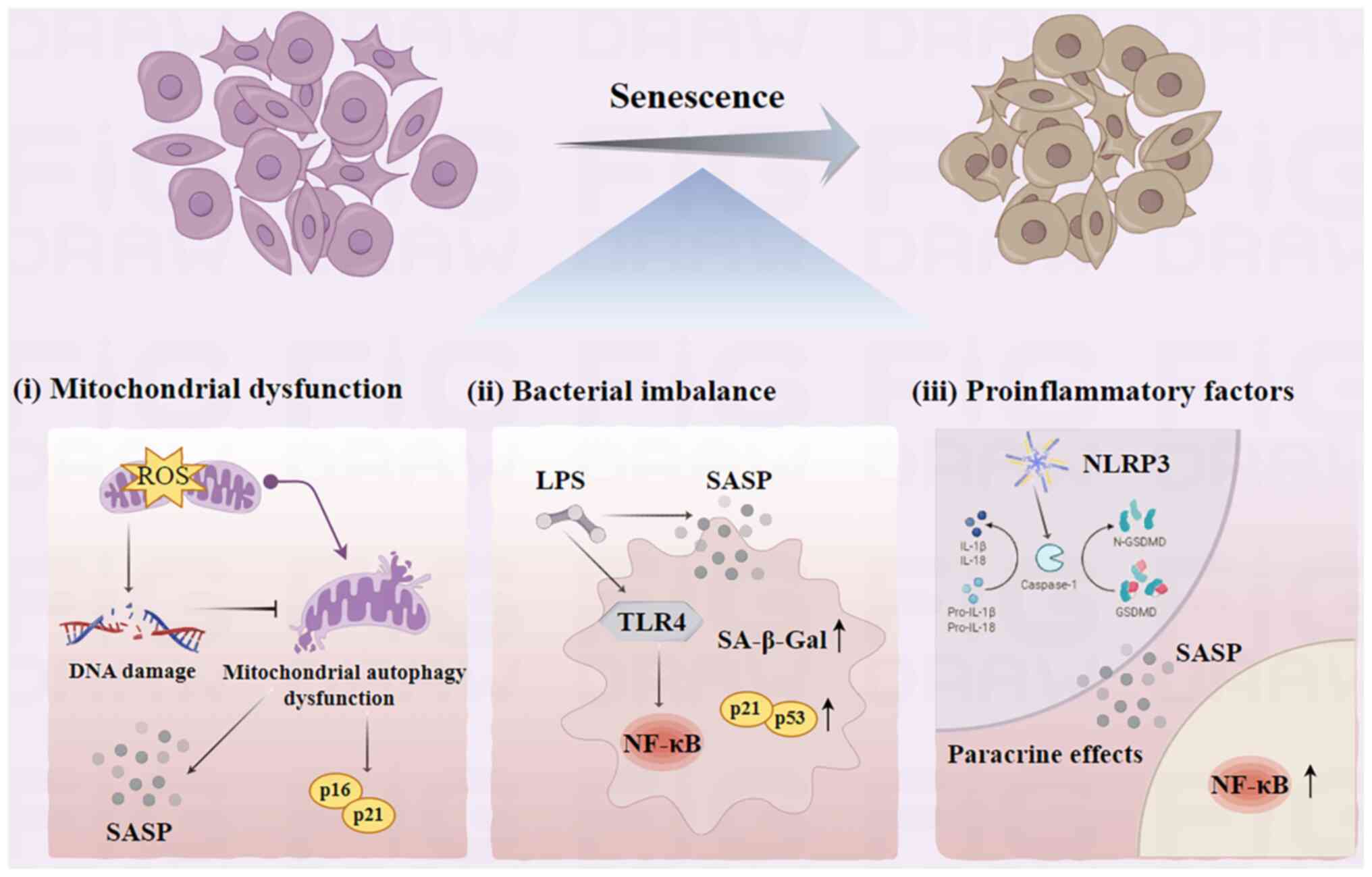
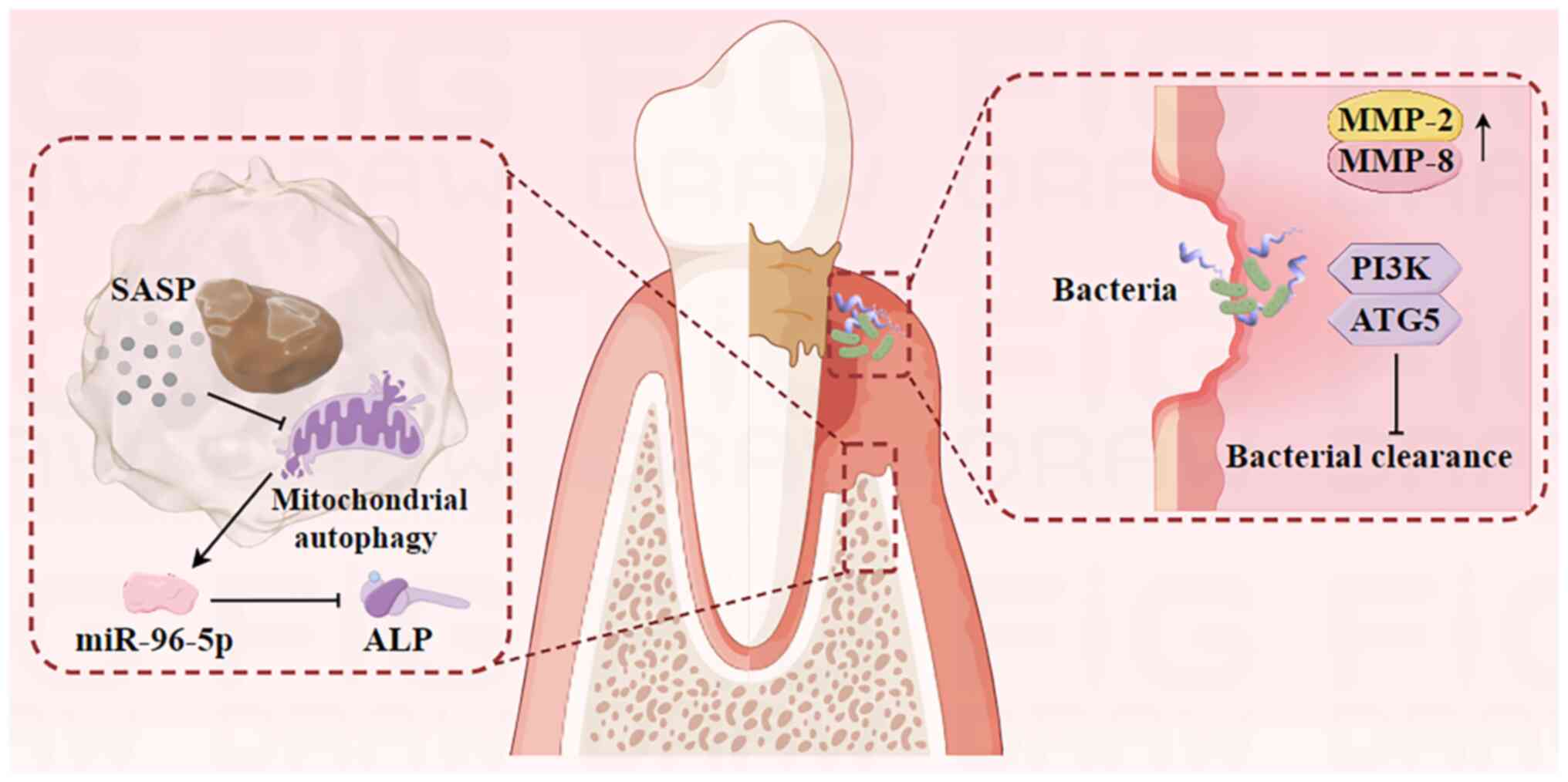
Aging encompasses a gradual decline in physiological
and pathological functions. Physiological aging involves the
natural deterioration of organs and systems over time, while
pathological aging is influenced by diseases, environmental factors
or other abnormal causes, including chronic disease, inflammation
and genetic mutation (1). There is
not an absolute boundary between physiological and pathological
aging and both can influence the onset of cellular senescence
(2,3). Cellular senescence is the
irreversible halting of cell division, which is induced by external
pro-senescence factors. IGFBP5 actively contributes to promoting
senescence and can induce senescence in neighboring cells. The
senescence-associated secretory phenotype (SASP) contributes to
this process through the secretion of interleukins (ILs), tumor
necrosis factor (TNF)-α, prostaglandin E2, interferon (INF)-β and
IFN-γ. Periodontitis, a chronic inflammatory disease,
disproportionately affects the patients aged 60–65 years, with
aging being a notable risk factor that exacerbates alveolar bone
and tooth loss (4). In developing
countries, periodontal disease is common in the elderly, with
62–97.00% having mild periodontitis and 20.00 to 48.00% having
severe periodontitis (5). Various
stimuli such as oxidative stress, proinflammatory factors,
microbial infections or activation of signaling pathways can induce
senescence in cells that is independent of telomere shortening. Due
to cell cycle arrest, senescent cells exhibit altered expression
profiles of proteins and transcription factors that are associated
with the regulation of the cell cycle (6). While bacteria may induce
inflammation, the immune response of the host is the primary driver
of periodontal tissue destruction. Cellular senescence can
compromise bacterial clearance and immune defenses (7). Targeting cellular senescence presents
a novel approach to understanding the mechanisms and clinical
management of periodontitis. The present review investigated the
mechanisms of cellular senescence and the characterization of
periodontal tissues during cellular senescence (Fig. 1).
Aging is characterized by progressive degeneration,
which is influenced by alterations in the oxidation-reduction
status and inflammatory responses triggered by oxidative stress
(8). This results in an increased
production of reactive oxygen species (ROS) within the body,
leading to abnormal levels of oxidative stress. Specific increases
in the levels of ROS are hypothesized to serve an important role in
initiating and perpetuating the cellular aging processes. The
accumulation of ROS can result in DNA damage and subsequent cell
cycle arrest (9). DNA damage can
further lead to telomere shortening, DNA methylation, histone
deacetylation and mitochondrial dysfunction, thereby instigating
transcriptome alterations that are associated with aging (10,11).
Aging is associated with alterations in the
composition of microbial communities. For example, in the gut, the
microbiota can influence the aging of gut cells by regulating the
intestinal epithelial barrier, gut immune system and gut
metabolism. In the skin, the microbiota can influence skin cell
senescence by regulating the integrity of the stratum corneum, the
skin immune system and skin metabolism. Previous studies reveal a
shift in the microbiome with age, demonstrating a positive
association between aging and the presence of gram-negative
bacteria such as Porphyromonas gingivalis (21,22).
Furthermore, aging is characterized by an increased responsiveness
of periodontal cells to the oral microbiota and mechanical stress
(23). A previous study indicates
that bacterial lipopolysaccharides (LPS) can induce cellular
senescence and increase expression levels of SASP-associated genes
(24). Prolonged exposure to P.
gingivalis LPS can prompt periodontal cell senescence due to
the activation of the cell cycle arrester molecule p53 (25). LPS also mediate microglia
activation and ischemic brain injury via the NF-κB signaling
pathway (26). Additionally, LPS
from Escherichia coli is indicated to increase the activity
of senescence-associated β-galactosidase, increase the expression
levels of cell cycle inhibitor proteins p21 and p53, and impede
human pulp stem cell growth (25,27,28).
Increased expression of LPS by P. gingivalis may stimulate
NF-κB activation in periodontal ligament stem cells (PDLSCs),
exacerbate the high glucose microenvironment and increase the
expression of proinflammatory cytokines (namely, ILs, TNF and INF)
and matrix metalloproteinases (MMPs) such as MMP-1 (29,30).
LPS can also trigger the downstream NF-κB pathway via Toll-like
receptor 4, prompting the release of inflammatory cytokines such as
TNF-α, IL-6 and IL-1β that contribute to cellular damage.
Activation of NF-κB also results in an increased expression of p53
and p21 in the PDLSC nucleus, which is pivotal in promoting
cellular aging (31,32).
A potential feedback loop exists between
inflammatory cytokines and cellular senescence, which could hasten
the progression of the senescence of inflammatory cell (33). Senescent cells are more vulnerable
to harm from external stimuli, including proinflammatory factors
and bacterial virulence factors. The NOD-like receptor pyrin
domain-containing protein 3 inflammasome recognizes
pathogen-associated molecular patterns or damage-associated
molecular patterns, activating cysteinyl aspartate specific
proteinases-1 (Caspase-1) (34).
Caspase-1 cleaves the protein Gasdermin D, which then converts the
precursor forms of IL-1β and IL-18 into mature forms, thereby
initiating pyroptosis (35,36).
IL-1β can promote paracrine cellular senescence and NF-κB
activation, activating a cascade of an inflammatory-induced
senescence (37). The accumulation
of pyroptotic macrophages (MΦs) in gingival tissue under
high-glucose conditions can stimulate IL-1β secretion and paracrine
senescence in neighboring cells (38). Furthermore, proinflammatory
cytokines (such as ILs, TNF and INF) generate the ROS themselves
and then the ROS induce epithelial cell senescence, the ROS
activate the Eotaxin-1/CCL11 pathway leading to fibroblast
senescence (39,40).
Senescent cells, with a SASP, release various
proinflammatory factors (such as IL-1α, IL-1β, IL-6 and IL-8),
growth factors (such as hepatocyte growth factor, TGF-β and
granulocyte macrophage colony-stimulating factor), chemokines [such
as chemokine (C-X-C motif) ligand (CXCL)-1/3 and CXCL-10] and MMP-8
and MMP-9. The SASP can promote chronic inflammation through
paracrine effects, affecting neighboring stem cells, fibroblasts,
immune cells, epithelial cells and endothelial cells via paracrine
and autocrine signaling mechanisms (41). In the aging heart, MMP-9 activates
the transition of the MΦ phenotype to the proinflammatory M1
subtype can induce inflammation due to its increased secretion of
TNF-α, IL-1β, IL-6 and other inflammatory factors (42). Activation of the SASP initiates a
milieu of chronic inflammation in addition to the age-induced
stressors, leading to a self-perpetuating cycle of senescent cell
accumulation (43).
PDLSCs possess self-renewal and multi-directional
differentiation capabilities, which are important for periodontal
tissue regeneration and osteogenic differentiation (44). The senescence of PDLSCs reduces
their osteogenic potential, leading to periodontal tissue
destruction via inflammation and the induction of the SASP
(Fig. 2). This process reduces the
regenerative capacity of periodontal tissues in periodontitis. In
patients with diabetes, oxidative stress induces telomere
dysfunction and PDLSC senescence, reducing periodontal bone tissue
regeneration and increasing bone loss in periodontitis (45–47).
Aging impacts the proliferation and differentiation
of bone marrow mesenchymal stem cells (BMSCs), with senescent cells
involved in senile osteoporosis-related bone loss (48). Sirtuin (SIRT), a NAD-dependent
deacetylase family member, is a potential therapeutic target for
age-related diseases (49). SIRT1,
an upstream regulator of mitochondrial autophagy, can inhibit
age-related degenerative changes in IVDD and OA (18,50).
The SIRT1/PTEN-induced kinase 1/Parkin pathway-mediated mitophagy
activation reduces renal tubular epithelial cell senescence
(51). The balance between
osteoblast and osteoclast activity maintains bone mass, and
osteoblasts are important in bone formation through matrix
synthesis, substance secretion and tissue mineralization.
Upregulation of SIRT1 expression levels activates the PI3K/Akt/mTOR
pathway to promote mitophagy, enhancing osteoblast proliferation
and viability (52).
Inhibition of micro (mi)RNA-96-5p expression levels
in osteoblasts markedly downregulates the mRNA of alkaline
phosphatase (the bone differentiation factor), which indicates that
miRNA-96-5p promotes osteoblast senescence, leading to decreased
bone differentiation. miRNA-96-5p is likely crucial in regulating
osteoblast degenerative changes during aging (53,54).
The accumulation of senescent cells in the alveolar bone can
stimulate the secretion of SASP-associated protein, which exerts a
potent paracrine effect on osteoblasts, inhibiting their function
and reducing bone regeneration in periodontitis (23,55).
Inflammatory bone loss in periodontitis, characterized by
osteoclast activity, is exacerbated by aging, which enhances
osteoclast production (56).
Osteoblast-secreted osteoprotegerin (OPG) mitigates bone loss by
inhibiting the receptor activator of NF-κB (RANK) ligand/RANK
pathway. However, the reduction in osteoblast numbers with aging
reduces OPG levels, contributing to osteoclast-induced bone loss
(57,58). These findings underscore the causal
role of senescent cells and their SASPs in alveolar bone loss and
suggest that targeting senescent cells could increase the alveolar
bone remodeling capabilities.
Gingival fibroblasts (GFs) are predominant cells
within the gingival connective tissue and serve an important role
in regulating periodontal inflammation. The proliferative capacity
of GFs and the mitotic activity of PDLSCs may reduce with age. This
reduction is associated with increased mRNA levels of MMP-2 and
MMP-8, leading to the increased degradation of the extracellular
matrix (59). Chronic bacterial
assaults can induce a reduction in the amount of epithelial growth,
compromising the integrity of the epithelial barrier (60). Once this barrier is breached,
bacteria and their virulence factors such as endotoxin and
exotoxin, can penetrate deeper connective tissues, exacerbating
periodontal tissue damage (61).
Neutrophils are important for pathogen clearance in
the immune response to periodontitis. Age-related neutrophil
dysfunction may disrupt the neutrophil homeostasis in periodontal
tissues, impairing bacterial clearance and resulting in subsequent
damage to periodontal tissues (62). Previous studies demonstrate
developmental endothelial locus-1 mRNA and protein expression
levels are downregulated in the periodontal tissue of aged mice
compared with that of young mice (63,64).
This downregulation leads to an excessive recruitment of
neutrophils and inflammatory bone resorption. Furthermore, in
elderly hosts aged >65 years, excessive or dysregulated PI3K
activity impairs the accuracy of neutrophil migration (65,66).
Aging mice have a deficiency in autophagy-related protein 5,
resulting in decreased autophagy levels and impaired release of
neutrophil extracellular traps (NETs) (67–69).
This impairment hinders the clearance of pathogenic bacteria and
their metabolites in the gingival sulcus by NETs. In summary,
cellular senescence reduces the clearance of existing periodontal
bacteria, compromising the integrity of the epithelial barrier and
exacerbating local periodontal inflammation.
MΦs serve an important role in the innate immunity
as the initial defense against pathogenic microorganisms.
Monitoring periodontitis activity may rely on the phenotypic
polarization of MΦs in order for them to modulate the immune
response to the subgingival biofilm and mitigate alveolar bone loss
(70). Aging-related M1 to M2
repolarization failure can lead to an increase in osteoclast
activation, a reduction in osteoblast formation, an increase in
bone resorption and a decrease in bone formation. Senescent MΦs are
implicated in perpetuating chronic inflammation during bone
remodeling, as well as inducing a senescent state in young BMSCs
and diminishing their osteogenic potential (71). MΦ M1 polarization is associated
with PDLSC senescence in diabetic periodontitis microenvironments.
By contrast, M2 polarization potentially increases the expression
levels of osteogenesis-related cytokines, such as runt-related
transcription factor 2, alkaline phosphatase and osteocalcin,
exerts anti-aging effects and facilitates the osteogenic
differentiation of PDLSCs (72,73).
Additionally, the paracrine factors released by PDLSCs can
stimulate the expression of CD163, a surface marker associated with
M2-MΦs, which contributes to macrophage M2 polarization (74).
Dendritic cells (DCs) represent a diverse cell
population and inhibiting their function results in reduced
responses of the adaptive immune system as well as an increased
susceptibility to periodontal disease. A previous study indicates
that the quantity of DCs in the peripheral blood fluctuates with
age, potentially contributing to the increased vulnerability of
older patients to infection compared with younger patients
(75). Furthermore, alterations in
the functionality of DCs contribute to immune dysregulation and the
onset of chronic inflammation. In vitro, infection of DCs by
the non-living oral pathogen P. gingivalis triggers
activation of the SASP and subsequent bone loss (76).
Langerhans cells (LCs), a subset of DCs located in
the oral mucosal epithelium, likely serve a role in initiating and
perpetuating periodontal diseases (77). A decrease in the number of
epithelial LCs and their dendritic structures in elderly
individuals aged >75 years with periodontitis, which are
distinct compared with those in adults, may compromise their
removal of pathogenic bacteria (78,79).
Reducing LCs in the aging epidermis impairs the immunoregulatory
functions of the skin and compromises barrier integrity,
antimicrobial defenses and overall cell protection capabilities
(80,81). Consequently, immune cell senescence
may disrupt the homeostasis of periodontal defense mechanisms.
In conclusion, the prevalence and severity of
periodontitis are associated with cellular senescence. The
intricate changes in cellular senescence can impair the effective
removal of pathogens by immune cells. The interaction of
intracellular and extracellular aging environments can lead to the
deterioration of cells and factors involved in bone metabolism,
tissue defense and immune response in periodontal tissues, which
exacerbates the progression of periodontitis. Furthermore, a
comprehensive literature review underscores the complexity of the
association between cellular senescence and inflammation. The
precise nature of the association between cellular senescence and
inflammation is yet to be completely understood, with the
possibility of a causal or bidirectional association.
The current understanding suggests that the cascade
of changes triggered by cellular senescence serves a role in the
development of periodontal disease, potentially explaining the
increased prevalence of this condition among the elderly aged
>65 years. Increasing the understanding of the association
between cellular senescence and periodontitis could offer a novel
approach to preventing and treating periodontal diseases in older
individuals. Further investigation is required to assess the
applicability of these mechanisms to other age-related diseases
such as hypertension, diabetes in clinical settings.
Not applicable.
The present study was funded by the National Natural Science
Foundation of China (grant no. 82201080), the High-level Talents
Project of Hainan Natural Science Foundation (grant no. 821RC687)
and the Innovative Scientific Research Project for Postgraduates of
Hainan Medical College (grant no. Qhys2022-280).
Not applicable.
ZG conceived and designed the present review. XL
wrote most of the manuscript. DS, JZ, YD and QY edited the
manuscript. All authors read and approved the final version of the
manuscript. Data authentication is not applicable.
Not applicable.
Not applicable.
The authors declare that they have no competing
interests.
|
1
|
Lu Y, Tang X, Wang W, Yang J and Wang S:
The role of deacetylase SIRT1 in allergic diseases. Front Immunol.
15:14225412024. View Article : Google Scholar
|
|
2
|
Jothi D and Kulka LAM: Strategies for
modeling aging and age-related diseases. NPJ Aging. 10:322024.
View Article : Google Scholar
|
|
3
|
López-Otín C, Blasco MA, Partridge L,
Serrano M and Kroemer G: Hallmarks of aging: An expanding universe.
Cell. 186:243–278. 2023. View Article : Google Scholar
|
|
4
|
Chen S, Zhou D, Liu O, Chen H, Wang Y and
Zhou Y: Cellular senescence and periodontitis: Mechanisms and
therapeutics. Biology (Basel). 11:14192022.
|
|
5
|
Verma SK, Singh N, Jha AK, Tigga C,
Noorani MK, Ekram S and Gupta V: Prevalence of periodontal disease
among patients reporting to tertiary care hospital in Ranchi. J
Pharm Bioallied Sci. 16 (Suppl 1):S838–S840. 2024. View Article : Google Scholar
|
|
6
|
Yan J, Chen S, Yi Z, Zhao R, Zhu J, Ding S
and Wu J: The role of p21 in cellular senescence and aging-related
diseases. Mol Cells. 18:1001132024. View Article : Google Scholar
|
|
7
|
Dong Z, Luo Y, Yuan Z, Tian Y, Jin T and
Xu F: Cellular senescence and SASP in tumor progression and
therapeutic opportunities. Mol Cancer. 23:1812024. View Article : Google Scholar
|
|
8
|
Li X, Li C, Zhang W, Wang Y, Qian P and
Huang H: Inflammation and aging: Signaling pathways and
intervention therapies. Signal Transduct Target Ther. 8:2392023.
View Article : Google Scholar
|
|
9
|
Rusciano D and Bagnoli P: Oxygen, the
Paradox of Life and the Eye. Front Biosci (Landmark Ed).
29:3192024. View Article : Google Scholar
|
|
10
|
Wang B, Wang Y, Zhang J, Hu C, Jiang J, Li
Y and Peng Z: ROS-induced lipid peroxidation modulates cell death
outcome: Mechanisms behind apoptosis, autophagy, and ferroptosis.
Arch Toxicol. 97:1439–1451. 2023. View Article : Google Scholar
|
|
11
|
Terao R, Ahmed T, Suzumura A and Terasaki
H: Oxidative stress-induced cellular senescence in aging retina and
age-related macular degeneration. Antioxidants (Basel).
11:21892022. View Article : Google Scholar
|
|
12
|
Wen J, Pan T, Li H, Fan H, Liu J, Cai Z
and Zhao B: Role of mitophagy in the hallmarks of aging. J Biomed
Res. 37:1–14. 2022. View Article : Google Scholar
|
|
13
|
Sun Y, Xu L, Li Y, Jia S, Wang G, Cen X,
Xu Y, Cao Z, Wang J, Shen N, et al: Mitophagy defect mediates the
aging-associated hallmarks in Hutchinson-Gilford progeria syndrome.
Aging Cell. 23:e141432024. View Article : Google Scholar
|
|
14
|
Ilie OD, Ciobica A, Riga S, Dhunna N,
McKenna J, Mavroudis I, Doroftei B, Ciobanu AM and Riga D:
Mini-Review on lipofuscin and aging: focusing on the molecular
interface, the biological recycling mechanism, oxidative stress,
and the gut-brain axis functionality. Medicina (Kaunas).
56:6262020. View Article : Google Scholar
|
|
15
|
Lee KS, Lin S, Copland DA, Dick AD and Liu
J: Cellular senescence in the aging retina and developments of
chemotherapies for age-related macular degeneration. J
Neuroinflammation. 18:322021. View Article : Google Scholar
|
|
16
|
Blasiak J: Senescence in the pathogenesis
of age-related macular degeneration. Cell Mol Life Sci. 77:789–805.
2020. View Article : Google Scholar
|
|
17
|
Kuang Y, Hu B, Feng G, Xiang M, Deng Y,
Tan M, Li J and Song J: Metformin prevents oxidative stress-induced
senescence in human periodontal ligament cells. Biogerontology.
21:13–27. 2020. View Article : Google Scholar
|
|
18
|
Sun K, Jing X, Guo J, Yao X and Guo F:
Mitophagy in degenerative joint diseases. Autophagy. 17:2082–2092.
2021. View Article : Google Scholar
|
|
19
|
Yu H and Song X: The relationship between
Alzheimer's disease and thyroiditis: A two-sample Mendelian
randomization study. Medicine (Baltimore). 102:e357122023.
View Article : Google Scholar
|
|
20
|
Xu Y, Sun B, Wang H, Cai Y, Chu D, Cao R
and Wang Z: Autophagy regulates age-related delayed jawbone
regeneration and decreased osteoblast osteogenesis by degrading
FABP3. FASEB J. 38:e238242024. View Article : Google Scholar
|
|
21
|
Ebersole JL, Nagarajan R, Kirakodu S and
Gonzalez OA: Oral microbiome and gingival gene expression of
inflammatory biomolecules with aging and periodontitis. Front Oral
Health. 2:7251152021. View Article : Google Scholar
|
|
22
|
Schwartz JL, Peña N, Kawar N, Zhang A,
Callahan N, Robles SJ, Griebel A and Adami GR: Old age and other
factors associated with salivary microbiome variation. BMC Oral
Health. 21:4902021. View Article : Google Scholar
|
|
23
|
Song Y and Chung J: Aging aggravates
periodontal inflammatory responses and alveolar bone resorption by
porphyromonas gingivalis infection. Curr Issues Mol Biol.
45:6593–6604. 2023. View Article : Google Scholar
|
|
24
|
Suzuki K, Susaki EA and Nagaoka I:
Lipopolysaccharides and cellular senescence: Involvement in
Atherosclerosis. Int J Mol Sci. 23:111482022. View Article : Google Scholar
|
|
25
|
Aquino-Martinez R, Rowsey JL, Fraser DG,
Eckhardt BA, Khosla S, Farr JN and Monroe DG: LPS-induced premature
osteocyte senescence: Implications in inflammatory alveolar bone
loss and periodontal disease pathogenesis. Bone. 132:1152202020.
View Article : Google Scholar
|
|
26
|
Jung YS, Park JH, Kim H, Kim SY, Hwang JY,
Hong KW, Bae SS, Choi BT, Lee SW and Shin HK: Probucol inhibits
LPS-induced microglia activation and ameliorates brain ischemic
injury in normal and hyperlipidemic mice. Acta Pharmacol Sin.
37:1031–1044. 2016. View Article : Google Scholar
|
|
27
|
Feng G, Zheng K, Cao T, Zhang J, Lian M,
Huang D, Wei C, Gu Z and Feng X: Repeated stimulation by LPS
promotes the senescence of DPSCs via TLR4/MyD88-NF-κB-p53/p21
signaling. Cytotechnology. 70:1023–1035. 2018. View Article : Google Scholar
|
|
28
|
Gao H, Nepovimova E, Heger Z, Valko M, Wu
Q, Kuca K and Adam V: Role of hypoxia in cellular senescence.
Pharmacol Res. 194:1068412023. View Article : Google Scholar
|
|
29
|
Nareika A, Im YB, Game BA, Slate EH,
Sanders JJ, London SD, Lopes-Virella MF and Huang Y: High glucose
enhances lipopolysaccharide-stimulated CD14 expression in U937
mononuclear cells by increasing nuclear factor kappaB and AP-1
activities. J Endocrinol. 196:45–55. 2008. View Article : Google Scholar
|
|
30
|
Gölz L, Memmert S, Rath-Deschner B, Jäger
A, Appel T, Baumgarten G, Götz W and rede S: Hypoxia and P.
gingivalis synergistically induce HIF-1 and NF-κB activation in PDL
cells and periodontal diseases. Mediators Inflamm. 2015:4380852015.
View Article : Google Scholar
|
|
31
|
Zusso M, Lunardi V, Franceschini D,
Pagetta A, Lo R, Stifani S, Frigo AC, Giusti P and Moro S:
Ciprofloxacin and levofloxacin attenuate microglia inflammatory
response via TLR4/NF-kB pathway. J Neuroinflammation. 16:1482019.
View Article : Google Scholar
|
|
32
|
Yin K, Patten D, Gough S, de Barros
Gonçalves S, Chan A, Olan I, Cassidy L, Poblocka M, Zhu H, Lun A,
et al: Senescence-induced endothelial phenotypes underpin
immune-mediated senescence surveillance. Genes Dev. 36:533–549.
2022. View Article : Google Scholar
|
|
33
|
Li Y, Yin H, Yuan H, Wang E, Wang C, Li H,
Geng X, Zhang Y and Bai J: IL-10 deficiency aggravates cell
senescence and accelerates BLM-induced pulmonary fibrosis in aged
mice via PTEN/AKT/ERK pathway. BMC Pulm Med. 24:4432024. View Article : Google Scholar
|
|
34
|
Tang H, Du Y, Tan Z, Li D and Xie J:
METTL14-mediated HOXA5 m6A modification alleviates osteoporosis via
promoting WNK1 transcription to suppress NLRP3-dependent macrophage
pyroptosis. J Orthop Translat. 48:190–203. 2024. View Article : Google Scholar
|
|
35
|
Liu Z, Gan L, Xu Y, Luo D, Ren Q, Wu S and
Sun C: Melatonin alleviates inflammasome-induced pyroptosis through
inhibiting NF-κB/GSDMD signal in mice adipose tissue. J Pineal Res.
63:2017. View Article : Google Scholar
|
|
36
|
Zhang Z, Li X, Wang Y, Wei Y and Wei X:
Involvement of inflammasomes in tumor microenvironment and tumor
therapies. J Hematol Oncol. 16:242023. View Article : Google Scholar
|
|
37
|
Martínez-Zamudio RI, Robinson L, Roux PF
and Bischof O: SnapShot: Cellular senescence pathways. Cell.
170:816–816.e1. 2017. View Article : Google Scholar
|
|
38
|
Zhao P, Yue Z, Nie L, Zhao Z and Wang Q,
Chen J and Wang Q: Hyperglycemia-associated macrophage pyroptosis
accelerates periodontal inflame-aging. J Clin Periodontol.
48:1379–1392. 2021. View Article : Google Scholar
|
|
39
|
Tsirpanlis G: Cellular senescence and
inflammation: A noteworthy link. Blood Purif. 28:12–14. 2009.
View Article : Google Scholar
|
|
40
|
Lavandoski P, Pierdoná V, Maurmann RM,
Grun LK, Guma F, Guma FTCR and Barbé-Tuana FM: Eotaxin-1/CCL11
promotes cellular senescence in human-derived fibroblasts through
pro-oxidant and proinflammatory pathways. Front Immunol.
14:12435372023. View Article : Google Scholar
|
|
41
|
Yue Z, Nie L, Zhao P, Ji N, Liao G and
Wang Q: Senescence-associated secretory phenotype and its impact on
oral immune homeostasis. Front Immunol. 13:10193132022. View Article : Google Scholar
|
|
42
|
Ma Y, Mouton AJ and Lindsey ML: Cardiac
macrophage biology in the steady-state heart, the aging heart, and
following myocardial infarction. Transl Res. 191:15–28. 2008.
View Article : Google Scholar
|
|
43
|
Gasek NS, Kuchel GA, Kirkland JL and Xu M:
Strategies for targeting senescent cells in human disease. Nat
Aging. 1:870–879. 2021. View Article : Google Scholar
|
|
44
|
Zhang J, An Y, Gao LN, Zhang YJ, Jin Y and
Chen FM: The effect of aging on the pluripotential capacity and
regenerative potential of human periodontal ligament stem cells.
Biomaterials. 33:6974–6986. 2012. View Article : Google Scholar
|
|
45
|
Li X, Zhang B, Wang H, Zhao X, Zhang Z,
Ding G and Wei F: The effect of aging on the biological and
immunological characteristics of periodontal ligament stem cells.
Stem Cell Res Ther. 11:3262020. View Article : Google Scholar
|
|
46
|
Tang L, Li T, Chang Y, Wang Z, Li Y, Wang
F and Sui L: Diabetic oxidative stress-induced telomere damage
aggravates periodontal bone loss in periodontitis. Biochem Biophys
Res Commun. 614:22–28. 2022. View Article : Google Scholar
|
|
47
|
Du TT, Liu N, Zhang W, Shi HG and Zhang T:
Effect of aging on proliferative and differentiation capacity of
human periodontal ligament stem cells. Nan Fang Yi Ke Da Xue Xue
Bao. 37:360–366. 2017.(In Chinese).
|
|
48
|
Föger-Samwald U, Kerschan-Schindl K,
Butylina M and Pietschmann P: Age Related osteoporosis: Targeting
cellular senescence. Int J Mol Sci. 23:27012022. View Article : Google Scholar
|
|
49
|
Cui Z and Zhao X, Amevor FK, Du X, Wang Y,
Li D, Shu G, Tian Y and Zhao X: Therapeutic application of
quercetin in aging-related diseases: SIRT1 as a potential
mechanism. Front Immunol. 13:9433212022. View Article : Google Scholar
|
|
50
|
Cao H, Zhou X, Xu B, Hu H, Guo J, Wang M,
Li N and Jun Z: Advances in the study of mitophagy in
osteoarthritis. J Zhejiang Univ Sci B. 25:197–211. 2024.(In
English, Chinese). View Article : Google Scholar
|
|
51
|
Liu T, Yang Q, Zhang X, Qin R, Shan W,
Zhang H and Chen X: Quercetin alleviates kidney fibrosis by
reducing renal tubular epithelial cell senescence through the
SIRT1/PINK1/mitophagy axis. Life Sci. 257:1181162020. View Article : Google Scholar
|
|
52
|
Yang X, Jiang T, Wang Y and Guo L: The
Role and Mechanism of SIRT1 in resveratrol-regulated osteoblast
autophagy in osteoporosis rats. Sci Rep. 9:184242019. View Article : Google Scholar
|
|
53
|
Wu BW, Guo JD, Wu MS, Liu Y, Lu M, Zhou YH
and Han HW: Osteoblast-derived lipocalin-2 regulated by
miRNA-96-5p/Foxo1 advances the progression of Alzheimer's disease.
Epigenomics. 12:1501–1513. 2020. View Article : Google Scholar
|
|
54
|
Yu H, Ji X and Ouyang Y: Unfolded protein
response pathways in stroke patients: A comprehensive landscape
assessed through machine learning algorithms and experimental
verification. J Transl Med. 21:7592023. View Article : Google Scholar
|
|
55
|
Aquino-Martinez R: The emerging role of
accelerated cellular senescence in periodontitis. J Dent Res.
102:854–862. 2023. View Article : Google Scholar
|
|
56
|
Wang Z, Zhang X, Cheng X, Ren T, Xu W, Li
J, Wang H and Zhang J: Inflammation produced by senescent
osteocytes mediates age-related bone loss. Front Immunol.
14:11140062023. View Article : Google Scholar
|
|
57
|
Hu S and Wang S: The Role of SIRT3 in the
Osteoporosis. Front Endocrinol (Lausanne). 13:8936782022.
View Article : Google Scholar
|
|
58
|
Feng Y and Tang X: FoxO1 as the critical
target of puerarin to inhibit osteoclastogenesis and bone
resorption. J Pharm Pharmacol. 76:813–823. 2024. View Article : Google Scholar
|
|
59
|
Ben-Eltriki M, Ahmadi AR, Nakao Y, Golla
K, Lakschevitz F, Häkkinen L, Granville DJ and Kim H: Granzyme B
promotes matrix metalloproteinase-1 (MMP-1) release from gingival
fibroblasts in a PAR1- and Erk1/2-dependent manner: A novel role in
periodontal inflammation. J Periodontal Res. 59:94–103. 2024.
View Article : Google Scholar
|
|
60
|
Ko YK, Hong S, Kim HM, Liu M, Moon E, Kim
P and Choi Y: Characterization of junctional structures in the
gingival epithelium as barriers against bacterial invasion. J
Periodontal Res. 57:799–810. 2022. View Article : Google Scholar
|
|
61
|
Jotwani R, Palucka AK, Al-Quotub M,
Nouri-Shirazi M, Kim J, Bell D, Banchereau J and Cutler CW: Mature
dendritic cells infiltrate the T cell-rich region of oral mucosa in
chronic periodontitis: In situ, in vivo, and in vitro studies. J
Immunol. 167:4693–4700. 2021. View Article : Google Scholar
|
|
62
|
Hajishengallis G, Moutsopoulos NM,
Hajishengallis E and Chavakis T: Immune and regulatory functions of
neutrophils in inflammatory bone loss. Semin Immunol. 28:146–158.
2016. View Article : Google Scholar
|
|
63
|
Kim TS and Moutsopoulos NM: Neutrophils
and neutrophil extracellular traps in oral health and disease. Exp
Mol Med. 56:1055–1065. 2024. View Article : Google Scholar
|
|
64
|
Eskan MA, Jotwani R, Abe T, Chmelar J, Lim
JH, Liang S, Ciero PA, Krauss JL, Li F, Rauner M, et al: The
leukocyte integrin antagonist Del-1 inhibits IL-17-mediated
inflammatory bone loss. Nat Immunol. 13:465–473. 2012. View Article : Google Scholar
|
|
65
|
Sapey E, Greenwood H, Walton G, Mann E,
Love A, Aaronson N, Insall RH, Stockley RA and Lord JM:
Phosphoinositide 3-kinase inhibition restores neutrophil accuracy
in the elderly: Toward targeted treatments for immunosenescence.
Blood. 123:239–248. 2014. View Article : Google Scholar
|
|
66
|
Bülow S, Ederer KU, Holzinger JM, Zeller
L, Werner M, Toelge M, Pfab C, Hirsch S, Göpferich F, Hiergeist A,
et al: Bactericidal/permeability-increasing protein instructs
dendritic cells to elicit Th22 cell response. Cell Rep.
43:1139292024. View Article : Google Scholar
|
|
67
|
Xu F, Zhang C, Zou Z, Fan EKY, Chen L, Li
Y, Billiar TR, Wilson MA, Shi X and Fan J: Aging-related Atg5
defect impairs neutrophil extracellular traps formation.
Immunology. 151:417–432. 2017. View Article : Google Scholar
|
|
68
|
Kim TS, Silva LM, Theofilou VI,
Greenwell-Wild T, Li L, Williams DW, Ikeuchi T, Brenchley L;
NIDCD/NIDCR Genomics and Computational Biology Core, ; Bugge TH, et
al: Neutrophil extracellular traps and extracellular histones
potentiate IL-17 inflammation in periodontitis. J Exp Med.
220:e202217512023. View Article : Google Scholar
|
|
69
|
Yu H, Si G and Si F: Mendelian
Randomization validates the immune landscape mediated by aggrephagy
in esophageal squamous cell carcinoma patients from the
perspectives of Multi-omics. J Cancer. 15:1940–1953. 2024.
View Article : Google Scholar
|
|
70
|
Sloniak MC, Lepique AP, Nakao LYS and
Villar CC: Alterations in macrophage polarization play a key role
in control and development of periodontal diseases. J Indian Soc
Periodontol. 27:578–582. 2023. View Article : Google Scholar
|
|
71
|
Bai L, Liu Y, Zhang X, Chen P, Hang R,
Xiao Y, Wang J and Liu C: Osteoporosis remission via an
anti-inflammaging effect by icariin activated autophagy.
Biomaterials. 297:1221252023. View Article : Google Scholar
|
|
72
|
Gong J, Ye C, Ran J, Xiong X, Fang X, Zhou
X, Yi Y, Lu X, Wang J, Xie C and Liu J: Polydopamine-Mediated
immunomodulatory patch for diabetic periodontal tissue regeneration
assisted by Metformin-ZIF System. ACS Nano. 17:16573–16586. 2023.
View Article : Google Scholar
|
|
73
|
Liu J, Chen B, Bao J, Zhang Y, Lei L and
Yan F: Macrophage polarization in periodontal ligament stem cells
enhanced periodontal regeneration. Stem Cell Res Ther. 10:3202019.
View Article : Google Scholar
|
|
74
|
Corradetti B, Taraballi F, Corbo C,
Cabrera F, Pandolfi L, Minardi S, Wang X, Van Eps J, Bauza G,
Weiner B and Tasciotti E: Immune tuning scaffold for the local
induction of a pro-regenerative environment. Sci Rep. 7:170302017.
View Article : Google Scholar
|
|
75
|
Reitsema RD, Kumawat AK, Hesselink BC, van
Baarle D and van Sleen Y: Effects of ageing and frailty on
circulating monocyte and dendritic cell subsets. NPJ Aging.
10:172024. View Article : Google Scholar
|
|
76
|
Elsayed R, Elashiry M, Liu Y, Morandini
AC, El-Awady A, Elashiry MM, Hamrick M and Cutler CW:
Microbially-Induced exosomes from dendritic cells promote paracrine
immune senescence: Novel mechanism of bone degenerative disease in
mice. Aging Dis. 14:136–151. 2023. View Article : Google Scholar
|
|
77
|
Sharawi H, Heyman O, Mizraji G, Horev Y,
Laviv A, Shapira L, Yona S, Hovav AH and Wilensky A: The prevalence
of gingival dendritic cell subsets in periodontal patients. J Dent
Res. 100:1330–1336. 2021. View Article : Google Scholar
|
|
78
|
de Vasconcelos Gurgel BC, Peixe PG,
Queiroz SIML, de Almeida Freitas R, de Aquino Martins ARL and
Duarte PM: Comparison of immunoexpression of dendritic cells, mast
cells and blood vessels in periodontal disease between adults and
elderly. Clin Oral Investig. 27:6823–6833. 2023. View Article : Google Scholar
|
|
79
|
Bodineau A, Coulomb B, Folliguet M,
Igondjo-Tchen S, Godeau G, Brousse N and Séguier S: Do Langerhans
cells behave similarly in elderly and younger patients with chronic
periodontitis? Arch Oral Biol. 52:189–194. 2007. View Article : Google Scholar
|
|
80
|
Lee HJ, Kim TG, Kim SH, Park JY, Lee M,
Lee JW, Lee SH and Lee MG: Epidermal barrier function is impaired
in langerhans cell-depleted mice. J Invest Dermatol. 139:1182–1185.
2019. View Article : Google Scholar
|
|
81
|
Oulee A, Ma F, Teles RMB, de Andrade Silva
BJ, Pellegrini M, Klechevsky E, Harman AN, Rhodes JW and Modlin RL:
Identification of genes encoding antimicrobial proteins in
langerhans cells. Front Immunol. 12:6953732021. View Article : Google Scholar
|