Introduction
Osteoarthritis (OA), a known degenerative disease,
is a chronic joint condition that mainly causes degenerative
changes in articular cartilage and secondary osteogenesis, and is
characterized by fibrosis, rhagadia, ulcers and attrition of
articular cartilage due to a number of factors (1). The main pathological manifestations
of OA include, but are not limited to, gradual loss of
extracellular matrix (ECM) proteins and proliferation of synovial
cells. OA can simultaneously destroy the articular cartilage
surface and narrow the articular space. The development of
subchondral osteosclerosis and osteophytes may gradually lead to
dysfunction of the entire joint. In early-stage OA, the cartilage
surface remains intact (2). The
first changes occur in the molecular composition of the tissues and
the ECM, which are characterized by a transient chondrocyte
proliferative response and an increase in ECM protein synthesis,
including collagen type II (Col II) and aggrecan (2). With the development of OA, the
synthesis of degrading proteases increases, leading to increased
catabolic activity and decreased proteoglycan content.
Subsequently, Col II is degraded and its fragments stimulate
several proteins associated with the catabolic state, such as MMPs
and aggrecanases, members of the a disintegrin and
metalloproteinase with thrombospondin motifs family (ADAMTSs),
which are accompanied by increased expression levels of
inflammatory cytokines (IL-1 and TNF-α), stress and apoptosis
markers (caspases-3 and −9 and Bcl-2), and transcription factors
[runt-related transcription factor 2 (Runx2) and Sox9] (3–5).
Changes in articular cartilage composition and structure further
stimulate chondrocytes into producing more catabolic factors
involved in cartilage degradation. As the proteoglycan and collagen
network breaks down, the integrity of the cartilage is disrupted
(6). Subsequently, apoptosis of
articular chondrocytes occurs, ultimately leading to the complete
loss of articular cartilage (Fig.
1) (2,3). Articular chondral injury is often
associated with pain, swelling and joint stiffness, or even
irreversible damage in certain patients, particularly adolescents,
severely compromising their quality of life (7). Articular cartilage tissue is
primarily composed of a large amount of ECM with embedded
chondrocytes. As the articular cartilage is a special type of
connective tissue without blood vessels, lymphatic vessels or
nerves, its primary nutrient sources are the synovial fluid in the
joint cavity and the blood supply from the subchondral bone.
Furthermore, its regenerative capacity is limited. Generally,
damage of 4 cm in diameter cannot be self-repaired (8). Current treatments focus on methods
for reducing pain and delaying disease progression. Furthermore,
preserving joint function and maintaining the quality of life of
the patients has attracted significant attention (1). At present, certain treatments, such
as sodium hyaluronate injection, etanercept and infliximab, are
used to alleviate symptoms, but with limited efficacy (7). Clinically, methods for repairing
chondral injury in the knee joint include microfracture, mosaics,
osteochondral grafting and autogenous chondrocyte grafting
(8). Although certain techniques
have progressed in treating cartilage defects, biological methods
for cartilage regeneration are still faced with a major challenge,
which lies in the limited regenerative capacity of cartilage tissue
(9). It has been reported
(10) that a new technological
method, percutaneous osmotic therapy, does not cause damage to the
digestive system, while it ensures good concentration and
utilization of drugs by the target organs. Another type of therapy,
intestinal microbial therapy, may contribute to the treatment of
systemic inflammation to a certain extent by restoring the
intestinal flora (11). Moreover,
a previous study reported that the regulation of microRNA (miRNA)
expression can inhibit the destruction of articular cartilage
caused by OA (12). However, the
aforementioned approaches have certain limitations (10). Tissue engineering aims to restore,
maintain and improve tissue performance, and it obtains
biocompatible and biologically functional tissues to achieve tissue
and organ regeneration or repair by combining stem cells and
biomaterial scaffolds (10).
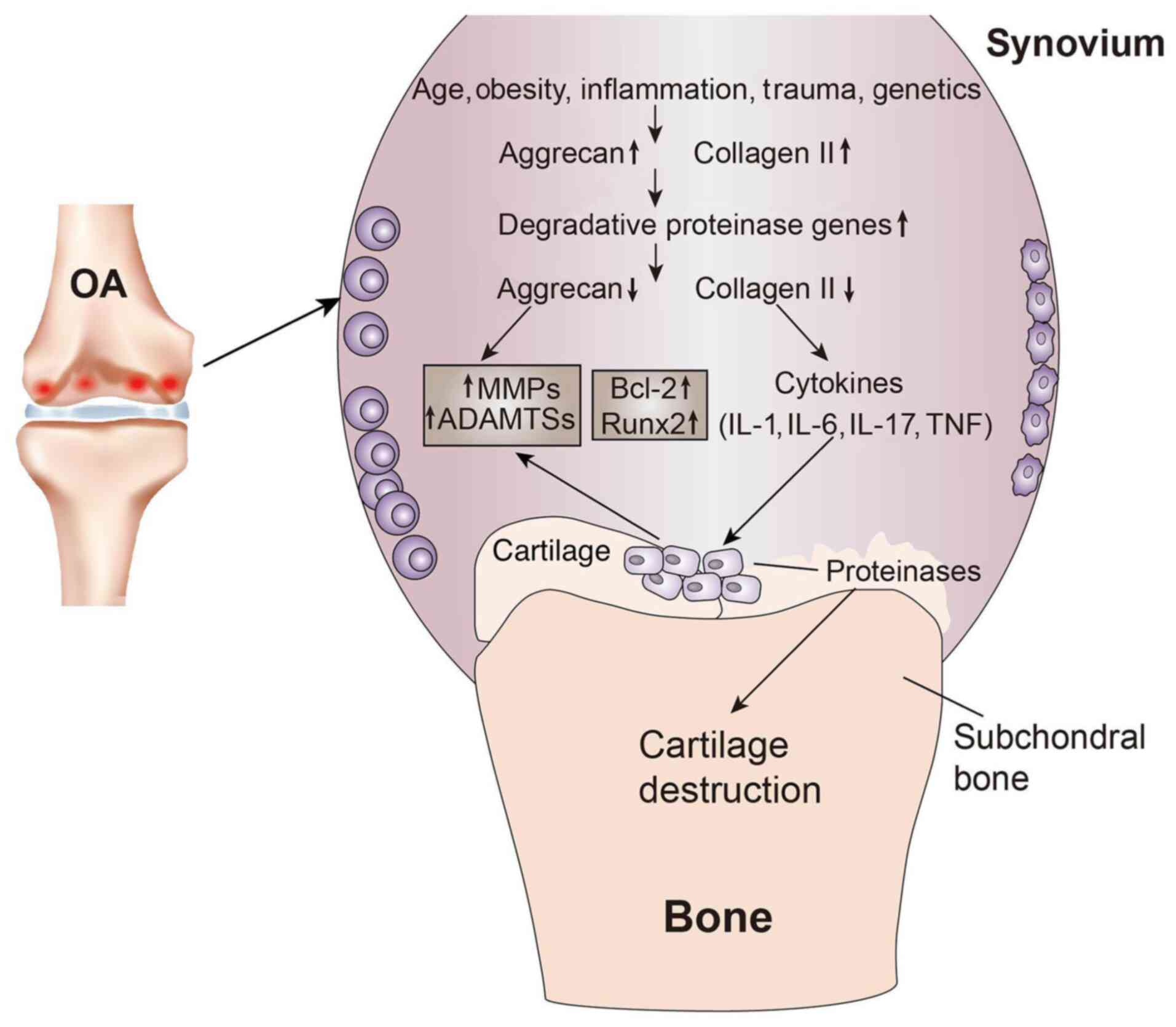 | Figure 1.Cartilage changes in OA. Age,
obesity, inflammation, trauma and genetics are common causes of OA.
In the early stages of OA, chondrocytes proliferate and further
extracellular matrix proteins, such as collagen type II, are
synthesized. As OA progresses, the gene expression of degradation
proteases increases, proteoglycans are gradually lost and,
subsequently, with collagen type II degradation, numerous proteins
associated with the catabolic state, such as MMPs and ADAMTSs, are
produced. With the production of proinflammatory cytokines (IL-1β
and TNF-α) and the increase in the expression of apoptotic markers
(Bcl-2 and Runx2) and transcription factors, the proinflammatory
cytokines produce more MMPs and ADAMTSs, as well as larger amounts
of proteases, which destroy the chondroprotein network, ultimately
leading to cartilage destruction. OA, osteoarthritis; ADAMTS, a
disintegrin and metalloproteinase with thrombospondin motif; Runx2,
runt-related transcription factor 2. |
Proinflammatory cytokines are endogenous
polypeptides that are primarily derived from immune system cells,
exert a variety of powerful biological effects and can mediate
various immune responses. Currently, the main proinflammatory
cytokines involved in OA are IL-1β, TNF-α, IL-6, IL-15, IL-17 and
IL-18 (13). The
anti-inflammatory cytokines in OA refer to cytokines that may
inhibit the actions of IL-1β and TNF-α, and any other cytokine that
primarily inhibits the cellular effects of the proinflammatory
cytokines in OA (14). At
present, the main anti-inflammatory cytokines in OA are IL-4,
insulin-like growth factor (IGF), IL-10 and TGF-β (15,16). During OA pathogenesis,
proinflammatory cytokines release inflammatory mediators by
regulating the NF-κB and MAPK signaling pathways, which triggers
the degeneration, destruction and degradation of articular
cartilage (17–19). Anti-inflammatory cytokines can
inhibit the actions of proinflammatory cytokines, downregulate the
expression levels of MMPs, inhibit inflammatory responses and lysis
of articular chondrocytes, promote chondrocyte proteoglycan and
collagen synthesis and delay OA progression (20). A possible approach for treating OA
is to block a single dominant inflammatory cytokine by employing a
similar strategy to target OA inflammatory signaling pathways and
maintaining their dynamic balance. Tissue engineering is also
critical for maintaining the balance between proinflammatory and
anti-inflammatory cytokines (21). Therefore, treating OA with a
combination of proinflammatory and anti-inflammatory cytokines,
along with tissue engineering, is a trending research topic in OA
treatment and it has highlighted a new treatment approach to this
disease. The aim of the present study was to therefore review the
role of combining proinflammatory and anti-inflammatory cytokines
in cartilage tissue engineering (CTE) for the treatment of OA.
Proinflammatory cytokines
As a group of compounds crucial for the pathogenesis
of OA, proinflammatory cytokines may cause the loss of articular
cartilage homeostasis via metabolic changes and markedly accelerate
joint damage (13). During
disease pathogenesis, these compounds affect most cells in the
joints and the production of cytokines and other inflammatory
compounds and enzymes via intracellular signal transduction
pathways. The most important proinflammatory cytokines are IL-1β,
TNF-α, IL-6, IL-15, IL-17 and IL-18 (13). How these compounds affect the
cells in the joints and the production of cytokines and other
inflammatory factors and enzymes is described in this section
(Table I).
 | Table I.Potential roles and origins of
proinflammatory cytokines. |
Table I.
Potential roles and origins of
proinflammatory cytokines.
| Cytokine | Source | Role | (Refs.) |
|---|
| IL-1β | Macrophages,
fibroblasts, chondrocytes, osteoblasts and osteoclasts | Potent inducer of
cartilage degradation and bone resorption | (16,17,20) |
| TNF-α | Macrophages,
fibroblasts and chondrocytes | Similar activity
profile to IL-1, but less potent | (30,39,40,44) |
| IL-6 | Macrophages,
fibroblasts, chondrocytes, osteoblasts and osteoclasts | IL-6 with Il-1β and
TNF-α have a synergistic effect, which may promote ECM degradation
of cartilage, leading to changes in subchondral bone | (57,59,61) |
| IL-15 | Macrophages,
fibroblasts and epidermal cells | Further activates
various cytokines, including TNF-α, IL-1, IL-6 and IL-17 | (62,69) |
| IL-17 | Macrophages | Promotes the
release of various proinflammatory cytokines and compounds,
including, IL-1β, TNF-α, NO and PGE2, as well as MMPs | (71,72,77) |
| IL-18 | Macrophages and
dendritic epithelial cells IFN-γ production and inhibit
angiogenesis | Recruits monocytes
and T lymphocytes to induce | (81,82,84) |
IL-1β
IL-1β is considered to be one of the key cytokines
involved in OA pathogenesis. It has been reported to cause
inflammatory responses and catabolic effects independently
(19). As one of the 11 members
of the IL-1 family (22), IL-1β
is primarily produced by activated type M1 mononuclear macrophages.
In patients with OA, IL-1β levels are elevated in the synovial
fluid, synovial membrane (SM), cartilage and subchondral bone
layers (23–26). IL-1β binds to its corresponding
receptors and promotes the release of proinflammatory cytokines,
such as IL-6 and IL-8, by regulating the NF-κB and MAPK signaling
pathways and increasing the MMP content in the matrices of
chondrocytes and fibroblast-like synoviocytes (FLS) (27). MMPs can irreversibly destroy
articular cartilage, bone and tendons. Among the MMPs, MMP-1 is
primarily expressed by synoviocytes in the joints and MMP-13 by
chondrocytes in cartilage. Both these MMPs can degrade collagen,
but MMP-13 can also degrade proteoglycan molecules and aggrecans,
which indicates MMP-13 serves a dual role in matrix degradation.
IL-1β also induces the expression of MMP-2, −3 and −9 in the
non-collagenous matrix components of the affected joint (28). Excluding MMP induction, IL-1β also
affects ADAMTS production in chondrocytes. ADAMTSs are responsible
for the proteolysis of aggregated proteoglycan molecules (29), with ADAMTS-4 being mainly
responsible for this effect (29,30). Moreover, the effects of IL-1β on
various enzymes and mediators secreted during the
pathophysiological process of OA have been observed. Relevant
compounds include inducible nitric oxide synthase (iNOS),
phospholipase A2, and cyclooxygenase-2 (COX-2) for nitric oxide
(NO) production, and prostaglandin E synthetase 2 for prostaglandin
(PG)E2 production (31,32). NO, as a free radical, promotes the
pathological progression of OA by inhibiting chondrocyte
proliferation and inducing chondrocyte apoptosis (33). Its catabolism in OA articular
cartilage is as follows: i) Inhibits Col II synthesis; ii) inhibits
proteoglycan synthesis; iii) inhibits IL-1 receptor antagonist (RA)
production; iv) interferes with integrin signals; v) induces
chondrocyte apoptosis; vi) stimulates MMP production and
activation; and vii) inactivates tissue inhibitor of
metalloproteinases (TIMP) (34).
PGE2, as one of the metabolites of arachidonic acid, has the
following actions: i) Absorbs subchondral bone; ii) affects bone
and cartilage anabolism; and iii) prevents the biochemical
synthesis of proteoglycans, leading to Col II destruction and
articular cartilage degeneration (35). Its catabolism in cartilage is
primarily achieved by activating MMPs. Furthermore, IL-1β
stimulates reactive oxygen species production, for example, the
formation of peroxides and hydroxylated radicals, and therefore
directly damages articular cartilage during disease progression. In
the affected joints, the intensification of this process is
reported to be associated with decreased oxidase expression
(36).
In summary, IL-1β binding to the IL-1R in SM cells,
activates transcription factors via the NF-κB and MAPK signaling
pathways to regulate inflammatory responses, resulting in the
production of inflammatory mediators in SM cells, such as MMPs,
COX-2, PGE2, NO and other catabolic factors, which may accelerate
cartilage degeneration. Therefore, blocking the aforementioned
signaling pathways mediated by inflammatory cytokines may be a
potential novel therapeutic strategy for treating OA.
TNF-α
Evidence suggests that TNF-α, one of the 19 ligands
in the TNF superfamily, is involved in the pathogenesis and
progression of OA (37). TNF-α is
also produced by activated type M1 mononuclear macrophages. It may
kill or inhibit tumor cells and promote the phagocytosis of
neutrophils (37). Its original
form is a type-II homotrimer transmembrane protein and under the
action of TNF convertase/ADAMTS17, free TNF-α is produced (38). This cytokine binds to two membrane
receptor isotypes, TNF-R1 (also known as p55, CD120a and TNF-R
super family member 1a) and TNF-R2 (also known as p75, CD120b and
TNF-R superfamily member 1b) on the surface of nearly all nucleated
cells (39,40). TNF-α binding to TNF-R1 leads to
the interaction of the TNFR1-associated death domain (DD) protein
adaptor protein with other adaptor proteins, such as TNF-R
associated factor 2, cellular inhibitor of apoptosis protein
(c-IAP)1, c-IAP2 and receptor-interacting protein (RIP)1 (41–43). The creation of the complex is
followed by the ubiquitination of the RIP1 protein, which also
binds to TGFβ-activated kinase (TAK)1, TAK1-binding protein (TAB)1
and TAB2, leading to phosphorylation of the IKK complex and
ultimately the activation of NF-κB, one of the most important
transcriptional signaling pathways in OA (44,45). TNF-α exerts its effects on
chondrocytes similar to IL-1β, including stimulating stromal
degradation proteases, NO and PGE2 production, as well as
inhibiting cartilage matrix synthesis (46–50). However, despite their similar
effects, IL-1β is 100- to 1,000-fold more potent compared with
TNF-α (51,52). The activity of these two cytokines
may produce a strong synergistic effect. For example,
intra-articular injection of recombinant IL-1 alone in rats, mice
and rabbits stimulates articular cartilage destruction.
Furthermore, when TNF-α and IL-1 are injected together, cartilage
damage may be far more severe compared with that caused by either
cytokine alone (53–55). Moreover, the effects of TNF-α
mediated via the PI3K/Akt axis may increase the expression levels
of cadherin-11, and this expression in FLS exhibits a positive
correlation with the severity of synovitis and cartilage
destruction (56). In an animal
experiment, Liu et al (57) demonstrated that TNF-α stimulation
increases cadherin-11 expression at the mRNA and protein levels in
OA FLS, whereby it also promotes the phosphorylation of Akt. The
intra-articular injection of anti-cadherin-11 antibody decreases
cadherin-11 expression in the SM and alleviates synovitis and
cartilage damage. These results indicated that TNF-α can induce
cadherin-11 expression and this protein may cause synovitis and
cartilage injury. Furthermore, cartilage injury may be treated by
inhibiting cadherin-11 expression, which may be achieved by
suppressing TNF-α expression (57).
In conclusion, free TNF-α binds to the corresponding
receptor and activates transcription factors via the NF-κB
signaling pathway to produce an inflammatory response, similar to
but weaker than IL-1β. These two cytokines can exert a synergistic
effect and further enhance the inflammatory response. Furthermore,
the effects of TNF-α can also be mediated via the PI3K/Akt axis,
which can increase the expression of cadherin-11 in FLS, resulting
in an increased degree of synovitis and cartilage destruction.
Unlike the aforementioned proinflammatory cytokines,
in OA pathogenesis, proinflammatory cytokines, including IL-6,
IL-15, IL-17 and IL-18, synergistically promote the development of
inflammation, upregulate the expression levels of MMPs, promote the
degradation of cartilage ECM and, eventually, destroy the normal
structure of the knee joint (58–62).
IL-6
IL-6 is a cytokine with numerous biological
functions that is produced by monocytic macrophages, endothelial
cells and lymphoid cells, and is crucial in the immune response,
acute phase response and hematopoietic regulation (63). Healthy cartilage tissue may
produce small quantities of IL-6, but a previous study has reported
abnormally elevated IL-6 levels in the synovial fluid or serum in
patients with OA (16). Highly
expressed IL-6 can induce MMP-3 and MMP-13 production and promote
cartilage ECM degradation, which therefore destroys the normal
joint structure (64).
Furthermore, IL-6 is considered to be the key cytokine that causes
changes in the subchondral bone layer, including promoting
inflammation in synovial tissue, increasing cartilage permeability,
accelerating osteoclast formation, as well as causing cartilage
absorption, degradation and destruction, ultimately leading to
enhanced FLS proliferation (65,66). Moreover, studies have demonstrated
a synergistic effect of IL-6 with IL-1β and TNF-α (65,67). IL-6 itself has no direct effect on
the synthesis of protease, PGE, or matrix proteins, but it may
stimulate TIMP synthesis, thereby exerting a protective effect on
cartilage (68). Proinflammatory
cytokines can also activate the MAPK signaling pathway in
synoviocytes and chondrocytes, which initiates a signaling cascade,
leading to the release of the inflammatory mediator IL-6 and damage
of articular cartilage (66).
IL-15
IL-15 is a cytokine with a wide range of biological
functions that is produced by numerous types of cells, including
activated monocytes, epidermal cells and FLS. IL-15 is a key
component of the body's immune response (69). IL-15 is a glycoprotein that exists
in the form of four interconnected α-helices and has a mass of
14–15 kDa (70,71). It acts primarily via stimulating
the differentiation and proliferation of T cells and natural killer
(NK) cells (72). Moreover, it is
one of the most well-documented cytokines involved in the
pathogenesis of rheumatoid arthritis (RA) (73,74). A previous study reported that the
levels of IL-15 and MMP-7 in the synovial fluid of patients with OA
were significantly increased, and higher levels were associated
with more severe OA manifestations, indicating an association
between IL-15 and OA (75).
Furthermore, IL-15 can upregulate MMP-1 expression levels and
promote the degradation of cartilage in the ECM (61). IL-15 can also induce the
activation of mononuclear macrophages and neutrophils, which
secrete a variety of proinflammatory cytokines, such as TNF-α,
IL-1, IL-6 and IL-17 (76).
Moreover, highly expressed proinflammatory cytokines, such as TNF-α
and IL-1β, may in turn promote IL-15 release (76). The proinflammatory cytokines
interact to promote inflammation and eventually destroy the normal
cartilage structure, which causes OA (77). Furthermore, a previous study
reported that increased serum IL-15 levels are be correlated with
pain perception and the severity of lesions on x-ray images
(78). IL-15 may also be involved
in bone destruction in RA and can be used as one of the indicators
for monitoring the damage of articular cartilage in patients with
RA (61).
IL-17
IL-17 is a proinflammatory cytokine produced by
CD4+ memory T lymphocytes and monocytes. It consists of
6 members (IL-17A-F), which interact via five types of receptors
(IL-17RA-E) (79,80). IL-17 has a variety of biological
effects. Studies have reported that it can cause cartilage
destruction primarily via stimulating FLS, chondrocytes and
macrophages to release enzymes that degrade cartilage (81–84). Another study indicated that IL-17
also serves an important role in the pathogenesis of OA and in
OA-related pain (85). IL-17
causes inflammatory cells to release various proinflammatory
factors and compounds, including IL-1β, TNF-α, NO and PGE2, as well
as MMPs, which may promote the development of OA and accelerate the
destruction of articular cartilage. Furthermore, this process is
associated with the release of proinflammatory cytokines, such as
IL-7, IL-6 and IL-23 (86). The
effect of IL-17 on the secretion of VEGF by chondrocytes and FLS is
also a known characteristic and it is conducive to the
overdevelopment of the vascular network in the synovium, which
leads to hypertrophy (87,88).
It has also been reported (89)
that IL-17A stimulation can alter chondrocyte morphology,
inhibiting the activity of chondrocytes and producing a series of
inflammatory responses, which may ultimately lead to chondrocyte
apoptosis.
IL-18
IL-18 is produced by activated macrophages,
dendritic cells and epithelial cells. It is a precursor of
pro-IL-18, consists of 192 amino acid residues, and is converted
into biologically active forms following activation by caspase-1 or
protease 3 (90,91). It is primarily involved in
mediating tissue inflammation and it is also crucial for regulating
immune networks. While mediating tissue inflammation, IL-18 is
associated with TNF-α and comes into direct contact with T
lymphocytes in the synovial fluid (92) and therefore stimulates monocytes
to increase the release of TNF-α and induces the production of
IFN-γ. The latter can significantly stimulate the release of IL-6,
NO and PGE2 in the synovial fluid, which are inflammatory factors
involved in the occurrence of OA (93). Furthermore, IL-18 can increase the
concentration of cartilage-degrading enzymes by inducing TNF-α,
which inhibits the synthesis of proteoglycans, aggrecan and Col II,
promotes their degradation and, ultimately, inhibits the
proliferation of chondrocytes, accelerating their apoptosis and
destroying articular cartilage (94–96). IL-18 also affects the chondrocytes
in patients with OA. IL-18 induces chondrocytes to exhibit typical
morphological changes of apoptotic cells via inducing the
upregulation of IL-18Rα expression on the chondrocyte surface and
upregulating the expression of MMP-1, MMP-3 and MMP-13 (97).
In summary, proinflammatory cytokines exert a
primarily destructive effect on articular cartilage. This is a
multi-layered effect, involving not only the induction of
chondrocyte senescence and apoptosis, but also a reduction in the
synthesis of key components of the ECM, such as proteoglycans and
Col II. Furthermore, inflammatory cytokines may promote the
synthesis and release of several proteolytic enzymes that break
down articular cartilage, including MMPs and ADAMTSs. Therefore, OA
may be treated at the molecular level via inhibition of a single
dominant proinflammatory cytokine, which may improve the efficacy
of OA treatment and the efficiency of cartilage repair by improving
both the internal and external factors of medication and
rehabilitation.
Anti-inflammatory cytokines
Anti-inflammatory cytokines in OA include cytokines
that inhibit at least one of the main proinflammatory cytokines
responsible for the development and progression of OA, such as
IL-1β and TNF-α. At present, the main anti-inflammatory cytokines
in OA are considered to be IL-4, IGF, IL-10 and TGF-β (20). In the pathogenesis of OA,
anti-inflammatory cytokines may inhibit the action of
proinflammatory cytokines, downregulate the expression levels of
MMPs, inhibit the inflammatory response and lysis of articular
chondrocytes, promote the synthesis of chondrocyte proteoglycans
and Col II and delay the progression of OA (20). The roles of these cytokines are
discussed below (Table II).
| Table II.Potential roles and origins of
anti-inflammatory cytokines. |
Table II.
Potential roles and origins of
anti-inflammatory cytokines.
| Cytokine | Source | Role | (Refs.) |
|---|
| IL-4 | Macrophages and TH2
cells | Reduces the
secretion of TGF-β receptors in the body by regulating IL-1, IL-6,
and TNF secretion | (93–98) |
| IGF | Macrophages,
fibroblasts, chondrocytes and osteoblasts | Stimulates the
production of cartilage matrix components. Also thought to
stimulate osteoclast cell lineage replication | (106,108–110) |
| IL-10 | Macrophages | Inhibits the
secretion of various proinflammatory cytokines and compounds,
including, IL-6, TNF-α and NO, and promotes Col II synthesis | (119–122) |
| TGF-β | Macrophages,
fibroblasts, chondrocytes, osteoblasts and osteoclasts | Stimulates the
production of cartilage matrix components. Promotes chondrogenic
differentiation of MSCs | (126,129–131) |
IL-4
IL-4 is a protein composed of 129 amino acids in the
form of four interlinked α-helices stabilized by three disulfide
bonds (98–100). IL-4 is a ligand (101) and as a cytokine produced by
activated T cells, it can regulate a variety of immune cells,
including B cells, T cells, mast cells, M2 macrophages and
hematopoietic cells (83,102). Previous studies have
demonstrated that IL-4 exerts a significant inhibitory effect on
the expression and release of proinflammatory cytokines and blocks
or inhibits monocyte-derived cytokines, including IL-1, TNF-α,
IL-6, IL-8 and the macrophage inflammatory protein-1a (103–106). It has also been reported to
inhibit macrophage cytotoxic activity, kill parasites and produce
macrophage-derived NO (107).
Furthermore, it can stimulate IL-1RA synthesis (108). IL-4 also exerts a strong
protective effect on cartilage and numerous studies have
demonstrated that IL-4 can induce the transcriptional regulator
CITED2, which can reduce variations in proteoglycan production
during the course of OA and prevent proteoglycan degradation in
articular cartilage by inhibiting MMP-13 (109–111). A study also demonstrated that
the protective effect of IL-4 on cartilage is attributed to NO
inhibition. In a previous study (112), in which recombinant rat IL-4 was
injected into OA rats, the results indicated that the recombinant
rat IL-4 downregulated the expression of iNOS mRNA and NO
production in chondrocytes induced by cyclic tensile stress.
Therefore, a limited but significant improvement in cartilage
destruction may be achieved, loss of aggrecan may be prevented and
the number of nitrotyrosine-positive chondrocytes may be reduced.
These effects are not dose-dependent and therefore OA-induced
cartilage destruction may be treated by IL-4 via inhibition of NO
production by chondrocytes (113). Furthermore, IL-4 can promote
macrophage transformation into the M2 phenotype by promoting an
immunomodulatory microenvironment, effectively removing the
proinflammatory debris of articular cartilage and maintaining the
stability of osteoclasts to maintain the health of the articular
cartilage (114). IL-4 can also
inhibit the expression of ADAMTS-4 and −5, delay ECM degradation
and promote the synthesis of chondrocyte proteoglycan and Col II in
articular cartilage (27).
In conclusion, IL-4 can inhibit inflammation by
decreasing the expression and release of inflammatory cytokines and
promoting the expression of IL-1RA. IL-4 serves a chondroprotective
role at the gene and protein levels by decreasing the expression of
NO, MMPs and ADAMTSs.
IGF
IGF is a peptide hormone that exerts the
physiological effect of growth stimulation. IGF regulates the
synthesis of glycogen and proteins, cell proliferation,
differentiation and apoptosis, and induces chondrogenesis (115). The IGF network is composed of
IGF-1, IGF-2 and their receptors (IGF-1R and IGF-2R), as well as
six IGF-binding proteins. As one of the main IGF factors, IGF-l can
be synthesized by a variety of cell types (115). It acts on targeted organs via
endocrine, paracrine or secretory actions, regulates glycogen and
protein synthesis and decomposition, and participates in the
processes of metabolism, cell proliferation, differentiation and
apoptosis, indicating that it serves an important role in repairing
numerous systems in the body (116).
OA is a skeletal motor system disease characterized
by cartilage degeneration. Chondrocytes, as the only cells in
cartilage, can proliferate under the influence of IGF-1, and
synthesize cartilage matrix proteoglycans and matrix collagen.
Physiologically, in infants, children and adolescents, IGF-l
promotes linear bone growth by stimulating chondrocyte
proliferation and shaping (117). In adults, IGF-1 inhibits
chondrocyte aging and death by stimulating chondrocytes to
synthesize matrix proteins, suggesting that IGF-1 may be used to
treat OA (118,119). In an in vitro test,
Yaeger et al (120)
demonstrated that after bovine articular chondrocytes were treated
with IGF-1, the collagen content was significantly increased when
compared with the untreated control. Furthermore, Madry et
al (121) treated
osteochondral defects in a rabbit model by using chondrocytes
transfected with a plasmid vector containing human IGF-1 in a
hydrogel. This study confirmed that at 14 weeks post-treatment, the
test group was significantly superior to the control group in terms
of histological grading (defect filling, integration and cell
morphology). In an equine model, Fortier et al (122) reported that adding IGF-1 to the
chondrocyte-fibrin complex enhanced cartilage formation in
cartilage defects, and 8 months after treatment compared with the
control group treated by chondrocyte-fibrin complex alone, the
severe defects in the repaired tissue were filled, with the mean
Col II content exhibiting a significant increase. These results
suggested that in the co-culture of IGF-1 and chondrocytes,
chondrocytes are regulated by IGF-1, which promotes not only
chondrocyte division and proliferation, but also cell function and
synthesis of proteoglycans and Col II. IGF-1 also exhibited a
positive effect when combined with other growth factors/cytokines.
Certain studies reported that, when IGF-1 and TGF-β were combined,
the production of cartilage matrix components (such as
proteoglycans) increased, the Col II and aggrecan gene expression
levels in chondrocytes were upregulated and the chondrogenic
differentiation of mesenchymal stem cells (MSCs) was enhanced
(123,124). Morisset et al (125) injected adenovirus-transfected
IGF-1 and IL-IRa into the joints of 12 horses with minor fractures
in the carpal bones caused by full-thickness cartilage defects. It
was discovered that the cartilage defects were completely restored.
The combination of IGF-1 and IL-1RA further reduced cartilage
degeneration when compared with IL-1RA alone. Therefore, the
combination of IGF-1 and IL-1 Ra could potentially reduce or even
reverse the loss of cartilage in OA, thus improving the effects of
OA treatment. Furthermore, IGF-1 may also inhibit chondrocyte
apoptosis. An in vitro chondrocyte culture demonstrated that
IGF-1 reversed the inhibitory effects of dexamethasone on
chondrocyte proliferation and prevented the apoptosis of
chondrocytes caused by collagenase (126). Moreover, IGF-1 may prevent
chondrocyte apoptosis via the P13K and MAPK signaling pathways
(127).
IL-10
IL-10 is a well-known anti-inflammatory and
immunomodulatory cytokine that is primarily released by cells of
the immune system, including monocytes, macrophages, T cells, NK
cells and B cells (128). IL-10
may also be produced by a few connective tissue cell types,
including chondrocytes, and is involved in processes such as ECM
remodeling in connective tissues (128). IL-10 can inhibit the synthesis
and secretion of related proinflammatory cytokines, such as IL-6,
thus regulating inflammatory immunity (129,130). IL-10 can also reduce iNOS2
expression, thereby reducing the release of NO, interfering with
the oxidative stress matrix and delaying joint degeneration
(131). Furthermore, IL-10
prevents early apoptosis induced by TNF-α by reducing the
expression of TNF-α-induced matrix-degrading enzymes, the release
of glycosaminoglycans and the formation of G1 fragment of the
proteoglycan aggrecan (132).
IL-10 can also activate the SMAD1/SMAD5/SMAD8 and ERK1/2/MAPK
signaling pathways and induce the expression of bone morphogenetic
protein (BMP)-2 and BMP-6. As members of the TGF-β family, BMP
proteins serve a crucial role in chondrogenesis (105). Through their signaling pathways,
they affect numerous genes and proteins regulating chondrocyte
mesenchymal cell transformation, such as NK3 homeobox 2/Sox9, Sox5
and Sox6 (133,134).
TGF-β
The TGF-β superfamily consists of >30
structurally-related members, including TGF-β1, 2 and 3 (135). TGF-β1 is a multifunctional
growth factor that regulates a wide range of biological processes,
including cell proliferation, survival, differentiation, migration
and ECM generation (136,137).
TGF-β1 signaling affects MSCs and their progenitors. Studies have
demonstrated that active TGF-β1 released by osteoclasts during bone
resorption may guide the migration of bone marrow MSCs (BMSCs) to
form new cartilage at resorption sites, induce Sox-9 expression in
BMSCs and increase the generation of ECM (138–140). Therefore, TGF-β1, as a soluble
factor, contributes to BMSC differentiation. Furthermore, TGF-β1 is
critical for maintaining homeostasis between subchondral bone and
articular cartilage (141).
TGF-β1 is a well-documented potent chondroblast factor that may
stimulate chondrocytes to synthesize and secrete proteoglycans and
Col II (141). One of the first
TGF-β1 activities to be discovered was the in vitro
induction of primitive rat mesenchymal cells to promote
chondrogenesis (142,143). Subsequently, TGF-β1-induced
chondroblasts have been observed in rabbit chondrocyte cultures
(144), chicken mesenchymal
cells, as well as bovine nose and articular chondrocyte cultures
(145,146).
With a biological function similar to that of
TGF-β1, TGF-β2 may regulate cartilage and osteogenic
differentiation (147). It has
been demonstrated that following TGF-β2 treatment of
dedifferentiated chondrocytes, the cells re-expressed glucosamine
and Col II and the chondrogenic phenotype was therefore restored.
It has also been reported that precursor cells isolated from the
perichondrium show the potential for chondrogenic differentiation
after being cultured in a medium containing TGF-β2 and IGF-1 before
producing a cartilage matrix (148).
As another important member of the TGF family of
cytokines, TGF-β3 can promote the proliferation and differentiation
of chondrocytes, accelerate the formation of ECM, inhibit the
activity of various inflammatory mediators, including IL-1, MMPs
and TNF-α, and reduce the immune response of the body, thus serving
a crucial role in wound healing, particularly in cartilage growth
and reconstruction (149).
TGF-β3 is expressed in a number of different types of cells and
tissues in the body, including the placenta, adipose tissue,
embryonic tissues, liver, bone and bone marrow, as well as in
tumors, and is secreted extracellularly via autocrine and paracrine
routes. TGF-β3 is an extracellular ligand and its effects are
mediated by binding to serine/threonine kinase receptors on the
cell surface. TGF-β3 activates the transcription factors
SMAD2/SMAD3 via the intracellular SMAD signaling pathway, and
activates and binds to SMAD4, forming a complex, which translocates
to the nucleus and regulates the transcriptional expression of the
downstream TGF-β gene and therefore its biological function
(150,151). Furthermore, it can inhibit
inflammatory responses and promote TIMP expression (150). It has also been reported that
the ability of TGF-β2 and TGF-β3 to induce chondrogenic
differentiation of MSCs may be superior to that of TGF-β1 (151).
In summary, the main effects of anti-inflammatory
cytokines involve inhibiting the synthesis of proinflammatory
cytokines, particularly IL-1β and TNF-α. The main effects of
anti-inflammatory cytokines include increasing the synthesis of
proteins and polysaccharides, inhibiting chondrocyte apoptosis and
reducing the synthesis and secretion of MMPs and ADAMTSs.
Anti-inflammatory cytokines can also act on target cells to
increase matrix degradation products dependent on cytokine
functions. The cytokines that inhibit or antagonize the activity of
catabolic cytokines are classified as anti-catabolic or inhibitory
cytokines (IL-4 and IL-10), and the cytokines that promote
chondrocyte proliferation and differentiation are classified as
anabolic cytokines (IGF and TGF-β). The effects of catabolic
cytokines should be viewed as inhibitory on proinflammatory
cytokines rather than having direct protective effects on
cartilage. In healthy joints, anti-inflammatory cytokines do not
exert significant anabolic effects but may have sufficient
biochemical dominance against proinflammatory cytokines. The
effects of anti-inflammatory cytokines may become apparent in the
joints affected by disease only in the presence of inflammatory
mediators. Anabolic cytokines mainly exert a protective effect on
chondrocytes and they also inhibit proinflammatory cytokines.
Maintaining a dynamic balance between proinflammatory and
anti-inflammatory cytokines is crucial for OA treatment. Thus, the
roles of proinflammatory and anti-inflammatory cytokines in OA must
be further elucidated and efforts must be made to design an
appropriate treatment method that may prevent or delay OA
progression.
CTE
CTE technology is used to culture and amplify
cartilage seed cells in vitro, and to implant them on
high-density scaffolds with good biocompatibility and
degradability, thus forming tissue-engineered cartilage using
regulatory factors. There are three main conditions for
constructing tissue-engineered cartilage: i) A sufficient quantity
of functioning seed cells; ii) a suitable cellular scaffold; and
iii) cytokines that can regulate cell proliferation and maintain
the phenotypic characteristics of the cells (152,153). Directed differentiation of
chondrocytes may be regulated by controlling relevant environmental
factors, therefore producing the desired cells. Cytokines can
significantly promote cell differentiation in chondrocytes, which
induces the relevant functions. Multiple cytokines jointly
stimulate the entire seed cell differentiation process to
chondrocytes (154). It has been
reported (153) that tissue
engineering can be used in the repair of the nose, trachea or
auricle, and certain patients have undergone successful repair of
nasal alar lobe defects via CTE. The development of CTE technology
is promising and it may enable successful cartilage repair.
Regulating proinflammatory and anti-inflammatory cytokines and
maintaining their balance during OA treatment has been proven to be
a useful treatment strategy (155). CTE technology is crucial for
regulating anti-inflammatory and proinflammatory cytokines and
maintaining the balance between the two, which may provide a new
approach to OA treatment. How CTE technology can be used to treat
OA by regulating proinflammatory and anti-inflammatory cytokines is
discussed in this section.
MSCs
MSCs are adult stem cells with multipotent
differentiation potential and the ability to self-renew, which were
first isolated from the bone marrow. MSCs are ideal seed cells for
tissue engineering, as they can differentiate into adipocytes,
osteoblasts and chondrocytes and are capable of transdermal
differentiation (156). A number
of studies have reported that MSCs can be used to repair and
regenerate myocardial tissue, damaged bone tissue, cartilage tissue
and tendons (157–159). The main mode of action for MSCs
is based on paracrine mechanisms (160). Exosomes are important carriers
of paracrine delivery factors promoting cell-to-cell conduction. As
the smallest known bilayer nanovesicles, exosomes are rich in
biologically active molecules, such as mRNAs, microRNAs (miRNAs)
and proteins, and they are involved in communication between cells
(155). All cells secrete
exosomes, which are present in bodily fluids, including blood,
saliva, amniotic fluid, ascitic and cerebrospinal fluid (161). During early-stage OA, exosomes
in the articular cavity mediate anti-inflammatory cytokines and
miRNAs for cartilage repair. However, in the intermediate or
advanced stages of the disease, they can induce related
proinflammatory cytokine and proinflammatory miRNA production to
accelerate relevant pathological changes (155). Therefore, exosomes can regulate
the expression of proinflammatory and anti-inflammatory cytokines,
which can therefore alter the pathological and/or physiological
processes implicated in certain diseases. A previous study
demonstrated (161) that the
exosomes of adipose MSCs (ADMSCs) can reduce the production of
proinflammatory cytokines, including TNF-α, IL-6 and IL-10, in OA
chondrocytes. Furthermore, following the downregulation of COX-2
and PGE1 expression levels, PGE2 production is decreased (155). Moreover, ADMSC exosomes inhibit
the activation of the NF-κB signaling pathway, serving an
anti-inflammatory role (33). It
has also been demonstrated that BMSC exosomes inhibit chondrocyte
destruction via inflammatory mediators and TNF-α-induced
inflammation. The chondrocytes treated by exosomes exhibit
significantly decreased levels of COX-2 expression and inflammatory
mediators, including IL-1, IL-6, IL-8 and IL-1 (162,163). As well as reducing the
inflammatory response of chondrocytes, MSC exosomes can also
regulate chondrocyte apoptosis and macrophage activity. For
example, Cosenza et al (163) investigated the protective effect
of MSC-derived exosomes in cartilage degradation in OA rats. The
results demonstrated that MSC-derived exosomes significantly
reduced IL-1β-induced chondrocyte apoptosis. Furthermore,
MSC-derived exosomes reduce the expression levels of CD86, major
histocompatibility complex II and CD40 in F4/80 macrophages. MSCs
co-cultured with macrophages can also reduce type M1 macrophages
and promote their transformation into type M2 macrophages (Fig. 2) (164). In a collagenase-induced OA
model, intra-articular injections of MSC-derived exosomes
significantly increase cartilage volume, surface volume ratio and
thickness (165). Therefore,
exosomes can alleviate inflammation and repair cartilage by
mediating the effects of proinflammatory and anti-inflammatory
cytokines.
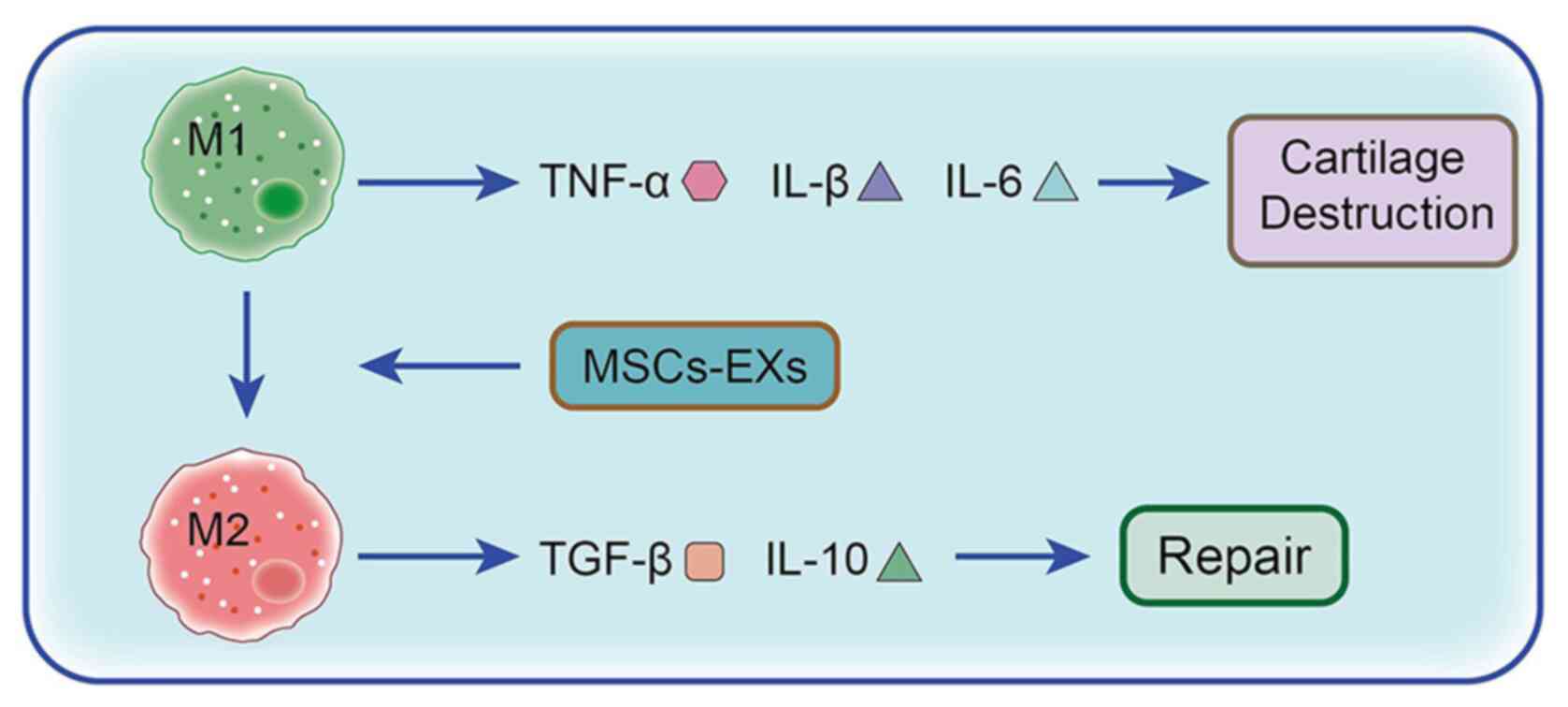 | Figure 2.Role and mechanism of MSCs in the
treatment of OA. When stimulated by stress, surgery and/or trauma,
M1 macrophages, under the induction of IFN and lipopolysaccharide,
produce the proinflammatory cytokines TNF-α, IL-1β and IL-6,
consequently promoting cartilage damage. However M2 macrophages,
under the induction of IL-13 and IL-3, produce the
anti-inflammatory cytokines IL-10 and TGF-β1 to promote cartilage
repair. MSCs promote the phenotypic transformation of synovial
macrophages from M1 to M2 via exosomes, which significantly reduces
the levels of the proinflammatory cytokines IL-1β, IL-6 and TNF-α,
while the significant increase in anti-inflammatory cytokine IL-10
and reduction in inflammation enable cartilage repair. OA,
osteoarthritis; MSC, mesenchymal stem cell; MSCs-EX, exosome. |
As important mediators of intercellular
communication, exosomes are involved in the occurrence of joint
diseases. Although exosomes may accelerate articular cartilage
destruction and promote the progression of joint diseases via
signaling pathways, they have also shown great potential in OA
pathogenesis and treatment. The exosomes released by MSCs can
promote anti-inflammatory cytokine secretion, delay joint disease
progression and repair damaged joints by inhibiting the secretion
of proinflammatory cytokines. Further research on these exosomes
may provide novel ideas for treating OA. Although studies on
exosomes have reported certain exciting findings, there remain
certain limitations, such as the high manufacturing costs,
difficulties in isolation and purification, and insufficient
efficacy and safety evaluation (163,166,167). Therefore, exosomes still require
in-depth studies to further explore their therapeutic
potential.
Scaffolding technology
The CTE bionic scaffold technology offers numerous
possibilities for cartilage repair and regeneration. Guided by
cytokines, autologous seed cells (stem cells/progenitor cells) can
be recruited from the injured site to repair the injured bone and
cartilage using scaffold materials to regulate their own
microenvironment (168).
Therefore, the exogenous seed cells required by traditional tissue
engineering technology may become unnecessary, and certain
limitations, such as the long duration for in vitro
reproduction of exogenous seed cells and potential immune system
rejection, may be avoided (163). The duration for treatment may be
significantly shortened and the treatment failure rate may be
reduced. Therefore, when treating bone and cartilage injuries, am
effective therapeutic may be produced (168,169). There are currently two main
categories of scaffold materials commonly used in CTE: i) Synthetic
polymer materials, such as polyglycolic acid, polylactic acid (and
their copolymer, poly lactic-co-glycolic acid) and polyvinyl oxide;
and ii) natural polymer materials, such as collagen, chitosan,
alginate, hyaluronic acid, fibrin and agarose. According to their
morphology, these can be divided into prefabricated 3D porous
scaffolds and hydrogel materials (170). The aforementioned materials can
endogenously regulate proinflammatory and anti-inflammatory
cytokines and can be used to treat OA by controlling the
microenvironment in vivo. Jiang et al (171) previously prepared a composite
electrospinning nanofiber composed of cartilage matrix components
(collagen or chondroitin sulfate) and
poly(ε-caprolactone)-polytetrahydrofuran. Moreover, experimental
results (137) have demonstrated
that this nanofiber can regulate the TNF and hypoxia-inducible
factor-1 signaling pathways by addition of chondroitin sulfate or
collagen, which upregulates BMP2 and IGF-1 and promotes cartilage
formation. Fasolino et al (172) proposed a bionic chitosan-based
scaffold and experimentally demonstrated that the biologically
activated scaffold could inhibit the synthesis of inflammatory
mediators, such as IL-1β, reduce oxidative stress metabolites and
promote the production of the anti-inflammatory marker IL-10 in
MSCs. The biologically activated scaffold also exhibited
anti-inflammatory activity in an in vitro co-culture.
Therefore the damaged cartilage microenvironment in vivo
could be efficiently simulated by it. Results have also
demonstrated that at the molecular level the cartilage scaffolds
can regulate the microenvironment in the joints, promote the
secretion of anti-inflammatory cytokines and inhibit the secretion
of proinflammatory cytokines, which promotes cartilage repair.
Bhardwaj et al (173)
co-cultured articular chondrocytes and ADMSCs in a 3D scaffold
based on silk fibroin proteins. Compared with the single control
group, the co-culture group exhibited significant increases in
TGF-β1 and IL-10 levels, thus upregulating the expression of the
bone formation markers aggrecan, Sox-9 and Col II, while the
expression levels of the hypertrophy gene collagen type X and
MMP-13 exhibited a significant decrease. Subsequently, inflammation
was controlled and the cartilage was repaired. According to these
results, cartilage scaffolds may be used as MSC carriers to further
promote the secretion of anti-inflammatory cytokines and enhance
anti-inflammatory effects in co-culture.
Existing studies have reported that cartilage
anabolic factors, such as TGF-β, fibroblast growth factor and
IGF-1, that can restore the function of chondrocytes,
cartilage-protective drugs and matrix components, such as
chondroitin sulfate and hyaluronic acid, can decrease
IL-1β-mediated NF-κB activation (174–176), thereby inhibiting cartilage
breakdown, delaying OA development and repairing damaged cartilage.
Park et al (177)
described an injectable microsphere made from genipin cross-linked
gelatin and discovered that the anti-inflammatory cytokines
released from the gelatin microspheres, mediated by inflammatory
cells, may alleviate inflammation in chondrocytes activated by
IL-1β and lipopolysaccharide. The results have demonstrated that
such biomaterial-based methods can be used to synchronize drug
release during the inflammatory process, thereby extending
anti-inflammatory cytokine therapeutic efficacy and retention
times. Moutos et al (178) produced two hemispheric scaffolds
with varying configurations constructed from a 3D polyester
(ε-caprolactone). A scaffold-mediated lentiviral transfecting
technique has been used for the genetic modification of ADMSCs in
the scaffold to produce high levels of anti-inflammatory cytokines
for treating OA in an exogenously regulated and induced manner.
Levinson et al (179)
observed that heparin can covalently combine with a variety of
hyaluronic acids to form hydrogels, therefore achieving continuous
release of TGF-β1 and enabling MSCs to mature into cartilage tissue
without the use of any additional growth factors. The results
demonstrated that cartilage scaffolds coated with drugs or
anti-inflammatory cytokines for the treatment of OA may strongly
induce anti-inflammatory cytokine expression or cause sustained
release of anti-inflammatory cytokines. Therefore, the application
of drugs, growth factors, anti-inflammatory cytokines and
proinflammatory cytokines is expected to become the focus of
studies on the clinical application of CTE scaffolds in the future.
Bionic scaffolds have the potential for cartilage repair and
regeneration, but they also have certain limitations. Even when
several materials are combined, the cartilage damage may not be
fully repaired (179).
Therefore, future research should focus on further optimizing
scaffold preparation technology and related processes, explore the
selection and proportion of different scaffold materials used, or
introduce different types of drugs, growth factors and cytokines,
in an attempt to overcome the current restrictions associated with
cartilage injury repair and regeneration. Furthermore, future CTE
should also aim to employ simpler preparations of scaffolding with
improved stability and more effective scaffold materials, therefore
realizing clinical transformation as soon as possible.
In summary, OA and relevant repair procedures may
be improved by regulating proinflammatory and anti-inflammatory
cytokines via CTE technology. However, CTE technology is still
faced with major obstacles, including the selection of source cells
and scaffolds, dedifferentiation and amplification procedures,
timing for chondrocyte expansion prior to implantation, loss of
chondrocytes and high costs. In the future, different methods may
be combined for the effective treatment of OA, including advanced
scaffolds, highly differentiated chondrocytes, 3D-printed
engineering structures, appropriate lubrication and improvement of
the proinflammatory environment. This will occur by regulating the
effects of proinflammatory and anti-inflammatory cytokines, which
may greatly promote the regeneration of articular cartilage
(180). However, the aim of CTE
is to ensure that the repaired cartilage will regain its basic
function, which is to support weight, by mimicking the properties
and structure of natural cartilage. A previous study demonstrated
that CTE can successfully repair cartilage or osteochondral defects
in pigs and meniscus defects in dogs (178). However, this has only been
tested on animal models, and the safety issues associated with the
extrapolation of animal experimental results to humans have not yet
been resolved. Thus, CTE has not been used in clinical practice to
date and it is difficult to evaluate its actual efficacy in
humans.
Conclusion
Proinflammatory cytokines contribute to the release
of other inflammatory mediators and trigger cartilage degeneration
and destruction. Anti-inflammatory cytokines inhibit the actions of
proinflammatory cytokines, promote chondrocyte proteoglycan and Col
II synthesis, and delay OA progression (2). The imbalance between OA-related
inflammatory cytokines may cause abnormal metabolism in the knee
cartilage, ultimately destroying the normal knee joint structure.
After further understanding of the regulation of chondrocytes and
synovium by proinflammatory and anti-inflammatory cytokines, it can
be inferred that early molecular cytological clinical intervention
in OA, such as inducing the expression of anabolic cytokines while
inhibiting that of catabolic cytokines, so that chondrocytes or
subchondral osteocytes and synovial cells can achieve a balance of
catabolism based on self-repair, will still represent an obstacle
in the future (7–9). As stated in the present review, it
has been hypothesized that blocking the IL-1β signaling pathway may
provide a new direction for the treatment of OA. Furthermore, TGF-β
may be a focus of future research, as it contributes to the
differentiation of MSCs.
The application of drugs, growth factors,
anti-inflammatory cytokines and CTE is expected to become the focus
of clinical research in the future. Moreover, scaffolds loaded with
growth factors, anti-inflammatory cytokines or drugs can
endogenously reduce the levels of inflammatory cytokines, promote
anti-inflammatory cytokine expression and create optimal conditions
for cartilage reproduction (181). However, previous studies often
paid attention to the function of a single cytokine, while they
ignored the potential synergistic effect of different cytokines
(182–184). In the cartilage
microenvironment, even small fluctuations in concentration due to
the decomposition of several cytokines may cause degradation and
destruction of the cartilage matrix (185). Furthermore, even partial
cytokines can either repair damaged cartilage or inhibit
chondrocyte function and destroy cartilage (186). Therefore, in CTE, the production
of cytokines, activation time, number of receptors and the
interactions between various cytokines must be further
investigated. Therefore, the role of cytokines should be carefully
considered in the study of OA, both as independent factors and in
combination. Investigating cytokines may help fully elucidate the
contributors and triggers involved, identify the relevant
conditions and effects resulting from complex interactions,
understand the main contradictions between various factors, enable
us to alter molecular mechanisms based on CTE techniques and
regulate the intra-articular microenvironment, thereby improving
the overall efficacy of OA treatment. This approach may lead to
important breakthroughs in OA research.
Acknowledgements
Not applicable.
Funding
The present review was supported by the National Natural Science
Foundation of China (grant no. 81672234).
Availability of data and materials
Not applicable.
Authors' contributions
SL was responsible for producing the tables and
searching the literature, as well as being a major contributor in
writing the manuscript. ZD was mainly responsible for revising the
manuscript. KC was responsible for making the figures. SJ
contributed to the conception of the study. FZ made substantial
contributions to conception and design. YY and ZF were involved in
drafting the manuscript or revising it critically for important
intellectual content. HX contributed significantly to manuscript
preparation, and approved the final version. JX and WZ were
responsible for funding support and reviewing the article. All
authors read and approved the final manuscript. Data authentication
is not applicable.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Abramoff B and Caldera FE: Osteoarthritis:
Pathology, diagnosis, and treatment options. Med Clin North Am.
104:293–311. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Goldring MB and Goldring SR: Articular
cartilage and subchondral bone in the pathogenesis of
osteoarthritis. Ann NY Acad Sci. 1192:230–237. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Xia B, Di Chen, Zhang J, Hu S, Jin H and
Tong P: Osteoarthritis pathogenesis: A review of molecular
mechanisms. Calcif Tissue Int. 95:495–505. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zhang Q, Ji Q, Wang X, Kang L, Fu Y, Yin
Y, Li Z, Liu Y, Xu X and Wang Y: SOX9 is a regulator of
ADAMTSs-induced cartilage degeneration at the early stage of human
osteoarthritis. Osteoarthritis Cartilage. 23:2259–2268. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tortorella MD and Malfait AM: Will the
real aggrecanase(s) step up: Evaluating the criteria that define
aggrecanase activity in osteoarthritis. Curr Pharm Biotechnol.
9:16–23. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Matsuo M, Nishida K, Yoshida A, Murakami T
and Inoue H: Expression of caspase-3 and −9 relevant to cartilage
destruction and chondrocyte apoptosis in human osteoarthritic
cartilage. Acta Med Okayama. 55:333–340. 2001.PubMed/NCBI
|
|
7
|
Abdel-Sayed P and Pioletti DP: Strategies
for improving the repair of focal cartilage defects. Nanomedicine
(Lond). 10:2893–2905. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Medvedeva EV, Grebenik EA, Gornostaeva SN,
Telpuhov VI, Lychagin AV, Timashev PS and Chagin AS: Repair of
damaged articular cartilage: Current approaches and future
directions. Int J Mol Sci. 19:23662018. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hunter DJ and Bierma-Zeinstra S:
Osteoarthritis. Lancet. 393:1745–1759. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Rhee SM, You HJ and Han SK: Injectable
tissue-engineered soft tissue for tissue augmentation. J Korean Med
Sci. 29 (Suppl 3):S170–S175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schott EM, Farnsworth CW, Grier A, Lillis
JA, Soniwala S, Dadourian GH, Bell RD, Doolittle ML, Villani DA,
Awad H, et al: Targeting the gut microbiome to treat the
osteoarthritis of obesity. JCI Insight. 3:e959972018. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kuo SJ, Yang WH, Liu SC, Tsai CH, Hsu HC
and Tang CH: Transforming growth factor β1 enhances heme oxygenase
1 expression in human synovial fibroblasts by inhibiting microRNA
519b synthesis. PLoS One. 12:e01760522017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang T and He C: Pro-inflammatory
cytokines: The link between obesity and osteoarthritis. Cytokine
Growth Factor Rev. 44:38–50. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kapoor M, Martel-Pelletier J, Lajeunesse
D, Pelletier JP and Fahmi H: Role of proinflammatory cytokines in
the pathophysiology of osteoarthritis. Nat Rev Rheumatol. 7:33–42.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nguyen L, Sharma A, Chakraborty C, Saibaba
B, Ahn ME and Lee SS: Review of prospects of biological fluid
biomarkers in osteoarthritis. Int J Mol Sci. 18:6012017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mabey T, Honsawek S, Tanavalee A,
Yuktanandana P, Wilairatana V and Poovorawan Y: Plasma and synovial
fluid inflammatory cytokine profiles in primary knee
osteoarthritis. Biomarkers. 21:639–644. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Boehme KA and Rolauffs B: Onset and
progression of human osteoarthritis-can growth factors,
inflammatory cytokines, or differential miRNA expression
concomitantly induce proliferation, ECM Degradation, and
inflammation in articular cartilage? Int J Mol Sci. 19:22822018.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Goldring MB and Goldring SR:
Osteoarthritis. J Cell Physiol. 213:626–634. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sirikaew N, Chomdej S, Tangyuenyong S,
Tangjitjaroen W, Somgird C, Thitaram C and Ongchai S:
Proinflammatory cytokines and lipopolysaccharides up regulate MMP-3
and MMP-13 production in Asian elephant (Elephas maximus)
chondrocytes: Attenuation by anti-arthritic agents. BMC Vet Res.
15:4192019. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ho YJ, Lu JW, Ho LJ, Lai JH, Huang HS, Lee
CC, Lin TY, Lien SB, Lin LC, Chen LW, et al: Anti-inflammatory and
anti-osteoarthritis effects of Cm-02 and Ck-02. Biochem Biophys Res
Commun. 517:155–163. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fearing BV and Van Dyke ME: In vitro
response of macrophage polarization to a keratin biomaterial. Acta
Biomater. 10:3136–3144. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dinarello CA: Overview of the
interleukin-1 family of ligands and receptors. Semin Immunol.
25:389–393. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Melchiorri C, Meliconi R, Frizziero L,
Silvestri T, Pulsatelli L, Mazzetti I, Borzì RM, Uguccioni M and
Facchini A: Enhanced and coordinated in vivo expression of
inflammatory cytokines and nitric oxide synthase by chondrocytes
from patients with osteoarthritis. Arthritis Rheum. 41:2165–2174.
1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Massicotte F, Lajeunesse D, Benderdour M,
Pelletier JP, Hilal G, Duval N and Martel-Pelletier J: Can altered
production of interleukin-1beta, interleukin-6, transforming growth
factor-beta and prostaglandin E(2) by isolated human subchondral
osteoblasts identify two subgroups of osteoarthritic patients.
Osteoarthritis Cartilage. 10:491–500. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Farahat MN, Yanni G, Poston R and Panayi
GS: Cytokine expression in synovial membranes of patients with
rheumatoid arthritis and osteoarthritis. Ann Rheum Dis. 52:870–875.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sohn DH, Sokolove J, Sharpe O, Erhart JC,
Chandra PE, Lahey LJ, Lindstrom TM, Hwang I, Boyer KA, Andriacchi
TP and Robinson WH: Plasma proteins present in osteoarthritic
synovial fluid can stimulate cytokine production via Toll-like
receptor 4. Arthritis Res Ther. 14:R72012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wojdasiewicz P, Poniatowski ŁA and
Szukiewicz D: The role of inflammatory and anti-inflammatory
cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm.
2014:5614592014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Burrage PS, Mix KS and Brinckerhoff CE:
Matrix metalloproteinases: Role in arthritis. Front Biosci.
11:529–543. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Verma P and Dalal K: ADAMTS-4 and
ADAMTS-5: Key enzymes in osteoarthritis. J Cell Biochem.
112:3507–3514. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Koshy PJ, Lundy CJ, Rowan AD, Porter S,
Edwards DR, Hogan A, Clark IM and Cawston TE: The modulation of
matrix metalloproteinase and ADAM gene expression in human
chondrocytes by interleukin-1 and oncostatin M: A time-course study
using real-time quantitative reverse transcription-polymerase chain
reaction. Arthritis Rheum. 46:961–967. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
El Mansouri FE, Chabane N, Zayed N, Kapoor
M, Benderdour M, Martel-Pelletier J, Pelletier JP, Duval N and
Fahmi H: Contribution of H3K4 methylation by SET-1A to
interleukin-1-induced cyclooxygenase 2 and inducible nitric oxide
synthase expression in human osteoarthritis chondrocytes. Arthritis
Rheum. 63:168–179. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gilman SC, Chang J, Zeigler PR, Uhl J and
Mochan E: Interleukin-1 activates phospholipase A2 in human
synovial cells. Arthritis Rheum. 31:126–130. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hardy MM, Seibert K, Manning PT, Currie
MG, Woerner BM, Edwards D, Koki A and Tripp CS: Cyclooxygenase
2-dependent prostaglandin E2 modulates cartilage proteoglycan
degradation in human osteoarthritis explants. Arthritis Rheum.
46:1789–1803. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lotz M: The role of nitric oxide in
articular cartilage damage. Rheum Dis Clin North Am. 25:269–282.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Haynes MK, Hume EL and Smith JB:
Phenotypic characterization of inflammatory cells from
osteoarthritic synovium and synovial fluids. Clin Immunol.
105:315–325. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Afonso V, Champy R, Mitrovic D, Collin P
and Lomri A: Reactive oxygen species and superoxide dismutases:
Role in joint diseases. Joint Bone Spine. 74:324–329. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Bodmer JL, Schneider P and Tschopp J: The
molecular architecture of the TNF superfamily. Trends Biochem Sci.
27:19–26. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hosseinzadeh A, Kamrava SK, Joghataei MT,
Darabi R, Shakeri-Zadeh A, Shahriari M, Reiter RJ, Ghaznavi H and
Mehrzadi S: Ap MMR-21332-279469 optosis signaling pathways in
osteoarthritis and possible protective role of melatonin. J Pineal
Res. 61:411–425. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
MacEwan DJ: TNF receptor subtype
signalling: Differences and cellular consequences. Cell Signal.
14:477–492. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Grell M, Douni E, Wajant H, Löhden M,
Clauss M, Maxeiner B, Georgopoulos S, Lesslauer W, Kollias G,
Pfizenmaier K and Scheurich P: The transmembrane form of tumor
necrosis factor is the prime activating ligand of the 80 kDa tumor
necrosis factor receptor. Cell. 83:793–802. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Hsu H, Xiong J and Goeddel DV: The TNF
receptor 1-associated protein TRADD signals cell death and
NF-kappaB activation. Cell. 81:495–504. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Hsu H, Huang J, Shu HB, Baichwal V and
Goeddel DV: TNF-dependent recruitment of the protein kinase RIP to
the TNF receptor-1 signaling complex. Immunity. 4:387–396. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Varfolomeev E, Goncharov T, Fedorova AV,
Dynek JN, Zobel K, Deshayes K, Fairbrother WJ and Vucic D: c-IAP1
and c-IAP2 are critical mediators of tumor necrosis factor alpha
(TNFalpha)-induced NF-kappaB activation. J Biol Chem.
283:24295–24299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
O'Donnell MA, Legarda-Addison D, Skountzos
P, Yeh WC and Ting AT: Ubiquitination of RIP1 Regulates an
NF-kappaB-Independent cell-death switch in TNF signaling. Curr
Biol. 17:418–424. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Ea CK, Deng L, Xia ZP, Pineda G and Chen
ZJ: Activation of IKK by TNF alpha requires site-specific
ubiquitination of RIP1 and polyubiquitin binding by NEMO. Mol Cell.
22:245–257. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Bunning RA and Russell RG: The effect of
tumor necrosis factor alpha and gamma-interferon on the resorption
of human articular cartilage and on the production of prostaglandin
E and of caseinase activity by human articular chondrocytes.
Arthritis Rheum. 32:780–784. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Campbell IK, Piccoli DS, Roberts MJ,
Muirden KD and Hamilton JA: Effects of tumor necrosis factor and on
resorption of human articular cartilage and production of
plasminogen activator by human articular chondrocytes. Arthritis
Rheum. 33:542–552. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lefebvre V, Peeters-Joris C and Vaes G:
Modulation by interleukin 1 and tumor necrosis factor of production
of collagenase, tissue inhibitor of metalloproteinases and collagen
types in differentiated and dedifferentiated articular
chondrocytes. Biochim Biophys Acta. 1052:366–378. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Meyer FA, Yaron I and Yaron M:
Synergistic, additive, and antagonistic effects of interleukin-1
beta, tumor necrosis factor alpha, and gamma-interferon on
prostaglandin E, hyaluronic acid, and collagenase production by
cultured synovial fibroblasts. Arthritis Rheum. 33:1518–1525. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Saklatvala J: Tumour necrosis factor alpha
stimulates resorption and inhibits synthesis of proteoglycan in
cartilage. Nature. 322:547–549. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
van den Berg WB: Anti-cytokine therapy in
chronic destructive arthritis. Arthritis Res. 3:18–26. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Brennan FM, Chantry D, Jackson AM, Maini
RN and Feldmann M: Cytokine production in culture by cells isolated
from the synovial membrane. J Autoimmun. 2 (Suppl 1):S177–S186.
1989. View Article : Google Scholar
|
|
53
|
O'Byrne E, Blancuzzi V, Wilson DE, Wong M
and Jeng AY: Elevated substance P and accelerated cartilage
degradation in rabbit knees injected with interleukin-1 and tumor
necrosis factor. Arthritis Rheum. 33:1023–1028. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Pettipher ER, Higgs GA and Henderson B:
Interleukin 1 induces leukocyte infiltration and cartilage
proteoglycan degradation in the synovial joint. Proc Natl Acad Sci
USA. 83:8749–8753. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
van Beuningen HM, Arntz OJ and van den
Berg WB: In vivo effects of interleukin-1 on articular cartilage.
Prolongation of proteoglycan metabolic disturbances in old mice.
Arthritis Rheum. 34:606–615. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Ding X, Zhang Y, Huang Y, Liu S, Lu H and
Sun T: Cadherin-11 involves in synovitis and increases the
migratory and invasive capacity of fibroblast-like synoviocytes of
osteoarthritis. Int Immunopharmacol. 26:153–161. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Liu S, Cao C, Zhang Y, Liu G, Ren W, Ye Y
and Sun T: PI3K/Akt inhibitor partly decreases TNF-α-induced
activation of fibroblast-like synoviocytes in osteoarthritis. J
Orthop Surg Res. 14:4252019. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Porée B, Kypriotou M, Chadjichristos C,
Beauchef G, Renard E, Legendre F, Melin M, Gueret S, Hartmann DJ,
Malléin-Gerin F, et al: Interleukin-6 (IL-6) and/or soluble IL-6
receptor down-regulation of human type II collagen gene expression
in Articular chondrocytes requires a decrease of Sp1·Sp3 ratio and
of the binding activity of both factors to the COL2A1 promoter. J
Biol Chem. 283:4850–4865. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Rowan AD, Koshy PJ, Shingleton WD, Degnan
BA, Heath JK, Vernallis AB, Spaull JR, Life PF, Hudson K and
Cawston TE: Synergistic effects of glycoprotein 130 binding
cytokines in combination with interleukin-1 on cartilage collagen
breakdown. Arthritis Rheum. 44:1620–1632. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Cawston TE, Curry VA, Summers CA, Clark
IM, Riley GP, Life PF, Spaull JR, Goldring MB, Koshy PJ, Rowan AD
and Shingleton WD: The role of oncostatin M in animal and human
connective tissue collagen turnover and its localization within the
rheumatoid joint. Arthritis Rheum. 41:1760–1771. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Scanzello CR, Umoh E, Pessler F,
Diaz-Torne C, Miles T, Dicarlo E, Potter HG, Mandl L, Marx R, Rodeo
S, et al: Local cytokine profiles in knee osteoarthritis: Elevated
synovial fluid interleukin-15 differentiates early from end-stage
disease. Osteoarthritis Cartilage. 17:1040–1048. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Honsawek S, Deepaisarnsakul B, Tanavalee
A, Yuktanandana P, Bumrungpanichthaworn P, Malila S and Saetan N:
Association of the IL-6-174G/C gene polymorphism with knee
osteoarthritis in a Thai population. Genet Mol Res. 10:1674–1680.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Attur MG, Patel RN, Abramson SB and Amin
AR: Interleukin-17 up-regulation of nitric oxide production in
human osteoarthritis cartilage. Arthritis Rheum. 40:1050–1053.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Ryu JH, Yang S, Shin Y, Rhee J, Chun CH
and Chun JS: Interleukin-6 plays an essential role in
hypoxia-inducible factor 2α-induced experimental osteoarthritic
cartilage destruction in mice. Arthritis Rheum. 63:2732–2743. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Kwan Tat S, Padrines M, Théoleyre S,
Heymann D and Fortun Y: IL-6, RANKL, TNF-alpha/IL-1: Interrelations
in bone resorption pathophysiology. Cytokine Growth Factor Rev.
15:49–60. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Chenoufi HL, Diamant M, Rieneck K, Lund B,
Stein GS and Lian JB: Increased mRNA expression and protein
secretion of interleukin-6 in primary human osteoblasts
differentiated in vitro from rheumatoid and osteoarthritic bone. J
Cell Biochem. 81:666–678. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Sakao K, Takahashi KA, Arai Y, Saito M,
Honjo K, Hiraoka N, Asada H, Shin-Ya M, Imanishi J, Mazda O and
Kubo T: Osteoblasts derived from osteophytes produce interleukin-6,
interleukin-8, and matrix metalloproteinase-13 in osteoarthritis. J
Bone Miner Metab. 27:412–423. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Ding X, Cao Y, Xing Y, Ge S, Lin M and Li
J: TIMP-1 mediates inflammatory and immune response to IL-6 in
adult orbital xanthogranulomatous disease. Ocul Immunol Inflamm.
28:288–297. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
De Hooge ASK, van de Loo FAJ, Bennink MB,
Bennink MB, Arntz OJ, de Hooge P and van den Berg WB: Male IL-6
gene Knock Out mice developed more advanced osteoarthritis upon
aging. Osteoarthritis Cartilage. 13:66–73. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Steel JC, Waldmann TA and Morris JC:
Interleukin-15 biology and its therapeutic implications in cancer.
Trends Pharmacol Sci. 33:35–41. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Perera LP: Interleukin 15: Its role in
inflammation and immunity. Arch Immunol Ther Exp (Warsz).
48:457–464. 2000.PubMed/NCBI
|
|
72
|
Waldmann TA and Tagaya Y: The multifaceted
regulation of interleukin-15 expression and the role of this
cytokine in NK cell differentiation and host response to
intracellular pathogens. Annu Rev Immunol. 17:19–49. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Baslund B, Tvede N, Danneskiold-Samsoe B,
Larsson P, Panayi G, Petersen J, Petersen LJ, Beurskens FJ,
Schuurman J, van de Winkel JG, et al: Targeting interleukin-15 in
patients with rheumatoid arthritis: A proof-of-concept study.
Arthritis Rheum. 52:2686–2692. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Mcinnes IB, al-Mughales J, Field M, Leung
BP, Huang FP, Dixon R, Sturrock RD, Wilkinson PC and Liew FY: The
role of interleukin-15 in T-cell migration and activation in
rheumatoid arthritis. Nat Med. 2:175–182. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Tao Y, Qiu X, Xu C, Sun B and Shi C:
Expression and correlation of matrix metalloproteinase-7 and
interleukin-15 in human osteoarthritis. Int J Clin Exp Pathol.
8:9112–9118. 2015.PubMed/NCBI
|
|
76
|
Santos Savio A, Machado Diaz AC, Chico
Capote A, Miranda Navarro J, Rodríguez Alvarez Y, Bringas Pérez R,
Estévez del Toro M and Guillen Nieto GE: Differential expression of
pro-inflammatory cytokines IL-15Ralpha, IL-15, IL-6 and TNFalpha in
synovial fluid from rheumatoid arthritis patients. BMC
Musculoskelet Disord. 16:512015. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Badolato R, Ponzi AN, Millesimo M,
Notarangelo LD and Musso T: Interleukin-15 (IL-15) induces IL-8 and
monocyte chemotactic protein 1 production in human monocytes.
Blood. 90:2804–2809. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Sun JM, Sun LZ, Liu J, Su BH and Shi L:
Serum interleukin-15 levels are associated with severity of pain in
patients with knee osteoarthritis. Dis Markers. 35:203–206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Chang SH and Dong C: Signaling of
interleukin-17 family cytokines in immunity and inflammation. Cell
Signal. 23:1069–1075. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Zhang X, Angkasekwinai P, Dong C and Tang
H: Structure and function of interleukin-17 family cytokines.
Protein Cell. 2:26–40. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Korn T, Bettelli E, Oukka M and Kuchroo
VK: IL-17 and Th17 cells. Annu Rev Immunol. 27:485–517. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Pawłowska J, Mikosik A, Soroczynska-Cybula
M, Jóźwik A, Łuczkiewicz P, Mazurkiewicz S, Lorczyński A, Witkowski
JM and Bryl E: Different distribution of CD4 and CD8 T cells in
synovial membrane and peripheral blood of rheumatoid arthritis and
osteoarthritis patients. Folia Histochem Cytobiol. 47:627–632.
2009.PubMed/NCBI
|
|
83
|
Ishii H, Tanaka H, Katoh K, Nakamura H,
Nagashima M and Yoshino S: Characterization of infiltrating T cells
and Th1/Th2-type cytokines in the synovium of patients with
osteoarthritis. Osteoarthritis Cartilage. 10:277–281. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
84
|
Suurmond J, Dorjée AL, Boon MR, Knol EF,
Huizinga TW, Toes RE and Schuerwegh AJ: Mast cells are the main
interleukin 17-positive cells in anticitrullinated protein
antibody-positive and -negative rheumatoid arthritis and
osteoarthritis synovium. Arthritis Res Ther. 13:R1502011.
View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Liu Y, Peng H, Meng Z and Wei M:
Correlation of IL-17 Level in synovia and severity of knee
osteoarthritis. Med Sci Monit. 21:1732–1736. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
86
|
Koenders MI, Marijnissen RJ, Devesa I,
Lubberts E, Joosten LA, Roth J, van Lent PL, van de Loo FA and van
den Berg WB: Tumor necrosis factor-interleukin-17 interplay induces
S100A8,interleukin-1β,and matrix metalloproteinases, and drives
irreversible cartilage destruction in murine arthritis: Rationale
for combination treatment during arthritis. Arthritis Rheum.
63:2329–2339. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Honorati MC, Cattini L and Facchini A:
VEGF production by osteoarthritic chondrocytes cultured in
micromass and stimulated by IL-17 and TNF-alpha. Connect Tissue
Res. 48:239–245. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Honorati MC, Neri S, Cattini L and
Facchini A: Interleukin-17, a regulator of angiogenic factor
release by synovial fibroblasts. Osteoarthritis Cartilage.
14:345–352. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Lubberts E, Joosten LA, van de Loo FA, van
den Gersselaar LA and van den Berg WB: Reduction of
interleukin-17-induced inhibition of chondrocyte proteoglycan
synthesis in intact murine articular cartilage by interleukin-4.
Arthritis Rheum. 43:1300–1306. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Okamura H, Tsutsui H, Komatsu T, Yutsudo
M, Hakura A, Tanimoto T, Torigoe K, Okura T, Nukada Y, Hattori K,
et al: Cloning of a new cytokine that induces IFN-gamma production
by T cells. Nature. 378:88–91. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Ghayur T, Banerjee S, Hugunin M, Butler D,
Herzog L, Carter A, Quintal L, Sekut L, Talanian R, Paskind M, et
al: Caspase-1 processes IFN-gamma-inducing factor and regulates
LPS-induced IFN-gamma production. Nature. 386:619–623. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
92
|
Malemud CJ: Cytokines as therapeutic
targets for osteoarthritis. BioDrugs. 18:23–35. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
93
|
Joosten LA, Radstake TR, Lubberts E, van
den Bersselaar LA, van Riel PL, van Lent PL, Barrera P and van den
Berg WB: Association of interleukin-18 expression with enhanced
levels of both interleukin-1beta and tumor necrosis factor alpha in
knee synovial tissue of patients with rheumatoid arthritis.
Arthritis Rheum. 48:339–347. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
94
|
Joosten LAB, Smeets RL, Koenders MI, van
den Bersselaar LA, Helsen MM, Oppers-Walgreen B, Lubberts E,
Iwakura Y, van de Loo FA and van den Berg WB: Interleukin-18
promotes joint inflammation and induces interleukin-1-driven
cartilage destruction. Am J Pathol. 165:959–967. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Inoue H, Hiraoka K, Hoshino T, Okamoto M,
Iwanaga T, Zenmyo M, Shoda T, Aizawa H and Nagata K: High levels of
serum IL-18 promote cartilage loss through suppression of aggrecan
synthesis. Bone. 42:1102–1110. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
John T, Kohl B, Mobasheri A, Ertel W and
Shakibaei M: Interleukin-18 induces apoptosis in human articular
chondrocytes. Histol Histopathol. 22:469–482. 2007.PubMed/NCBI
|
|
97
|
Dai SM, Shan ZZ, Nishioka K and Yudoh K:
Implication of interleukin 18 in production of matrix
metalloproteinases in Articular chondrocytes in arthritis: Direct
effect on chondrocytes may not be pivotal. Ann Rheum Dis.
64:735–742. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Powers R, Garrett DS, March CJ, Frieden
EA, Gronenborn AM and Clore GM: The high-resolution,
three-dimensional solution structure of human interleukin-4
determined by multidimensional heteronuclear magnetic resonance
spectroscopy. Biochemistry. 32:6744–6762. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Wlodawer A, Pavlovsky A and Gustchina A:
Crystal structure of human recombinant interleukin-4 at 2.25 A
resolution. FEBS Lett. 309:59–64. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Carr C, Aykent S, Kimack NM and Levine AD:
Disulfide assignments in recombinant mouse and human interleukin 4.
Biochemistry. 30:1515–1523. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Mueller TD, Zhang JL, Sebald W and Duschl
A: Structure, binding, and antagonists in the IL-4/IL-13 receptor
system. Biochim Biophys Acta. 1592:237–250. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
102
|
Schlaak JF, Pfers I, Meyer Zum
Büschenfelde KH and Märker-Hermann E: Different cytokine profiles
in the synovial fluid of patients with osteoarthritis, rheumatoid
arthritis and seronegative spondylarthropathies. Clin Exp
Rheumatol. 14:155–162. 1996.PubMed/NCBI
|
|
103
|
Brown MA and Hural J: Functions of IL-4
and control of its expression. Crit Rev Immunol. 17:1–32. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Wang P, Wu P, Siegel MI, Egan RW and
Billah MM: Interleukin (IL)-10 inhibits nuclear factor kappa B (NF
kappa B) activation in human monocytes. IL-10 and IL-4 suppress
cytokine synthesis by different mechanisms. J Biol Chem.
270:9558–9563. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
te Velde AA, Huijbens RJ, Heije K, de
Vries JE and Figdor CG: Interleukin-4 (IL-4) inhibits secretion of
IL-1beta, tumor necrosis factor alpha, and human IL-6 by human
monocytes. Blood. 76:1392–1397. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Paul WE: Interleukin-4: A prototypic
immunoregulatory lymphokine. Blood. 77:1859–1870. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
Vannier E, Miller MC and Dinarello CA:
Coordinated antiinflammatory effects of interleukin 4: Interleukin
4 suppresses interleukin 1 production but up-regulates gene
expression and synthesis of interleukin 1 receptor antagonist. Proc
Natl Acad Sci USA. 89:4076–4080. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Hart PH, Vitti GF, Burgess DR, Whitty GA,
Piccoli DS and Hamilton JA: Potential anti-inflammatory effects of
interleukin 4: Suppression of human monocyte tumor necrosis factor
alpha, interleukin 1, and prostaglandin E2. Proc Natl Acad Sci USA.
86:3803–3807. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Yeh LA, Augustine AJ, Lee P, Riviere LR
and Sheldon A: Interleukin-4, an inhibitor of cartilage breakdown
in bovine articular cartilage explants. J Rheumatol. 22:1740–1746.
1995.PubMed/NCBI
|
|
110
|
Van Meegeren ME, Roosendaal G, Jansen NW,
Wenting MJ, van Wesel AC, van Roon JA and Lafeber FP: IL-4 alone
and in combination with IL-10 protects against blood-Induced
cartilage damage. Osteoarthritis Cartilage. 20:764–772. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
111
|
van Lent PL, Holthuysen AE, Slöetjes A,
Lubberts E and van den Berg WB: Local overexpression of adeno-viral
IL-4 protects cartilage from metallo proteinase-induced destruction
during immune complex-mediated arthritis by preventing activation
of pro-MMPs. Osteoarthritis Cartilage. 10:234–243. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Doi H, Nishida K, Yorimitsu M, Komiyama T,
Kadota Y, Tetsunaga T, Yoshida A, Kubota S, Takigawa M and Ozaki T:
Interleukin-4 downregulates the cyclic tensile stress-induced
matrix metalloproteinases-13 and cathepsin B expression by rat
normal chondrocytes. Acta Medica Okayama. 62:119–126.
2008.PubMed/NCBI
|
|
113
|
Yorimitsu M, Nishida K, Shimizu A, Doi H,
Miyazawa S, Komiyama T, Nasu Y, Yoshida A, Watanabe S and Ozaki T:
Intra-articular injection of interleukin-4 decreases nitric oxide
production by chondrocytes and ameliorates subsequent destruction
of cartilage in instability-induced osteoarthritis in rat knee
joints. Osteoarthritis Cartilage. 16:764–771. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
von Kaeppler EP, Wang Q, Raghu H, Bloom
MS, Wong H and Robinson WH: Interleukin 4 promotes
anti-inflammatory macrophages that clear cartilage debris and
inhibits osteoclast development to protect against osteoarthritis.
Clin Immunol. 229:1087842021. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Shah SS and Mithoefer K: Current
Applications of growth factors for knee cartilage repair and
osteoarthritis treatment. Curr Rev Musculoskelet Med. 13:641–650.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
McAlindon TE, Teale JD and Dieppe PA:
Levels of insulin related growth factor 1 in osteoarthritis of the
knee. Ann Rheum Dis. 52:229–231. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
117
|
McQuillan DJ, Handley CJ, Campbell MA,
Bolis S, Milway VE and Herington AC: Stimulation of proteoglycan
biosynthesis by serum and insulin-like growth factor-I in cultured
bovine articular cartilage. Biochem J. 240:423–430. 1986.
View Article : Google Scholar : PubMed/NCBI
|
|
118
|
van Osch GJ, van den Berg WB, Hunziker EB,
Hunziker EB and Häuselmann HJ: Differential effects of IGF-1 and
TGF beta-2 on the assembly of proteoglycans in pericellular and
territorial matrix by cultured bovine articular chondrocytes.
Osteoarthritis Cartilage. 6:187–195. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Sah RL, Chen AC, Grodzinsky AJ and Trippel
SB: Differential effects of bFGF and IGF-I on matrix metabolism in
calf and adult bovine cartilage explants. Arch Biochem Biophys.
308:137–147. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Yaeger PC, Masi TL, de Ortiz JL, Binette
F, Tubo R and McPherson JM: Synergistic action of transforming
growth factor-beta and insulin-like growth factor-I induces
expression of type II collagen and aggrecan genes in adult human
articular chondrocytes. Exp Cell Res. 237:318–325. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Madry H, Kaul G, Cucchiarini M, Stein U,
Zurakowski D, Remberger K, Menger MD, Kohn D and Trippel SB:
Enhanced repair of articular cartilage defects in vivo by
transplanted chondrocytes overexpressing insulin-like growth factor
I (IGF-I). Gene Ther. 12:1171–1179. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Fortier LA, Mohammed HO, Lust G and Nixon
AJ: Insulin-like growth factor-I enhances cell-based repair of
articular cartilage. J Bone Joint Surg Br. 84:276–288. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Davies LC, Blain EJ, Gilbert SJ, Caterson
B and Duance VC: The potential of IGF-1 and TGFbeta1 for promoting
‘adult’ articular cartilage repair: An in vitro study. Tissue Eng
Part A. 14:1251–1261. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Longobardi L, O'Rear L, Aakula S,
Johnstone B, Shimer K, Chytil A, Horton WA, Moses HL and Spagnoli
A: Effect of IGF-I in the chondrogenesis of bone marrow mesenchymal
stem cells in the presence or absence of TGF-beta signaling. J Bone
Miner Res. 21:626–636. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Morisset S, Frisbie DD, Robbins PD, Nixon
AJ and McIlwraith CW: IL-1ra/IGF-1 gene therapy modulates repair of
microfractured chondral defects. Clin Orthop Relat Res.
462:221–228. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Mushtaq T, Bijman P, Ahmed SF and
Farquharson C: Insulin-like growth factor-I augments chondrocyte
hypertrophy and reverses glucocorticoid-mediated growth retardation
in fetal mice metatarsal cultures. Endocrinology. 145:2478–2486.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
127
|
Koike M, Yamanaka Y, Inoue M, Tanaka H,
Nishimura R and Seino Y: Insulin-like growth factor-1 rescues the
mutated FGF receptor 3 (G380R) expressing ATDC5 cells from
apoptosis through phosphatidylinositol 3-kinase and MAPK. J Bone
Miner Res. 18:2043–2051. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Fytili P, Giannatou E, Karachalios T,
Malizos K and Tsezou A: Interleukin-10G and interleukin-10R
microsatellite polymorphisms and osteoarthritis of the knee. Clin
Exp Rheumatol. 23:621–627. 2005.PubMed/NCBI
|
|
129
|
Jansen NWD, Roosendaal G, Hooiveld MJJ,
Bijlsma JW, van Roon JA, Theobald M and Lafeber FP: Interleukin-10
protects against blood-induced joint damage. Br J Haematol.
142:953–961. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Wang Y and Lou S: Direct protective effect
of interleukin-10 on articular chondrocytes in vitro. Chin Med J
(Engl). 114:723–725. 2001.PubMed/NCBI
|
|
131
|
Behrendt P, Preusse-Prange A, Klüter T,
Haake M, Rolauffs B, Grodzinsky AJ, Lippross S and Kurz B: IL-10
reduces apoptosis and extracellular matrix degradation after
injurious compression of mature articular cartilage. Osteoarthritis
Cartilage. 24:1981–1988. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Rudwaleit M, Yin Z, Siegert S, Grolms M,
Radbruch A, Braun J and Sieper J: Response to methotrexate in early
rheumatoid arthritis is associated with a decrease of T cell
derived tumour necrosis factor alpha, increase of interleukin 10,
and predicted by the initial concentration of interleukin 4. Ann
Rheum Dis. 59:311–314. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Zeng L, Kempf H, Murtaugh LC, Sato ME and
Lassar AB: Shh establishes an Nkx3.2/Sox9 autoregulatory loop that
is maintained by BMP signals to induce somitic chondrogenesis.
Genes Dev. 16:1990–2005. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Chimal-Monroy J, Rodriguez-Leon J, Montero
JA, Gañan Y, Macias D, Merino R and Hurle JM: Analysis of the
molecular cascade responsible for mesodermal limb chondrogenesis:
Sox genes and BMP signaling. Dev Biol. 257:292–301. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Horbelt D, Denkis A and Knaus P: A
portrait of transforming growth factor β superfamily signalling:
Background matters. Int J Biochem Cell Biol. 44:469–474. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
136
|
Massagué J, Blain SW and Lo RS: TGFbeta
signaling in growth control, cancer, and heritable disorders. Cell.
103:295–309. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
137
|
Siegel PM and Massagué J: Cytostatic and
apoptotic actions of TGF-beta in homeostasis and cancer. Nat Rev
Cancer. 3:807–821. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Santo VE, Gomes ME, Mano JF and Reis RL:
Controlled release strategies for bone, cartilage, and
osteochondral engineering-part I: Recapitulation of native tissue
healing and variables for the design of delivery systems. Tissue
Eng Part B Rev. 19:308–326. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Cals FL, Hellingman CA, Koevoet W,
Baatenburg de Jong RJ and van Osch GJ: Effects of transforming
growth factor-β subtypes on in vitro cartilage production and
mineralization of human bone marrow stromal-derived mesenchymal
stem cells. J Tissue Eng Regen Med. 6:68–76. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Fortier LA, Barker JU, Strauss EJ,
McCarrel TM and Cole BJ: The role of growth factors in cartilage
repair. Clin Orthop Relat Res. 469:2706–2715. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Zhang RK, Li GW, Zeng C, Lin CX, Huang LS,
Huang GX, Zhao C, Feng SY and Fang H: Mechanical stress contributes
to osteoarthritis development through the activation of
transforming growth factor beta 1 (TGF-β1). Bone Joint Res.
7:587–594. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Rosen DM, Stempien SA, Thompson AY,
Brennan JE, Ellingsworth LR and Seyedin SM: Differentiation of rat
mesenchymal cells by cartilage-inducing factor. Enhanced phenotypic
expression by dihydrocytochalasin B. Exp Cell Res. 165:127–138.
1986. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Seyedin SM, Thompson AY, Bentz H, Rosen
DM, McPherson JM, Conti A, Siegel NR, Galluppi GR and Piez KA:
Cartilage-inducing factor-A. Apparent identity to transforming
growth factor-beta. J Biol Chem. 261:5693–5695. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Kato Y, Iwamoto M, Koike T, Suzuki F and
Takano Y: Terminal differentiation and calcification in rabbit
chondrocyte cultures grown in centrifuge tubes: Regulation by
transforming growth factor beta and serum factors. Proc Natl Acad
Sci USA. 85:9552–9556. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Leonard CM, Fuld HM, Frenz DA, Downie SA,
Massagué J and Newman SA: Role of transforming growth factor-beta
in chondrogenic pattern formation in the embryonic limb:
Stimulation of mesenchymal condensation and fibronectin gene
expression by exogenenous TGF-beta and evidence for endogenous
TGF-beta-like activity. Dev Biol. 145:99–109. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Xu C, Oyajobi BO, Frazer A, Kozaci LD,
Russell RG and Hollander AP: Effects of growth factors and
interleukin-1 alpha on proteoglycan and type II collagen turnover
in bovine nasal and articular chondrocyte pellet cultures.
Endocrinology. 137:3557–3565. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Ito T, Sawada R, Fujiwara Y, Seyama Y and
Tsuchiya T: FGF-2 suppresses cellular senescence of human
mesenchymal stem cells by down-regulation of TGF-beta2. Biochem
Biophys Res Commun. 359:108–114. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Lin Y, Luo E, Chen X, Liu L, Qiao J, Yan
Z, Li Z, Tang W, Zheng X and Tian W: Molecular and cellular
characterization during chondrogenic differentiation of adipose
tissue-derived stromal cells in vitro and cartilage formation in
vivo. J Cell Mol Med. 9:929–939. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Bian L, Zhai DY, Tous E, Rai R, Mauck RL
and Burdick JA: Enhanced MSC chondrogenesis following delivery of
TGF-β3 from alginate microspheres within hyaluronic acid hydrogels
in-vitro and in vivo. Biomaterials. 2:6425–6434. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Bhang SH, Jeon JY, La WG, Seong JY, Hwang
JW, Ryu SE and Kim BS: Enhanced chondrogenic marker expression of
human mesenchymal stem cells by interaction with both TGF-β3 and
hyaluronic acid. Biotechnol Appl Biochem. 58:271–276. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
151
|
Barry F, Boynton RE, Liu B and Murphy JM:
Chondrogenic differentiation of mesenchymal stem cells from bone
marrow: Differentiation-dependent gene expression of matrix
components. Exp Cell Res. 268:189–200. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Dave K and Gomes VG: Interactions at
scaffold interfaces: Effect of surface chemistry, structural
attributes and bioaffinity. Mater Sci Eng C Mater Biol Appl.
105:1100782019. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Huang BJ, Hu JC and Athanasiou KA:
Cell-based tissue engineering strategies used in the clinical
repair of articular cartilage. Biomaterials. 98:1–22. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Yang J, Zhang YS, Yue K and Khademhosseini
A: Cell-laden hydrogels for osteochondral and cartilage tissue
engineering. Acta Biomater. 57:1–25. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Kim YG, Choi J and Kim K: Mesenchymal stem
cell-derived exosomes for effective cartilage tissue repair and
treatment of osteoarthritis. Biotechnol J. 15:e20000822020.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Pittenger MF, Mackay AM, Beck SC, Jaiswal
RK, Douglas R, Mosca JD, Moorman MA, Simonetti DW, Craig S and
Marshak DR: Multilineage potential of adult human mesenchymal stem
cells. Science. 284:143–147. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Traggiai E, Volpi S, Schena F, Gattorno M,
Ferlito F, Moretta L and Martini A: Bone marrow-derived mesenchymal
stem cells induce both polyclonal expansion and differentiation of
B cells isolated from healthy donors and systemic lupus
erythematosus patients. Stem Cells. 6:562–569. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Rasmusson I, Le Blanc K, Sundberg B and
Ringdén O: Mesenchymal stem cells stimulate antibody secretion in
human B cells. Scand J Immunol. 65:336–343. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Spaggiari GM, Capobianco A, Becchetti S,
Mingari MC and Moretta L: Mesenchymal stem cell-natural killer cell
interactions: Evidence that activated NK cells are capable of
killing MSCs, whereas MSCs can inhibit IL-2-induced NK-cell
proliferation. Blood. 107:1484–1490. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Liang X, Ding Y, Zhang Y, Tse HF and Lian
Q: Paracrine mechanisms of mesenchymal stem cell-based therapy:
Current status and perspectives. Cell Transplant. 23:1045–1059.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Théry C, Witwer KW, Aikawa E, Alcaraz MJ,
Anderson JD, Andriantsitohaina R, Antoniou A, Arab T, Archer F,
Atkin-Smith GK, et al: Minimal information for studies of
extracellular vesicles 2018 (MISEV2018): A position statement of
the international society for extracellular vesicles and update of
the MISEV2014 guidelines. J Extracell Vesicles. 7:15357502018.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Li X, Ellman M, Muddasani P, Wang JH,
Cs-Szabo G, van Wijnen AJ and Im HJ: Prostaglandin E2 and its
cognate EP receptors control human adult articular cartilage
homeostasis and are linked to the pathophysiology of
osteoarthritis. Arthritis Rheum. 60:513–523. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
163
|
Cosenza S, Ruiz M, Toupet K, Jorgensen C
and Noël D: Mesenchymal stem cells derived exosomes and
microparticles protect cartilage and bone from degradation in
osteoarthritis. Sci Rep. 7:162142017. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Song Y, Zhang TJ, Li Y and Gao Y:
Mesenchymal stem cells decrease M1/M2 ratio and alleviate
inflammation to improve limb ischemia in mice. Med Sci Monit.
26:e9232872020. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Toh WS, Zhang B, Lai RC and Lim SK: Immune
regulatory targets of mesenchymal stromal cell exosomes/small
extracellular vesicles in tissue regeneration. Cytotherapy.
20:1419–1426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Chang YH, Wu KC, Harn HJ, Lin SZ and Ding
DC: Exosomes and stem cells in degenerative disease diagnosis and
therapy. Cell Transplant. 27:349–363. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Wang Y, Yu D, Liu Z, Zhou F, Dai J, Wu B,
Zhou J, Heng BC, Zou XH, Ouyang H and Liu H: Exosomes from
embryonic mesenchymal stem cells alleviate osteoarthritis through
balancing synthesis and degradation of cartilage extracellular
matrix. Stem Cell Res Ther. 8:1892017. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Temenoff JS and Mikos AG: Review: Tissue
engineering for regeneration of articular cartilage. Biomaterials.
21:431–440. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Hutmacher DW: Scaffolds in tissue
engineering bone and cartilage. Biomaterials. 21:2529–2543. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Zuluaga M, Gregnanin G, Cencetti C, Di Meo
C, Gueguen V, Letourneur D, Meddahi-Pellé A, Pavon-Djavid G and
Matricardi P: PVA/Dextran hydrogel patches as delivery system of
antioxidant astaxanthin: A cardiovascular approach. Biomed Mater.
13:0150202017. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Jiang T, Heng S, Huang X, Zheng L, Kai D,
Loh XJ and Zhao J: Biomimetic
poly(poly(ε-caprolactone)-Polytetrahydrofuran urethane) based
nanofibers enhanced chondrogenic differentiation and cartilage
regeneration. J Biomed Nanotechnol. 15:1005–1017. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Fasolino I, Raucci MG, Soriente A, Demitri
C, Madaghiele M, Sannino A and Ambrosio L: Osteoinductive and
anti-inflammatory properties of chitosan-based scaffolds for bone
regeneration. Mater Sci Eng C Mater Biol Appl. 105:1100462019.
View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Bhardwaj N, Singh YP and Mandal BB: Silk
fibroin scaffold-based 3D Co-culture model for modulation of
chondrogenesis without hypertrophy via reciprocal Cross-talk and
paracrine signaling. ACS Biomater Sci Eng. 5:5240–5254. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Campo GM, Avenoso A, Campo S, D'Ascola A,
Traina P and Calatroni A: Chondroitin-4-sulphate inhibits NF-kB
translocation and caspase activation in collagen-induced arthritis
in mice. Osteoarthritis Cartilage. 16:1474–1483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
175
|
Avenoso A, D'Ascola A, Scuruchi M,
Mandraffino G, Calatroni A, Saitta A, Campo S and Campo GM:
Hyaluronan in the experimental injury of the cartilage: Biochemical
action and protective effects. Inflamm Res. 67:5–20. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
176
|
Chen L, Liu J, Guan M, Zhou T, Duan X and
Xiang Z: Growth factor and its polymer scaffold-based delivery
system for cartilage tissue engineering. Int J Nanomedicine.
15:6097–6111. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Park E, Hart ML, Rolauffs B, Stegemann JP
and T Annamalai R: Bioresponsive microspheres for on-demand
delivery of anti-inflammatory cytokines for articular cartilage
repair. J Biomed Mater Res A. 108:722–733. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
178
|
Moutos FT, Glass KA, Compton SA, Ross AK,
Gersbach CA, Guilak F and Estes BT: Anatomically shaped
tissue-engineered cartilage with tunable and inducible anticytokine
delivery for biological joint resurfacing. Proc Natl Acad Sci USA.
113:E4513–E4522. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Levinson C, Lee M, Applegate LA and
Zenobi-Wong M: An injectable heparin-conjugated hyaluronan scaffold
for local delivery of transforming growth factor β1 promotes
successful chondrogenesis. Acta Biomater. 99:168–180. 2019.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Armiento AR, Stoddart MJ, Alini M and
Eglin D: Biomaterials for articular cartilage tissue engineering:
Learning from biology. Acta Biomater. 65:1–20. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Qasim M, Chae DS and Lee NY: Advancements
and frontiers in nano-based 3D and 4D scaffolds for bone and
cartilage tissue engineering. Int J Nanomedicine. 14:4333–4351.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Zhao Y, Li Y, Qu R, Chen X, Wang W, Qiu C,
Liu B, Pan X, Liu L, Vasilev K, et al: Cortistatin binds to TNF-α
receptors and protects against osteoarthritis. EBioMedicine.
41:556–570. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
Cui Z, Crane J, Xie H, Jin X, Zhen G, Li
C, Xie L, Wang L, Bian Q, Qiu T, et al: Halofuginone attenuates
osteoarthritis by inhibition of TGF-β activity and H-type vessel
formation in subchondral bone. Ann Rheum Dis. 75:1714–1721. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Chavez RD and Serra R: Scaffoldless
tissue-engineered cartilage for studying transforming growth factor
beta-mediated cartilage formation. Biotechnol Prog. 36:e28972020.
View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Goldring MB, Otero M, Plumb DA, Dragomir
C, Favero M, El Hachem K, Hashimoto K, Roach HI, Olivotto E, Borzì
RM and Marcu KB: Roles of inflammatory and anabolic cytokines in
cartilage metabolism: Signals and multiple effectors converge upon
MMP-13 regulation in osteoarthritis. Eur Cell Mater. 21:202–220.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Contentin R, Demoor M, Concari M, Desancé
M, Audigié F, Branly T and Galéra P: Comparison of the chondrogenic
potential of mesenchymal stem cells derived from bone marrow and
umbilical cord blood intended for cartilage tissue engineering.
Stem Cell Rev Rep. 16:126–143. 2020. View Article : Google Scholar : PubMed/NCBI
|














