Introduction
In spite of continuing advances in the treatment of
acute myeloid leukemia (AML), the 5-year event-free survival rate
remains low, and the majority of patients succumb to the disease
due to relapse and chemotherapy resistance. Therefore, the
development of novel therapeutic strategies is urgently required.
Experiments and clinical trials have demonstrated vascular
endothelial growth factor (VEGF) and VEGF receptors (VEGFRs) are
overexpressed in patients with acute or chronic leukemia, and are
closely associated with the occurrence, development and prognosis
of the disease (1–4). Therefore, the direct or indirect
suppression of VEGF and its receptors are therefore promising novel
therapeutic approaches to improve patient outcome.
VEGFR-1 is widely expressed in hematological
malignancies, and the functional role of VEGFR-1 is complex and
dependent on both the developmental stage and cell type (4). Although it has been reported that
VEGFR-1 regulates the distribution of leukemia cells in the bone
marrow of patients with acute lymphoblastic leukemia, and, in
conjunction with VEGF and placental derived growth factor modulate
the survival and exportation of leukemia cells (5), studies specifically defining the role
of VEGFR-1 in AML are limited.
RNA interference (RNAi) is a novel strategy for the
specific suppression of target gene expression and has been applied
in the development of antiviral and antitumor drugs (6–9),
making it possible to elucidate the role of VEGFR-1. In the present
study, we used a lentivirus-mediated shRNA expression system to
specifically inhibit VEGFR-1 expression in the U937 leukemia cell
line. Our results showed that shRNA reduced the proliferation and
migration of U937 cells. In addition, RNA interference targeting
VEGFR-1 in combination with bevacizumab did not exert synergistic
inhibitory effects on the U937 cells. However, the knockdown of
VEGFR-1 by shRNA made the U937 cells more sensitive to cytarabine.
These findings suggest that lentivirus-mediated RNA interference
targeting VEGFR-1 specifically inhibits the expression of VEGFR-1,
resulting in a series of changes in the biological characteristics
of leukemia cells, and may be applied as a novel antitumor
therapeutic strategy for the treatment of leukemia.
Materials and methods
Cell culture and primary leukemia cell
lines
The leukemia cell lines, HL-60, HEL, NB4, U937 and
THP-1, and lentivirus packaging cell line (293T cells) were
purchased from the Shanghai Institute for Biological Sciences,
Chinese Academy of Sciences, Shanghai, China. The 293T cells were
grown in DMEM containing 10% fetal bovine serum (FBS) and the other
cells were maintained in RPMI-1640 medium supplemented with 10%
FBS. Cells were incubated at 37°C in humidified 5% CO2
in air and those in the logarithmic growth phase were used for
further experiments. The diagnosis of primary leukemia in 100
patients was confirmed by bone marrow biopsy and flow cytometry.
After obtaining informed consent, 5 ml of heparin anti-coagulated
bone marrow were collected and mononuclear cells were separated
with Ficoll, and were then cultured under regular conditions or
stored at −80°C for further use. In all samples, the proportion of
leukemia cells was >80% and trypan blue excluding fraction was
>98%. For the control group, peripheral venous blood was
obtained from 15 healthy volunteer donors who also provided written
informed consent before the samples were obtained.
Reagents
Lentiviral vector for RNA interference (Shanghai
Innovation Biotechnology Co., Ltd., Shanghai, China), RPMI-1640
medium, DMEM (Gibco, Carlsbad, CA, USA), FBS (HyClone, Logan, UT,
USA), TRIzol for RNA extraction (Invitrogen, Carlsbad, CA, USA),
the reverse transcription kit (Promega, Madison, WI, USA), the
SYBR-Green Real-time PCR kit (Takara, Shiga, Japan), the cell
counting kit-8 (CCK-8; Dojindo Laboratories, Kumamoto, Japan), the
Annexin V-PE/7-AAD apoptosis detection kit (Nanjing KeyGen Biotech
Co., Ltd., Nanjing, China), cell cycle staining solution
(MultiSciences Biotech Co., Ltd., Shanghai, China), VEGFR-1
monoclonal antibody (Abcam, Cambridge, MA, USA), secondary
antibodies (R&D Systems, Minneapolis, MN, USA), cytarabine
(Pharmacia, Kalamazoo, MI, USA) and bevacizumab (Genentech, South
San Francisco, CA, USA) were used in the present study.
RT-PCR of VEGF, VEGFR-1 and VEGFR-2 in
leukemia cell lines
The leukemia cells were first analyzed for VEGF and
VEGFR expression by RT-PCR. Total cellular RNA was extracted, and
cDNA was synthesized following conventional protocols. PCR was
performed using a PCR thermal cycler (GeneAmp PCR System 9700; Life
Technologies, Carslbad, CA, USA). The primers used were synthesized
as follows: VEGF forward, 5′-AGGGAAGAGGAGGAGATGAG-3′ and reverse,
5′-GCTGGGTTTGTCGGTGTT-3′ (anticipated size, 148 bp); VEGFR-1
forward, 5′-AGACTAGATAGCGTCA CCAGC-3′ and reverse,
5′-GAAACCGTCAGAATCCTCC-3′ (anticipated size, 102 bp); VEGFR-2
forward, 5′-CCAATAAT CAGAGTGGCAGTG-3′ and reverse, 5′-CATAGACAT
AAATGACCGAGGC-3′ (anticipated size, 162 bp). The cycling conditions
for PCR were 38 cycles of denaturation (94°C for 20 sec), annealing
(58 or 60°C for 20 sec) and extension (72°C for 30 sec). A
pre-heating step at 94°C for 5 min and a final extension step
consisting of 7 min at 72°C were also carried out. The primer
sequences for β-actin were: forward, 5′-CCTGTACGCCAACACAGTGC-3′ and
reverse, 5′-ATACTCCTGCTTGCTGATCC-3′. The temperature for annealing
was 56°C and a total of 30 cycles were conducted. The other
conditions were as mentioned above. The experiment was performed 3
times.
Real-time PCR of VEGF, VEGFR-1 and
VEGFR-2
Fluorescence quantitative PCR instrument
(LightCycler 1.5, Roche, Basel, Switzerland) and the SYBR-Green
Master Mix kit were used for the detection of the expression of
target genes and β-actin was used as an internal reference. The
primers of VEGF and VEGFRs were as mentioned above. The temperature
for annealing was 58 or 60°C and a total of 38 cycles were
conducted. β-actin served as an internal reference and the
2−ΔΔCT method was employed to determine the relative
expression of the target genes and the experiment was repeated 3
times.
Western blot analysis for VEGFR-1
Cells were harvested and incubated with lysis buffer
for protein extraction followed by the determination of protein
concentration. The proteins were separated by SDS-PAGE and
transferred onto polyvinylidene fluoride (PVDF) membranes. The
membranes were incubated with primary antibodies (VEGFR-1, 1:500;
β-actin/GAPDH, 1:5,000) followed by secondary antibodies conjugated
to horseradish peroxidase. Color development was performed with the
ECL kit. The experiment was performed 3 times.
Detection of VEGF content by ELISA
The leukemia cell lines (1×106) were
grown in 1 ml of serum-free RPMI-1640 medium at 37°C in humidified
5% CO2 in air for 48 h. Following centrifugation, the
supernatant was obtained and the VEGF content was determined by
ELISA. Each experiment was performed in duplicate according to the
manufacturer’s instructions. The experiment was repeated 3
times.
Construction of lentiviral vector
carrying VEGFR-1 shRNA, transfection of U937 cells and
determination of the efficiency of RNA interference
According to the principles for the design of the
RNA interference sequence, 3 sequences targeting the VEGFR-1 gene
(NM_002019) were designed: -sh1: 704–723 bp,
5′-CACACCACGCCCAGTCAAA-3′;-sh2: 3221–3240 bp,
5′-AATCTACAGCACCAAGAGC-3′; and -sh3: 2639–2658 bp,
5′-AATCTTGACCCACATTGGC-3′. The shRNAs targeting the VEGFR-1 gene
were then connected to the lentiviral vector, pRNAT-U6.2/Lenti,
yielding recombinant vectors. Following PCR and sequencing, the
recombinant vectors together with lentiviral packaging plasmids (at
a ratio of 1:1:1) was transfected into the 293T cells using
Lipofectamine 2000. The viral solution was prepared and named
sh-VEGFR-1 KD. In addition, the siRNA sequence unrelated to VEGFR-1
(5′-TTCTCCGAAC GTGTCACGT-3′) was synthesized and served as the
negative control (shRNA-NC). Lentiviral particles were conjugated
to green fluorescent protein (GFP). The serial dilution method and
flow cytometry were performed to determine the lentivirus titer.
The lentivirus in the sh-VEGFR-1 KD and shRNA-NC group was used to
transfect the U937 cells at the appropriate titer and named
sh-KD-U937 (KD group) and sh-NC-U937 (NC group), respectively. In
addition, untreated U937 cells were used as the controls (CON
group). The proportion of GFP-positive cells was observed under a
fluorescence microscope 72 h later. When the proportion was
>90%, the cells were used for the following experiments. These
cells were harvested at 120 h after being transfected, real-time
PCR and western blot analysis were performed to determine the
efficiency of RNA interference at the mRNA level and protein level.
The experiment was repeated 3 times.
Growth curve assay
Twenty-four hours after transfection, 3 groups of
cells (5×103) in the logarithmic growth phase were
seeded in 96-well plates (100 μl/well) and each experiment was
performed in sextuplicate followed by incubation for 7 days. The
medium was refreshed every 48 h. The cells were then incubated with
CCK-8 solution (5 mg/ml, 10 μl/well) for 4 h followed by blending.
The absorbance (A) was determined at 450 nm with a microplate
reader, which represents the cell proliferation rate. The
experiment was repeated 3 times.
Transwell migration assay
Migration assays were performed in Transwell plates
(Costar, Cambridge, MA, USA) of 6.5 mm in diameter, with 5-μm pore
filters, as previously described by Fragoso et al(5). In brief, cell lines or different
groups of U937 cells (1×106 cells/ml) were placed in
serum-free medium with or without bevacizumab (1 μg/ml) for 1 h.
Cell aliquots (100 μl) were subsequently added to the upper
compartments, and 600-μl of serum-free medium with or without VEGF
(50 ng/ml) were added to the lower compartments. The transwell
plates were incubated at 37°C, 5% CO2 for 4 h. Cell
migration was determined after 4 h by counting the number of
migrated cells in 6 high-power fields (x400 magnification) under an
optical microscope.
Drug treatment
Seventy-two hours after transfection, 3 groups of
U937 cells in the logarithmic growth phase were harvested and
seeded in 96-well plates at a density of 1×105 cells/ml
(100 μl/well). These cells were then treated with cytarabine or
bevacizumab (10 μl) at various concentrations in RPMI-1640 medium
containing 10% FBS. For each concentration, the experiment was
performed in sextuplicate. Additionally, a normal control and blank
group were also prepared. After 48 h of incubation, cells were
incubated with CCK-8 solution (10 μl) for 4 h followed by blending
(5 min). The absorbance (A) was determined at 450 nm with a
microplate reader and the cell proliferation and inhibition rate
were calculated as follows: proliferation rate/inhibition rate (%)
= [(Acontrol −
Aexperiment)/Acontrol − Ablank)]
×100%. The experiment was repeated 3 times.
Flow cytometry for apoptosis
analysis
After 48 h of treatment with different drugs, 3
groups of U937 cells were collected followed by washing with PBS
twice, then resuspended in PBS (1X) at a density of
1×106 cells/ml. Subsequently, 1 μl of Annexin V-PE and 5
μl of 7-AAD were added followed by blending. A blank group, Annexin
V-PE group and 7-AAD group were also prepared. Cells were incubated
for 15 min followed by flow cytometry for the apoptotic rate.
Flow cytometry for cell cycle
analysis
Three groups of U937 cells were seeded in 24-well
plates at a density of 1×106 cells/ml and maintained in
medium containing different drugs for 48 h. Cells
(5×105) were then collected and washed with PBS (4°C)
twice. The cells were then resuspended in 1 ml of PBS and added to
4 ml of cold absolute ethanol, which was blended and fixed at −20°C
for 15 min followed by centrifugation. The supernatant was removed
and 5 ml of PBS were added to the sediment followed by incubation
for 15 min. Centrifugation was then performed and the supernatant
was removed. The sediment was stained with DNA staining solution
[propidium iodide (PI), 50 mg/l and RNase A, 10 mg/ml] in the dark
for 30 min. In addition, a blank group and PI group were also
prepared. Flow cytometry was performed within 1 h.
Statistical analysis
Statistical analysis was performed using SPSS 14.0
statistical software, and the t-test was used for comparisons
between two means. The χ2 test was conducted for
comparisons between apoptotic rates. A value of P<0.05 was
considered to indicate a statistically significant difference.
Results
Expression of VEGF, VEGFR-1 and VEGFR-2
in acute leukemia cell lines and primary leukemia samples
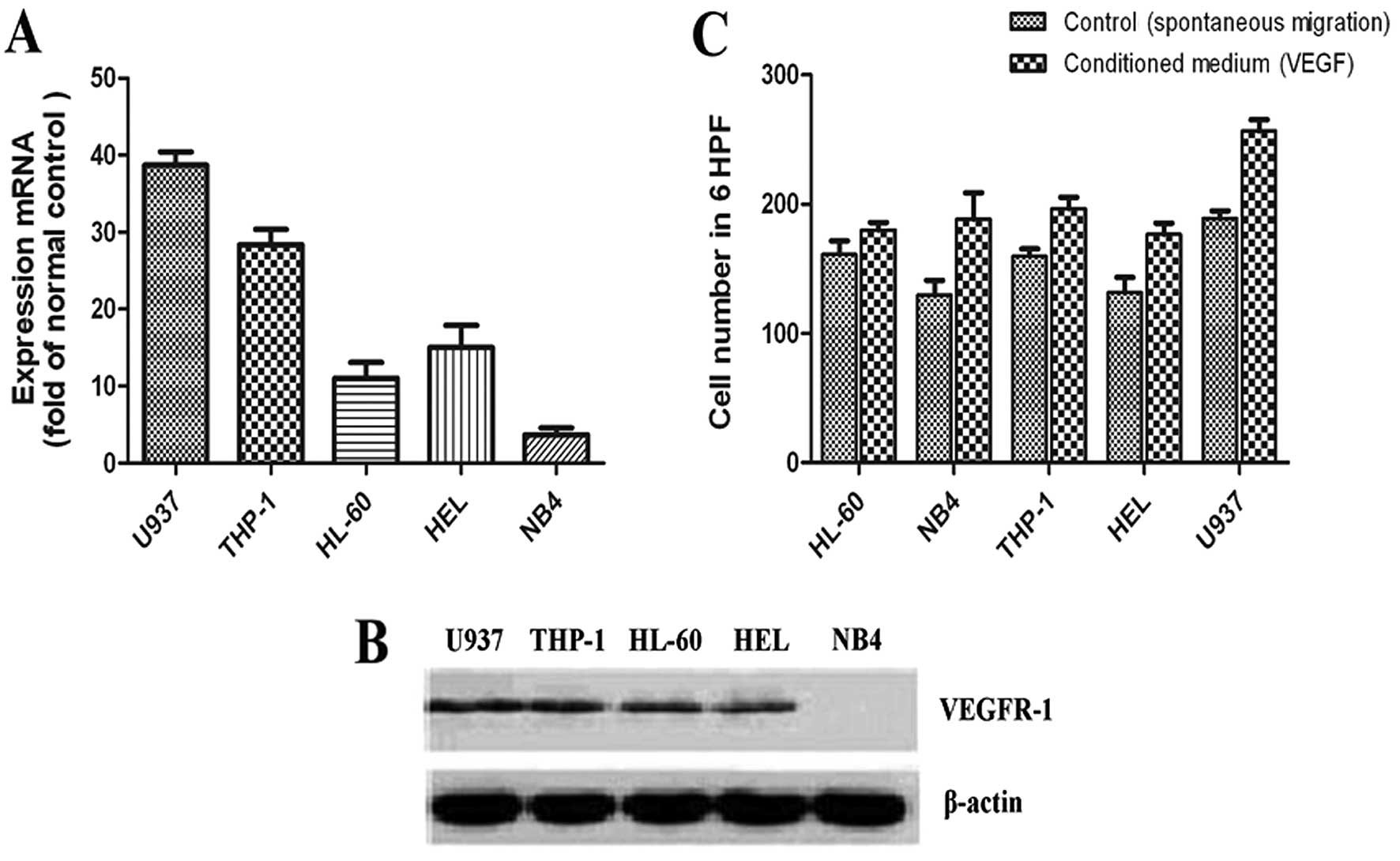
The expression of VEGF, VEGFR-1 and VEGFR-2 was
detected by RT-PCR in 5 AML cell lines. As shown in Table I, VEGF and VEGFR-1 were expressed in
all 5 AML cell lines, whereas only HEL cells expressed VEGFR-2. We
also detected the VEGF protein levels in the cell supernatants by
ELISA (Table I). The U937 cell line
had the highest VEGF protein level and the THP-1 cell line the
second highest (P<0.05 for both). Furthermore, quantitative PCR
and western blot analysis were used to detect VEGFR-1 expression in
the 5 AML cell lines. As shown in Fig.
1A and B, the expression of VEGFR-1 was the highest in the U937
cell line and the lowest in the NB4 cell line.
 | Table IExpressions of VEGF and VEGFRs in 5
AML cell lines. |
Table I
Expressions of VEGF and VEGFRs in 5
AML cell lines.
| Cell line | Cell stage | VEGF | VEGFR-1 | VEGFR-2 | VEGF (ng/ml) |
|---|
| HL-60 | Promyelocytic
leukemia | + | + | − | 0.81±0.13 |
| NB4 | Promyelocytic
leukemia | + | + | − | 0.71±0.09 |
| U937 | Monocytic cells | + | + | − | 1.77±0.07a |
| THP-1 | Monocytic cells | + | + | − | 1.59±0.12a |
| HEL | Erythroleukemia | + | + | + | 0.94±0.11 |
In order to determine whether the migratory behavior
of the leukemia cells is associated with the high expression of
VEGF/VEGFR-1, VEGF-induced cell migration was performed in
transwell migration assays with the 5 AML cell lines. As shown in
Fig. 1C, the U937 cells showed the
most potent chemotactic response to VEGF (with a 2.35-fold ratio of
VEGF-migrated cells/control; P<0.05), the THP-1 cell line showed
the second most potent chemotactic response to VEGF (with a
1.75-fold ratio of VEGF-migrated cells/control; P<0.05), with
the remaining 3 cell lines showing comparable migration
capacity.
As regards the primary leukemia samples, we detected
the mRNA expression of VEGF, VEGFR-1 and VEGFR-2 in 100 patients
with AML. VEGF was expressed in 92% of the AML samples, and VEGFR-1
and VEGFR-2 were expressed in 87 and 37% of the samples,
respectively. The patients were divided into 2 groups according to
the French-American-British (FAB) classification system of acute
leukemia (Table II). The
quantification of VEGF and VEGFR-1 mRNA levels demonstrated
significantly higher levels in the M4/M5 vs. Ml, M2 and M3 FAB
subtypes (P<0.05 for both). In addition, the significantly
higher expression of VEGF and VEGFR-1 mRNA was observed in patients
with extramedullary infiltration vs. those with no extramedullary
infiltration, whereas VEGFR-2 levels did not significantly differ
between the 2 groups.
| Table IIVEGF and VEGFR mRNA expression in
different groups of patients with acute myeloid leukemia. |
Table II
VEGF and VEGFR mRNA expression in
different groups of patients with acute myeloid leukemia.
| Group | VEGF mRNA | VEGFR-1 mRNA | VEGFR-2 mRNA |
|---|
| Extramedullary
infiltration |
| Yes | 1.325±0.23a | 0.987±0.16b | 0.623±0.22 |
| No | 0.976±0.25 | 0.625±0.18 | 0.60±0.13 |
| FAB subtype |
| M4 + M5 | 1.225±0.09c | 1.105±0.16d | 0.645±0.27 |
| Favorable FAB | 0.876±0.25 | 0.587±0.13 | 0.512±0.26 |
Given the high expression level of VEGF and VEGFR-1
in the U937 cells and the M4 and M5 subtypes of AML samples
characteristic of a higher migration rate or extramedullary
infiltration, we hypothesized that VEGF-VEGFR-1 may play
significant roles in the regulation of leukemia cell migration and
other biological characteristics. In the following experiments, the
U937 cells were used as target cells, and lentivirus-mediated RNA
interference was performed to suppress the VEGFR-1 expression in
the U937 cell line.
Lentivirus-mediated RNA interference
targeting VEGFR-1 effectively inhibits the VEGFR-1 expression in
U937 cells
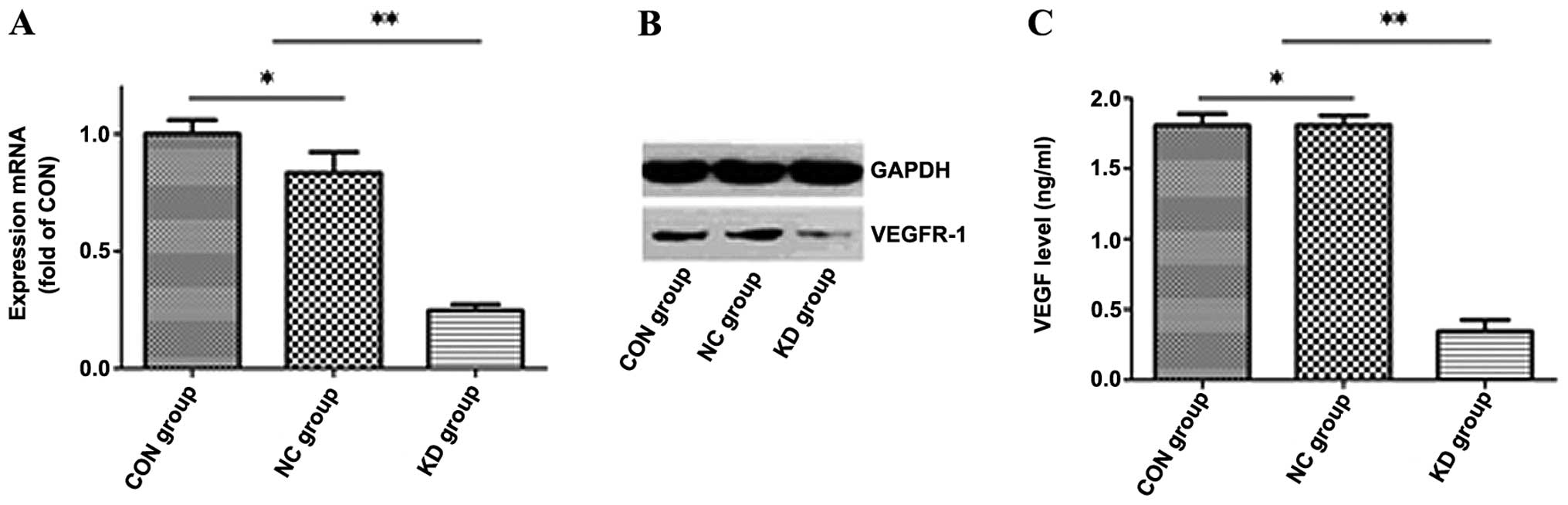
The shRNA-NC and sh-VEGFR-1 KD lentiviral vectors
were used to transfect the U937 cells at a multiplicity of
infection (MOI) of 10. Seventy-two hours after transfection, the
transfection efficiency of the U937 cells was >90%, as observed
under an inverted fluorescence microscope. One hundred and twenty
hours after transfection, we collected the cells and used
quantitative PCR to detect VEGFR-1 mRNA expression. VEGFR-1 mRNA
expression decreased by 73.53±0.026% in the KD group when compared
with the NC group (Fig. 2A).
Western blot analysis showed that the VEGFR-1 protein expression in
the KD group decreased by 94.14% (P<0.05) when compared with the
NC group (Fig. 2B). No significant
difference in the mRNA and protein expression of VEGFR-1 was
observed between the CON and NC group.
Furthermore, we performed ELISA to demonstrate that
there was no significant difference in VEGF production in the U937
cell supernatants in the CON (1.69±0.17 ng/ml) and NC group
(1.65±0.04 ng/ml), while VEGF production in the U937 cell
supernatants in the KD group (0.41±0.06 ng/ml) decreased
dramatically by >75% (P<0.05) when compared to the CON and NC
group (Fig. 2C). These results
indicated that lentivirus-mediated RNA interference suppressed the
autocrine loop of VEGF/VEGFR-1 in the transfected U937 cells.
These results demonstrate the feasibility of
suppressing VEGFR-1 expression by lentivirus-mediated RNA
interference, which inhibits the mRNA and protein expression of
VEGFR-1 and blocks the VEGF/VEGFR-1 autocrine loop in U937
cells.
Lentivirus-mediated RNA interference
targeting VEGFR-1 suppresses the proliferation and migration of
U937 cells
To determine the effects of VEGFR-1 gene silencing
on the biological characteristics of U937 cells, a series of in
vitro experiments were conducted, including cell proliferation
and migration assays, cell cycle analysis and determination of the
apoptotic rate.
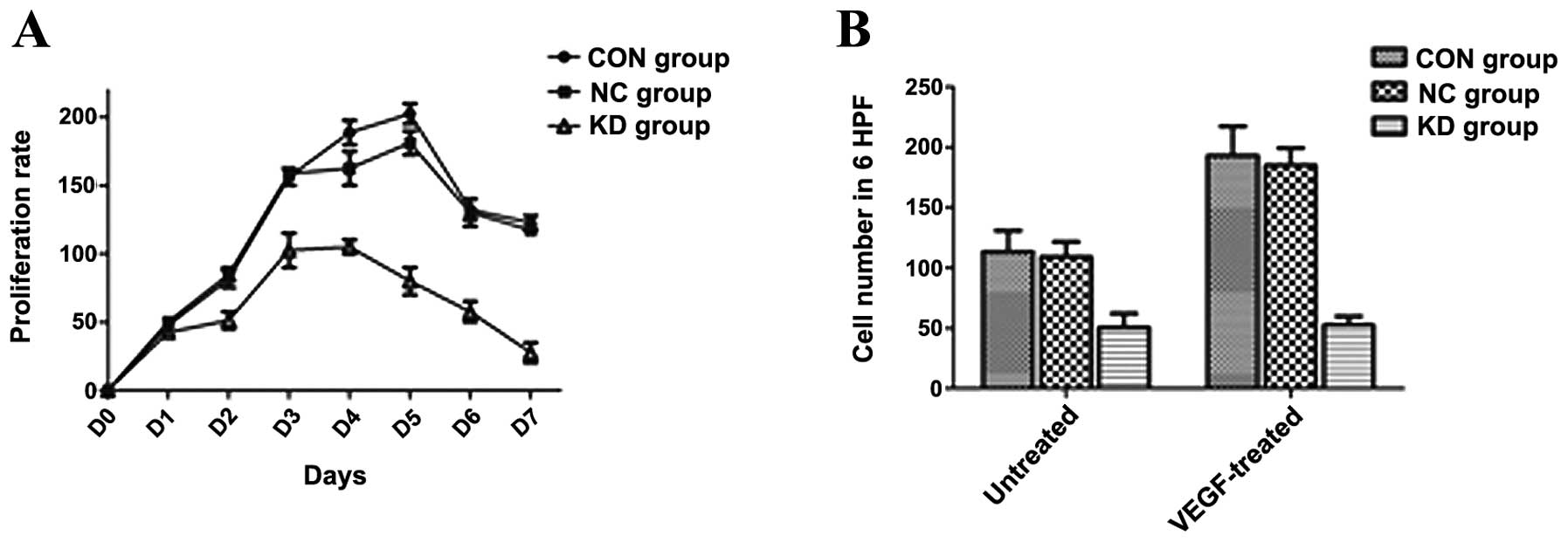
Cells from 3 different groups were grown for 7
consecutive days and the growth curve was delineated according to
the proliferation inhibition rate. As shown in Fig. 3A, the proliferation rate of the U937
cells in the KD group was markedly slower than that of the cells in
the NC and CON group after 3 days of culture (P<0.05), and this
tendency continued for at least 7 days.
Seventy two hours after transfection, 3 groups of
U937 cells were seeded in transwell plates for cell migration
assay. As shown in Fig. 3B, before
drug treatment, the number of migrated cells in the KD group was
markedly lower than that in the NC and CON group (with a 2.5-fold
ratio of VEGF-migrated cells/control; P<0.05). Following
treatment with VEGF, the number of migrated cells in the NC and CON
group was markedly increased, while this phemomenon was not
observed in the KD group.
The apoptotic rate of the U937 cells in the KD group
was slightly higher than that in the CON and NC group (Fig. 4C). No significant difference in cell
cycle distribution was observed among the 3 groups of U937 cells by
cell cycle analysis prior to drug treatment (Fig. 5).
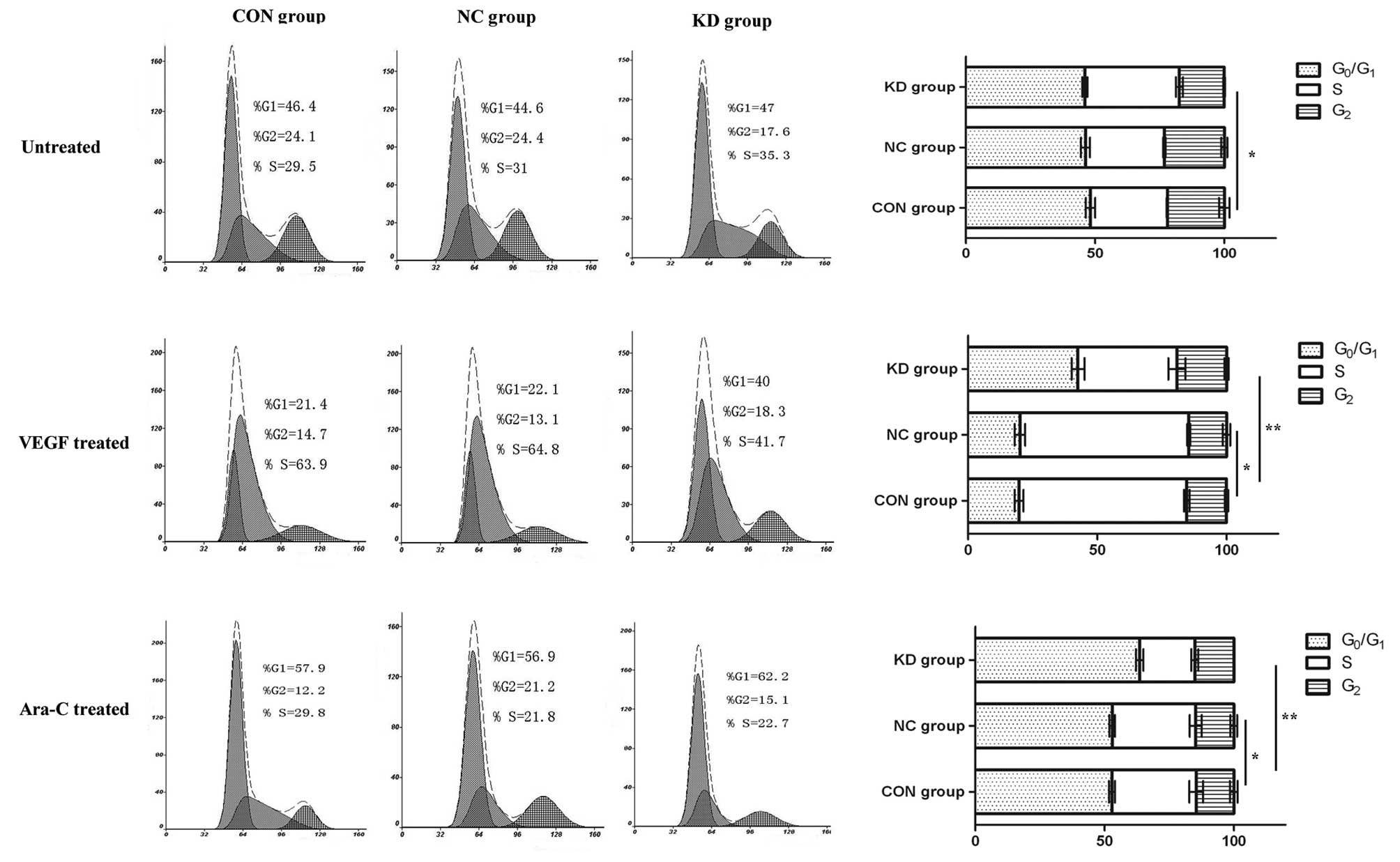 | Figure 5Changes in the cell cycle distribution
of U937 cells in the different groups. Following treatment with
VEGF, the proportion of cells in the S + G2 phase in the
CON and NC group, increased from 53.6 to 78.6% and from 55.4 to
77.9%, respectively, and that in the G1 phase decreased
from 46.4 to 21.4% and from 44.6 to 22.1%, respectively. However,
in the KD group, the proportion of cells in the S + G2
phase increased from 52.9 to 60% and that in the G1
phase decreased from 47 to 40%. Flow cytometry analysis showed that
in the KD group, there were not marked changes in the number of
cells in the S + G2 phase, and the G1 phase
following treatment with VEGF. Ara-C,
cytarabine.*P>0.05, indicates that there is no
statistically significant difference between the groups.
**P<0.05, indicates that there is a statistically
significant difference between the groups. |
Following treatment with VEGF (Fig. 5), the proportion of cells in the S +
G2 phase and G1 phase increased and
decreased, respectively, in the CON and NC group. However, in the
KD group, no marked changes in the proportion of cells in the S +
G2 phase or G1 phase were observed following
treatment with VEGF. These results demonstrate that VEGF enhances
the colony-forming ability of the U937 cells by increasing the
proportion of cells in the S + G2 phase. However, after
VEGFR-1 gene silencing, these effects conferred by VEGF were
blocked, accompanied by a suppressed colony-forming ability. These
findings suggest that VEGFR-1 plays a pivotal role in the
proliferation of leukemia cells.
VEGFR-1 gene silencing in combination
with cytarabine results in increased inhibitory effects on
proliferation and a higher apoptotic rate of U937 cells
As demonstrated above, VEGFR-1 gene silencing
profoundly inhibits the in vitro proliferation and migration
of the U937 cells, but does not directly increase apoptotic rate of
the U937 cells. In the following experiments, VEGFR-1 gene
silencing in combination with bevacizumab or cytarabine were used
to explore their synergistic effects on the inhibition of cell
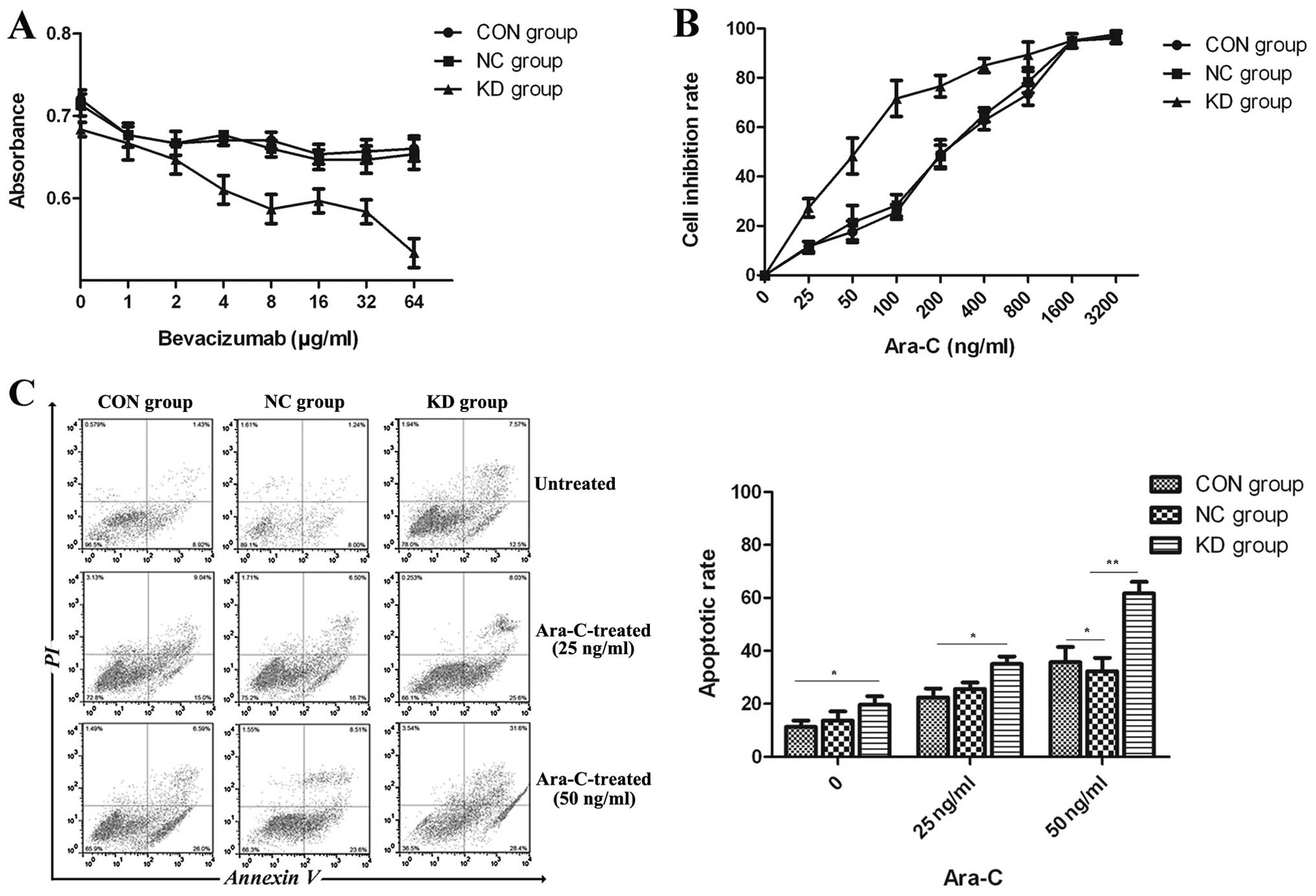
proliferation and apoptotic effects. As shown in Fig. 4A, bevacizumab at various
concentrations did not profoundly affect the proliferation of U937
cells in the CON, NC and KD group. Flow cytometry also showed that
the apoptotic rate in the 3 groups of U937 cells was not affected
by bevacizumab (data not shown). These findings showed that in
vitro VEGFR-1 gene silencing in combination with bevacizumab
did not confer suppressive effects on the proliferation and did not
increase the apoptosis of U937 cells. Following treatment with
various concentrations of cytarabine, a higher number of shrunken
and ruptured cells was observed in the KD group. As indicated by
the growth curve (Fig. 4B), the
inhibition rate increased with the increase in the cytarabine
concentration in each group. When the cytarabine concentration
reached ≥25 ng/ml, the inhibition rate in the KD group dramatically
increased compared to the CON and NC group (P<0.05). The
IC50 of cytarabine in the U937 cells in the KD, CON and
NC group was 50, 200 and 200 ng/ml, respectively. These results
revealed that the sensitivity of U937 cells to cytarabine increased
by VEGFR-1 gene silencing.
The apoptotic rate in the different groups was
detected with a flow cytometer following treatment with cytarabine.
As shown in Fig. 4C, the apoptotic
rate in the KD group was higher than that in the CON and NC group.
When the cytarabine concentration reached 25 or 50 ng/ml, a
significant difference in the apoptotic rate was observed between
the KD group and the other 2 groups (CON and NC group) (P<0.05
for both). Furthermore, the apoptotic rate increased with the
increase in the cytarabine concentration.
Changes in cell cycle distribution and
anti-apoptotic gene expression are associated with the synergistic
effects of VEGFR-1 gene silencing in combination with cytarabine on
U937 cells
To elucidate the mechanisms underlying the
inhibitory effects of VEGFR-1 gene silencing in combination with
cytarabine on the proliferation and increased apoptosis of U937
cells, we further examined the cell cycle distribution in the 3
groups of U937 cells following treatment with cytarabine (50 ng/m).
As shown in Fig. 5, the proportion
of G1 phase cells in the KD group increased from 47 to
62.2%, and that in the S + G2 phase decreased from 53 to
37.8%. However, the proportion of cells in the G1 phase
increased from 46.4 to 51.7% and from 44.6 to 51.9% in the CON and
NC group, respectively, and that in the S + G2 phase
decreased from 53.6 to 48.3% and from 55.4 to 48.1%, respectively.
These results suggested that, after VEGFR-1 gene silencing,
cytarabine treatment markedly decreased the number of cells in the
S + G2 phase and that these cells were more sensitive to
cytarabine when compared with those in the CON and NC group.
We further analyzed the expression of anti-apoptotic
genes in the 3 groups of U937 cells following treatment with
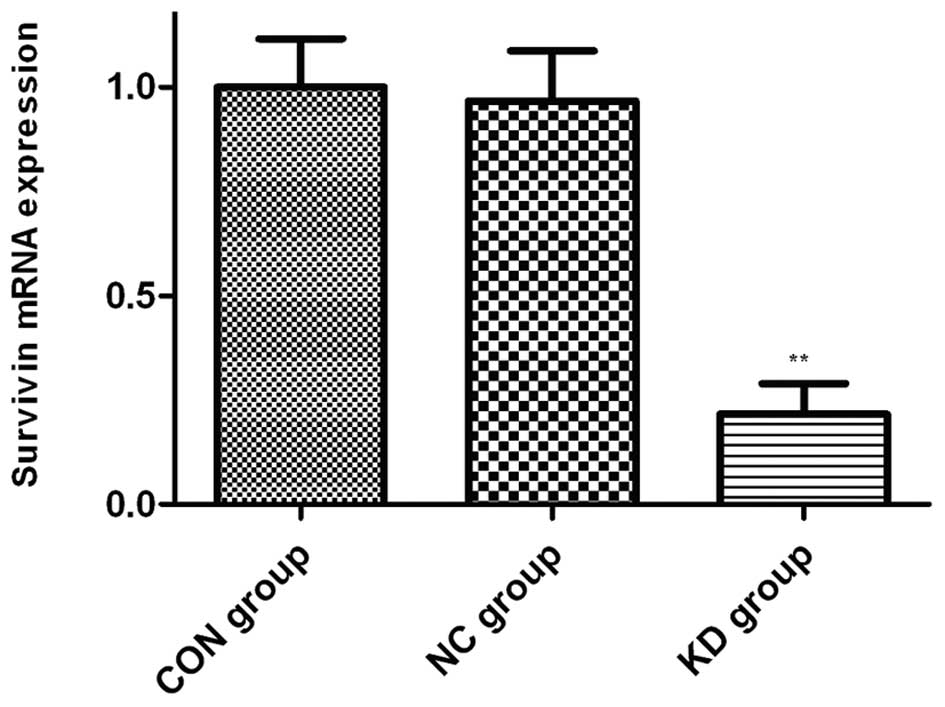
cytarabine. As shown in Fig. 6,
following treatment with cytarabine (50 ng/ml), the expression of
survivin in the KD group was dramatically lower than that in the
CON and NC group. These findings suggest that VEGFR-1 gene
silencing downregulates the expression of anti-apoptotic genes.
Discussion
VEGF and VEGFRs not only promote angiogenesis under
physiological and pathophysiological conditions, but also enhance
the proliferation, survival and migration of leukemia and myeloma
cells, thus resulting in resistance to chemotherapy-induced
apoptosis (4). Previous studies
have demonstrated that the expression of VEGF and its receptors can
be used to evaluate the progression of leukemia, as well as to
predict the outcome of leukemia and monitor minimal residual
leukemia cells (1–3).
Since its discovery, RNAi has been widely used as a
rapid reverse-genetic approach for gene function analysis, as well
as for the ablation of specific genes for therapeutic purposes
(10). Due to their important roles
in tumor neovascularization, VEGF and its receptors have been the
ideal target genes of RNAi. It has been shown that the RNAi
targeting of VEGF in combination with chemotherapy inhibits the
proliferation of cancer cells and promotes their apoptosis
(11). Shen et al(12) used a vector-based siRNA expression
system to specifically inhibit VEGF165 expression in leukemia
cells, leading to decreased tumor vascularity and growth in
vivo.
To date, little is known concerning the role of
VEGFR-1 in AML. In the present study, we used real-time PCR to
detect the mRNA expression of VEGF, VEGR-1, VEGFR-2 in leukemia
cell lines and patients diagnosed with AML. We demonstrated that
the U937 cell line, with the highest migration rate under both
normal and VEGF-induced physiological conditions, presents the
highest expression of VEGF and VEGFR-1. In addition, we
demonstrated that patients with extramedullary disease express
significantly higher levels of VEGF and VEGFR-1 mRNA comapred to
their non-extramedullary migration counterparts. This phenomenon
was also observed in leukemia cells derived from AML M4/M5 compared
to non-M4/M5 subtypes. Therefore, we hypothesized that VEGF and
VEGFR-1 may modulate leukemia survival, proliferation and
migration. Therefore, we used a lentiviral vector-based shRNA
expression system, which induced stable and long-term gene
silencing (13), to elucidate the
role of VEGFR-1 in leukemia biology.
We came to the conclusion that the mRNA and protein
expression of VEGFR-1 was inhibited by pRNAT/sh-VEGFR-1
transfection. In addition, we concluded that transfection with
pRNAT/sh-VEGFR-1 in vitro suppressed the proliferation of
U937 cells and blocked the promotive effects conferred by VEGF. The
suppressed proliferation occurred at 72 h after transfection and
continued for at least 1 week. We hypothesized that there may be a
positive feedback between VEGF and VEGFR-1. Following VEGFR-1 gene
silencing, the expression of VEGFR-1 was suppressed and the
positive stimulatory effect of VEGF/VEGFR-1 was blocked. The amount
of secreted VEGF decreased and the autocrine loop of VEGF was
interrupted, resulting in suppressed proliferation. These results
are in agreement with those from a previous study on
myelodysplastic syndrome (14).
In our study, we observed that in vitro
transfection with pRNAT/sh-VEGFR-1 suppressed the migration of U937
cells and blocked the VEGF-induced cell migration, while Fragoso
et al(5) demonstrated in
vitro and in vivo that VEGFR-1 (FLT-1) activation
results mainly in leukemia cell migration. These results suggest
that VEGFR-1 is associated with the migration of leukemia cells and
that the specific suppression of VEGFR-1 may delay extramedullary
infiltration in leukemia patients.
Bevacizumab is an anti-VEGF monoclonal antibody that
targets VEGF-A isoform and blocks its binding to the VEGF
receptors. Increasing evidence has shown that bevacizumab exhibits
clinical activity against solid tumors when administered with
cytotoxic chemotherapy (15).
However, few studies have investigated bevacizumab treatment in
hematological malignancies. Karp et al(16) conducted a phase II clinical trial of
bevacizumab administered after chemotherapy to adults with
refractory or relapsed AML, showing that cytotoxic chemotherapy
followed by bevacizumab yields a favorable complete response (CR)
rate and duration in adults with AML that is resistant to
traditional treatment approaches. However, other studies (17,18)
have revealed that bevacizumab had no effect on leukemia cells both
in vitro and in vivo. In our study, we demonstrate
in vitro that VEGFR-1 gene silencing in combination with
bevacizumab does not confer synergistic effects on the suppression
of cell proliferation and does not increase the apoptosis of U937
cells, which was consistent with a previous study which indicated
treatment with the anti-VEGF antibody alone had little effect on
cell survival (19). However,
VEGFR-1 gene silencing in combination with cytarabine increased the
sensitivity of U937 cells to cytarabine. As regards the mechanism
by which sh-VEGFR-1 in combination with cytarabine inhibits cell
prolifereation, we demonstrated that transfection with
pRNAT/sh-VEGFR-1 decreased the IC50 of cytarabine in the
U937 cells in the KD group to 25% of that in the CON group, and
increased the apoptotic rate in the KD group, leading to the
enhanced sensitivity of U937 cells in the KD group to chemotherapy.
These effects may be related to the suppressed expression of
VEGFR-1 and VEGF, and the elevated oxygenation process, which
facilitated the entry of the drugs to the tumors and endothelial
cells, resulting in the abolishment of VEGF-induced resistance to
chemotherapy (4). Additionally,
cell cycle assay indicated, following treatment with cytarabine,
that the proportion of cells in the S + G2 phase was
decreased and that the expression of survivin was suppressed by
90%. These findings suggest that the changes in the cell cycle and
survivin expression also play a crucial role in the
pRNAT/sh-VEGFR-1-induced apoptosis. Of note, similar effects were
not observed after treatment with cytarabine in combination with
bevacizumab.
The treatment of tumors with RNAi is still at the
premature stage. A number of issues, such as increasing the
transfection efficiency and ensuring the stability and safety of
shRNA, should be resolved before this approach can be applied in
clinical practice. Therefore, it is necessary to explore effective
approaches for the target-specific suppression of VEGF and
receptors, to construct safe and stable vectors for RNAi, and to
further determine the effects of the RNAi targeting of VEGF and its
receptors on the therapy of leukemia.
In conclusion, in this study, we used a
lentivirus-mediated siRNA expression system to specifically inhibit
VEGFR-1 expression in leukemia cells. The siRNA that we synthesized
reduced the proliferation and migration of the U937 cells and
enhanced their sensitivity to cytarabine. Our data suggest that the
use of sh-VEGFR-1 in the treatment of leukemia, may be of
therapeutic benefit.
Acknowledgements
We thank Zheng Weiping, Sun Xianghua, Zhang Yi and
Zhang Xiaoping for their technical assistance. The present study
was partly supported by the China National Natural Science
Foundation (grant no. 30572161).
References
|
1
|
Aref S, El Sherbiny M, Goda T, Fouda M, Al
Askalany H and Abdalla D: Soluble VEGF/sFLt1 ratio is an
independent predictor of AML patient out come. Hematology.
10:131–134. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
de Bont ES, Fidler V, Meeuwsen T, Scherpen
F, Hahlen K and Kamps WA: Vascular endothelial growth factor
secretion is an independent prognostic factor for relapse-free
survival in pediatric acute myeloid leukemia patients. Clin Cancer
Res. 8:2856–2861. 2002.PubMed/NCBI
|
|
3
|
Liang AB, Li L, Xie XT, et al: Preliminary
study of VEGF and its receptor expression on childhood acute
lymphoblastic leukemia and its relativity to clinical
manifestations. Zhonghua Xue Ye Xue Za Zhi. 26:489–492. 2005.(In
Chinese).
|
|
4
|
Podar K and Anderson KC: The
pathophysiologic role of VEGF in hematologic malignancies:
therapeutic implications. Blood. 105:1383–1395. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fragoso R, Pereira T, Wu Y, Zhu Z,
Cabeçadas J and Dias S: VEGFR-1 (FLT-1) activation modulates acute
lymphoblastic leukemia localization and survival within the bone
marrow, determining the onset of extramedullary disease. Blood.
107:1608–1616. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elbashir SM, Harborth J, Weber K and
Tuschl T: Analysis of gene function in somatic mammalian cells
using small interfering RNAs. Methods. 26:199–213. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hannon GJ: RNA interference. Nature.
418:244–251. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
McManus MT and Sharp PA: Gene silencing in
mammals by small interfering RNAs. Nat Rev Genet. 3:737–747. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Shi Y: Mammalian RNAi for the masses.
Trends Genet. 19:9–12. 2003. View Article : Google Scholar
|
|
10
|
Hannon GJ and Rossi JJ: Unlocking the
potential of the human genome with RNA interference. Nature.
431:371–378. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Filleur S, Courtin A, Ait-Si-Ali S, et al:
SiRNA-mediated inhibition of vascular endothelial growth factor
severely limits tumor resistance to antiangiogenic thrombospondin-1
and slows tumor vascularization and growth. Cancer Res.
63:3919–3922. 2003.
|
|
12
|
Shen HL, Xu W, Wu ZY, Zhou LL, Qin RJ and
Tang HR: Vector-based RNAi approach to isoform-specific
downregulation of vascular endothelial growth factor (VEGF)165
expression in human leukemia cells. Leuk Res. 31:515–521. 2007.
View Article : Google Scholar
|
|
13
|
Manjunath N, Wu H, Subramanya S and
Shankar P: Lentiviral delivery of short hairpin RNAs. Adv Drug
Deliv Rev. 61:732–745. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bellamy WT, Richter L, Sirjani D, et al:
Vascular endothelial cell growth factor is an autocrine promoter of
abnormal localized immature myeloid precursors and leukemia
progenitor formation in myelodysplastic syndromes. Blood.
97:1427–1434. 2001. View Article : Google Scholar
|
|
15
|
Mukherji SK: Bevacizumab (Avastin). AJNR
Am J Neuroradiol. 31:235–236. 2010. View Article : Google Scholar
|
|
16
|
Karp JE, Gojo I, Pili R, et al: Targeting
vascular endothelial growth factor for relapsed and refractory
adult acute myelogenous leukemias: therapy with sequential
1-β-d-arabinofuranosylcytosine, mitoxantrone, and bevacizumab. Clin
Cancer Res. 10:3577–3585. 2004.PubMed/NCBI
|
|
17
|
Kim KJ, Li B, Winer J, et al: Inhibition
of vascular endothelial growth factor-induced angiogenesis
suppresses tumour growth in vivo. Nature. 362:841–844. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zahiragic L, Schliemann C, Bieker R, et
al: Bevacizumab reduces VEGF expression in patients with relapsed
and refractory acute myeloid leukemia without clinical antileukemic
activity. Leukemia. 21:1310–1312. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Santos SC and Dias S: Internal and
external autocrine VEGF/KDR loops regulate survival of subsets of
acute leukemia through distinct signaling pathways. Blood.
103:3883–3889. 2004. View Article : Google Scholar : PubMed/NCBI
|