Introduction
Acute myocardial infarction (MI) leads to myocardial
ischemia, and reperfusion of ischemic myocardium is the primary
treatment strategy for acute MI (1). Reperfusion therapy is a treatment that
reopens completely blocked blood vessels using thrombolytic drugs,
interventional therapy, or surgery, improving outcome for patients
with myocardial infarction (MI) (2). Although the ischemic myocardium can be
restored to normal perfusion, reperfusion can increase the
pathological process of patients with MI, resulting in tissue
damage and cardiac dysfunction, called myocardial
ischemia-reperfusion injury (MIRI) (3). Increased apoptosis of myocardial cells
during MI is accentuated by the subsequent reperfusion-induced
inflammatory response (4,5). MI caused by coronary artery
obstruction leads to myocardial tissue damage and ischemic death,
and is a serious threat to human health. A Sprague-Dawley rat model
is often used to study the inflammatory response following MI.
Macrophages are the main inflammatory cell type that trigger and
regulate the inflammatory response following MI (6,7).
Macrophages phagocytose apoptotic or necrotic cells and activate
the anti-inflammatory process by releasing cytokines (8). The quantity of inflammatory cytokines
released is proportional to cardiac function damage and the level
of myocardial apoptosis following ischemia (9). Notably, inflammatory responses are
activated during MI and reperfusion can significantly increase the
myocardial inflammatory response; MIRI is common in clinical
practice (10). An effective and
feasible therapeutic treatment that can protect ischemic
myocardium, limiting MI-associated damage and reducing MIRI, is
therefore necessary. Over the last decade, drug preconditioning has
received extensive attention in multiple diseases, including MI
(11–13).
Traditional Chinese medicines have been extensively
investigated because of their potential curative effects (14,15),
low toxicity, limited side effects and capacity to act on multiple
molecular interactions associated with disease (16). Crocin is an active component found
in Gardenia jasminoides Ellis and Crocus sativus L.
(17–19). Crocin reportedly exhibits
cardioprotective (20–22) and antitumor properties, and serves
an important pharmacological role in the treatment of central
nervous diseases (23). Xu et
al (24) revealed that crocin
suppresses the apoptosis of bovine endothelial cells through
increasing the ratio of Bcl-2/Bax, leading to an
anti-atherosclerotic role. In addition, Bcl-2/Bax mRNA expression
has been reported to be increased in response to crocin, thus
inhibiting cell apoptosis (25). A
previous study investigating platelet apoptosis induced by
Agkistrodon pallas venom demonstrated that crocin may
inhibit the expression of caspase-3 and caspase-9 (26). Furthermore, crocin has been shown to
effectively inhibit cardiovascular cell and platelet apoptosis
(27,28). However, the pharmacological
mechanism of crocin remains unclear.
The BTB-kelch protein family is involved in numerous
cellular functions, including transcriptional regulation,
cytoskeleton formation, ion channels, protein ubiquitination,
angiogenesis and apoptosis (29).
Kelch repeat and BTB domain-containing protein 7 (KBTBD7) is
located on chromosome 13q14.11, has a cDNA length of 3,008 bases,
an open reading frame of 2,052 bases and encodes a protein of 684
amino acid residues (30). The
KBTBD7 protein contains a conserved BTB domain and a series of
kelch repeats. Homologous proteins have been found in a range of
species, from yeast to fruit flies to mice, displaying a high level
of inter-species protein conservation. Fragment analysis has
demonstrated that the BTB, BTB and C-terminal kelch, and kelch
repeat domains exhibit transcriptional activation activity
(31). However, few cardiovascular
studies have reported on KBTBD7. Moreover, the role of KBTBD7 in
MIRI and whether crocin functions through KBTBD7 remain
unclear.
In the present study, a rat model of acute MI was
established to investigate the protective effect of crocin on acute
MI aggravated by reperfusion injuries. The interaction between
crocin and KBTBD7 was also examined. The present study has provided
the foundations for further MI research and the development of new
therapeutics.
Materials and methods
Animal model of MI
Clean-grade Sprague Dawley rats (male; age, 8–10
weeks; weight, 200–220 g; n=40) were purchased from Beijing Vital
River Laboratory Animal Technology Co., Ltd. Animals were housed at
21–25°C and 50–60% humidity, under a 12-h light/dark cycle, with
free access to food and water. Sprague-Dawley rats were randomly
divided into four groups (n=10): Sham operation group, MI model
group, MI + crocin (Amyjet Scientific, Inc.) low-dose group (100
mg/kg) and MI + crocin high-dose group (200 mg/kg). Rats in the MI
+ crocin low-dose group and MI + crocin high-dose group were
intraperitoneally injected with crocin once per day (volume 0.2
ml/100 g) for 14 days. Rats in the sham operation and MI groups
were intraperitoneally injected with an equal volume of normal
saline for 14 days. The rats were anesthetized 30 min after the
last administration by intraperitoneal injection of 2% sodium
pentobarbital (40 mg/kg). Following endotracheal intubation, an
artificial respirator was connected (tidal volume, 1.5–2 ml/100 g;
frequency, 48–54 times/min). The right common carotid artery was
carefully separated and the left ventricle was intubated. An
incision was made between ribs 3 and 4 on the left side of the
chest of rats. The pericardium was excised to completely expose the
heart. With the exception of the sham operation group, the left
anterior descending coronary artery was ligated. After ligation for
30 min, the ligature was loosened for reperfusion for 120 min. Left
ventricular end-diastolic pressure, left ventricular systolic
pressure and cardiac function were all evaluated following ischemia
and reperfusion in each group. At the end of the experiment, rats
were euthanized via carbon dioxide; rats were placed within a box
and 100% CO2 was used at a displacement rate of 30%
vol/min. After 10 min, the rats were observed to no longer be
breathing and had discolored eyes. Minimum CO2 flow was
maintained for a further 2 min following respiratory arrest. Animal
death was then determined. All animal experiments were approved by
the Ethics Committee of Zhongda Hospital Southeast University
(approval no. 190501017; Nanjing, China).
Cell culture
The mouse leukemia RAW264.7 cell line was purchased
from the American Type Culture Collection and seeded in RPMI 1640
media (Gibco; Thermo Fisher Scientific, Inc.) containing 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.). The cells
were cultured in an incubator at 37°C with 5% CO2. After
24 h, the culture medium was changed to serum-free medium
overnight. An inflammatory response model was established using
recombinant mouse high mobility group box 1 (rmHMGB1; 1 µg/ml; cat.
no. ab255799; Abcam) and recombinant mouse heat shock protein 60
(rmHSP60; 1 mol/ml; cat. no. ab92364; Abcam) for 24 h at 37°C.
Crocin (low dose, 25 µg/ml; high dose, 50 µg/ml) was used to
pretreat RAW264.7 cells 1 h before establishment of the
inflammatory response model. The cells in the control group were
not treated with crocin, rmHMGB1 or rmHSP60.
Cell transfection
RAW264.7 cells (2×105 cells/well) were
transfected with 50 nM small interfering RNA (si)-negative control
(NC; 5′-UUCUCCGAACGUGUCACGU-3′) and 50 nM si-KBTBD7
(5′-GGAUUAAUAUAGGCACCAU-3′), synthesized by Shanghai GenePharma
Co., Ltd. Lipofectamine® 2000 (Thermo Fisher Scientific,
Inc.) reagent was used for transfection according to the
manufacturer's protocol. The transfection was performed at 37°C for
24 h.
TUNEL assay
Cardiac tissue apoptosis was detected using the
TUNEL assay (cat. no.; 40307ES50; Shanghai Yeasen Biotechnology
Co., Ltd.) according to the manufacturer's protocol. Briefly,
RAW264.7 cells were fixed with 4% paraformaldehyde at 4°C for 25
min and permeabilized using 0.2% Triton X-100 at room temperature
for 5 min. After washing with PBS, the cells were stained with 50
µl TdT buffer at 37°C for 1 h. The nuclei were stained with 2 µg/ml
DAPI at room temperature for 5 min in the dark. Images were
captured using a confocal microscope and 10 randomly-selected
high-magnification fields were observed. The apoptotic rate was
calculated as follows: Apoptotic nuclei/total nuclei ×100.
Reverse transcription quantitative-PCR
(RT-qPCR)
Total RNA was extracted from rat myocardium and
RAW264.7 cells using TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.). An ultraviolet spectrophotometer was used
to determine the concentration and purity of the RNA. Total RNA was
reverse transcribed into cDNA using the following components:
High-Fidelity Script reverse transcriptase, dNTPs, primers, RNA, RT
buffer, dithiothreitol and RNase-free water (Invitrogen; Thermo
Fisher Scientific, Inc.). RT was performed at 37°C for 15 min
followed by 85°C for 5 sec. qPCR was subsequently performed using
the UltraSYBR Mixture (CoWin Biosciences) according to the
manufacturer's protocol. The following thermocycling conditions
were used for qPCR: Pre-denaturation at 95°C for 10 min; 35 cycles
of denaturation at 95°C for 15 sec, and annealing and elongation at
60°C for 1 min; followed by final extension at 95°C for 15 sec,
60°C for 1 min, 95°C for 15 sec and 60°C for 15 sec. The following
primer pairs were used for qPCR: KBTBD7 forward (F),
5′-AGCTCAGTTGTATCGGTAGCA-3′ and reverse (R),
5′-CCGGAATAAGGGTCGTAACAGA-3′; interleukin (IL)-1β F,
5′-GGTGTGTGACGTTCCCATTAGAC-3′ and R,
5′-CATGGAGAATATCACTTGTTGGTTGA-3′; IL-6 F,
5′-TAGTCCTTCCTACCCCAATTTCC-3′ and R, 5′-TTGGTCCTTAGCCACTCCTTC-3′;
tumor necrosis factor α (TNFα) F, 5′-AAGCCTGTAGCCCACGTCGTA-3′ and
R, 5′-GGCACCACTAGTTGGTTGTCTTTG-3′; Bax F,
5′-AGTGTCTCAGGCGAATTGGC-3′ and R, 5′-CACGGAAGAAGACCTCTCGG-3′; Bcl-2
F, 5′-ACTGAGTACCTGAACCGGCATC-3′ and R,
5′-GGAGAAATCAAACAGAGGTCGC-3′; and GAPDH F,
5′-AGGTCGGTGTGAACGGATTTG-3′ and R, 5′-TGTAGACCATGTAGTTGAGGTCA-3′.
mRNA expression levels were quantified using the 2−∆∆Cq
method (32) and normalized to the
internal reference gene GAPDH.
Western blotting
After 30 min of ischemia and 120 min of reperfusion,
100 mg myocardial tissue was immediately extracted from the
ischemic area in the left ventricle of the rats. Tissue was
subsequently washed with 1X PBS before tissue lysis solution was
added. The tissue was then incubated with RIPA buffer (Beyotime
Institute of Biotechnology) on ice for 30 min. Total protein
content was determined by bicinchoninic acid (BCA) assay (Thermo
Fisher Scientific, Inc.). In addition, RAW264.7 cells were
collected, cells were cleared on ice, and total protein was
extracted using RIPA buffer and was quantified using a BCA assay.
Total protein (20 µg) was separated by SDS-PAGE on 12% gels. After
blocking with 5% skim-milk at room temperature for 1 h, the PVDF
membranes were incubated with primary antibodies against HMGB1
(1:1,000; cat. no. ab227526; Abcam), HSP60 (1:1,000; cat no.
ab190828; Abcam), p38 (1:1,000: cat. no. 8690; Cell Signaling
Technology, Inc.), phosphorylated (p)-p38 (1:1,000; cat. no. 4511;
Cell Signaling Technology, Inc.), JNK (1:1,000; cat. no. 9252; Cell
Signaling Technology, Inc.), p-JNK (1:1,000; cat. no. 9251; Cell
Signaling Technology, Inc.), p65 (1:1,000; cat. no. 8242; Cell
Signaling Technology, Inc.), p-p65 (1:1,000; cat. no. 3033; Cell
Signaling Technology, Inc.) and GAPDH (1:1,000; cat. no. 5174; Cell
Signaling Technology, Inc.) overnight at 4°C. Horseradish
peroxidase-labeled secondary antibodies (1:3,000; cat. no. 7074;
Cell Signaling Technology, Inc.) were then added to incubate with
the membranes at room temperature for 2 h. Protein bands were
visualized using ECL (Shanghai Yeasen Biotechnology Co., Ltd.). The
ratio of gray values was used for semi-quantitative analysis with
GAPDH as the loading control using ImageJ software (version 1.8.0;
National Institutes of Health).
ELISA
The RAW 264.7 cell supernatant solution was
collected. ELISA was performed according to the manufacturers'
instructions. The levels of cytokines in each treatment group were
determined using the standard curve and optical density. The
following ELISA kits were used: IL-1β ELISA Kit [cat. no. EK201B/3;
Hangzhou Multi Sciences (Lianke) Biotech, Co., Ltd.], TNFα ELISA
Kit [cat. no. EK282/4; Hangzhou Multi Sciences (Lianke) Biotech,
Co., Ltd.], IL-6 ELISA Kit [cat. no. EK206/3; Hangzhou Multi
Sciences (Lianke) Biotech, Co., Ltd.], HSP60 ELISA Kit (cat. no.
EM1136; FineTest), and HMGB1 ELISA Kit (cat. no. EM0382;
FineTest).
Statistical analysis
Data were analyzed using SPSS 19.0 software (IBM
Corporation) from three independent experiments. Data are presented
as the mean ± standard deviation. Unpaired Student's t-test was
used for comparisons between two groups. One-way ANOVA and Tukey's
post-hoc test was used for comparisons between multiple groups
(33–35). P<0.05 was considered to indicate
a statistically significant difference.
Results
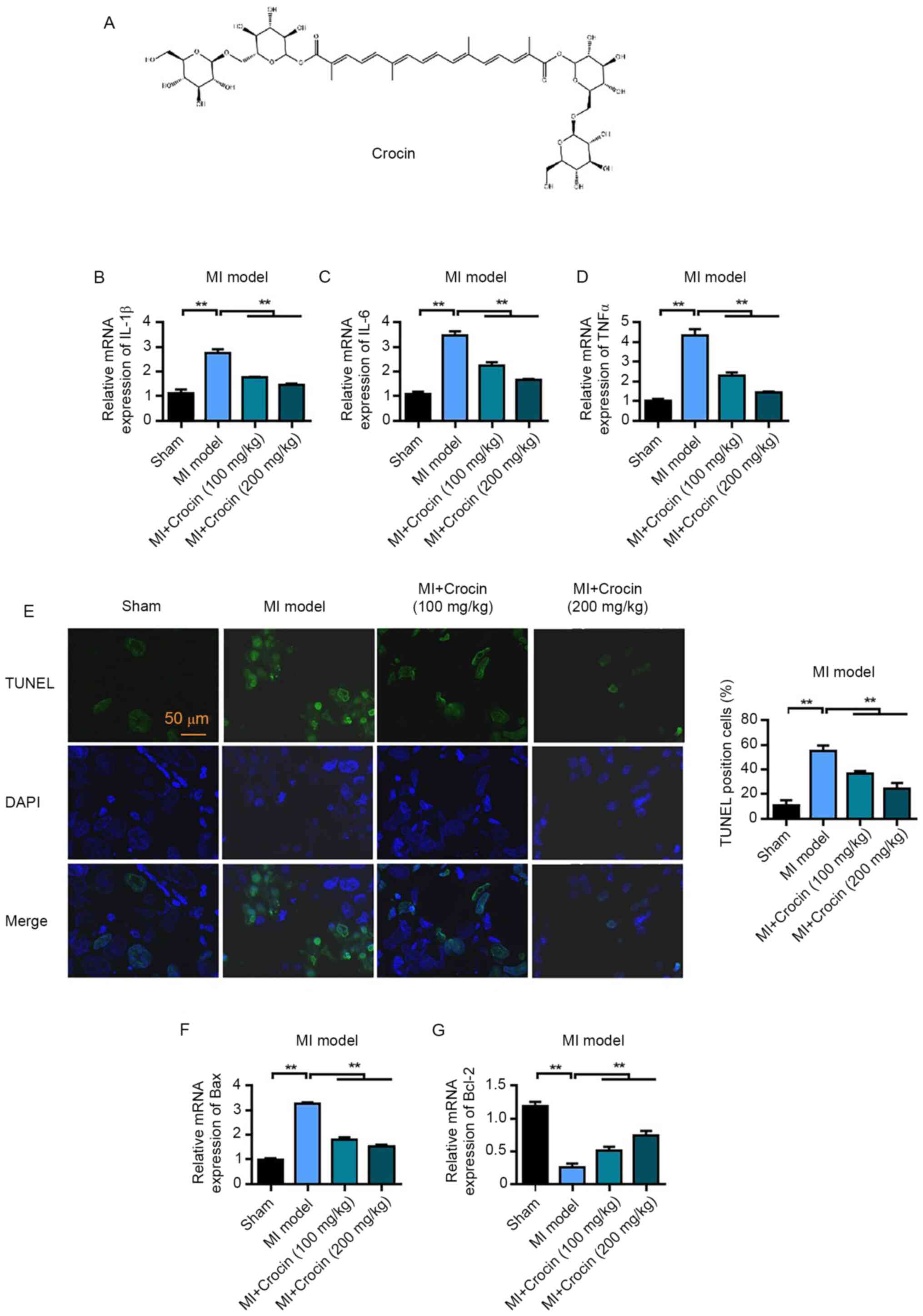
Crocin inhibits the inflammatory
response and myocardial apoptosis caused by MIRI
A rat model of MIRI was constructed to study the
pharmacological effects of crocin, the chemical structure of which
is shown in Fig. 1A. To assess the
inflammatory response and the degree of myocardial injury, mRNA
expression levels of IL-1β, IL-6 and TNFα in myocardial tissue were
determined. The mRNA expression levels of IL-1β, IL-6 and TNFα were
significantly increased in the MI group compared with those in the
sham group. However, the mRNA expression levels of IL-1β, IL-6 and
TNFα were significantly decreased following crocin treatment
compared with in the MI group (Fig.
1B-D). Furthermore, the TUNEL assay was used to detect
apoptosis. Apoptosis was observed following myocardial I/R in the
MI group, this was significant compared with the sham group;
however, the apoptotic rate in the low and high crocin treatment
groups was significantly reduced compared with that in the MI group
(Fig. 1E). The mRNA expression
levels of the apoptosis-related protein Bax and antiapoptotic
protein Bcl-2 were also detected. The mRNA expression levels of Bax
in the myocardial tissue of the MI group were significantly
increased compared with those in the sham group; however, Bax
expression was significantly reduced following treatment with high
and low doses of crocin compared with in the MI group (Fig. 1F). The mRNA expression levels of
Bcl-2 were significantly downregulated in the MI group compared
with those in the sham group, and were significantly upregulated
following high and low levels of crocin treatment compared with in
the MI group (Fig. 1G). These
results therefore suggested that crocin could inhibit myocardial
apoptosis caused by MIRI.
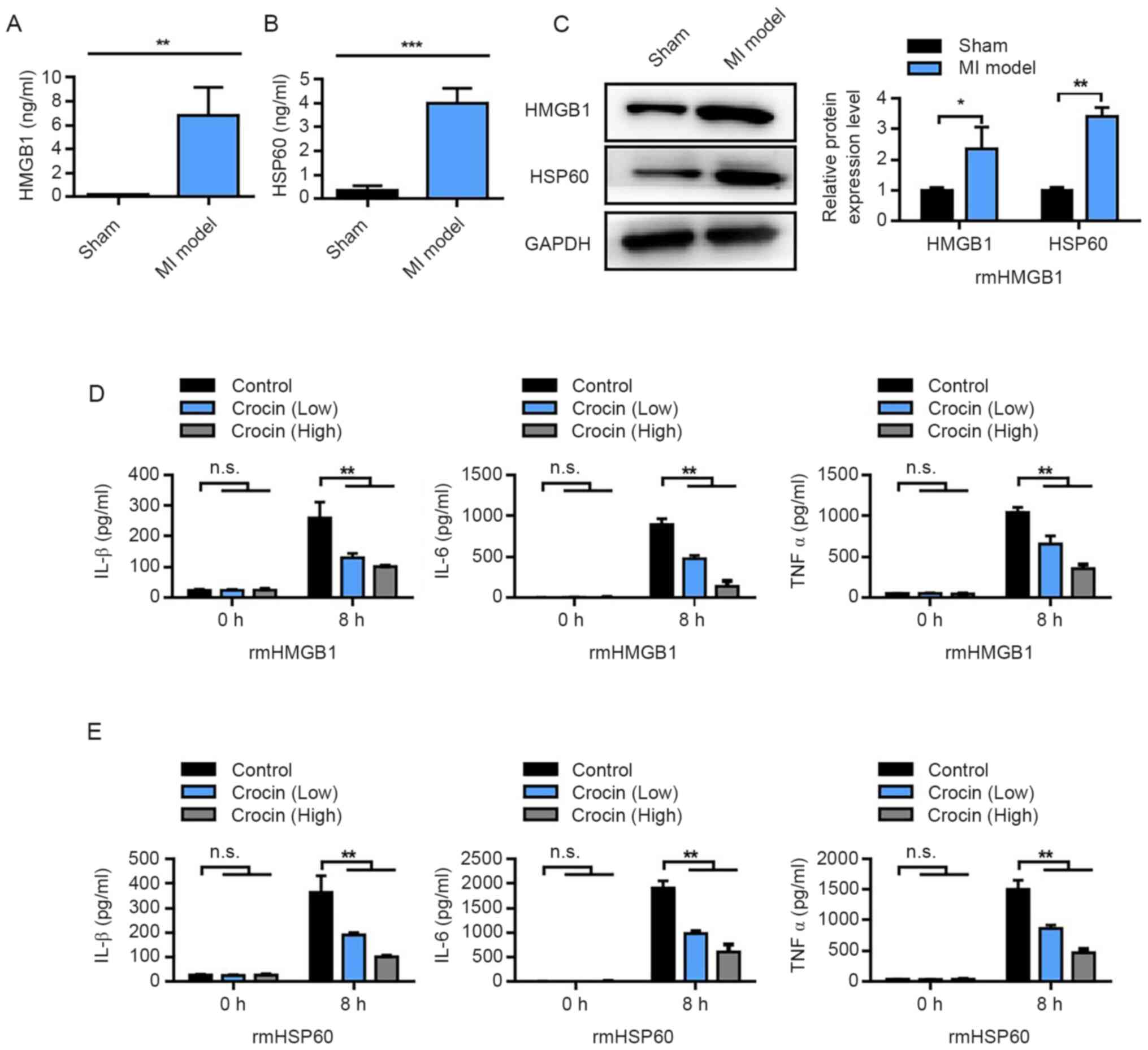
Crocin inhibits the production of
damage-associated molecular pattern (DAMP)-triggered inflammatory
cytokines
Changes in the levels of HMGB1 and HSP60 in
myocardial tissue following MI were determined. HMGB1 and HSP60
levels, measured using ELISA, were significantly increased in the
MI group compared with those in the sham group (Fig. 2A and B). Western blotting results
also demonstrated that the protein expression levels of HMGB1 and
HSP60 were significantly increased in the MI group compared with
those in the sham group (Fig. 2C).
Subsequently, rmHMGB1 and rmHSP60 were used to stimulate an
inflammatory response in RAW264. 7 cells and the effect of crocin
on these macrophages was observed. The results demonstrated that
the levels of IL-1β, IL-6 and TNFα following rmHMGB1 treatment were
significantly downregulated by the addition of crocin, in a
dose-dependent manner, compared with the control (no crocin
treatment) (Fig. 2D). These results
were further reflected upon RAW 264. 7 cells treatment with rmHSP60
(Fig. 2E). These results indicated
that crocin may inhibit the inflammatory response triggered by
HMGB1 and HSP60.
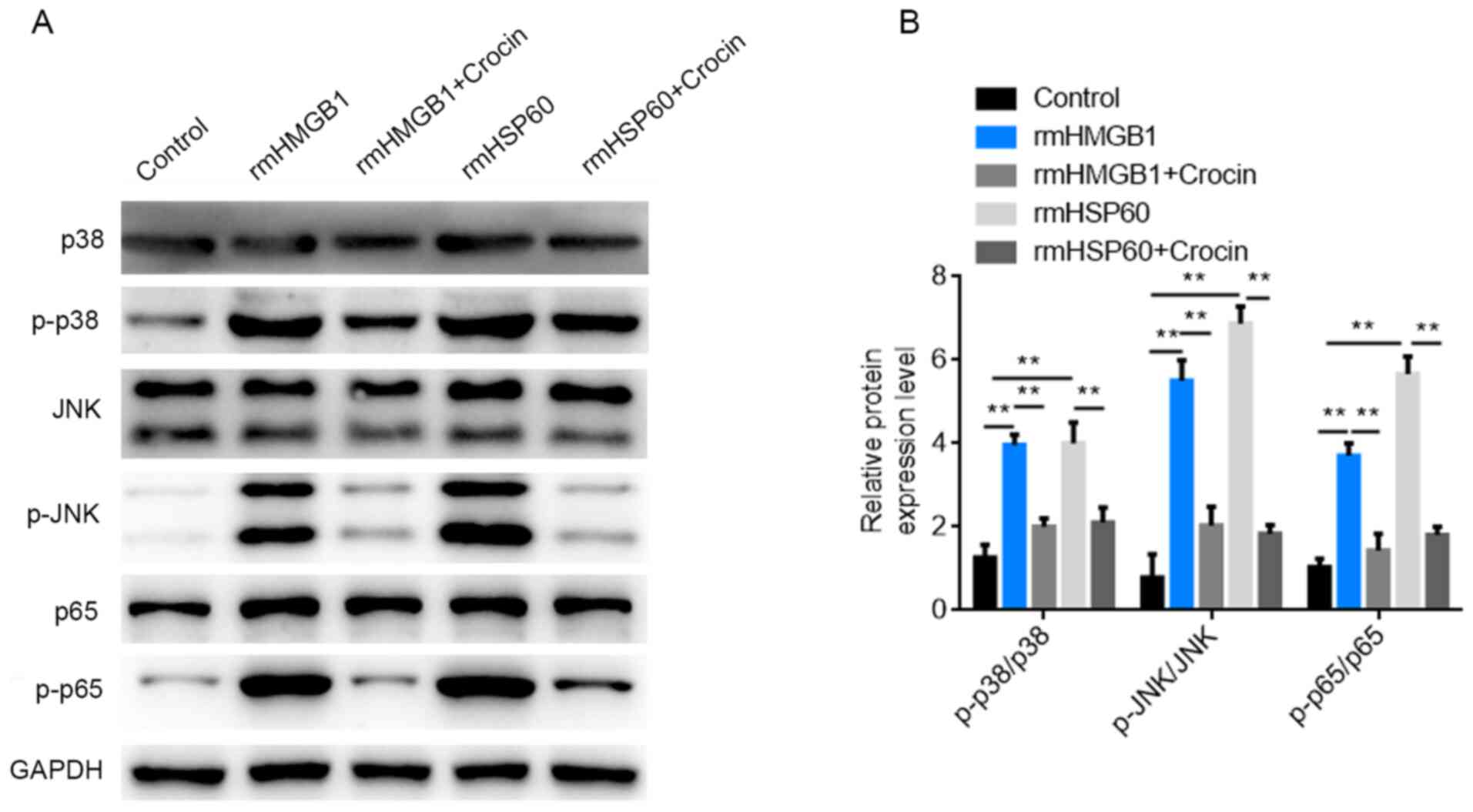
Activation of MAPK and NF-κB signaling
pathways is inhibited by crocin
Western blotting was used to detect the expression
levels of proteins involved in the MAPK and NF-κB signaling
pathways following treatment with rmHMGB1, rmHSP60 and crocin. The
results demonstrated that the protein expression levels of p-p38,
p-JNK and p-p65 were significantly increased following treatment
with rmHMGB1 and rmHSP60 compared with those in the control group.
However, the protein expression levels of p-p38, p-JNK and p-p65
were significantly reduced following high-dose crocin treatment
compared with cells that received no crocin (Fig. 3A and B). Therefore, the results
indicated that rmHMGB1 and rmHSP60 macrophage stimulation may
activate the p38 and NF-κB signaling pathways. The results also
suggested that crocin may suppress the production of inflammatory
cytokines through inhibition of the MAPK and NF-κB signaling
pathways.
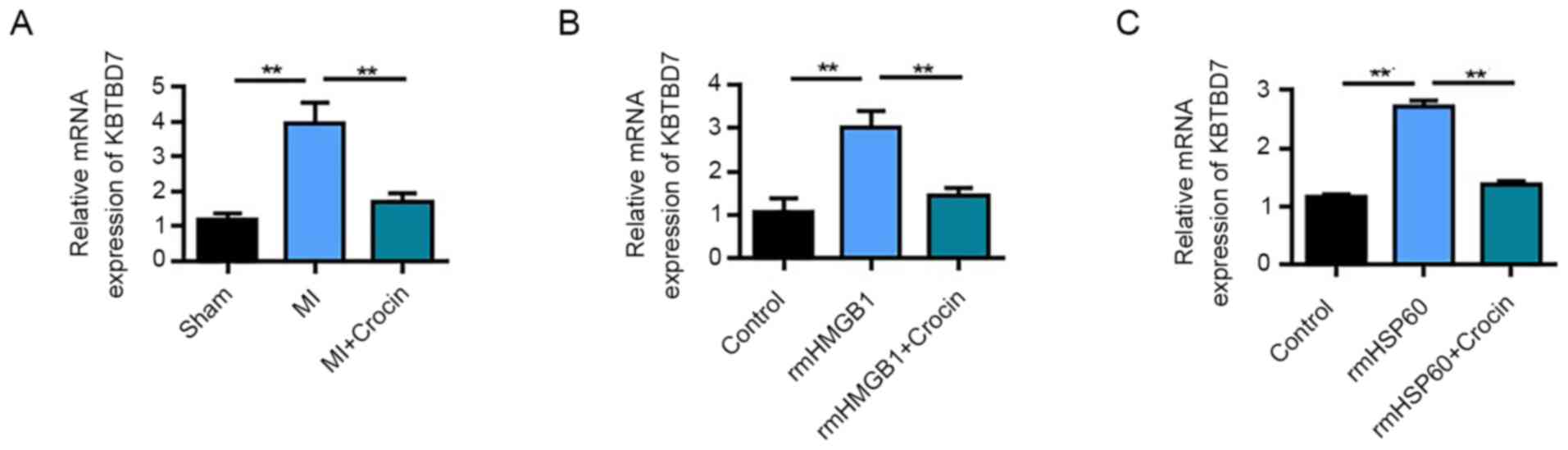
Crocin targets and inhibits
KBTBD7
KBTBD7 plays a crucial role in MI, especially in the
response to inflammation and regulation of myocardial function
(6). Therefore, the present study
investigated whether crocin could function through regulating
KBTBD7. qPCR detection demonstrated that KBTBD7 was significantly
upregulated following MI compared with in the sham group.
Furthermore, high-dose crocin treatment significantly downregulated
KBTBD7 mRNA expression levels compared with those in the MI group
(Fig. 4A). In RAW264.7 cells,
rmHMGB1 induction significantly upregulated KBTBD7 mRNA expression
levels compared with in the control, whereas crocin inhibited
KBTBD7 mRNA expression levels in rmHMGB1-treated cells (Fig. 4B). Similar results were also
determined in the rmHSP60-induced RAW264.7 cells (Fig. 4C). These results therefore suggested
that KBTBD7 is a target of crocin.
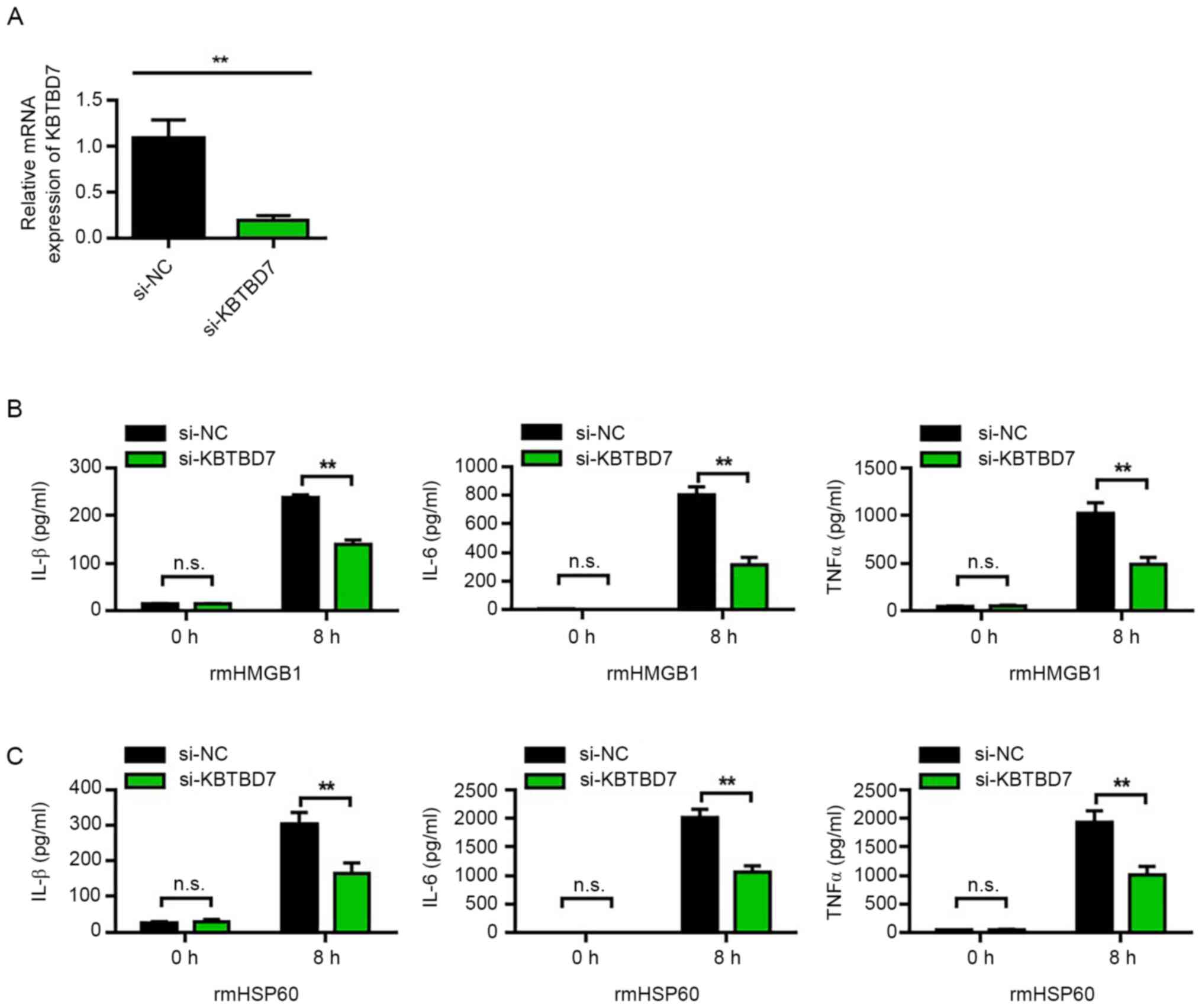
KBTBD7 knockdown reduces the
production of DAMP-induced inflammatory cytokines
The effect of KBTBD7 on DAMP-induced inflammatory
cytokines in RAW264.7 cells was further observed. The transfection
efficiency of si-KBTBD7 was first analyzed. The results
demonstrated that si-KBTBD7 could significantly reduce mRNA
expression levels of KBTBD7 compared with si-NC (Fig. 5A). Furthermore, the results
demonstrated that KBTBD7 knockdown significantly decreased the
levels of the rmHMGB1- and rmHSP60-induced inflammatory cytokines
compared with si-NC (Fig. 5B and
C).
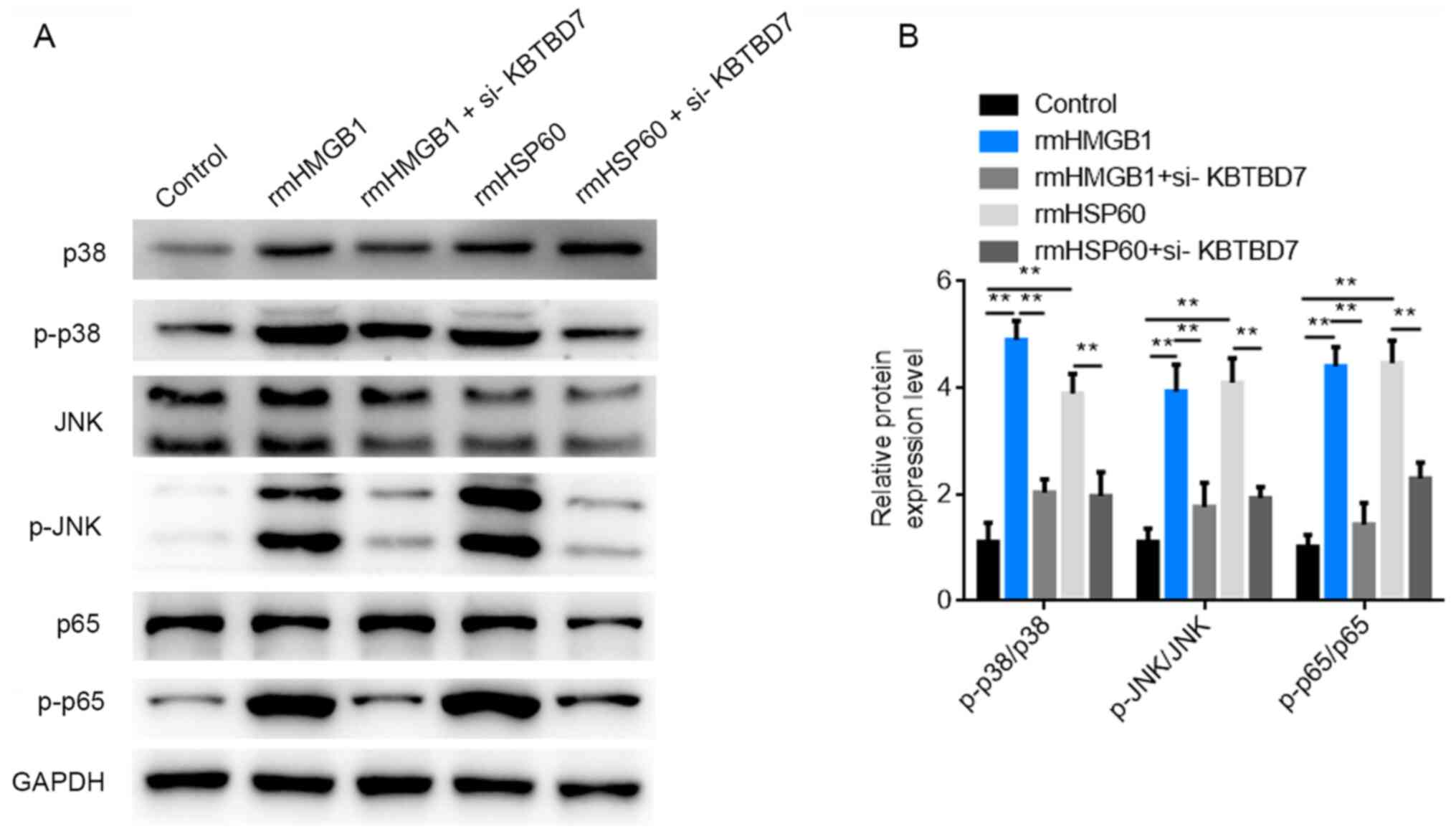
KBTBD7 knockdown reduces DAMP-induced
p38 and NF-κB signaling activation
The molecular mechanism by which KBTBD7 affects the
inflammatory response was investigated. Western blotting was used
to detect the expression levels of proteins associated with the
MAPK and NF-κB pathways following rmHMGB1, rmHSP60 and si-KBTBD7
treatments. The results demonstrated that the protein expression
levels of p-p38, p-JNK and p-p65 were significantly increased
following rmHMGB1 and rmHSP60 treatment compared with those in the
control group. However, KBTBD7 knockdown significantly reduced the
protein expression levels of p-p38, p-JNK and p-p65 compared with
the rmHMGB1 and rmHSP60 only treatments (Fig. 6A and B). These results indicated
that rmHMGB1 and rmHSP60 stimulation activated the p38 and NF-κB
signaling pathways, whereas KBTBD7 knockdown potentially inhibited
the production of inflammatory cytokines.
Discussion
Ischemic cardiomyopathy is a complex heart disease
worldwide (36). The primary and
most common cause of ischemic cardiomyopathy is coronary
atherosclerosis, which causes lumen stenosis or occlusion (37). Early reperfusion therapy is an
important strategy to save dying myocardial tissues; however,
reperfusion itself can also cause injury. Reperfusion can induce
the production of high levels of reactive oxygen species, resulting
in the peroxidation damage of proteins, lipids and chromosomes,
calcium overload, leukocyte accumulation, abnormal pH and ATP
energy metabolism disorder (38).
The inflammatory response plays an important role in this
phenomenon by triggering the release of numerous pro-inflammatory
factors, such as TNFα, IL-6 and IL-1β, which can lead to tissue
damage and in severe cases to apoptosis and necrosis (39). Therefore, inhibiting the
inflammatory response can reduce MIRI to a certain extent (40).
Crocin is a traditional Chinese medicine extract
with strong bioactivity (17).
Crocin has been reported to exhibit a variety of pharmacological
activities, such as antioxidative, anticancer, anti-inflammatory
and anti-atherosclerosis effects, and cardiovascular protection
(41). Crocin has also been shown
to act on mitochondria and inhibit mitochondrial dysfunction in
cardiovascular-related tissues and cells (42–44).
Crocin has previously been reported to reduce the hydrogen
peroxide-induced mitochondrial membrane potential of platelets
(44) and increase the release rate
of cytochrome c into rat heart tissues (42). Crocin may also act on cardiovascular
diseases via reducing platelet aggregation and apoptosis, oxidative
stress and cardiovascular apoptosis (45,46).
In the present study, the literature and preliminary experiments
were used to determine the concentrations of crocin to use in
animals and cells (47–50). The preliminary results demonstrated
that the current dose of crocin (200 mg/kg in mice) would reflect
the therapeutic effect of crocin on myocardial ischemia.
Following MIRI, DAMPs are released, which
consequently activate toll-like receptors expressed in macrophages,
triggering a series of inflammatory responses. Notably, HMGB1 and
HSP60 are major DAMP triggers (51,52).
HMGB1 is a highly conserved and widely present protein in the
nuclei of eukaryotic cells (53,54).
Under normal physiological conditions, HMBG1 has the function of a
nuclear-binding protein, whereas in the pathological state, it can
enter the intercellular space from the nucleus through passive
release and active secretion, thus triggering an inflammatory
response (55). As a
pro-inflammatory protein, HMGB1 plays an important role in MIRI and
is associated with inflammatory factors, such as TNFα, IL-6 and
inducible nitric oxide synthase (56). Inhibition of HMGB1 activity has
demonstrated the protective effects of HMGB1 on cardiopulmonary
function injury (57). The present
study further investigated the inhibitory effects of crocin on
HMGB1 and HSP60-induced inflammatory responses. The results
demonstrated that treatment with HMGB1 and HSP60 markedly
upregulated the levels of IL-1β, IL-6, and TNFα in RAW 264.7 cells.
Conversely, the levels of these pro-inflammatory cytokines were
significantly inhibited by crocin. The cardiac protective effect of
crocin may be related to the inhibition of inflammatory responses.
The release of inflammatory factors is closely related to the
occurrence and development of MIRI (58). During and after MIRI, a high level
of inflammatory cytokines, such as TNFα and IL-6, are released into
the blood, which can cause neutrophils to adhere to endothelial
cells (59). This phenomenon
results in no reflow after reperfusion and a large amount of oxygen
free radicals are generated in response (60). In addition, macrophages are
encouraged to secrete a large amount of inflammatory factors, which
eventually cause or aggravate the degree of MIRI (61). In the present study, crocin
treatment reduced the levels of TNFα and IL-6 in the serum of rats
with MIRI. This result suggested that crocin may protect against
MIRI in rats by reducing inflammatory responses.
The MAPK and NF-κB signaling pathways serve key
roles in inflammatory responses in MIRI; therefore, inhibition of
the MAPK and NF-κB signaling pathways serves an important role in
reducing the inflammatory response (62,63).
The present study revealed that crocin suppressed the production of
inflammatory cytokines, which may be mediated through inactivation
of the MAPK and NF-κB signaling pathways.
The BTB protein is a transcription factor in
eukaryotes, which is closely related to the development of various
multicellular organisms, including fruit flies and vertebrates.
Yang et al (6) demonstrated
that microRNA-21 inhibited the inflammatory response in the early
stages of MI by targeting KBTBD7, and inhibiting p38 and NF-κB
signaling activation, thereby preventing excessive scarring and
improving cardiac function. In the present study, crocin reduced
the inflammation caused by MIRI and the apoptosis of myocardial
cells. The protective effect of crocin may be realized by
inhibiting the expression of KBTBD7.
In conclusion, the present study demonstrated that
crocin protects against MIRI by inhibiting myocardial inflammatory
response and regulating the expression of inflammatory factors. The
anti-inflammatory effects of crocin were possibly related to the
inhibition of KBTBD7. The present study has provided a new
therapeutic approach for MIRI and lays the foundations for further
research on the myocardial-protective mechanism of crocin and the
development of novel treatments. Moreover, KBTBD7 may be a
potential target of MI therapy.
Acknowledgements
Not applicable.
Funding
Funding: No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CY designed the study and wrote the manuscript. CY,
ZC and QZ performed all experiments. ZC is responsible for data
analysis. CY and QZ confirm the authenticity of all the raw data.
All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All animal experiments were operated according to
the Guide for the Care and Use of Laboratory Animals (64) and were approved by the Ethics
Committee of Zhongda Hospital Southeast University (approval no.
190501017; Nanjing, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mui D and Zhang Y: Mitochondrial scenario:
Roles of mitochondrial dynamics in acute myocardial
ischemia/reperfusion injury. J Recept Signal Transduct Res. 41:1–5.
2021. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rout A, Tantry US, Novakovic M, Sukhi A
and Gurbel PA: Targeted pharmacotherapy for ischemia reperfusion
injury in acute myocardial infarction. Expert Opin Pharmacother.
21:1851–1865. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hausenloy DJ and Yellon DM: Myocardial
ischemia-reperfusion injury: A neglected therapeutic target. J Clin
Invest. 123:92–100. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Turer AT and Hill JA: Pathogenesis of
myocardial ischemia-reperfusion injury and rationale for therapy.
Am J Cardiol. 106:360–368. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Davidson SM, Ferdinandy P, Andreadou I,
Bøtker HE, Heusch G, Ibáñez B, Ovize M, Schulz R, Yellon DM,
Hausenloy DJ, et al: Multitarget strategies to reduce myocardial
ischemia/reperfusion injury: JACC review topic of the week. J Am
Coll Cardiol. 73:89–99. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yang L, Wang B, Zhou Q, Wang Y, Liu X, Liu
Z and Zhan Z: MicroRNA-21 prevents excessive inflammation and
cardiac dysfunction after myocardial infarction through targeting
KBTBD7. Cell Death Dis. 9:7692018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Frantz S and Nahrendorf M: Cardiac
macrophages and their role in ischaemic heart disease. Cardiovasc
Res. 102:240–248. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Lambert JM, Lopez EF and Lindsey ML:
Macrophage roles following myocardial infarction. Int J Cardiol.
130:147–158. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zhang Y, Wang Y, Xu J, Tian F, Hu S, Chen
Y and Fu Z: Melatonin attenuates myocardial ischemia-reperfusion
injury via improving mitochondrial fusion/mitophagy and activating
the AMPK-OPA1 signaling pathways. J Pineal Res. 66:e125422019.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ye B, Chen X, Dai S, Han J, Liang X, Lin
S, Cai X, Huang Z and Huang W: Emodin alleviates myocardial
ischemia/reperfusion injury by inhibiting gasdermin D-mediated
pyroptosis in cardiomyocytes. Drug Des Devel Ther. 13:975–990.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ryu SM, Kim HJ, Cho KR and Jo WM:
Myocardial protective effect of tezosentan, an endothelin receptor
antagonist, for ischemia-reperfusion injury in experimental heart
failure models. J Korean Med Sci. 24:782–788. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xi X, Liu N, Wang Q, Chu Y, Yin Z, Ding Y
and Lu Y: ACT001, a novel PAI-1 inhibitor, exerts synergistic
effects in combination with cisplatin by inhibiting PI3K/AKT
pathway in glioma. Cell Death Dis. 10:7572019. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang H, Zhong W, Zhao J, Zhang H, Zhang Q,
Liang Y, Chen S, Liu H, Zong S, Tian Y, et al: Oleanolic acid
inhibits epithelial-mesenchymal transition of hepatocellular
carcinoma by promoting iNOS dimerization. Mol Cancer Ther.
18:62–74. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhong W, Sun B, Gao W, Qin Y, Zhang H,
Huai L, Tang Y, Liang Y, He L, Zhang X, et al: Salvianolic acid A
targeting the transgelin-actin complex to enhance vasoconstriction.
EBioMedicine. 37:246–258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong W, Hou H, Liu T, Su S, Xi X, Liao Y,
Xie R, Jin G, Liu X, Zhu L, et al: Cartilage oligomeric matrix
protein promotes epithelial-mesenchymal transition by interacting
with transgelin in colorectal cancer. Theranostics. 10:8790–8806.
2020. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhong W, Yang W, Qin Y, Gu W, Xue Y, Tang
Y, Xu H, Wang H, Zhang C, Wang C, et al: 6-Gingerol stabilized the
p-VEGFR2/VE-cadherin/β-catenin/actin complex promotes microvessel
normalization and suppresses tumor progression. J Exp Clin Cancer
Res. 38:2852019. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen L, Qi Y and Yang X: Neuroprotective
effects of crocin against oxidative stress induced by
ischemia/reperfusion injury in rat retina. Ophthalmic Res.
54:157–168. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lee IA, Lee JH, Baek NI and Kim DH:
Antihyperlipidemic effect of crocin isolated from the fructus of
Gardenia jasminoides and its metabolite crocetin. Biol Pharm
Bull. 28:2106–2110. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hashemzaei M, Mamoulakis C, Tsarouhas K,
Georgiadis G, Lazopoulos G, Tsatsakis A, Shojaei Asrami E and
Rezaee R: Crocin: A fighter against inflammation and pain. Food
Chem Toxicol. 143:1115212020. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Hashemzaei M, Rezaee R, Nabatzehi M,
Tsarouhas K, Konstantinos Nikolouzakis T, Lazopoulos G, A Spandidos
D, Tsatsakis A and Shahraki J: Anti-hypertensive effect of crocin
and hesperidin combination in high-fat diet treated rats. Exp Ther
Med. 19:3840–3844. 2020.PubMed/NCBI
|
|
21
|
Rezaee R, Mahmoudi M, Abnous K, Zamani
Taghizadeh Rabe S, Tabasi N, Hashemzaei M and Karimi G: Cytotoxic
effects of crocin on MOLT-4 human leukemia cells. J Complement
Integr Med. 10:1–8. 2013.PubMed/NCBI
|
|
22
|
El-Baz FK, Aly HF and Abd-Alla HI: The
ameliorating effect of carotenoid rich fraction extracted from
Dunaliella salina microalga against inflammation-associated cardiac
dysfunction in obese rats. Toxicol Rep. 7:118–124. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tian Y, Pu X, Yu H, Ji A, Gao R, Hu Y, Xu
Z and Wang H: Genome-wide characterization and analysis of bHLH
transcription factors related to crocin biosynthesis in Gardenia
jasminoides ellis (rubiaceae). Biomed Res Int.
2020:29038612020. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu G, Gong Z, Yu W, Gao L, He S and Qian
Z: Increased expression ratio of Bcl-2/Bax is associated with
crocin-mediated apoptosis in bovine aortic endothelial cells. Basic
Clin Pharmacol Toxicol. 100:31–35. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ochiai T, Ohno S, Soeda S, Tanaka H,
Shoyama Y and Shimeno H: Crocin prevents the death of rat
pheochromyctoma (PC-12) cells by its antioxidant effects stronger
than those of alpha-tocopherol. Neurosci Lett. 362:61–64. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sebastin Santhosh M, Hemshekhar M,
Thushara RM, Devaraja S, Kemparaju K and Girish KS: Vipera russelli
venom-induced oxidative stress and hematological alterations:
Amelioration by crocin a dietary colorant. Cell Biochem Funct.
31:41–50. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ordoudi SA, Befani CD, Nenadis N, Koliakos
GG and Tsimidou MZ: Further examination of antiradical properties
of Crocus sativus stigmas extract rich in crocins. J Agric
Food Chem. 57:3080–3086. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Oruc S, Gönül Y, Tunay K, Oruc OA, Bozkurt
MF, Karavelioğlu E, Bağcıoğlu E, Coşkun KS and Celik S: The
antioxidant and antiapoptotic effects of crocin pretreatment on
global cerebral ischemia reperfusion injury induced by four vessels
occlusion in rats. Life Sci. 154:79–86. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Genau HM, Huber J, Baschieri F, Akutsu M,
Dötsch V, Farhan H, Rogov V and Behrends C: CUL3-KBTBD6/KBTBD7
ubiquitin ligase cooperates with GABARAP proteins to spatially
restrict TIAM1-RAC1 signaling. Mol Cell. 57:995–1010. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hu J, Yuan W, Tang M, Wang Y, Fan X, Mo X,
Li Y, Ying Z, Wan Y, Ocorr K, et al: KBTBD7, a novel human
BTB-kelch protein, activates transcriptional activities of SRE and
AP-1. BMB Rep. 43:17–22. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu YT, Liu F, Cao L, Xue L, Gu WT, Zheng
YZ, Tang H, Wang Y, Yao H, Zhang Y, et al: The KBTBD6/7-DRD2 axis
regulates pituitary adenoma sensitivity to dopamine agonist
treatment. Acta Neuropathol. 140:377–396. 2020. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Gao Y, Wang C, Wang Z, Li W, Liu Y, Shou S
and Chai Y: Semaphorin 3A contributes to sepsis-induced
immunosuppression by impairing CD4+ T cell anergy. Mol
Med Rep. 23:3022021. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li J, Wang X, Ma C, Xu S, Xu M, Yang J,
Wang R and Xue L: Dual PI3K/mTOR inhibitor NVP-BEZ235 decreases the
proliferation of doxorubicin-resistant K562 cells. Mol Med Rep.
23:3012021. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wu H, Tao Y, Zhang W, Wang G and Zhang Q:
circ-0000212 promotes cell proliferation of colorectal cancer by
sponging miR-491 and modulating FOXP4 expression. Mol Med Rep.
23:3002021. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liu NB, Wu M, Chen C, Fujino M, Huang JS,
Zhu P and Li XK: Novel molecular targets participating in
myocardial ischemia-reperfusion injury and cardioprotection.
Cardiol Res Pract. 2019:69351472019. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mozaffari MS, Liu JY, Abebe W and Baban B:
Mechanisms of load dependency of myocardial ischemia reperfusion
injury. Am J Cardiovasc Dis. 3:180–196. 2013.PubMed/NCBI
|
|
38
|
Fischesser DM, Bo B, Benton RP, Su H,
Jahanpanah N and Haworth KJ: Controlling reperfusion injury with
controlled reperfusion: Historical perspectives and new paradigms.
J Cardiovasc Pharmacol Ther. 26:504–523. 2021. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liu H, Wu X, Luo J, Wang X, Guo H, Feng D,
Zhao L, Bai H, Song M, Liu X, et al: Pterostilbene attenuates
astrocytic inflammation and neuronal oxidative injury after
ischemia-reperfusion by inhibiting NF-κB phosphorylation. Front
Immunol. 10:24082019. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang C, Sun H, Song Y, Ma Z, Zhang G, Gu X
and Zhao L: Pterostilbene attenuates inflammation in rat heart
subjected to ischemia-reperfusion: Role of TLR4/NF-κB signaling
pathway. Int J Clin Exp Med. 8:1737–1746. 2015.PubMed/NCBI
|
|
41
|
Farkhondeh T and Samarghandian S: The
effect of saffron (Crocus sativus L.) and its ingredients on
the management of diabetes mellitus and dislipidemia. Afr J Pharm
Pharmacol. 8:541–549. 2014. View Article : Google Scholar
|
|
42
|
Razavi BM, Hosseinzadeh H, Movassaghi AR,
Imenshahidi M and Abnous K: Protective effect of crocin on diazinon
induced cardiotoxicity in rats in subchronic exposure. Chem Biol
Interact. 203:547–555. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Sarshoori JR, Asadi MH and Mohammadi MT:
Neuroprotective effects of crocin on the histopathological
alterations following brain ischemia-reperfusion injury in rat.
Iran J Basic Med Sci. 17:895–902. 2014.PubMed/NCBI
|
|
44
|
Thushara RM, Hemshekhar M, Santhosh MS,
Jnaneshwari S, Nayaka SC, Naveen S, Kemparaju K and Girish KS:
Crocin, a dietary additive protects platelets from oxidative
stress-induced apoptosis and inhibits platelet aggregation. Mol
Cell Biochem. 373:73–83. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Vakili A, Einali MR and Bandegi AR:
Protective effect of crocin against cerebral ischemia in a
dose-dependent manner in a rat model of ischemic stroke. J Stroke
Cerebrovasc Dis. 23:106–113. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang X, Fan Z and Jin T: Crocin protects
against cerebral-ischemia-induced damage in aged rats through
maintaining the integrity of blood-brain barrier. Restor Neurol
Neurosci. 35:65–75. 2017.PubMed/NCBI
|
|
47
|
Naghizadeh B, Boroushaki MT, Vahdati
Mashhadian N and Mansouri MT: Protective effects of crocin against
cisplatin-induced acute renal failure and oxidative stress in rats.
Iran Biomed J. 12:93–100. 2008.PubMed/NCBI
|
|
48
|
He SY, Qian ZY, Tang FT, Wen N, Xu GL and
Sheng L: Effect of crocin on experimental atherosclerosis in quails
and its mechanisms. Life Sci. 77:907–921. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Roshankhah S, Salahshoor MR, Jalili F,
Karimi F, Sohrabi M and Jalili C: Crocin effects on the
nicotine-induce ovary injuries in female rat. Int J Life Sci Pharm
Res. 7:1–8. 2017.
|
|
50
|
Hussain MA, Abogresha NM, AbdelKader G,
Hassan R, Abdelaziz EZ and Greish SM: Antioxidant and
anti-inflammatory effects of crocin ameliorate doxorubicin-induced
nephrotoxicity in rats. Oxid Med Cell Longev. 2021:88417262021.
View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Khwaja B, Thankam FG and Agrawal DK:
Mitochondrial DAMPs and altered mitochondrial dynamics in OxLDL
burden in atherosclerosis. Mol Cell Biochem. 476:1915–1928. 2021.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yang H, Wang H and Andersson U: Targeting
Inflammation Driven by HMGB1. Front Immunol. 11:4842020. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Andrassy M, Volz HC, Riedle N, Gitsioudis
G, Seidel C, Laohachewin D, Zankl AR, Kaya Z, Bierhaus A,
Giannitsis E, et al: HMGB1 as a predictor of infarct transmurality
and functional recovery in patients with myocardial infarction. J
Intern Med. 270:245–253. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Ding HS, Yang J, Chen P, Yang J, Bo SQ,
Ding JW and Yu QQ: The HMGB1-TLR4 axis contributes to myocardial
ischemia/reperfusion injury via regulation of cardiomyocyte
apoptosis. Gene. 527:389–393. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Hu X, Fu W and Jiang H: HMGB1: A potential
therapeutic target for myocardial ischemia and reperfusion injury.
Int J Cardiol. 155:4892012. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Li Q, Xu M, Li Z, Li T, Wang Y, Chen Q,
Wang Y, Feng J, Yin X and Lu C: Maslinic acid attenuates
ischemia/reperfusion injury-induced myocardial inflammation and
apoptosis by regulating HMGB1-TLR4 axis. Front Cardiovasc Med.
8:7689472021. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Giallauria F, Cirillo P, D'agostino M,
Petrillo G, Vitelli A, Pacileo M, Angri V, Chiariello M and
Vigorito C: Effects of exercise training on high-mobility group
box-1 levels after acute myocardial infarction. J Card Fail.
17:108–114. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Vinten-Johansen J, Jiang R, Reeves JG,
Mykytenko J, Deneve J and Jobe LJ: Inflammation, proinflammatory
mediators and myocardial ischemia-reperfusion Injury. Hematol Oncol
Clin North Am. 21:123–145. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Hou M, Wu X, Zhao Z, Deng Q, Chen Y and
Yin L: Endothelial cell-targeting, ROS-ultrasensitive drug/siRNA
co-delivery nanocomplexes mitigate early-stage neutrophil
recruitment for the anti-inflammatory treatment of myocardial
ischemia reperfusion injury. Acta Biomater. 143:344–355. 2022.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Gao C, Liu Y, Yu Q, Yang Q, Li B, Sun L,
Yan W, Cai X, Gao E, Xiong L, et al: TNF-α antagonism ameliorates
myocardial ischemia-reperfusion injury in mice by upregulating
adiponectin. Am J Physiol Heart Circ Physiol. 308:H1583–H1591.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Cao J, Xie H, Sun Y, Zhu J, Ying M, Qiao
S, Shao Q, Wu H and Wang C: Sevoflurane post-conditioning reduces
rat myocardial ischemia reperfusion injury through an increase in
NOS and a decrease in phopshorylated NHE1 levels. Int J Mol Med.
36:1529–1537. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Xie C, Li X, Zhu J, Wu J, Geng S and Zhong
C: Magnesium isoglycyrrhizinate suppresses LPS-induced inflammation
and oxidative stress through inhibiting NF-κB and MAPK pathways in
RAW264.7 cells. Bioorg Med Chem. 27:516–524. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Wang HW, Liu HJ, Cao H, Qiao ZY and Xu YW:
Diosgenin protects rats from myocardial inflammatory injury induced
by ischemia-reperfusion. Med Sci Monit. 24:246–253. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
National Research Council (US) Committee
for the Update of the Guide for the Care and Use of Laboratory
Animals, . Guide for the care and use of laboratory animals. 8th
edition. Washington (DC): National Academies Press (US); 2011
|