Introduction
Human gastric cancer is one of the most common
malignancies in the world, with 934,000 new cases occurring each
year. The overall 5-year survival rate of gastric cancer remains
less than 20%. Advanced disease stages and tumor metastasis
contribute to the high mortality rate in patients with gastric
cancer (1), which is associated
with abnormalities in tumor intracellular signaling networks.
During tumor development, environmental and genetic factors
interact to contribute to the dysregulation of gene signaling
pathways, which include JAK-STAT, Wnt, MAPK and PI3K (2–6).
Studies have shown that abnormal expression of
microRNAs (miRNAs, miRs), small non-coding RNAs ~18–22 nucleotides
in length that post-transcriptionally regulate expression of target
genes, is one of the key mechanisms in cancer development, invasion
and metastasis (7–9). Many studies have shown altered
expression of various miRNAs in human carcinogenesis. For example,
miR-451, miR-101, let-7 and miR-17–92 have been reported to
regulate the invasion and metastasis of tumor cells by targeting
the PI3K/Akt and MAPK pathways (10–13).
In gastric cancer, miRNAs have been found to play
important roles in tumor initiation and progression, including
miR-20, miR-34b, miR-128, miR-129, miR-148, miR-150 and miR-21
(14,15). Other miRNAs (miR-15b, miR-16,
miR-372 and miR-150) affected gastric cancer apoptosis and drug
resistance (16–18). The miRNAs miR-218, miR-141, miR-375,
miR-101, miR-21, miR-449 and miR-107 are implicated in the
regulation of gastric cancer cell differentiation, invasion and
metastasis (19–25). Further investigation of aberrant
miRNA expression in gastric cancer cells may provide insightful
information for a better understanding of gastric cancer and the
mechanisms underlying gastric carcinogenesis. Such investigations
could help us to develop novel strategies for future prevention,
diagnosis and treatment of gastric cancer.
To this end, in the present study we first profiled
differentially expressed miRNAs in gastric cancer cell lines
relative to a normal stomach mucosal epithelial cell line. We then
investigated miR-7 expression in gastric cancer tissue specimens
and the effects of miR-7 on growth, invasion and metastasis of
gastric cancer cells and the underlying molecular events. The miR-7
gene is localized at the intron region of the heterogeneous
ribonucleic protein k gene on human chromosome 9, and altered miR-7
expression is associated with the progression of numerous types of
tumors since miR-7 can target and inhibit expression of the
epidermal growth factor receptor (EGFR), Pak1, Bcl-2 and IGF1R in
breast cancer, Schwann cell tumors, tongue squamous cell carcinoma
and non-small cell lung cancer (26–29).
Materials and methods
Cell lines and culture
The human gastric adenocarcinoma cell lines SGC-7901
(moderately differentiated) and MKN-45 (poorly differentiated) and
the normal human gastric epithelial cell line GES-1 were obtained
from the Cell Bank of Xiangya Central Laboratory, Central South
University (Changsha, Hunan, China). The human gastric carcinoma
cell lines NCI-N87 (well differentiated) and MGC-803 (poorly
differentiated) were obtained from the Cell Bank of the Chinese
Academy of Sciences (Shanghai, China).
MGC-803, MKN-45 and SGC-7901 cells were maintained
in RPMI-1640 medium. NCI-N87 and GES-1 cells were maintained in
Dulbecco’s modified Eagle’s medium (DMEM; HyClone-Pierce, Logan,
UT, USA) supplemented with 10% fetal bovine serum (FBS; Invitrogen,
Carlsbad, CA, USA), penicillin (100 U/ml; Sigma Chemicals, St.
Louis, MO, USA), and streptomycin (100 μg/ml; Sigma Chemicals). All
cells were cultured in a humidified incubator with 5%
CO2 at 37°C.
RNA isolation and miRNA microarray
profiling
Total cellular RNA was isolated from cells of each
cell line using TRIzol reagent (Invitrogen) and purified with an
RNeasy Mini kit (Qiagen, Hilden, Germany). After being labeled
using the miRCURY Array Power Labeling kit (Exiqon, Vedbaek,
Denmark), these RNA samples were used as probes to hybridize with
the miRCURY microRNA Array kit (Exiqon) in a Hybridization Chamber
II (Ambion, Austin, TX, USA) in accordance with the manufacturer’s
instructions. Hybridized miRNA microarrays were then scanned using
a GenePix 4000B microarray scanner (Axon Instruments, Inc., Union
City, CA, USA), and the data were analyzed using GenePix Pro
software v6.0 (Axon). Differential miRNA expression (fold change
>1.50 or ratio <0.667) and cluster analysis were performed
using MeV software (v4.6, TIGR). Some of these differentially
expressed miRNAs were validated using qRT-PCR.
Gastric cancer tissue specimens
Gastric cancer tissue samples were obtained from 23
patients who underwent partial gastrectomy between March and
November 2011 at the First Affiliated Hospital and Second
Affiliated Hospital of the University of South China (Hengyang,
China). Patients included 12 men aged 60.9±9 years and 11 women
aged 52.8±9.5 years. These patients did not receive any
radiotherapy and chemotherapy before surgery.
Gastric cancer tissues and matched non-tumor tissues
were obtained from surgical resections and were each divided into
two parts: one for pathological diagnosis and another for
extracting total RNA. Pathological diagnoses were performed
independently by two pathologists in accordance with the WHO
standard for pathological diagnosis of gastric cancer. Nine of
these cases were histologically diagnosed as well/moderately
differentiated gastric cancers, while 14 were diagnosed as poorly
differentiated gastric cancers.
The present study was approved by our institutional
review boards of the First and Second Affiliated Hospitals of the
University of South China. Each patient provided informed consent
before participation in the study.
qRT-PCR
Total cellular RNA was isolated from the gastric
cell lines and tissues using TRIzol reagent (Invitrogen) and then
reverse transcribed to cDNA using a Hairpin-it miRNA qPCR
quantitation kit (Shanghai GenePharma Co., Ltd., Shanghai, China)
in accordance with the instructions. The PCR primers were:
hsa-miR-7 (to generate an 82-bp PCR product),
5′-CCACGTTGGAAGACTAGTGATTT-3′ and 5′-TATGGTTGTTCTGCTCTCTGTCTC-3′;
human epidermal growth factor receptor (EGFR; to generate a 93-bp
PCR product), 5′-AAAGAATACCATGCAGAAGGAGG-3′ and
5′-GACATCACTCTGGTGGGTATAGA-3′; U6 (as an internal control, to
generate a 70-bp PCR product), 5′-ATTGGAA CGATACAGAGAAGATT-3′ and
5′-GGAACGCTTCACGA ATTTG-3′, and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH, an internal control, to generate a 113-bp PCR
product), 5′-CATGAGAAGTATGACAACAGCCT-3′ and
5′-AGTCCTTCCACGATACCAAAGT-3′. All of the primers were obtained from
Shanghai GenePharma. The qRT-PCR conditions consisted of an initial
95°C for 3 min; and 40 cycles of 95°C for 30 sec and 62°C for 40
sec. To generate the melting curves, the PCR amplification was
carried out by slowly heating at 0.1°C/sec increments from 60 to
94°C.
Construction of the miR-7 expression
vector and gene transfection
To construct the miR-7 expression vector, we
synthesized a mature miR-7 sequence (5′-TGGAAGACTAG
TGATTTTGTTGT-3′) and inserted these double-stranded
oligonucleotides into a linearized PcDNATM6.2-GW/EmGFP-miR vector
(Invitrogen). After amplification, we sequenced this vector to
confirm successful cloning. We also constructed a negative control
vector using non-sense sequences (5′-AAATGTACTGCGCGTGGAGAC-3′) to
insert into a pcDNA 6.2-GW/miR vector. For gene transfection, we
grew MGC-803 cells and transfected the above vectors into this cell
line using GBfectene-Elite (Genebank Biosciences, Suzhou, China) in
accordance with the manufacturer’s instructions.
The following experimental groups were created:
untreated, transfection reagent only, negative control vector and
miR-7 vector. The transfection rate was confirmed using an inverted
fluorescence microscope and qRT-PCR 48 h after transfection.
Cell viability CCK-8 assay
Cells in each group were suspended and inoculated
into 96-wells at 1,000 cells/well. After reaching 80% confluency,
the cells were transfected with plasmids (0.05 μg/well) using
GBfectene-Elite in accordance with the manufacturer’s instructions,
and cultured for 72 h. At the end of the experiment, 10 μl of the
CCK-8 solution (Yiyuan Biotechnology, Guangzhou, China) was added
into each well and the cells were further incubated at 37°C in a
humidified incubator with 5% CO2 for 1 h. Optical
densities were measured at an absorbance of 450 nm using an ELX800
enzyme-linked immunoassay analyzer (BioTek, Winooski, VT, USA). The
average and standard deviation of each group were calculated and
the data are presented in a histogram.
Transwell tumor cell migration and
invasion assays
A Transwell chamber (PengBo Biotech, Changsha,
China) was used to detect gastric cancer cell migration and
invasion capacity. For invasion, the membrane in the Transwell
chamber was coated with Matrigel basement membrane matrix (BD
BioCoat, Bedford, MA, USA). The miR-7-transfected or control
vector-transfected cells were suspended at 1.0×106/ml in
DMEM containing 1.0% BSA. One hundred microliters was added into
the upper chamber of the Transwell, and 500 μl DMEM containing 20%
FBS was added into the lower chamber. The cells were incubated for
48 h. At the end of the experiments, the cells on the upper side of
the membrane were carefully wiped out, and the cells that had
migrated or invaded into the lower side of the membrane were fixed
with 4% paraformaldehyde and stained with
4′,6-diamidino-2-phenylindole (DAPI). Images were then captured,
and the number of cells was counted in 5 random fields for each
group and summarized as mean ± standard deviation (SD) for
statistical analysis.
Protein extraction and western blot
analysis
Cells were washed 3 times with phosphate-buffered
saline (PBS) and lysed in RIPA buffer (Beyotime Institute of
Biotechnology, Shanghai, China) to extract the total cellular
protein. Equal amounts of protein samples were resolved via 8%
sodium dodecyl sulfate-polyacrylamide gel electrophoresis
(SDS-PAGE) and transferred onto a polyvinylidene difluoride (PVDF)
membrane (Beyotime Institute of Biotechnology). After blocking with
a blocking buffer (GenePharma) for 1 h, the membrane was incubated
with a primary antibody against EGFR (Proteintech Group, Chicago,
IL, USA) or GADPH (Sigma-Aldrich) at a dilution of 1:500 at 4°C
overnight. The next day the membranes were washed with PBS thrice
and then further incubated with a horseradish peroxidase-conjugated
secondary antibody (Jackson ImmunoResearch, West Grove, PA, USA) at
a dilution of 1:1,000 at 37°C for 1 h. After washing with PBS
thrice, positive protein bands were visualized using SuperSignal
West Pico chemiluminescence substrates (Thermo Fisher Scientific,
Rockford, IL, USA). Image analysis was performed with Alpha Imager
2200 (UVP, Upland, CA, USA).
Dual-luciferase reporter gene assay
To assess the direct binding of miR-7 to EFGR cDNA,
we performed a luciferase assay. In brief, the cells were
co-transfected with EGFR 3′UTR-pmirGLO plasmid and hsa-mir-7 mimic
vector or the negative control plasmid hsa-miR-NC using
Lipofectamine 2000 (Invitrogen) in accordance with the
manufacturer’s instructions. Twenty-four or 48 h after
transfection, the cells were prepared for the luciferase assay
using the Dual-luciferase reporter assay system (Promega, Madison,
WI, USA) in accordance with the kit instructions. Luciferase
activity was detected with a GloMax™20/20 Luminometer (Promega).
The relative luciferase activity was defined as the ratio of
firefly luciferase activity to Renilla luciferase activity.
Δ luciferase activity was used for statistical analysis of data,
where Δ luciferase activity = (Firefly/Renilla)
sample/(Firefly/Renilla) control.
Statistical analysis
Statistical analyses were performed using the
statistical software SPSS 16.0 (SPSS, Inc., Chicago, IL, USA). All
data are expressed as mean ± standard deviation. Differences
between groups were analyzed with one-way ANOVA and the least
significant difference test; P<0.05 was considered to indicate a
statistically significant result.
Results
Differentially expressed miRNAs in
gastric cancer cells
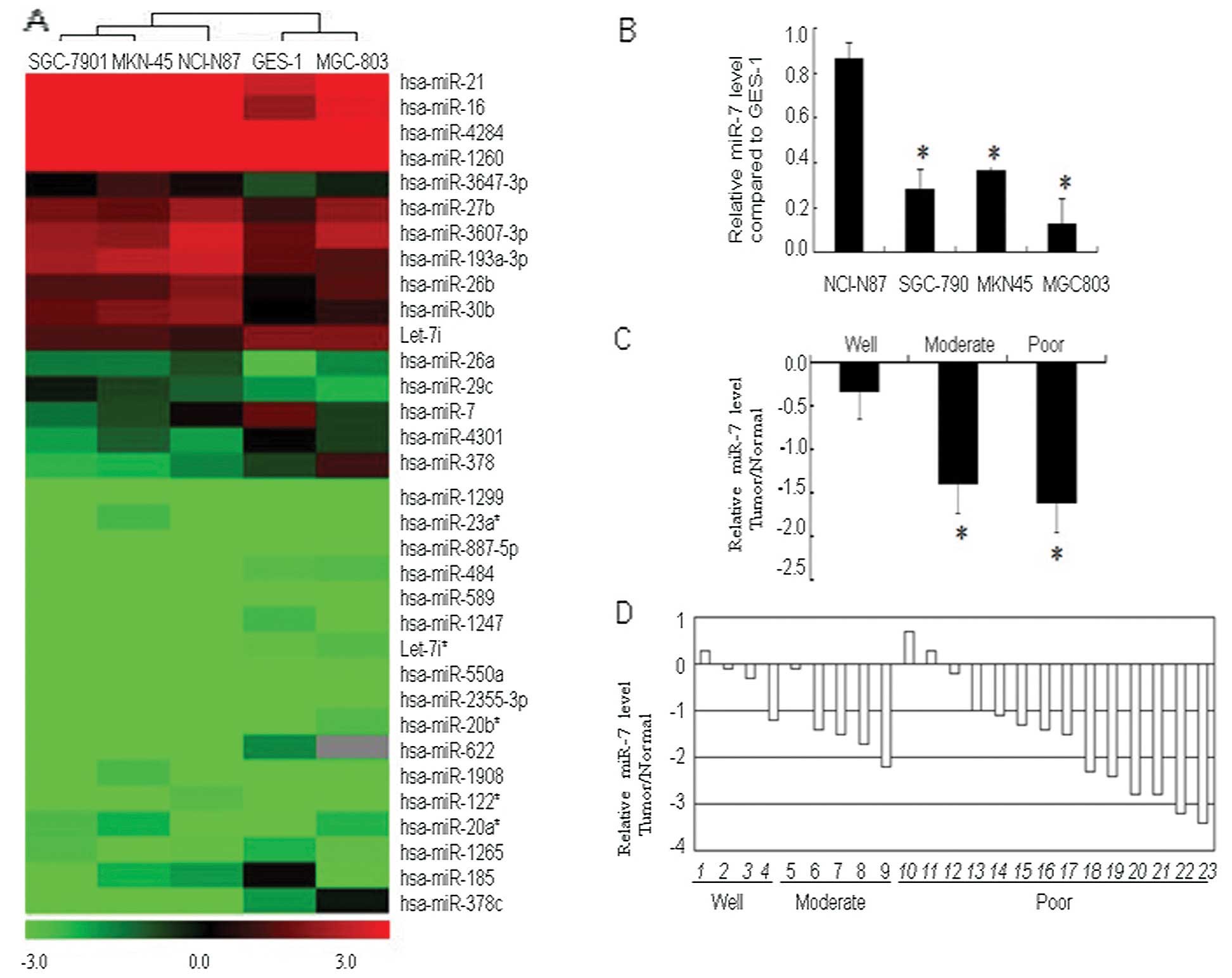
We performed a microRNA microarray assay to identify
differentially expressed miRNAs in gastric cancer cell lines
(MGC-803, MKN-45, SGC-7901 and NCI-N87) that varied in the degrees
of tumor differentiation, as well as in the normal gastric mucosal
epithelial cell line GES-1 (Fig.
1A). In these gastric cancer cell lines, we found 14 miRNAs
that were overexpressed relative to the normal gastric mucosal
GES-1 cells (ratio >1.5 between tumor and normal cells). These
overexpressed miRNAs were: hsa-miR-23a, hsa-miR-27b, hsa-miR-1908,
hsa-miR-26b, hsa-miR-3607-3p, hsa-miR-20a*, hsa-miR-21,
hsa-miR-122*, hsa-miR-193a-3p, hsa-miR-26a, hsa-miR-29c,
hsa-miR-16, hsa-miR-30b and hsa-miR-3647-3p. In contrast, 19 miRNAs
were reduced by 1.5-fold or more (i.e., ratio <0.667 between
tumor and normal cells). The miRNAs that were underexpressed were:
hsa-miR-1247, hsa-miR-20b*, hsa-miR-4301, hsa-miR-1299,
hsa-miR-484, hsa-miR-885-5p, hsa-miR-1265, hsa-miR-589,
hsa-miR-550a, let-7i*, hsa-miR-2355-3p, hsa-miR-4284, let-7i,
hsa-miR-7, hsa-miR-1260, hsa-miR-622, hsa-miR-185, hsa-miR-378 and
hsa-miR-378c.
The miRNA hsa-miR-7 was downregulated in all four
gastric cancer cell lines comprising well-, moderately and poorly
differentiated gastric cancer cells, relative to normal GES-1
cells. Thus, we chose hsa-miR-7 for further investigation. The
underexpression of hsa-miR-7 in the NCI-N87, SGC-7901, MKN-45 and
MGC-803 cell lines was confirmed via qRT-PCR (Fig. 1B).
We then assessed miR-7 expression in gastric cancer
tissue specimens, and found that relative to the normal tissues,
expression of miR-7 was decreased in 86.9% of gastric cancer
tissues. Levels of miR-7 expression were significantly lower in
moderately and poorly differentiated cancer tissues when compared
with that in the well-differentiated gastric cancer tissues
(Fig. 1C and D).
Expression of miR-7 inhibits gastric
cancer cell viability and invasion
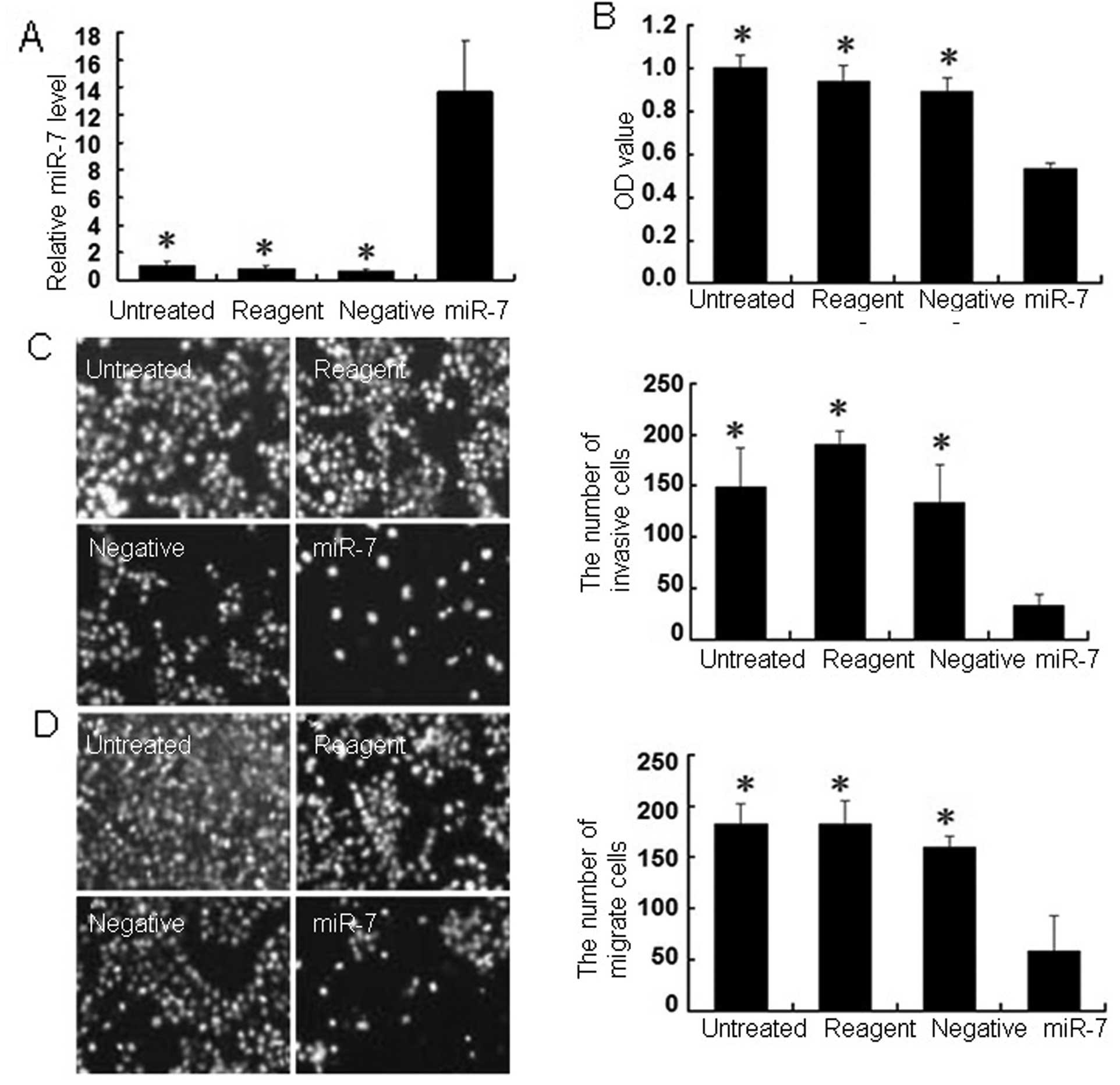
To further assess the role of miR-7 in gastric
cancer, we constructed a miR-7 expression vector for gene
transfection and found that the transfection efficiency of the
miR-7 plasmid and negative control groups was >80% in MGC803
cells after a 48-h transfection, via observation under a
fluorescence microscope. The qRT-PCR data showed that levels of
miR-7 expression were significantly greater 48 h after miR-7
plasmid transfection compared with the negative control cells
(Fig. 2A). Restoration of miR-7
expression reduced gastric cancer cell viability (Fig. 2B) compared with the controls.
We used a Transwell chamber to detect the effects of
miR-7 on the invasion and migration abilities of MGC803 gastric
cancer cells. The miR-7-transfected cells exhibited significantly
reduced invasion and migration abilities (33±11 and 59±34,
respectively) compared with the negative control cells (135±37 and
160±11), transfection reagent only cells (191±12 and 183±23), and
the parental cells (149±39 and 182±20; Fig. 2C and D).
Expression of the target gene EGFR is
inhibited by miR-7
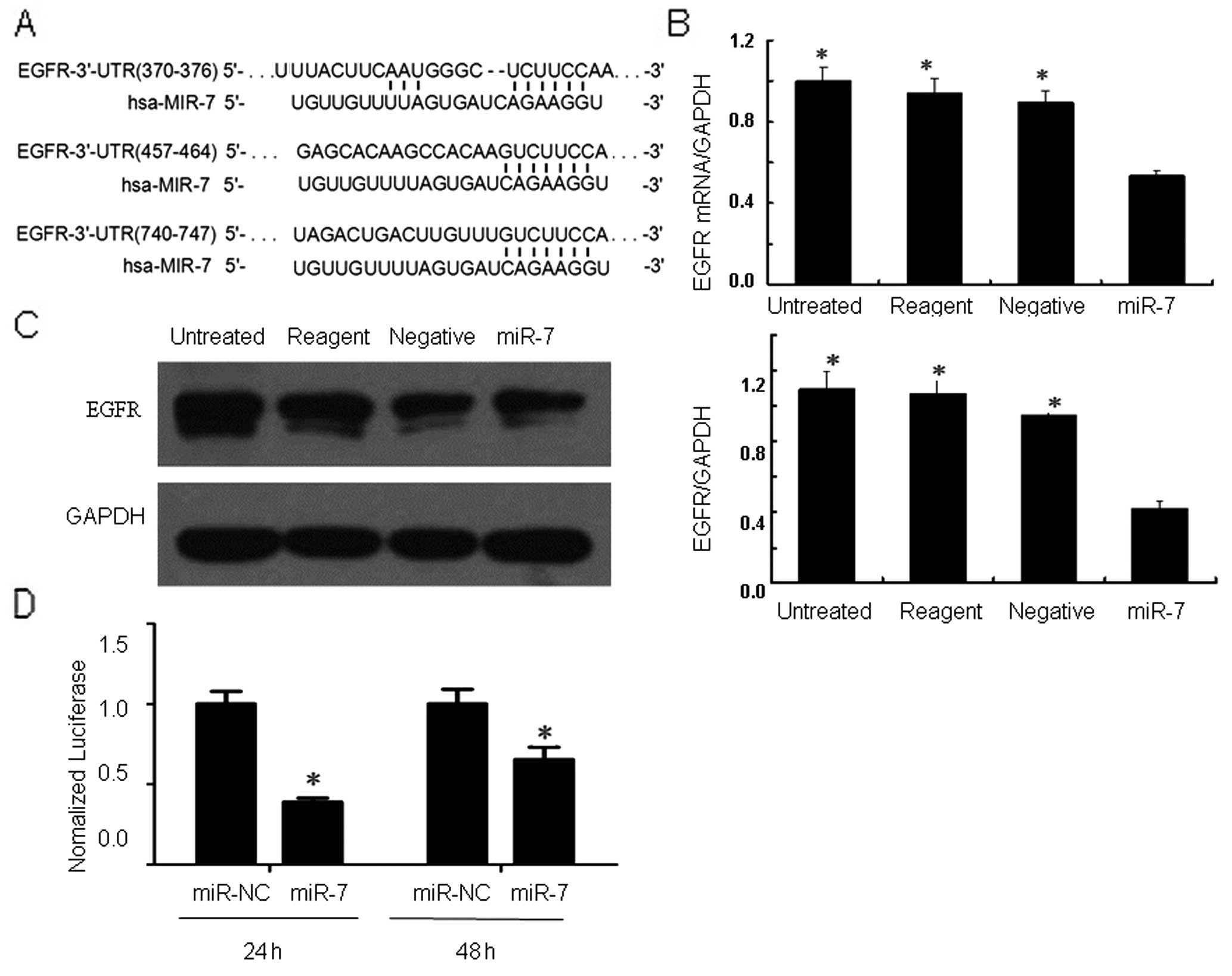
To explore miR-7 targeting (binding) genes, we
performed a bioinformatic analysis using TargetScan, PicTar and
miRanda software and found that EGFR could be a target gene
(Fig. 3A). To confirm whether miR-7
regulates EGFR expression by binding to the EFGR 3′UTR, we
performed a dual luciferase reporter assay. The results showed that
24 and 48 h after co-transfection of EGFR 3′UTR-pmirGLO plasmid and
hsa-miR-7, fluorescent activity was significantly reduced when
compared to that of the miR-NC control group (Fig. 3D).
We then performed qRT-PCR and western blot analysis
and found that levels of EGFR mRNA and protein were significantly
less in the miR-7-transfected cells when compared with the levels
in the negative control, transfection reagent only cells and the
parental cells (Fig. 3B and C).
These data suggest that miR-7-induced inhibition of gastric cancer
viability and invasion acts through degradation of EGFR mRNA and a
decrease in EGFR protein expression.
Discussion
In the present study, we first profiled
differentially expressed miRNAs in gastric cancer cells relative to
normal cells, since a great number of studies have shown that
altered miRNA expression is closely associated with gastric cancer
development and progression (14–25).
Indeed, our data revealed a group of miRNAs that were either
upregulated or downregulated in cells of the gastric cancer cell
lines of various differentiation levels relative to cells of the
normal mucosal cell line. We then focused on miR-7, which was
downregulated in all four of the gastric cancer cell lines tested.
Consistent with this finding, miR-7 expression was reduced in the
gastric cancer tissues, particularly in moderately and poorly
differentiated gastric cancer tissues compared to
well-differentiated tumors. Furthermore, restoration of miR-7
expression inhibited tumor cell viability, migration and invasion
ability. At the gene level, miR-7 was able to bind to the EGFR
3′UTR and suppress expression of EGFR mRNA and EGFR protein. The
data from the present study indicate that miRNA expression was
altered in gastric cancer cells and tissue specimens and that
restoration of miR-7 expression inhibited gastric cancer cell
viability, migration and invasion. These results suggest the role
of aberrant miR-7 expression in gastric cancer progression and the
underlying molecular pathways.
Previous studies have shown that differential
expression of various miRNAs not only contributes to carcinogenesis
and cancer progression, but can also predict the histological type
of cancer, drug resistance and clinical outcomes (7). One study utilized 200 miRNAs to
molecularly classify human cancers (30), which may help clinicians to more
effectively treat cancer patients. However, to date, there has been
no miRNA expression profile study in gastric cancer, while an
individual miRNA study showed controversial data (31). In the present study, we selected
gastric cancer cell lines with different degrees of differentiation
(i.e., well, moderate and poor) relative to normal gastric mucosal
cells, to profile aberrant expression of miRNAs and then verify
them in gastric cancer cell lines and tissue samples. The purpose
of this approach was to identify miRNAs that may be related to
gastric cancer progression (such as high metastatic potential). Our
data showed that compared with normal gastric mucosal cells, 14
miRNAs were upregulated, whereas 19 miRNAs were downregulated in
the four gastric cancer cell lines tested. The altered expression
of miR-7, miR-16, miR-21, hsa-miR-30b, let-7i, hsa-miR-622 and
hsa-miR-378 found in the present study was consistent with the
findings of others (14,32–37),
yet not the altered expression of miR-185 and miR-29c (22,38).
However, there have been no previous reports of the differential
expression of miR-27b, miR-378c and miR-484 in gastric cancer
cells. Although data regarding aberrant expression of miR-124,
miR-126 and miR-204 have been reported for gastric cancer cells
(39–41), the present study did not find any
alterations in gastric cancer cell lines. This may be because of
differences in the cell lines tested, particularly in the level of
cancer cell differentiation and the screening criteria for
differential miRNA expression.
The miRNA microarray and RT-PCR data of the present
study showed that miR-7 expression was significantly lower in the
NCI-N87, SGC-7901, MGC-803 and MKN-45 cell lines, which was further
confirmed in gastric carcinoma tissues and matched adjacent
non-tumor tissues. Restoration of miR-7 expression reduced gastric
cancer cell viability, migration and invasion ability. Indeed, the
current emphasis regarding the clinical control of human cancer
concerns the suppression of cancer cell proliferation, invasion and
metastasis, all of which affect tumor classification, treatment and
clinical outcomes. Poorly differentiated gastric cancer is more
aggressive and is defined by increased proliferation, invasion and
metastasis (42), making this type
of cancer difficult to treat resulting in a poor patient prognosis.
It has been reported that transfection of miR-7 precursor into
human gastric cancer AZ521 and Kato III cell lines significantly
inhibited gastric cancer cell proliferation (33). Overexpression of miR-7 in the
gastric cancer GC9811-P cell line suppressed tumor cell migration
and invasion (43). This study is
consistent with these previous studies, although different tumor
cell lines were used. These data together indicate that miR-7 may
play an important role in gastric cancer progression.
Molecularly, miR-7 has been shown to downregulate
EGFR expression in glioma, and cancers of the prostate, lung, and
breast (44,45), but there has been no previous report
of the role of miR-7 in the regulation of EGFR expression in
gastric cancer. As we know, the expression of miRNAs are
tissue-specific and time dependent, with different tissues or cells
at different stages of biological development expressing various
miRNAs (8). In the present study,
we performed a bioinformatic analysis and predicted that EGFR cDNA
contains hsa-MIR-7 targeting sequences. We then performed a
luciferase assay to confirm the interaction between miR-7 and the
EGFR 3′UTR in human gastric cancer cell lines. Western blot
analysis and qRT-PCR data further revealed that miR-7 suppressed
levels of EGFR mRNA and protein in human gastric cancer cell
lines.
EGFR is involved in the regulation of tumor cell
proliferation, angiogenesis, invasion, metastasis and inhibition of
apoptosis through downstream signal transduction pathways, and is
closely correlated with tumor initiation, progression and poor
prognosis of gastric cancer (46–52).
The downstream of EGFR signal transduction pathways include the
Ras/Raf/MEK/ERK-MAPK and PI3K/Akt/mTOR pathways (4,53). The
ERK-MAPK signaling pathway was found to induce cell tumor
proliferation by inactivating cell cycle inhibitory protein p27KIP
via phosphorylation (54) or by
activating matrix metalloproteinases to enhance hydrolysis of the
extracellular matrix and invasion of tumor cells (55), in addition to participation in the
regulation of apoptosis of tumor cells. Furthermore, the PI3K/Akt
pathway can activate mTOR signaling networks to promote translation
of key proteins for cell growth (56,57).
The present study is proof-of-principle and future
studies will be carried out to investigate these gene pathways in
the control of gastric cancer development and progression. We will
explore whether miR-7 offers a novel strategy for gastric cancer
therapy.
Acknowledgements
We thank Dr Xiaoyong Lei of the Life Science and
Technology College of the University of South China, Dr Rongfang He
of the Pathology Department of the First Affiliated Hospital of the
University of South China and Dr Yong Fang of the Pathology
Department of the Second Affiliated Hospital of the University of
South China for their technical support and for providing tissue
specimens for the present study. We also thank Medjaden Bioscience
Ltd., Hong Kong, China, for assisting in the editing of this
manuscript. The study was supported in part by a grant from the
National Natural Science Foundation of China (#81071965).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Ben-Chetrit N, Tarcic G and Yarden Y:
ERK-ERF-EGR1, a novel switch underlying acquisition of a motile
phenotype. Cell Adh Migr. 7:33–37. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Thu KL, Radulovich N, Becker-Santos DD, et
al: SOX15 is a candidate tumor suppressor in pancreatic cancer with
a potential role in Wnt/β-catenin signaling. Oncogene. 33:279–288.
2014.PubMed/NCBI
|
|
4
|
Hill KS, Erdogan E, Khoor A, et al:
Protein kinase Cα suppresses Kras-mediated lung tumor formation
through activation of a p38 MAPK-TGFβ signaling axis. Oncogene. Apr
22–2013.(Epub ahead of print).
|
|
5
|
Nojima M, Suzuki H, Toyota M, et al:
Frequent epigenetic inactivation of SFRP genes and
constitutive activation of Wnt signaling in gastric cancer.
Oncogene. 26:4699–4713. 2007.PubMed/NCBI
|
|
6
|
Xu G, Zhang W, Bertram P, Zheng XF and
McLeod H: Pharmacogenomic profiling of the PI3K/PTEN-AKT-mTOR
pathway in common human tumors. Int J Oncol. 24:893–900.
2004.PubMed/NCBI
|
|
7
|
Krützfeldt J, Poy MN and Stoffel M:
Strategies to determine the biological function of microRNAs. Nat
Genet. 38:S14–S19. 2006.
|
|
8
|
Bartel DP: MicroRNAs: genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Carthew RW: Gene regulation by microRNAs.
Curr Opin Genet Dev. 16:203–208. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hayashita Y, Osada H, Tatematsu Y, et al:
A polycistronic microRNA cluster, miR-17–92, is overexpressed in
human lung cancers and enhances cell proliferation. Cancer Res.
65:9628–9632. 2005.
|
|
11
|
Lee YS and Dutta A: The tumor suppressor
microRNA let-7 represses the HMGA2 oncogene. Genes Dev.
21:1025–1030. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nan Y, Han L, Zhang A, et al: miRNA-451
plays a role as tumor suppressor in human glioma cells. Brain Res.
1359:14–21. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang JG, Guo JF, Liu DL, Liu Q and Wang
JJ: MicroRNA-101 exerts tumor-suppressive functions in non-small
cell lung cancer through directly targeting enhancer of zeste
homolog 2. J Thorac Oncol. 6:671–678. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chan SH, Wu CW, Li AF, Chi CW and Lin WC:
miR-21 microRNA expression in human gastric carcinomas and its
clinical association. Anticancer Res. 28:907–911. 2008.PubMed/NCBI
|
|
15
|
Katada T, Ishiguro H, Kuwabara Y, et al:
microRNA expression profile in undifferentiated gastric cancer. Int
J Oncol. 34:537–542. 2009.PubMed/NCBI
|
|
16
|
Xia L, Zhang D, Du R, et al: miR-15b and
miR-16 modulate multidrug resistance by targeting BCL2 in human
gastric cancer cells. Int J Cancer. 123:372–379. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cho WJ, Shin JM, Kim JS, et al: miR-372
regulates cell cycle and apoptosis of ags human gastric cancer cell
line through direct regulation of LATS2. Mol Cells. 28:521–527.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu Q, Jin H, Yang Z, et al: MiR-150
promotes gastric cancer proliferation by negatively regulating the
pro-apoptotic gene EGR2. Biochem Biophys Res Commun. 392:340–345.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Varambally S, Cao Q, Mani RS, et al:
Genomic loss of microRNA-101 leads to overexpression of histone
methyltransferase EZH2 in cancer. Science. 322:1695–1699. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang Z, Li Z, Gao C, et al: miR-21 plays
a pivotal role in gastric cancer pathogenesis and progression. Lab
Invest. 88:1358–1366. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Du Y, Xu Y, Ding L, Yao H, Yu H, Zhou T
and Si J: Down-regulation of miR-141 in gastric cancer and its
involvement in cell growth. J Gastroenterol. 44:556–561. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Takahashi Y, Forrest AR, Maeno E,
Hashimoto T, Daub CO and Yasuda J: miR-107 and miR-185 can induce
cell cycle arrest in human non small cell lung cancer cell lines.
PLoS One. 4:e66772009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ding L, Xu Y, Zhang W, et al: MiR-375
frequently downregulated in gastric cancer inhibits cell
proliferation by targeting JAK2. Cell Res. 20:784–793. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tie J, Pan Y, Zhao L, et al: MiR-218
inhibits invasion and metastasis of gastric cancer by targeting the
Robo1 receptor. PLoS Genet. 6:e10008792010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
BouKheir T, Futoma-Kazmierczak E, Jacobsen
A, et al: miR-449 inhibits cell proliferation and is down-regulated
in gastric cancer. Mol Cancer. 10:292011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Reddy SD, Ohshiro K, Rayala SK and Kumar
R: MicroRNA-7, a homeobox D10 target, inhibits p21-activated kinase
1 and regulates its functions. Cancer Res. 68:8195–8200. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Saydam O, Senol O, Würdinger T, et al:
miRNA-7 attenuation in Schwannoma tumors stimulates growth by
upregulating three oncogenic signaling pathways. Cancer Res.
71:852–861. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiong S, Zheng Y, Jiang P, Liu R, Liu X
and Chu Y: MicroRNA-7 inhibits the growth of human non-small cell
lung cancer A549 cells through targeting BCL-2. Int J Biol Sci.
7:805–814. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang L, Liu X, Chen Z, et al: MicroRNA-7
targets IGF1R (insulin-like growth factor 1 receptor) in tongue
squamous cell carcinoma cells. Biochem J. 432:199–205. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lu J, Getz G, Miska EA, et al: MicroRNA
expression profiles classify human cancers. Nature. 435:834–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yao Y, Suo AL, Li ZF, et al: MicroRNA
profiling of human gastric cancer. Mol Med Rep. 2:963–970.
2009.PubMed/NCBI
|
|
32
|
Guo XB, Jing CQ, Li LP, et al:
Down-regulation of miR-622 in gastric cancer promotes cellular
invasion and tumor metastasis by targeting ING1 gene. World J
Gastroenterol. 17:1895–1902. 2011.PubMed/NCBI
|
|
33
|
Kong D, Piao YS, Yamashita S, et al:
Inflammation-induced repression of tumor suppressor miR-7 in
gastric tumor cells. Oncogene. 31:3949–3960. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shin VY, Jin H, Ng EK, et al: NF-κB
targets miR-16 and miR-21 in gastric cancer: involvement of
prostaglandin E receptors. Carcinogenesis. 32:240–245. 2011.
|
|
35
|
Inoue T, Iinuma H, Ogawa E, Inaba T and
Fukushima R: Clinicopathological and prognostic significance of
microRNA-107 and its relationship to DICER1 mRNA expression in
gastric cancer. Oncol Rep. 27:1759–1764. 2012.PubMed/NCBI
|
|
36
|
Liu K, Qian T, Tang L, Wang J, Yang H and
Ren J: Decreased expression of microRNA let-7i and its association
with chemotherapeutic response in human gastric cancer. World J
Surg Oncol. 10:2252012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Deng H, Guo Y, Song H, et al: MicroRNA-195
and microRNA-378 mediate tumor growth suppression by epigenetical
regulation in gastric cancer. Gene. 518:351–359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Matsuo M, Nakada C, Tsukamoto Y, et al:
MiR-29c is downregulated in gastric carcinomas and regulates cell
proliferation by targeting RCC2. Mol Cancer. 12:152013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Xia J, Wu Z, Yu C, et al: miR-124 inhibits
cell proliferation in gastric cancer through down-regulation of
SPHK1. J Pathol. 227:470–480. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Feng R, Chen X, Yu Y, et al: miR-126
functions as a tumour suppressor in human gastric cancer. Cancer
Lett. 298:50–63. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sacconi A, Biagioni F, Canu V, et al:
miR-204 targets Bcl-2 expression and enhances responsiveness of
gastric cancer. Cell Death Dis. 3:e4232012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Catalano V, Labianca R, Beretta GD, Gatta
G, de Braud F and Van Cutsem E: Gastric cancer. Crit Rev Oncol
Hematol. 71:127–164. 2009. View Article : Google Scholar
|
|
43
|
Zhao X, Dou W, He L, et al: MicroRNA-7
functions as an anti-metastatic microRNA in gastric cancer by
targeting insulin-like growth factor-1 receptor. Oncogene.
32:1363–1372. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Webster RJ, Giles KM, Price KJ, Zhang PM,
Mattick JS and Leedman PJ: Regulation of epidermal growth factor
receptor signaling in human cancer cells by microRNA-7. J Biol
Chem. 284:5731–5741. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Giles KM, Barker A, Zhang PM, Epis MR and
Leedman PJ: MicroRNA regulation of growth factor receptor signaling
in human cancer cells. Methods Mol Biol. 676:147–163. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Nicholson RI, Gee JM and Harper ME: EGFR
and cancer prognosis. Eur J Cancer. 37:S9–S15. 2001. View Article : Google Scholar
|
|
47
|
Saif MW: Colorectal cancer in review: the
role of the EGFR pathway. Expert Opin Investig Drugs. 19:357–369.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lieto E, Ferraraccio F, Orditura M, et al:
Expression of vascular endothelial growth factor (VEGF) and
epidermal growth factor receptor (EGFR) is an independent
prognostic indicator of worse outcome in gastric cancer patients.
Ann Surg Oncol. 15:69–79. 2008. View Article : Google Scholar
|
|
49
|
Arkenau HT: Gastric cancer in the era of
molecularly targeted agents: current drug development strategies. J
Cancer Res Clin Oncol. 135:855–866. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
She QB, Solit DB, Ye Q, O’Reilly KE, Lobo
J and Rosen N: The BAD protein integrates survival signaling by
EGFR/MAPK and PI3K/Akt kinase pathways in PTEN-deficient tumor
cells. Cancer Cell. 8:287–297. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Corcoran RB, Ebi H, Turke AB, et al:
EGFR-mediated re-activation of MAPK signaling contributes to
insensitivity of BRAF mutant colorectal cancers to RAF inhibition
with vemurafenib. Cancer Discov. 2:227–235. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim MA, Lee HS, Lee HE, Jeon YK, Yang HK
and Kim WH: EGFR in gastric carcinomas: prognostic significance of
protein overexpression and high gene copy number. Histopathology.
52:738–746. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Toulany M, Dittmann K, Krüger M, Baumann M
and Rodemann HP: Radioresistance of K-Ras mutated human tumor cells
is mediated through EGFR-dependent activation of PI3K-AKT pathway.
Radiother Oncol. 76:143–150. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Donovan JC, Milic A and Slingerland JM:
Constitutive MEK/MAPK activation leads to p27Kip1
deregulation and antiestrogen resistance in human breast cancer
cells. J Biol Chem. 276:40888–40895. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Stamenkovic I: Matrix metalloproteinases
in tumor invasion and metastasis. Semin Cancer Biol. 10:415–433.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Pires MM, Hopkins BD, Saal LH and Parsons
RE: Alterations of EGFR, p53 and PTEN that mimic changes found in
basal-like breast cancer promote transformation of human mammary
epithelial cells. Cancer Biol Ther. 14:246–253. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Britschgi A, Bill A, Brinkhaus H, et al:
Calcium-activated chloride channel ANO1 promotes breast cancer
progression by activating EGFR and CAMK signaling. Proc Natl Acad
Sci USA. 110:E1026–E1034. 2013. View Article : Google Scholar : PubMed/NCBI
|