. Biochem Biophys Res Commun.
516:1159–1166. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tessitore A, Cicciarelli G, Del Vecchio F,
Gaggiano A, Verzella D, Fischietti M, Mastroiaco V, Vetuschi A,
Sferra R, Barnabei R, et al: MicroRNA expression analysis in high
fat diet-induced NAFLD-NASH-HCC progression: Study on C57BL/6J
mice. BMC Cancer. 16:32016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tasci I, Mas MR, Vural SA, Deveci S,
Comert B, Alcigir G, Mas N, Akay C, Bozdayi M, Yurdaydin C, et al:
Pegylated interferon-alpha plus taurine in treatment of rat liver
fibrosis. World J Gastroenterol. 13:3237–3244. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Brunt EM, Kleiner DE, Wilson LA, Belt P
and Neuschwander-Tetri BA; NASH Clinical Research Network (CRN), :
Nonalcoholic fatty liver disease (NAFLD) activity score and the
histopathologic diagnosis in NAFLD: Distinct clinicopathologic
meanings. Hepatology. 53:810–820. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Pigeon C, Ilyin G, Courselaud B, Leroyer
P, Turlin B, Brissot P and Loréal O: A new mouse liver-specific
gene, encoding a protein homologous to human antimicrobial peptide
hepcidin, is overexpressed during iron overload. J Biol Chem.
276:7811–7819. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Angulo P: GI epidemiology: Nonalcoholic
fatty liver disease. Aliment Pharmacol Ther. 25:883–889. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mofrad P, Contos MJ, Haque M, Sargeant C,
Fisher RA, Luketic VA, Sterling RK, Shiffman ML, Stravitz RT and
Sanyal AJ: Clinical and histologic spectrum of nonalcoholic fatty
liver disease associated with normal ALT values. Hepatology.
37:1286–1292. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Vogel A, Cervantes A, Chau I, Daniele B,
Llovet JM, Meyer T, Nault JC, Neumann U, Ricke J, Sangro B, et al:
Hepatocellular carcinoma: ESMO clinical practice guidelines for
diagnosis, treatment and follow-up. Ann Oncol. 30:871–873. 2019.
View Article : Google Scholar
|
|
32
|
Deffieux T, Gennisson JL, Bousquet L,
Corouge M, Cosconea S, Amroun D, Tripon S, Terris B, Mallet V,
Sogni P, et al: Investigating liver stiffness and viscosity for
fibrosis, steatosis and activity staging using shear wave
elastography. J Hepatol. 62:317–324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Khov N, Sharma A and Riley TR: Bedside
ultrasound in the diagnosis of nonalcoholic fatty liver disease.
World J Gastroenterol. 20:6821–6825. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Myers RP, Pomier-Layrargues G, Kirsch R,
Pollett A, Beaton M, Levstik M, Duarte-Rojo A, Wong D, Crotty P and
Elkashab M: Discordance in fibrosis staging between liver biopsy
and transient elastography using the FibroScan XL probe. J Hepatol.
56:564–570. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Myers RP, Pomier-Layrargues G, Kirsch R,
Pollett A, Duarte-Rojo A, Wong D, Beaton M, Levstik M, Crotty P and
Elkashab M: Feasibility and diagnostic performance of the FibroScan
XL probe for liver stiffness measurement in overweight and obese
patients. Hepatology. 55:199–208. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Angulo P: Long-term mortality in
nonalcoholic fatty liver disease: Is liver histology of any
prognostic significance? Hepatology. 51:373–375. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dongiovanni P, Meroni M, Longo M, Fargion
S and Fracanzani AL: miRNA Signature in NAFLD: A turning point for
a non-invasive diagnosis. Int J Mol Sci. 19:39662018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dorairaj V, Sulaiman S, Abu N and Abdul
Murad N: Extracellular vesicles in the development of the
non-alcoholic fatty liver disease: An update. Biomolecules.
10:14942020. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Banales JM, Feldstein AE, Sanger H,
Lukacs-Kornek V, Szabo G and Kornek M: Extracellular vesicles in
liver diseases: Meeting report from the international liver
congress 2018. Hepatol Commun. 3:305–315. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Garcia-Martinez I, Alen R, Rada P and
Valverde AM: Insights into extracellular vesicles as biomarker of
NAFLD pathogenesis. Front Med (Lausanne). 7:3952020. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Otsuka M, Kishikawa T, Yoshikawa T,
Yamagami M, Ohno M, Takata A, Shibata C, Ishibashi R and Koike K:
MicroRNAs and liver disease. J Hum Genet. 62:75–80. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Povero D, Eguchi A, Li H, Johnson CD,
Papouchado BG, Wree A, Messer K and Feldstein AE: Circulating
extracellular vesicles with specific proteome and liver microRNAs
are potential biomarkers for liver injury in experimental fatty
liver disease. PLoS One. 9:e1136512014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Povero D, Eguchi A, Niesman IR, Andronikou
N, de Mollerat du Jeu X, Mulya A, Berk M, Lazic M, Thapaliya S,
Parola M, et al: Lipid-induced toxicity stimulates hepatocytes to
release angiogenic microparticles that require Vanin-1 for uptake
by endothelial cells. Sci Signal. 6:ra882013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Csak T, Bala S, Lippai D, Satishchandran
A, Catalano D, Kodys K and Szabo G: microRNA-122 regulates
hypoxia-inducible factor-1 and vimentin in hepatocytes and
correlates with fibrosis in diet-induced steatohepatitis. Liver
Int. 35:532–541. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Madhavan D, Zucknick M, Wallwiener M, Cuk
K, Modugno C, Scharpff M, Schott S, Heil J, Turchinovich A, Yang R,
et al: Circulating miRNAs as surrogate markers for circulating
tumor cells and prognostic markers in metastatic breast cancer.
Clin Cancer Res. 18:5972–5982. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Mori MA, Ludwig RG, Garcia-Martin R,
Brando BB and Kahn CR: Extracellular miRNAs: From biomarkers to
mediators of physiology and disease. Cell Metab. 30:656–673. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Jiang F, Chen Q, Wang W, Ling Y, Yan Y and
Xia P: Hepatocyte-derived extracellular vesicles promote
endothelial inflammation and atherogenesis via microRNA-1. J
Hepatol. 72:156–166. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Cermelli S, Ruggieri A, Marrero JA,
Ioannou GN and Beretta L: Circulating microRNAs in patients with
chronic hepatitis C and non-alcoholic fatty liver disease. PLoS
One. 6:e239372011. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Cheung O, Puri P, Eicken C, Contos MJ,
Mirshahi F, Maher JW, Kellum JM, Min H, Luketic VA and Sanyal AJ:
Nonalcoholic steatohepatitis is associated with altered hepatic
MicroRNA expression. Hepatology. 48:1810–1820. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lv LL, Cao YH, Ni HF, Xu M, Liu D, Liu H,
Chen PS and Liu BC: MicroRNA-29c in urinary exosome/microvesicle as
a biomarker of renal fibrosis. Am J Physiol Renal Physiol.
305:F1220–F1227. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Murakami Y, Toyoda H, Tanahashi T, Tanaka
J, Kumada T, Yoshioka Y, Kosaka N, Ochiya T and Taguchi YH:
Comprehensive miRNA expression analysis in peripheral blood can
diagnose liver disease. PLoS One. 7:e483662012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Devhare PB and Ray RB: Extracellular
vesicles: Novel mediator for cell to cell communications in liver
pathogenesis. Mol Aspects Med. 60:115–122. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Quan H, Li B and Yang J: MicroRNA-504
functions as a tumor suppressor in hepatocellular carcinoma through
inhibiting Frizzled-7-mediated-Wnt/β-catenin signaling. Biomed
Pharmacother. 107:754–762. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Tian Y, Mok MT, Yang P and Cheng AS:
Epigenetic activation of Wnt/β-catenin signaling in
NAFLD-associated hepatocarcinogenesis. Cancers (Basel). 8:762016.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Li C, Miao R, Zhang J, Qu K and Liu C:
Long non-coding RNA KCNQ1OT1 mediates the growth of hepatocellular
carcinoma by functioning as a competing endogenous RNA of miR-504.
Int J Oncol. 52:1603–1612. 2018.PubMed/NCBI
|
|
56
|
Duan S, Dong X, Hai J, Jiang J, Wang W,
Yang J, Zhang W and Chen C: MicroRNA-135a-3p is downregulated and
serves as a tumour suppressor in ovarian cancer by targeting CCR2.
Biomed Pharmacother. 107:712–720. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Inamoto T, Uehara H, Akao Y, Ibuki N,
Komura K, Takahara K, Takai T, Uchimoto T, Saito K, Tanda N, et al:
A Panel of MicroRNA signature as a tool for predicting survival of
patients with urothelial carcinoma of the bladder. Dis Markers.
2018:54686722018. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Esquejo RM, Salatto CT, Delmore J,
Albuquerque B, Reyes A, Shi Y, Moccia R, Cokorinos E, Peloquin M,
Monetti M, et al: Activation of Liver AMPK with PF-06409577
Corrects NAFLD and lowers cholesterol in rodent and primate
preclinical models. EBioMedicine. 31:122–132. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Watt MJ, Miotto PM, De Nardo W and
Montgomery MK: The liver as an endocrine organ-linking NAFLD and
insulin resistance. Endoc Rev. 40:1367–1393. 2019. View Article : Google Scholar : PubMed/NCBI
|