Introduction
Lung cancer is the most prevalent type of cancer in
the world, accounting for ~1.8 million new lung cancer cases and
1.6 million lung cancer-associated deaths annually (1). The 5-year survival rate of patients
with lung cancer varies from 4–17%, based on their pathological
stage and region (2). Lung cancer
can be categorized as either non-small cell lung cancer or small
cell lung cancer according to its pathological features. The early
symptoms of lung cancer are not obvious, usually resulting in
diagnosis in the middle or later stages of its development
(3). The etiological complexity and
frequent metastasis of lung cancer are the primary factors
underlying the difficulty in treatment and the high mortality rates
associated with it (4). Tumor
metastasis is a complex biological process, and its mechanism is
poorly understood. The epithelial-mesenchymal transition (EMT)
process is considered an early and critical step in the tumor
metastatic cascade (5). Cigarette
smoking is the leading cause of all histological types of lung
cancer (6). Our previous work on
the carcinogenic potential of cigarette smoke (CS) in normal human
bronchial epithelial (BEAS-2B) cells showed aberrant DNA
methylation and transcriptional profiles, as well as an increase in
tumorgenicity and migratory ability (7–9).
MicroRNAs (miRNAs/miRs) are endogenous
single-stranded non-coding RNAs of ~20–24 nucleotides in length
(10). It has been widely reported
that miRNAs negatively regulate the expression of various mRNAs by
binding to the 3′-untranslated regions (3′-UTR) of mRNAs (11). miRNAs exhibit unique and diverse
expression patterns and modulate several cellular processes and
signaling pathways related to tumorigenesis, cell proliferation,
metastasis, invasion and apoptosis (12,13).
Our previous study identified multiple miRNAs that are aberrantly
expressed in CS-induced malignantly transformed cells (9). The present study focused on one of
these miRNAs, miR-200b, as the aberrant expression of the miR-200
family has been reported to be associated with the occurrence and
development of various types of malignant tumors, such as
hepatocellular carcinoma (14),
colon cancer (15), breast cancer
(16), ovarian cancer (17) and lung cancer (18). The miR-200 family is comprised of
five members, miR-200a, miR-200b, miR-200c, miR-429 and miR-141.
Accumulating evidence suggests that miR-200b serves an important
role in EMT, cancer stem cell maintenance, apoptosis and in the
cell cycle (19). miR-200b has also
been reported to be upregulated in lung cancer cells, where it is
associated with cell proliferation and metastasis (20). However, the exact role of miR-200b
in mediating lung cancer cell migration and invasion, as well as
the underlying molecular mechanisms involved remain to be
determined.
The aim of the present study was to identify the
biological function and regulatory mechanism of miR-200b in the
CS-induced malignant transformed BEAS-2B cells to explore the
molecular mechanisms underlying CS-induced lung cancer.
Bioinformatics analysis and in vitro experiments were used
to analyze and verify the function and target gene of miR-200b.
Materials and methods
Data source and bioinformatics
analysis
Log2 transformed RNA-seq expression data in lung
cancer tissues, including lung adenocarcinoma [The Cancer Genome
Atlas (TCGA) LUAD] and lung squamous cell carcinoma (TCGA LUSC),
were obtained from TCGA database through the University of
California Santa Cruz (UCSC) Xena Browser (xena.ucsc.edu) (21).
Additionally, four datasets for the analysis of miR-200b-3p
(miR-200b), accession nos. GSE62182 (22), GSE74190 (23), GSE51853 (24) and GSE48414 (25), and two datasets for ETS
proto-oncogene 1 transcription factor (ETS1) analysis,
accession nos. GSE19804 (26) and
GSE27262 (27), were downloaded
from the Gene Expression Omnibus (GEO) database (ncbi.nlm.nih.gov/geo/). The protein levels were
determined using the ‘CPTAC lung adenocarcinoma’ dataset, which was
downloaded from the Clinical Proteomic Tumor Analysis Consortium
(CPTAC) data portal (cptac-data-portal.georgetown.edu/cptacpublic)
(28). The Pearson correlation
analysis of miR-200b with ETS1 was analyzed using the
webtool ‘LinkedOmics’ (linkedomics.org/admin.php). Enrichment analysis was
performed using the webtool Database for Annotation, Visualization
and Integrated Discovery (DAVID; david.ncifcrf.gov/).
Cell culture
Normal human bronchial epithelial cells (BEAS-2B),
as well as lung adenocarcinoma cells (A549 and H1299) and lung
squamous carcinoma cells (H226) were obtained from the American
Type Culture Collection and cultured in high-glucose DMEM
(Biological Industries; Sartorius AG) supplemented with 10% FBS
(Biological Industries; Sartorius AG) in a 37°C incubator with 5%
CO2.
In vitro cell model for CS-induced
malignant transformation
The BEAS-2B cells were used to establish an in
vitro model of CS-induced malignant transformation, the
detailed procedure of which has been described previously (7). Briefly, aliquots of exponentially
growing BEAS-2B cells (1×105) were plated onto a
Transwell membrane (0.4 µm pore; Corning, Inc.). An automatic
smoking machine was used to produce CS, which was then pumped into
an inhalation chamber where the BEAS-2B cells were directly exposed
to CS for 10 min every other day at a smoke concentration of 20%.
This procedure of exposing cells to CS was continued for 10, 20 and
30 passages, and such cells were referred to as experimental S10,
S20 and S30 cells, respectively. BEAS-2B cells that were not
exposed to CS were used as the control cells.
Transfection
Aliquots containing 2×105 BEAS-2B cells
were seeded in each well of 6-well plates and cultured for 24 h.
Then, the cells were transfected with: i) 50 nM mimic of miR-200b
(UAAUACUGCCUGGUAAUGAUGA); ii) mimic negative control (NC,
UUCUCCGAACGUGUCA); iii) 100 nM inhibitor of miR-200b
(UCAUCAUUACCAGGCGGUAUUA); or iv) 5-carboxyfluorescein (FAM) labeled
inhibitor NC (CAGUACUUUUGUGUAGUACAA). The four oligos were obtained
from Guangzhou RiboBio Co., Ltd. Transfection was performed using
Lipofectamine® 6000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol.
Transfection efficiency was evaluated by reverse
transcription-quantitative (RT-q)PCR and fluorescence of
FAM-inhibitor NC transfected cells. The cells were harvested 24 h
after transfection and used for wound healing and Transwell assays,
as well as for RNA and protein extraction, as described below.
Wound healing assay
Aliquots of 2×105 exponentially growing
mimic- and inhibitor-transfected cells were seeded into separate
6-well plates and cultured under standard conditions until they
reached 100% confluency. Subsequently, a scratch was made on each
plate using a P10 pipette tip. The culture medium was replaced with
fresh serum-free DMEM and the cells were cultured further. Images
of the wound were obtained at 0 and 24 h using an inverted
microscope (magnification, ×200; Olympus Corporation) and
quantitated using ImageJ version 1.8.0 (National Institutes of
Health).
Transwell migration assay
Aliquots of exponentially growing transfected cells
(2×105 cells in 500 µl serum-free medium) were seeded
into the upper chambers of the Transwell inserts (Corning, Inc.),
whereas the lower chambers were filled with 1 ml complete medium
with 10% FBS. All plates were incubated at 37°C for 24 h.
Subsequently, the cells on the membrane of the upper chamber were
removed with a cotton swab. Cells in the lower chamber were fixed
with 4% paraformaldehyde at room temperature for 15 min and stained
with 0.5% crystal violet solution at room temperature for 15 min.
Images of the cells were taken using an inverted microscope
(magnification, ×200; Olympus Corporation) and analyzed using
ImageJ version 1.8.0 (National Institutes of Health).
RNA extraction and RT-qPCR
Total RNA was extracted using TRIzol®
reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions from cell samples. From each sample,
~1.5 µg RNA was reverse transcribed into cDNA using a RevertAid
First Strand cDNA Synthesis Kit for mRNA detection (Thermo Fisher
Scientific, Inc.), or a Mir-X™ miRNA First-Strand Synthesis kit for
miRNA detection (Clontech Laboratories, Inc.) according to the
manufacturer's instructions. qPCR was performed using a
NovoScript® SYBR Two-Step RT-qPCR kit (Novoprotein
Scientific, Inc.) on a QuantStudio™ 6 Flex RT-qPCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). Thermocycling
conditions were initial hold at 95°C for 10 min, 40 cycles of 95°C
for 15 sec and 60°C for 60 sec. The sequences of the primer pairs
used for qPCR are listed in Table
SI. GAPDH and U6 were used as the internal controls. The
relative expression of each target was analyzed using the
2−ΔΔCq method (29).
Protein extraction and western blot
analysis
Exponentially growing cells were washed with
ice-cold PBS and lysed in ice-cold RIPA buffer (Beyotime Institute
of Biotechnology) for protein extraction. Total protein was
quantified using BCA reagent (Beyotime Institute of Biotechnology).
Then, ~30 µg of each protein sample was mixed with a quarter volume
of 5X loading buffer and boiled at 95°C for 10 min. The lysates
were loaded on a 10% sodium dodecyl sulfate (SDS)-gel, resolved
using SDS-polyacrylamide gel electrophoresis, and subsequently the
proteins were transferred to PVDF membranes (EMD Millipore). The
membranes were blocked using TBS with 0.1% Tween-20 (TBST) buffer
containing 5% BSA (Beyotime Institute of Biotechnology) at room
temperature for 2 h, and then probed with specific primary
antibodies overnight at 4°C. After washing with TBST, membranes
were incubated at room temperature for 2 h with mouse or rabbit
anti-rabbit IgG antibody. The antibodies used in this experiment
were: Epithelial marker, E-cadherin (CDH1; 1:1,000; cat. no.
20874-1-AP; ProteinTech Group, Inc.), mesenchymal marker,
N-cadherin (CDH2; 1:1,000; cat. no. 22018-1-AP; ProteinTech Group,
Inc.), GAPDH (1:1,000; cat. no. 5174; Cell Signaling Technology,
Inc.), horseradish peroxidase (HRP)-conjugated anti-rabbit IgG
secondary antibody (1:20,000; cat. no. ab7090; Abcam) and
HRP-tagged anti-mouse IgG secondary antibody (1:3,000; cat. no.
7076; Cell Signaling Technology, Inc.). Signals were visualized
using enhanced chemiluminescence (ECL) plus reagents (Invitrogen;
Thermo Fisher Scientific, Inc.) and a GeneTools GBox system
(Syngene Europe). The intensity of each band was semi-quantified
using ImageJ.
Dual-luciferase reporter assays
To verify whether ETS1 was a direct target of
miR-200b, the complete sequence of the ETS1 wild-type 3′-UTR
(ETS1_UTR_WT) was amplified by PCR from human genomic DNA using the
forward primer, 5′-GCTCTAGAGCTATCACTCTAGTTTTGAAGC-3′ and reverse
primer, 5′-GCTCTAGAGCCTTTCATTGTGACAGAATCC-3′. Both primers
contained the recognition sequence for the XbaI restriction
enzyme at the 5′ ends. Subsequently, the sequence was cloned into
the pGL3 (cat. no. 48743; Addgene) vector downstream of the
luciferase open reading frame. Additionally, the site-directed
mutagenesis for ETS1 3′-UTR was performed using a
site-directed mutagenesis kit (Beyotime Institute of Biotechnology)
to remove the miR-200b binding site.
Aliquots of 2×105 exponentially growing
BEAS-2B cells were seeded into 24-well plates and co-transfected
with the mimics (miR-200b) and the constructed pGL3 plasmids, as
well as the Renilla luciferase plasmid (pRL, Addgene, cat. no.
27163, which was used as the internal control) at a ratio of 5:5:1
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). Lysates were collected 48 h after transfection.
Firefly and Renilla luciferase activities were measured
using a Firefly luciferase Reporter Gene Assay kit (Beyotime
Institute of Biotechnology).
Target gene prediction
The target genes of miRNAs were predicted using
miRDB (mirdb.org/miRDB/), miRTarBase
(mirtarbase.cuhk.edu.cn/php/index.php) and miRanda
(microrna.org/) online analysis tools. To further
enhance the reliability of bioinformatics analysis, the overlapping
target genes were identified using a Venn diagram generated with
VennDiagram package (version: 1.6.20) in R (version: 4.0.4).
ETS1 plasmid construction and rescue
testing
The complete coding sequence of ETS1 was
amplified by PCR from human genomic DNA using the forward primer,
5′- GGAATTCC GCCACCATGAGCTACTTTGTGGATTCT-3′ and reverse primer, 5′-
ACGCGTCGACTCACTCGTCGGCATCTGG CTT-3′. The ETS1 plasmid was
constructed by inserting the CDS into a mammalian expression
vector, pCDH-EF1-copGFP (Addgene, Inc.), which contained a CMV
promoter driving the expression of GFP and ETS1. The negative
control used in this study was the empty vector. A total of 2.5 µg
pCDH-EF1-copGFP-ETS1 (pCDH ETS1) or empty vector (pCDH Blank) was
transfected into BEAS-2B cells (105 cells per well of
6-well plate) to rescue the low-expression induced by mimic
transfection. Transfection was performed using
Lipofectamine® 6000 reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. After
48 h, transfected cells were harvested for further detection.
Statistical analysis
All results are presented as the mean ± standard
deviation (n=3 replicates for each experiment) and were analyzed
using SPSS version 22.0 (IBM Corp.). Histograms were plotted using
GraphPad Prism version 7 (GraphPad Software, Inc.). Differences
between groups were compared using an unpaired Student's t-test or
a one-way ANOVA with a Tukey's post hoc test where appropriate.
P<0.05 was considered to indicate a statistically significant
difference.
Results
miR-200b expression is upregulated in
CS-exposed cells and lung cancer cell lines
Our previous study identified several differentially
expressed miRNAs, including miR-200b-3p (miR-200b), based on the
miRNA sequencing results (8)
(Fig. 1A). The results from RT-qPCR
analysis indicated that miR-200b was significantly upregulated in
S10, S20 and S30 cells (Fig. 1B),
as well as in human lung cancer cells, including A549, H1299 and
H226 (Fig. 1C). Next, the
expression of miR-200b in six lung cancer datasets was analyzed.
The results showed that miR-200b was significantly increased in
lung cancer tissues when compared with normal tissues (Fig. 1D-I).
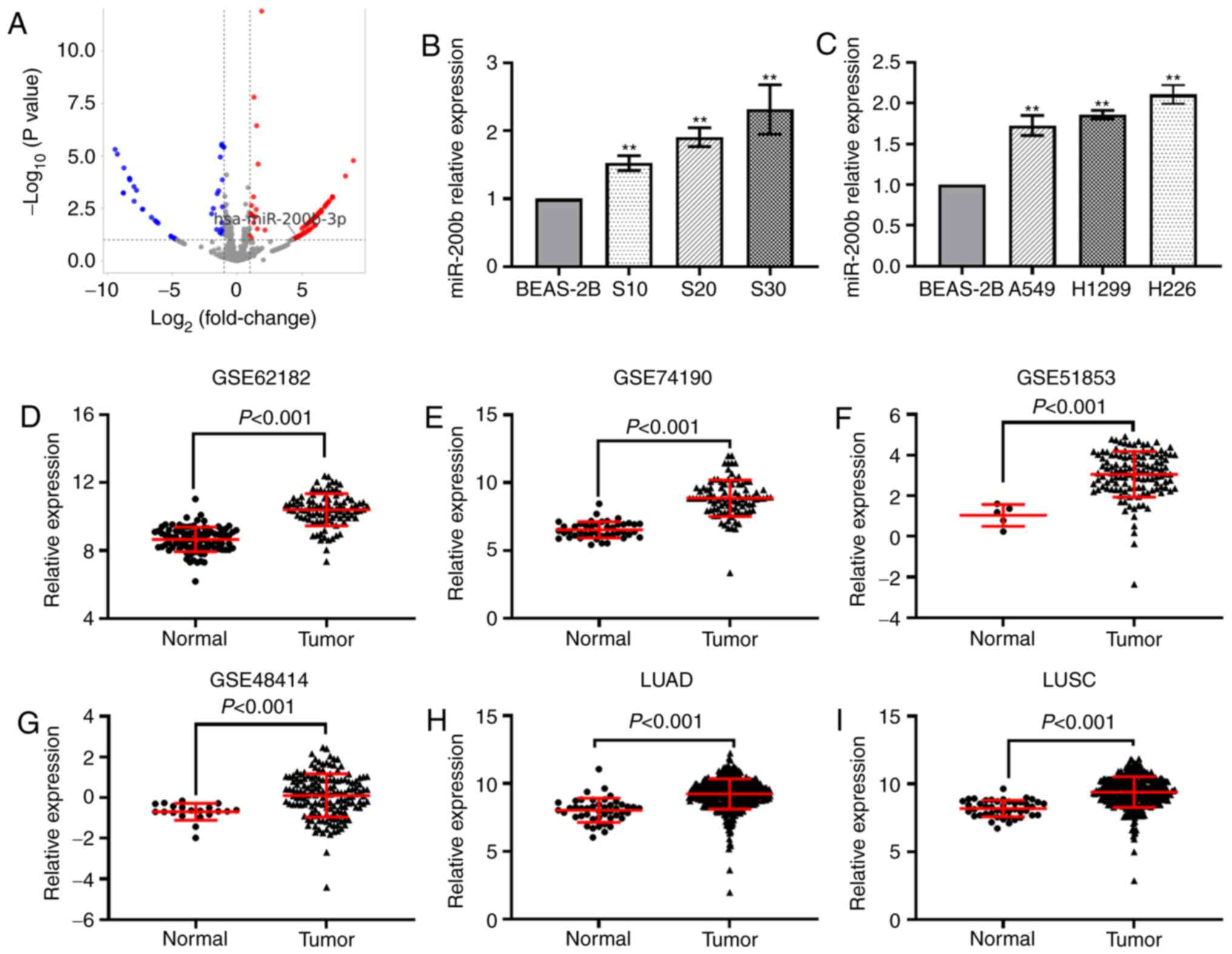 | Figure 1.miR-200b is upregulated in CS-exposed
cells and lung cancer. (A) Reverse transcription-quantitative PCR
analysis of miR-200b in (B) CS-exposed BEAS-2B cells and (C) lung
cancer cells. Differential analysis of miR-200b expression in six
datasets, (D) GSE62182, (E) GSE74190, (F) GSE51853, (G) GSE48414,
as well as in the (H) LUAD and (I) LUSC datasets in The Cancer
Genome Atlas. **P<0.01 vs. normal BEAS-2B cells. miR, microRNA;
CS, cigarette smoke; qPCR, quantitative PCR; LUAD, lung
adenocarcinoma; LUSC, lung squamous cell carcinoma. |
miR-200b promotes migration of lung
cancer cells
Using the LinkedOmics online tool, positively and
negatively correlated genes of miR-200b-5p in LUAD and LUSC were
obtained (Fig. S1A and B). A total
of 380 and 818 overlapping genes were significantly positively and
negatively correlated with miR-200b in both LUAD and LUSC datasets,
respectively (Fig. S1C and D).
Additionally, further enrichment analysis demonstrated that these
correlated genes were significantly enriched in several
cancer-related signaling pathways, particularly cell
migration-related pathways, including ‘Focal adhesion’,
‘ECM-receptor interaction’, ‘Cell adhesion molecules’ and ‘NF-kappa
B’ and ‘Jak-STAT’ signaling pathways (Fig. S1E). Transfection efficiency was
evaluated by RT-qPCR and FAM-inhibitor NC. RT-qPCR results revealed
that miR-200b-5p levels were elevated significantly in the
mimic-transfected cells, compared with mimic NC-transfected cells
(Fig. S2A). Fluorescent imaging
demonstrated that the FAM-inhibitor NC became abundant within the
cells, suggesting that the miRNA oligos used in this study were
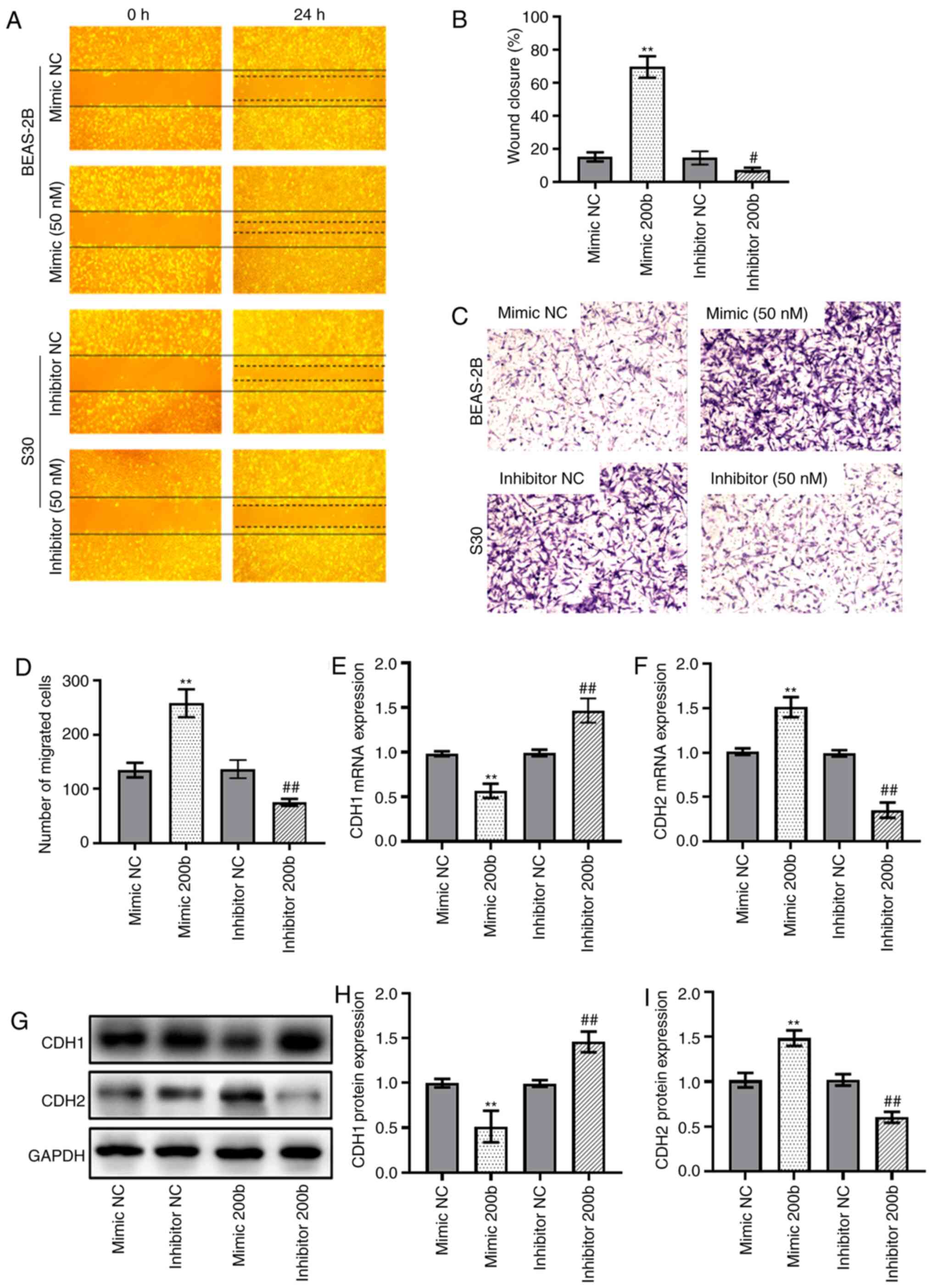
successfully transfected into BEAS-2B cells s(Fig. S2B). The results from the Transwell
and wound-healing assays showed that transfection of cells with
miR-200b mimic enhanced the migration of BEAS-2B cells. Conversely,
transfection with the inhibitor of miR-200b significantly reduced
the migratory capability of BEAS-2B cells (Fig. 2A-D). The results also showed that
the mRNA expression levels of CDH1 were downregulated in cells
transfected with miR-200b mimic, whereas they were upregulated in
cells transfected with the miR-200b inhibitor (Fig. 2E). By contrast, CDH2 mRNA expression
was upregulated in cells transfected with miR-200b mimic and
downregulated in cells transfected with miR-200b inhibitors
(Fig. 2F). The changes in protein
levels of CDH1 and CDH2 in the transfected cells were consistent
with the changes in mRNA expression (Fig. 2G-I).
ETS1 is a target of miR-200b in lung
cancer cells
Using the TargetScan, miRanda, miRWalk and
miRTarBase online tools, a total of six intersected target genes of
miR-200b were obtained, including ETS1, ZFPM2, RND3, E2F3,
QKI and XIAP (Fig. 3A).
The pathological analysis indicated that in TCGA LUAD dataset,
ETS1 expression was downregulated in lung cancer tissues at
advanced invasion depth (T2) compared with tissues at stage T1
(Fig. 3B). The decreased expression
of ETS1 in lung cancer tissues of patients was found in TCGA
LUAD (Fig. 3C), TCGA LUSC (Fig. 3D), GSE19804 (Fig. 3E), and GSE27262 (Fig. 3F) datasets, as well as in the
proteomics dataset, CPTAC_LUAD (Fig.
3G). Additionally, the RT-qPCR and western blot analysis showed
significant downregulation of ETS1 in S20 and S30 cells
(Fig. 3H and I).
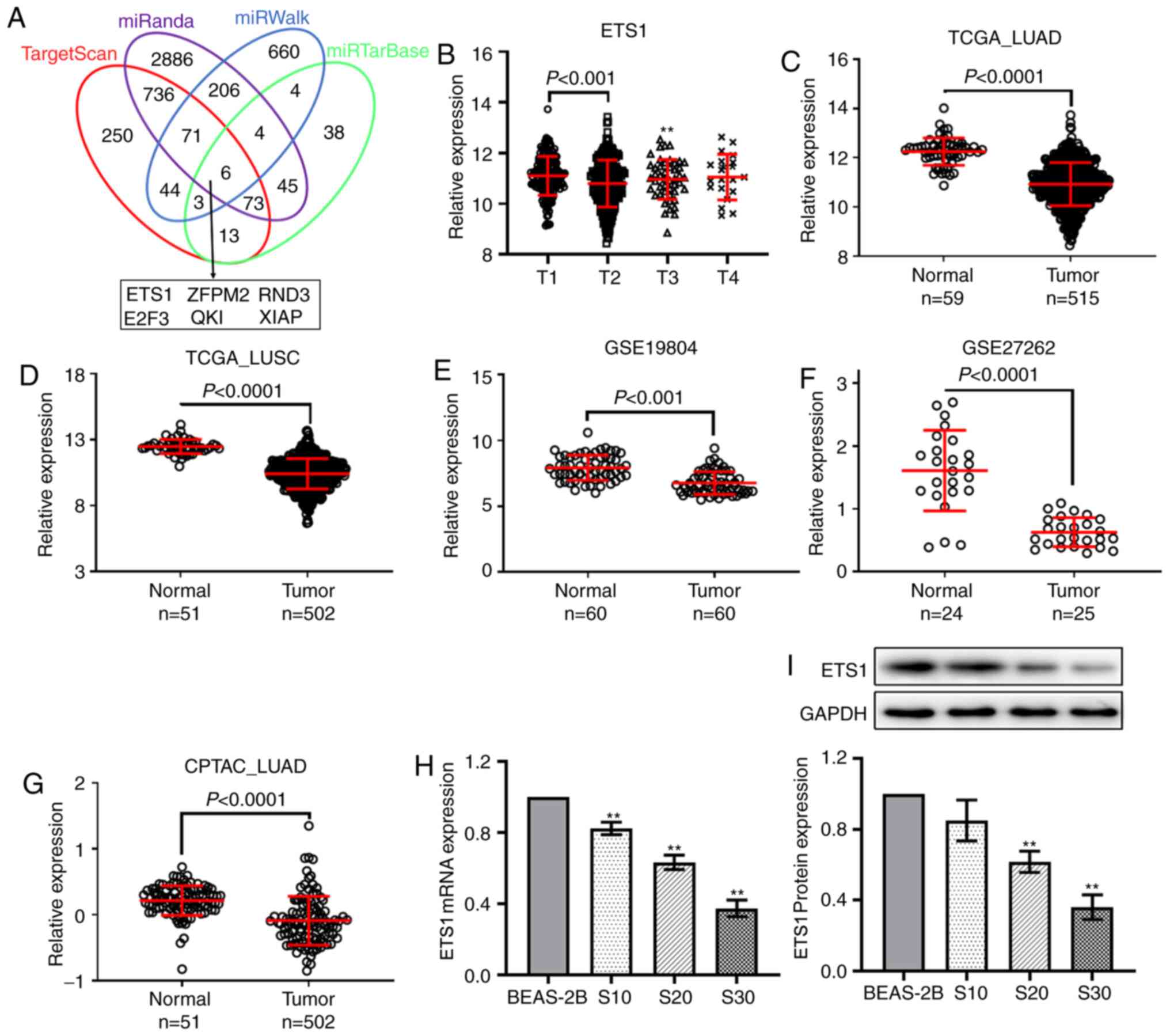 | Figure 3.ETS1 is a downstream target of
miR-200b. (A) Venn diagram showing the intersection of the target
genes predicted by four online tools, TargetScan, miRanda, miRWalk
and miRTarBase. (B) In TCGA LUAD dataset, ETS1 was
downregulated in lung cancer tissues at stage T2 compared with
tissues at stage T1. Differential analysis of ETS1
expression in four datasets, (C) TCGA LUAD, (D) TCGA LUSC, (E)
GSE19804 and (F) GSE27262, as well as the (G) CPTAC LUAD proteomics
dataset. (H) Reverse transcription-quantitative PCR analysis showed
ETS1 expression was downregulated in CS-exposed cells. (I)
Western blotting results showed ETS1 protein expression was
downregulated in CS-exposed cells. **P<0.01 vs. normal BEAS-2B
cells. TCGA, The Cancer Gene Atlas; CPTAC, Clinical Proteomic Tumor
Analysis Consortium; LUAD, lung adenocarcinoma; LUSC, lung squamous
cell carcinoma; CS, cigarette smoke; miR, microRNA; ETS1, ETS
proto-oncogene 1 transcription factor; ZFPM2, zinc finger protein
ZFPM2; RND3, rho-related GTP-binding protein RhoE; E2F3,
transcription factor E2F3; QKI, protein quaking; XIAP, E3
ubiquitin-protein ligase XIAP. |
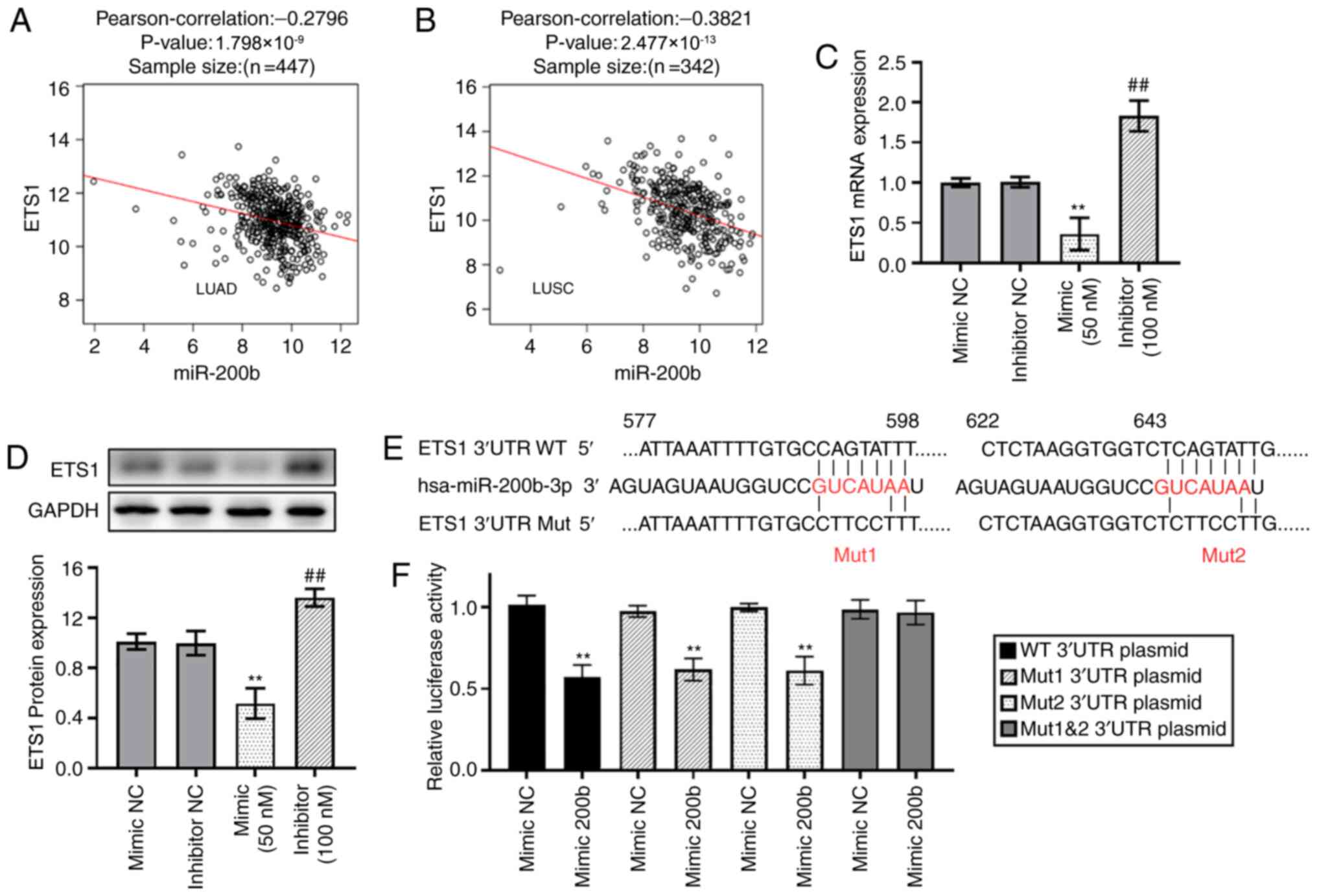
In addition, correlation analysis revealed that the
expression levels of miR-200b were weakly and moderately negatively
correlated with ETS1 mRNA expression in TCGA LUAD (Fig. 4A) and TCGA LUSC (Fig. 4B) datasets respectively. The mRNA
and protein expression levels of ETS1 were significantly
downregulated in cells transfected with miR-200b mimics, whereas
expression was upregulated in cells transfected with inhibitors
(Fig. 4C and D). As shown in
Fig. 4E, two binding sites for
miR-200b were present in the 3′UTR of ETS1. The relative
luciferase activity of the reporter gene in BEAS-2B cells
co-transfected with pGL3-ETS1 WT 3′UTR and miR-200b mimic was
significantly decreased compared with the control (co-transfected
with pGL3-ETS1 3′UTR and mimic NC). Mutating one of the two binding
sites alone can still result in downregulation of relative
luciferase activity when compared with mimic NC groups, whereas
mutating both sites simultaneously avoids this downregulation
(Fig. 4F).
ETS1 mediates the pro-migration effect
of miR-200b
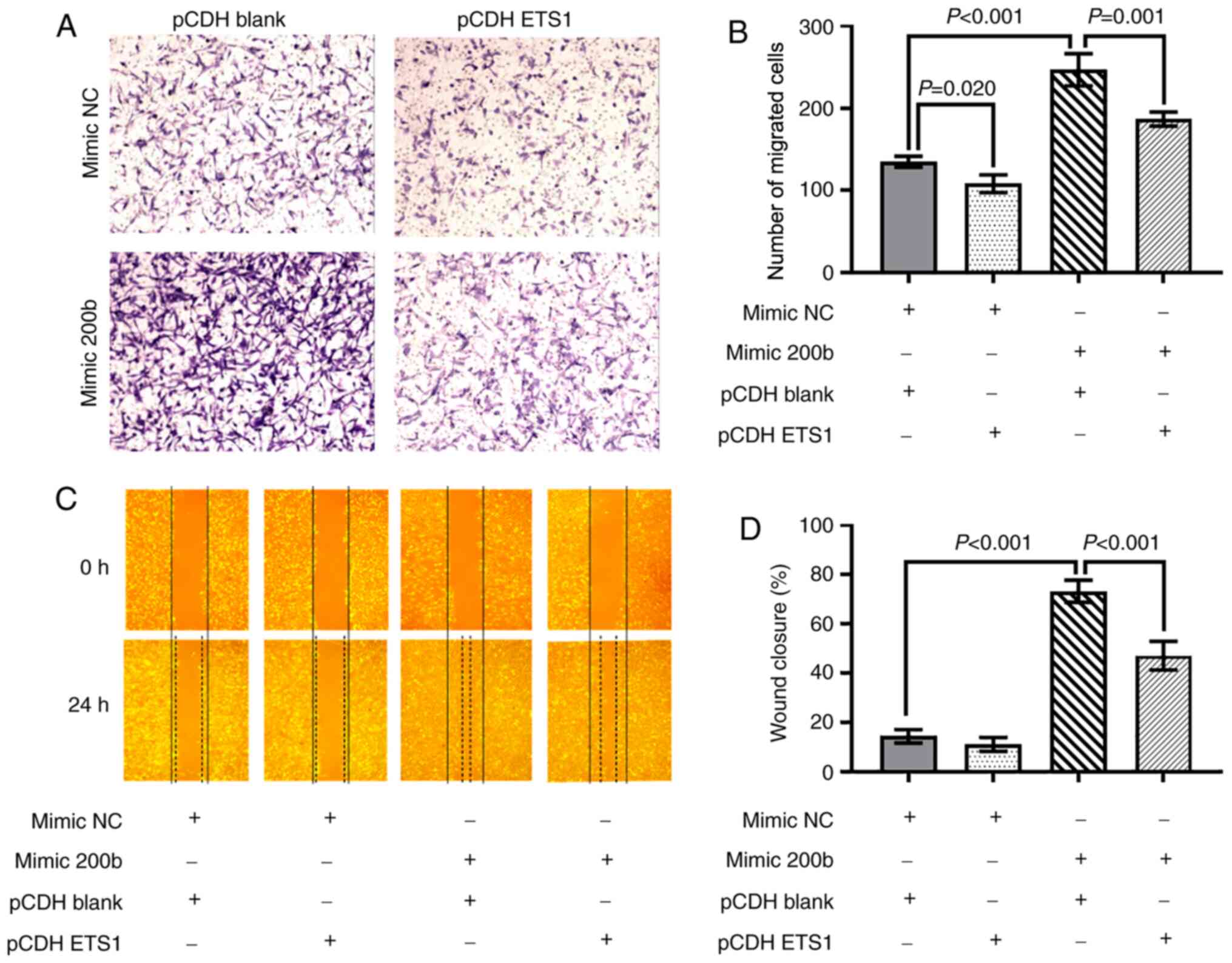
To further confirm that the effects of miR-200b were
mediated by repression of ETS1 in BEAS-2B cells, rescue
experiments in which ETS1 was overexpressed were performed. The
transfection efficiency of ETS1 overexpression vectors was verified
by RT-qPCR (Fig. S2C) and western
blot (Fig. S2D and E). The
enhanced migratory ability of BEAS-2B cells induced by miR-200b
mimic was reduced by transfection of the ETS1 overexpression
vector. Compared with the cells transfected with mimic NC and blank
vector, BEAS-2B cells transfected with miR-200b mimic and blank
vector exhibited increased migration ability. However, the enhanced
migratory ability induced by the mimic miR-200b was downregulated
by co-transfection with the ETS1 vector (Fig. 5A and B). In the wound healing
assays, a similar trend was observed (Fig. 5C and D).
Discussion
Previously, it has been reported that certain miRNAs
directly activate EMT transcription factors, whereas others are
able to reverse the EMT process by targeting several signaling
pathways (10). In the present
study, miR-200b was demonstrated to be involved in the migration
and EMT process of lung cancer cells. miR-200b is a member of the
miR-200 family and has been reported to serve contrasting roles in
different types of cancer. Studies reported that its expression is
downregulated in several malignant tumors, exhibiting a
tumor-suppressing role (30–33).
For example, miR-200b was identified as a repressor of invasiveness
in esophageal squamous cell carcinoma by modulating multiple key
cell cycle regulators and the Wnt/β-catenin signaling pathways
(34). Conversely, miR-200b has
also been found to promote cellular proliferation and serve a
tumor-promoting role in several other tumors, such as cervical
cancer and colorectal cancer, as well as in lung cancer (20,35–37).
It has been reported that miRNA dysregulation may serve as a
biomarker of damage caused by acute and chronic environmental
exposure (38). Our previous miRNA
high throughput sequencing analysis identified several
differentially expressed miRNAs in the CS-induced malignantly
transformed BEAS-2B cells (9). In
the present study, elevated miR-200b levels were found in the
CS-induced malignant BEAS-2B cells and several lung cancer cell
lines, which was corroborated by analysis of external lung cancer
tissue databases. Furthermore, it was demonstrated that miR-200b
overexpression significantly enhanced the cellular migratory
ability and EMT of cells as demonstrated by the reduction in CDH1
expression and increase in CDH2 expression levels. Importantly,
these alterations in cell behavior have also been demonstrated in
CS-induced malignantly transformed cells (9).
ETS1 is a 54-kDa nuclear protein that serves
a major role in the regulation of transcription factors (39). Transcription factors of the ETS
family are involved in normal cell development, proliferation and
differentiation (40). Previously,
studies have found multiple miRNAs that downregulate ETS1
expression by directly targeting the 3′UTR region of ETS1
(41). For example, in human
hepatocellular carcinoma cells, miR-1, miR-129-5p, miR-193b and
miR-499 can induce the downregulation of ETS1 expression, and thus
reduce migration and invasion (42–44).
Similar to a previous study (45), it was presented in the present study
that ETS1 was a target gene of miR-200b, through multiple
target gene prediction databases. The dual-luciferase reporter
assay confirmed this targeting regulatory relationship between
miR-200b and ETS1. However, multiple aspects distinguish the
present study from previously published literature. In the present
study, not only did we show the downregulation of ETS1
expression in lung cancer cell lines in vitro and in tumor
tissue samples, but we also further confirmed this relationship
using a controlled model of long-term CS-exposed cells. Moreover,
in addition to establishing the importance of the miR-200b-ETS1
cascade in elevating the migratory ability of cells, we further
showed a reversal of the miR-200b-induced increase in migration
when ETS1 overexpression plasmids were transfected into the
miR-200b overexpressing BEAS-2B cells. Similarly, this increase in
migratory ability was dampened by transfection of miR-200b
inhibitors.
The present study had some limitations that are
worth mentioning. First, only one cell line, the immortalized
bronchial epithelial cell BEAS-2B, was used to investigate the role
of miR-200b in promoting migration, while no lung cancer cell lines
were used for comparative analysis. The present study also lacks
the necessary animal experiments for miR-200b function. Further
studies will shed more light on its function with more in
vitro or in vivo experiments.
In summary, the present study demonstrated the
upregulation of miR-200b in CS-exposed BEAS-2B cells, lung cancer
cell lines and tumor tissue samples. Downstream, miR-200b was shown
to serve a carcinogenic role by targeting the 3′-UTR of
ETS1, in-turn promoting cell migration and EMT. These
insights into the miR-200b-ETS1 axis may facilitate the
development of treatments for CS-induced lung cancer. With the
development of improved delivery vehicles for these therapeutics,
we hypothesize that miRNA therapies will soon become a clinical
reality in the treatment of cancer.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable.
Funding
This study was supported by the National Natural
Science Foundation of China (grant no. 81573178), Jiangsu Key
Laboratory of Preventive and Translational Medicine for Geriatric
Diseases, the Priority Academic Program Development of Jiangsu
Higher Education Institutions (PAPD) and Postgraduate Research
& Practice Innovation Program of Jiangsu Province (grant no.
KYCX20_2681).
Availability of data and materials
The TCGA LUAD and LUSC datasets analyzed during the
current study are available in the UCSC Xena repository,
[https://xenabrowser.net/datapages/].
The datasets analyzed during the current study are available in the
GEO repository, [https://ncbi.nlm.nih.gov/geo/].
Authors' contributions
JL and JW conceived and designed the study. RY, JW
and YL retrieved and downloaded the datasets. JW, RY, QL and YL
analyzed the datasets. RY, QL, LT, JT, BJ and NO performed the
experiments and analyzed the data. JL and JW confirm the
authenticity of all the raw data. JW and RY drafted the manuscript.
QL, JL, YL and JT edited the manuscript. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hirsch FR, Scagliotti GV, Mulshine JL,
Kwon R, Curran WJ Jr, Wu YL and Paz-Ares L: Lung cancer: Current
therapies and new targeted treatments. Lancet. 389:299–311. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Goebel C, Louden CL, McKenna R Jr, Onugha
O, Wachtel A and Long T: Diagnosis of non-small cell lung cancer
for early stage asymptomatic patients. Cancer Genomics Proteomics.
16:229–244. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hay ED: An overview of
epithelio-mesenchymal transformation. Acta Anat (Basel). 154:8–20.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lamouille S, Xu J and Derynck R: Molecular
mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell
Biol. 15:178–196. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Du H, Sun J, Chen Z, Nie J, Tong J and Li
J: Cigarette smoke-induced failure of apoptosis resulting in
enhanced neoplastic transformation in human bronchial epithelial
cells. J Toxicol Environ Health A. 75:707–720. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Huang H, Ji Y, Zhang J, Su Z, Liu M, Tong
J, Ge C, Chen T and Li J: Aberrant DNA methylation in radon and/or
cigarette smoke-induced malignant transformation in BEAS-2B human
lung cell line. J Toxicol Environ Health A. 80:1321–1330. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wang J, Yu XF, Ouyang N, Zhao S, Yao H,
Guan X, Tong J, Chen T and Li JX: MicroRNA and mRNA Interaction
network regulates the malignant transformation of human bronchial
epithelial cells induced by Cigarette smoke. Front Oncol.
9:10292019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zaravinos A: The regulatory role of
MicroRNAs in EMT and cancer. J Oncol. 2015:8658162015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gros J and Tabin CJ: Vertebrate limb bud
formation is initiated by localized epithelial-to-mesenchymal
transition. Science. 343:1253–1256. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Correa-Costa M, Andrade-Oliveira V, Braga
TT, Castoldi A, Aguiar CF, Origassa CS, Rodas AC, Hiyane MI,
Malheiros DM, Rios FJ, et al: Activation of platelet-activating
factor receptor exacerbates renal inflammation and promotes
fibrosis. Lab Invest. 94:455–466. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Grande MT, Sánchez-Laorden B, López-Blau
C, De Frutos CA, Boutet A, Arévalo M, Rowe RG, Weiss SJ,
López-Novoa JM and Nieto MA: Snail1-induced partial
epithelial-to-mesenchymal transition drives renal fibrosis in mice
and can be targeted to reverse established disease. Nat Med.
21:989–997. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dhayat SA, Mardin WA, Köhler G, Bahde R,
Vowinkel T, Wolters H, Senninger N, Haier J and Mees ST: The
microRNA-200 family - a potential diagnostic marker in
hepatocellular carcinoma? J Surg Oncol. 110:430–438. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hur K, Toiyama Y, Takahashi M, Balaguer F,
Nagasaka T, Koike J, Hemmi H, Koi M, Boland CR and Goel A:
MicroRNA-200c modulates epithelial-to-mesenchymal transition (EMT)
in human colorectal cancer metastasis. Gut. 62:1315–1326. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bojmar L, Karlsson E, Ellegård S, Olsson
H, Björnsson B, Hallböök O, Larsson M, Stål O and Sandström P: The
role of microRNA-200 in progression of human colorectal and breast
cancer. PLoS One. 8:e848152013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kan CW, Hahn MA, Gard GB, Maidens J, Huh
JY, Marsh DJ and Howell VM: Elevated levels of circulating
microRNA-200 family members correlate with serous epithelial
ovarian cancer. BMC Cancer. 12:6272012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pacurari M, Addison JB, Bondalapati N, Wan
YW, Luo D, Qian Y, Castranova V, Ivanov AV and Guo NL: The
microRNA-200 family targets multiple non-small cell lung cancer
prognostic markers in H1299 cells and BEAS-2B cells. Int J Oncol.
43:548–560. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Feng B, Wang R and Chen LB: Review of
miR-200b and cancer chemosensitivity. Biomed Pharmacother.
66:397–402. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu K, Zhang W, Tan J, Ma J and Zhao J:
miR-200b-3p functions as an oncogene by targeting ABCA1 in lung
adenocarcinoma. Technol Cancer Res Treat. 18:15330338198925902019.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Goldman MJ, Craft B, Hastie M, Repečka K,
McDade F, Kamath A, Banerjee A, Luo Y, Rogers D, Brooks AN, et al:
Visualizing and interpreting cancer genomics data via the Xena
platform. Nat Biotechnol. 38:675–678. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Vucic EA, Thu KL, Pikor LA, Enfield KS,
Yee J, English JC, MacAulay CE, Lam S, Jurisica I and Lam WL:
Smoking status impacts microRNA mediated prognosis and lung
adenocarcinoma biology. BMC Cancer. 14:7782014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Huang W, Jin Y, Yuan Y, Bai C, Wu Y, Zhu H
and Lu S: Validation and target gene screening of hsa-miR-205 in
lung squamous cell carcinoma. Chin Med J (Engl). 127:272–278.
2014.PubMed/NCBI
|
|
24
|
Arima C, Kajino T, Tamada Y, Imoto S,
Shimada Y, Nakatochi M, Suzuki M, Isomura H, Yatabe Y, Yamaguchi T,
et al: Lung adenocarcinoma subtypes definable by lung
development-related miRNA expression profiles in association with
clinicopathologic features. Carcinogenesis. 35:2224–2231. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bjaanaes MM, Halvorsen AR, Solberg S,
Jørgensen L, Dragani TA, Galvan A, Colombo F, Anderlini M,
Pastorino U, Kure E, et al: Unique microRNA-profiles in
EGFR-mutated lung adenocarcinomas. Int J Cancer. 135:1812–1821.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lu TP, Tsai MH, Lee JM, Hsu CP, Chen PC,
Lin CW, Shih JY, Yang PC, Hsiao CK, Lai LC, et al: Identification
of a novel biomarker, SEMA5A, for non-small cell lung carcinoma in
nonsmoking women. Cancer Epidemiol Biomarkers Prev. 19:2590–2597.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wei TY, Juan CC, Hisa JY, Su LJ, Lee YC,
Chou HY, Chen JM, Wu YC, Chiu SC, Hsu CP, et al: Protein arginine
methyltransferase 5 is a potential oncoprotein that upregulates G1
cyclins/cyclin-dependent kinases and the phosphoinositide
3-kinase/AKT signaling cascade. Cancer Sci. 103:1640–1650. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Whiteaker JR, Halusa GN, Hoofnagle AN,
Sharma V, MacLean B, Yan P, Wrobel JA, Kennedy J, Mani DR,
Zimmerman LJ, et al Clinical proteomic tumor analysis consortium
(CPTAC), : CPTAC assay portal: A repository of targeted proteomic
assays. Nat Methods. 11:703–704. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yao Y, Hu J, Shen Z, Yao R, Liu S, Li Y,
Cong H, Wang X, Qiu W and Yue L: MiR-200b expression in breast
cancer: A prognostic marker and act on cell proliferation and
apoptosis by targeting Sp1. J Cell Mol Med. 19:760–769. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shinozaki A, Sakatani T, Ushiku T, Hino R,
Isogai M, Ishikawa S, Uozaki H, Takada K and Fukayama M:
Downregulation of microRNA-200 in EBV-associated gastric carcinoma.
Cancer Res. 70:4719–4727. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
He M, Liu Y, Deng X, Qi S, Sun X, Liu G,
Liu Y, Liu Y and Zhao M: Down-regulation of miR-200b-3p by low p73
contributes to the androgen-independence of prostate cancer cells.
Prostate. 73:1048–1056. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Li Y, Guan B, Liu J, Zhang Z, He S, Zhan
Y, Su B, Han H, Zhang X, Wang B, et al: MicroRNA-200b is
downregulated and suppresses metastasis by targeting LAMA4 in renal
cell carcinoma. EBioMedicine. 44:439–451. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang HF, Alshareef A, Wu C, Jiao JW,
Sorensen PH, Lai R, Xu LY and Li EM: miR-200b induces cell cycle
arrest and represses cell growth in esophageal squamous cell
carcinoma. Carcinogenesis. 37:858–869. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Xing T, Chen Y and Xiao J:
Exosome-mediated miR-200b promotes colorectal cancer proliferation
upon TGF-β1 exposure. Biomed Pharmacother. 106:1135–1143. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zeng F, Xue M, Xiao T, Li Y, Xiao S, Jiang
B and Ren C: MiR-200b promotes the cell proliferation and
metastasis of cervical cancer by inhibiting FOXG1. Biomed
Pharmacother. 79:294–301. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Fu Y, Liu X, Zhou N, Du L, Sun Y, Zhang X
and Ge Y: MicroRNA-200b stimulates tumour growth in TGFBR2-null
colorectal cancers by negatively regulating p27/kip1. J Cell
Physiol. 229:772–782. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Vrijens K, Bollati V and Nawrot TS:
MicroRNAs as potential signatures of environmental exposure or
effect: A systematic review. Environ Health Perspect. 123:399–411.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang C, Kam RKT, Shi W, Xia Y, Chen X, Cao
Y, Sun J, Du Y, Lu G, Chen Z, et al: The proto-oncogene
transcription factor Ets1 regulates neural crest development
through histone deacetylase 1 to mediate output of bone
morphogenetic protein signaling. J Biol Chem. 290:21925–21938.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shaikhibrahim Z and Wernert N: ETS
transcription factors and prostate cancer: The role of the family
prototype ETS-1 (Review). Int J Oncol. 40:1748–1754.
2012.PubMed/NCBI
|
|
41
|
Dittmer J: The role of the transcription
factor Ets1 in carcinoma. Semin Cancer Biol. 35:20–38. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu C, Liu S, Fu H, Li S, Tie Y, Zhu J,
Xing R, Jin Y, Sun Z and Zheng X: MicroRNA-193b regulates
proliferation, migration and invasion in human hepatocellular
carcinoma cells. Eur J Cancer. 46:2828–2836. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Wei W, Hu Z, Fu H, Tie Y, Zhang H, Wu Y
and Zheng X: MicroRNA-1 and microRNA-499 downregulate the
expression of the ets1 proto-oncogene in HepG2 cells. Oncol Rep.
28:701–706. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Ma N, Chen F, Shen SL, Chen W, Chen LZ, Su
Q, Zhang LJ, Bi J, Zeng WT, Li W, et al: MicroRNA-129-5p inhibits
hepatocellular carcinoma cell metastasis and invasion via targeting
ETS1. Biochem Biophys Res Commun. 461:618–623. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Chan YC, Khanna S, Roy S and Sen CK:
miR-200b targets Ets-1 and is down-regulated by hypoxia to induce
angiogenic response of endothelial cells. J Biol Chem.
286:2047–2056. 2011. View Article : Google Scholar : PubMed/NCBI
|