Introduction
Epidemiological studies have revealed an increased
risk of cardiovascular disease associated with high local doses of
ionizing radiation to the heart. This can be observed in patients
who have received thoracic radiotherapy treatment for breast
cancer, Hodgkin's disease, or childhood cancers (1,2). A
significant increase in mortality from cardiovascular disease has
been reported among patients treated by radiotherapy (3,4).
Typically, cardiac symptoms appear late, sometimes decades after
exposure to radiation, and include increased infiltration of
inflammatory cells and fibrosis of the myocardium, especially after
receiving high radiation doses (5).
In addition to vascular abnormalities, recent studies have
emphasized cardiac fibrosis as a major causal factor for
radiation-induced heart disease (6),
a condition that is accompanied by an accumulation of fibroblasts
and fibroblast-to-myofibroblast transition (FMT) (7).
One area of interest is the potential for
mesenchymal stem cells (MSCs) to repair injured cardiac tissue.
MSCs possess the capacity for self-renewal, multipotency, and
cytokine secretion (8,9), characteristics which render them ideal
for tissue repair or regeneration applications (10,11).
Recent studies have indicated that MSCs could potentially exert
anti-fibrotic effects in the context of cardiac disease, which may
allow for repair of the damaged myocardium (12). Previous studies have revealed that
MSCs pre-treated with hypoxia secrete a variety of cytokines such
as vascular endothelial growth factor, hepatocyte growth factor
(HGF), basic fibroblast growth factor, interleukin-10, as well as
others, which act to inhibit cardiac fibrosis (13,14).
TGF-β is considered a ‘master’ cytokine/growth
factor produced within injured or diseased tissues, where it
activates fibroblasts and facilitates the production of
extracellular matrix (ECM), leading to tissue fibrosis (15). TGF-β binds a heterodimeric receptor in
the plasma membrane consisting of the TGF-β type I and type II
half-receptors, which together induce phosphorylation of Smad3
transcription factors in the canonical signaling pathway (16). Smad7 acts as an inhibitor of TGF-β
signaling by interacting with the TGF-β type I receptor, preventing
the phosphorylation and activation of Smad3 (17). It has been reported that Smad7 can
block TGF-β-induced fibroblast accumulation, inhibiting FMT, which
can further inhibit fibrosis (18).
The pathogenesis of cardiac fibrosis involves
alterations in the cellular and neuro-humoral environments, leading
to changes in fibroblast activity and ECM turnover (19,20). The
oxidative stress that is induced by radiation is further increased
in response to mechanical and metabolic stress, triggering
increased activation and proliferation of fibroblasts, which in
turn leads to tissue fibrosis (21,22).
Notably, abundant data have revealed that MSCs have a strong
capacity to suppress oxidative stress, leading to an inhibition of
cardiac fibrosis and protective effects (23).
Based on the aforementioned observation, it was
hypothesized that hypoxia pre-conditioned MSCs would be superior to
MSCs or hypoxia in isolation in inhibiting radiation-induced
cardiac fibrosis.
Materials and methods
Cell culture
Normal human cardiac fibroblasts (HCFs) were
purchased from Cell Applications, Inc. Fibroblasts were maintained
in low glucose Dulbecco's modified Eagle's media (DMEM)
supplemented with 10% fetal bovine serum (FBS), 1%
penicillin-streptomycin (Beyotime Institute of Biotechnology), and
100 µg/ml streptomycin (Life Technologies; Thermo Fisher
Scientific, Inc.).
Bone marrow-derived human MSCs (BM-hMSCs) were
purchased from ATCC, and seeded at 5,000 cells/cm2 in
DMEM-F12 growth media with 1% Glutamax (Sigma-Aldrich; Merck KGaA)
and 1% penicillin-streptomycin, and were grown to 80–85%
confluency.
Cell treatment
In order to induce radiation exposure,
1×105 fibroblasts grown in 6-well plates were irradiated
in a 15×15 cm2 square field with 5 Gy (6-MeV electron
rays; current, 6 A; dose rate, 2 Gy/min) (X RAD 225; PRECISION
X-ray, Inc.).
A Transwell system was used to prevent direct
contact between hMSCs and HCFs. hMSCs and HCFs were placed in the
upper and lower layers of the Transwell plate, respectively, at a
density of 1×106 cells/well.
Hypoxia pre-conditioning was performed by incubating
MSCs for 30 min in serum-free media in a controlled atmosphere
(anaerobic chamber) glove box (855-AC; Plas Labs, Inc.) to scavenge
for free oxygen.
To block the HGF function, we used anti-HGF antibody
to neutralize the HGF. The anti-HGF antibody was added one hour
before cell treatment, and this antibody was purchased from Sigma
Aldrich; Merck KGaA (product no. H0652).
Cell proliferation assay
Fibroblast proliferation was determined using an MTT
Cell Proliferation Assay kit (ATCC) according to the manufacturer's
instructions. Briefly, 300 µl of MTT reagent was added to each well
(1×105 cells) 3 h prior to harvesting. Absorbance at 540
nm was recorded using an enzyme-linked immunosorbent assay (ELISA)
plate reader. Three repeats were performed.
Quantitative reverse transcription
polymerase chain reaction (qRT-PCR)
Total RNA was isolated from cardiac fibroblasts with
TRIzol (Ambion; ThermoFisher Scientific, Inc.). An ultraviolet
spectrophotometer was used to quantify the RNA. cDNA was
synthesized from 1 µg of total RNA using Superscript II reverse
transcriptase (Invitrogen), as per the manufacturer's protocol. The
qPCR was carried out using the Fast Start Universal SYBR Master and
a fluorescence quantitative PCR system. RT-PCR was conducted as
previously described (24). The
relative expression levels were calculated with the
2−∆∆Cq method (25), using
GAPDH as the internal control. Primer sets (Invitrogen;
ThermoFisher Scientific, Inc.) used are listed in Table I.
 | Table I.Primer sequences. |
Table I.
Primer sequences.
| Genes | Sequences |
|---|
| Col1 | F:
5′-CCTGGAAAGAATGGAGATGATG-3′ |
|
| R:
5′-ATCCAAACCACTGAAACCTCTG-3′ |
| α-SMA | F:
5′-TGACAATGGCTCTGGGCTCTGTAA-3′ |
|
| R:
5′-TTCGTCACCCACGTAGCTGTCTTT-3′ |
| HGF | F:
5′-ACCCTGGTGTTTCACAAGCA-3′ |
|
| R:
5′-GCAAGAATTTGTGCCGGTGT-3′ |
| TGF-β1 | F:
5′-GGACACCAACTATTGCTTCAG-3′ |
|
| R:
5′-TCCAGACTCCAAATGTAG-3 |
| GAPDH | F:
5′-GGGCTGCTTTTAACTCTGGT-3′ |
|
| R:
5′-GCAGGTTTTTCTAGACGG-3′ |
Western blot analysis
Cardiac fibroblasts were lysed with ice-cold lysis
buffer (Beyotime Institute of Biotechnology, Haimen, China) to
obtain total protein. Protein samples (20 µg) were quantified using
BCA Protein Assay kit, and separated by 10% SDS-PAGE, and then the
proteins were transferred to polyvinylidene difluoride membranes
(EMD Millipore). Then, the membranes were blocked with 5% milk in
Tris-buffered saline, followed by incubation with the appropriate
primary antibodies: Smad7 (1:1,000; product code ab90086; Abcam);
Smad3 (1:1,000; product no. 9513); p-Smad3 (1:1,000; product no.
9520); and β-actin, (1:2,000; product no. 4970; all from CST)
overnight at 4°C. Membranes were then incubated with horseradish
peroxidase-conjugated secondary antibodies (1:2,000 dilution; cat.
no. 7074; from Cell Signaling Technology, Danvers, MA, USA), and
developed using High Sensitivity ECL Substrate kit (product code
ab133406; Abcam). The stained protein bands were visualized using
Bio-Rad ChemiDoc XRS equipment, and quantified and analyzed using
Quantity One software. Three repeats were performed.
Immunofluorescence
After treatment, cardiac fibroblasts were seeded in
cell culture dishes, fixed in 4% paraformaldehyde, and
permeabilized with 0.1% Triton X-100, followed by blocking in 3%
bovine serum albumin (BSA), washed by phosphate-buffered saline
(PBS). They were then immunolabeled with specific primary
antibodies [alpha smooth muscle actin (α-SMA); 1:200; product code
ab32575; Abcam] overnight at 4°C. After rinsing, the cells were
incubated with the corresponding FITC-conjugated secondary
antibodies (1:250) (product code ab7086; Abcam) in 1% BSA for 1 h
at 37°C. Nuclei were stained with DAPI for 5 min at room
temperature. Fluorescence was detected under a fluorescence
microscope.
ELISA
The concentration of HGF in the supernatant was
determined using an HGF Human ELISA kit (Abcam). Samples from each
group (groups as follows: Control; Radiation; Radiation +
MSCsHypoxia; Radiation + MSCsNormoxia) were
collected in sterile tubes and centrifuged at 1,500 × g for 15 min
to obtain supernatants. The supernatants were analyzed according to
the manufacturer's instructions.
Small interfering (si)RNA
transfection
siRNAs were used to knock down Smad7 in fibroblasts.
A non-targeting siRNA was used as a negative control (Invitrogen;
ThermoFisher Scientific, Inc.). The target sequences were as
follows: Smad7, GCAGCCTAACCAGACCTTT; Control, AACCTGCGGGAAGAAGTGG.
Transfection efficiency was detected by western blotting.
Microarray
Cardiac fibroblasts irradiated were immediately
lysed in 500 µl TRIzol (Thermo Fisher Scientific, Inc.) and stored
at −80°C before purification by a standard phenol-chloroform
extraction protocol using the RNAqueous Micro kit (Thermo Fisher
Scientific, Inc.). The transcriptome was analyzed by microarray
using an Affymetrix human array (Thermo Fisher Scientific, Inc.)
and normalized based upon quantiles.
Measurement of reactive oxygen species
(ROS) production
Cells were detached from culture plates using 0.25%
trypsin-EDTA, collected in 5-ml round-bottom polystyrene tubes, and
washed with 1X Wash Buffer (PBS). The cell suspension was
centrifuged for 5 min at 400 × g at room temperature, and the
supernatant was discarded. The cell pellet was resuspended in 500
µl of ROS/Superoxide Detection Solution. Cells were incubated for
30 min at 37°C in the dark. Data were acquired on a FACScan
instrument (BD Biosciences) and analyzed using CellQuest software
(BD Biosciences).
Superoxide dismutase (SOD)
activity
SOD activity in cells was determined using a
colorimetric assay kit (Abcam), according to the manufacturer's
instructions. Briefly, protein was isolated from cells using lysis
buffer (supplied by the kit), and SOD activity was measured using
10 µg of total protein extract. Absorbance was measured at 450
nm.
Lipid peroxidation assay
Lipid peroxidation was measured using an assay kit
(Abcam) that measures the formation of malondialdehyde (MDA),
according to the manufacturer's instructions. Briefly, fibroblasts
(1×106 cells) were homogenized on ice in 300 µl of MDA
lysis buffer (containing 3 µl of 100X butylated hydroxytoluene,
supplied by the kit), then centrifuged (13,000 × g for 10 min) to
remove insoluble material. The supernatant (200 µl) was added to
600 µl of thiobarbituric acid and incubated at 95°C for 60 min.
Samples were then cooled to room temperature in an ice bath for 10
min, and the absorbance at 532 nm was measured on a
spectrophotometer.
Determination of 4-hydroxynonenal
(4-HNE) levels
The level of 4-HNE was evaluated using commercially
available Lipid Peroxidation (4-HNE) Assay kit (product code
ab238538), according to the manufacturer's instructions.
Statistical analysis
Data are expressed as the mean ± standard deviation
(SD). Differences between groups were tested using a one-way
analysis of variance followed by Tukey's multiple comparisons test,
and comparisons between two groups were evaluated using a Student's
t-test. Analyses were performed using the SPSS package v19.0
(SPSS, Inc.). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of hypoxia-treated MSCs
(MSCsHypoxia) on radiation-induced fibroblast
proliferation and FMT
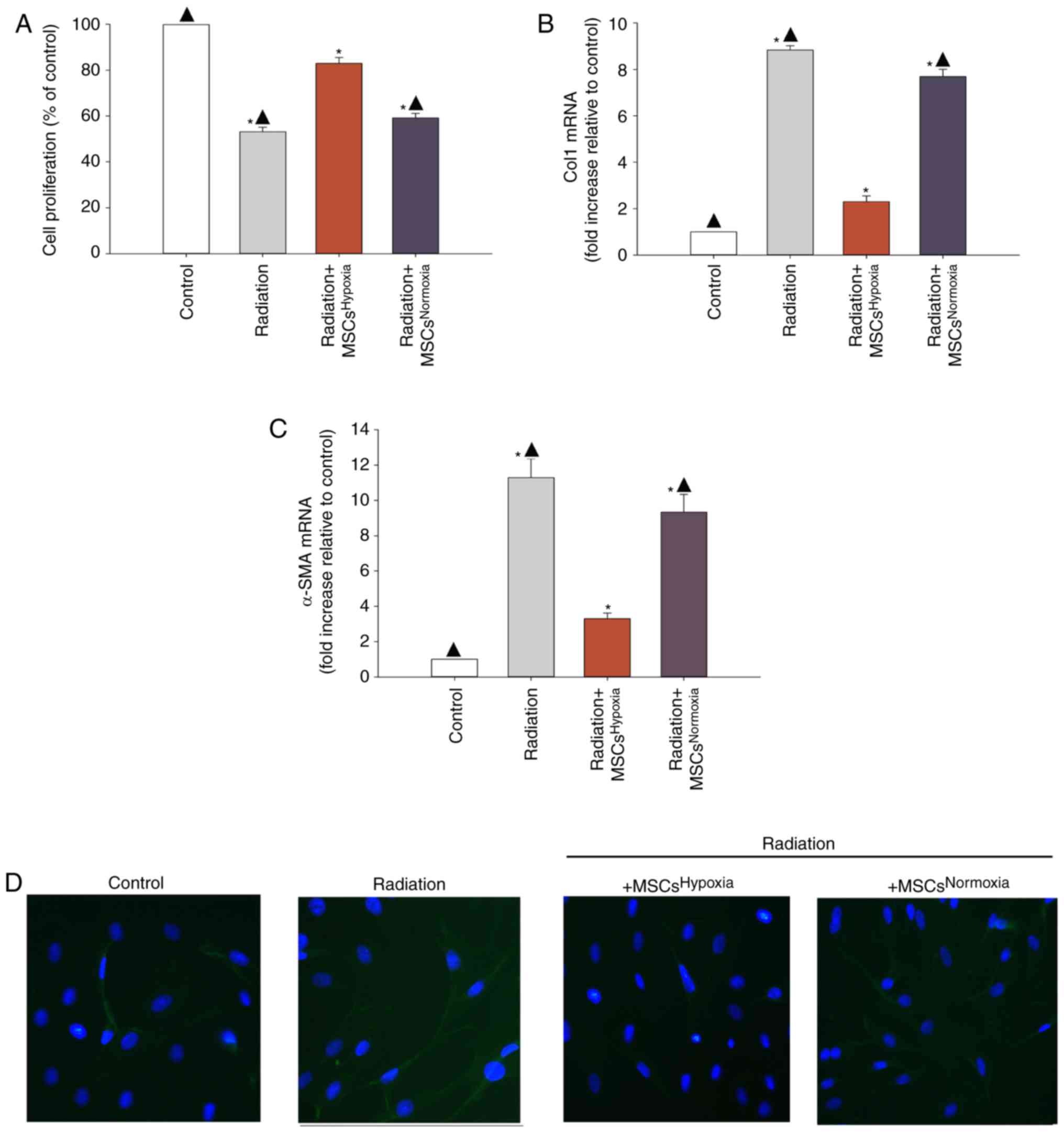
Proliferating fibroblasts and FMT-derived
myofibroblasts are the major source of collagen and other ECM
proteins during pathological matrix remodeling (26). When irradiated fibroblasts were
co-cultured with MSCsHypoxia, fibroblasts exhibited a
lower proliferation rates compared to those that were not exposed
to MSCs. Additionally, normoxia-conditioned MSCs
(MSCsNormoxia) resulted in no inhibition of fibroblast
proliferation, compared with the radiation group (Fig. 1A).
In order to investigate whether
MSCsHypoxia inhibited FMT, the mRNA levels of various
markers of fibrosis including type I collagen (Col1) and α-SMA were
measured. It was revealed that irradiated fibroblasts exhibited a
significant increase in the levels of Col1 and α-SMA mRNA, whereas
co-culturing with MSCsHypoxia reversed this increase
(Fig. 1B and C). In addition,
fluorescence staining revealed increased α-SMA staining in
irradiated fibroblasts compared to those co-cultured with
MSCsHypoxia. No increase in α-SMA expression was
observed in irradiated fibroblasts co-cultured with
MSCsNormoxia (Fig.
1D).
Involvement of HGF in
MSCHypoxia-driven inhibition of irradiation-induced
fibroblast proliferation and FMT
Since MSCs obtain cytoprotection through their
paracrine function, it was explored whether paracrine signaling was
involved in the observed anti-fibrotic effects of
MSCsHypoxia. An ELISA was conducted to examine the
release of HGF, and the results revealed that
MSCsHypoxia released more HGF compared to
MSCsNormoxia (Fig. 2A).
However, the levels of HGF mRNA did not differ between fibroblasts
co-cultured with MSCsHypoxia and those co-cultured with
MSCsNormoxia (Fig.
2B).
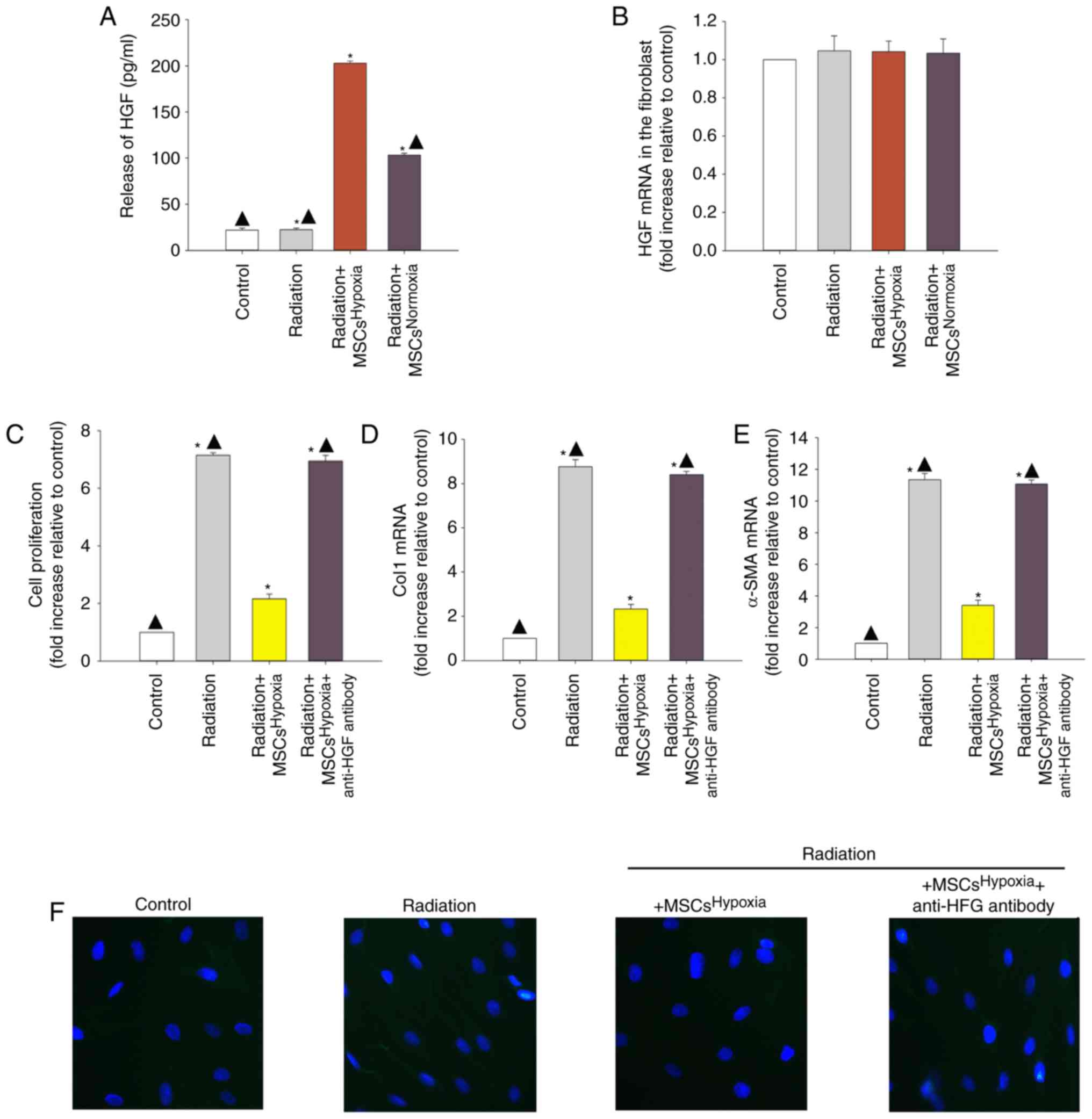 | Figure 2.Involvement of HGF in
MSCsHypoxia-driven inhibition of fibroblast
proliferation and radiation-induced FMT. To explore the role of the
anti-fibrotic effect of MSCsHypoxia on radiation-treated
fibroblast cells, a Transwell co-culture system was used. (A) ELISA
measuring the release of HGF into the condition media of
fibroblasts, fibroblasts treated with radiation, and fibroblasts
co-cultured with MSCsHypoxia or MSCsNormoxia in the
presence of radiation. (B) HGF mRNA levels in fibroblasts,
fibroblasts treated with radiation, and fibroblasts co-cultured
with MSCsHypoxia or MSCsNormoxia in the presence of
radiation were analyzed by RT-qPCR. Each column represents the mean
± SD from three independent experiments; *P<0.05 vs. the
Control; ▲P<0.05 vs. Radiation + MSCsHypoxia. In
fibroblasts, fibroblasts treated with radiation, and fibroblasts
co-cultured with MSCsHypoxia or MSCsHypoxia +
anti-HGF antibody in the presence of radiation, (C) proliferation
growth curves were determined using an MTT assay. (D and E) Col1
and α-SMA mRNA levels as analyzed by RT-qPCR. (F) Expression of
α-SMA was measured using immunofluorescence staining. Each column
represents the mean ± SD from three independent experiments;
*P<0.05 vs. the Control; ▲P<0.05 vs. Radiation +
MSCsHypoxia. HGF, hepatocyte growth factor; MSCs,
mesenchymal stem cells; FMT, fibroblast-to-myofibroblast
transition; Col1, type I collagen; α-SMA, α-smooth muscle
actin. |
To confirm whether MSC-derived HGF inhibited
fibroblast proliferation and radiation-induced FMT, a neutralizing
HGF antibody was used to neutralize the HGF protein. As revealed in
Fig. 2C, radiation induced the
proliferation of fibroblasts, and co-culturing with
MSCsHypoxia decreased their proliferation. The addition
of the anti-HGF antibody diminished the inhibitory effect of
MSCsHypoxia on fibroblast proliferation. Moreover, the
ability of MSCsHypoxia co-culture to reduce the
radiation-induced increase in of Col1 and α-SMA mRNA levels was
also impaired by the addition of the anti-HGF antibody (Fig. 2D and E). Additionally, the anti-HGF
antibody abolished the inhibitory effect of MSCsHypoxia
on α-SMA protein expression in fibroblasts (Fig. 2F).
Identification of TGF-β1 as a
modulator of radiation-induced fibroblast proliferation and
FMT
Microarray analysis was conducted to compare gene
expression between fibroblasts treated or not treated with
radiation (Fig. 3A). Genes with a
≥1.5-fold increase in expression were considered upregulated. It
was revealed that TGF-β1 expression was increased upon exposure to
radiation. Co-culture with MSCsHypoxia decreased the
radiation-induced expression of TGF-β1, while the addition of the
anti-HGF antibody increased the expression of TGF-β1 (Fig. 3B).
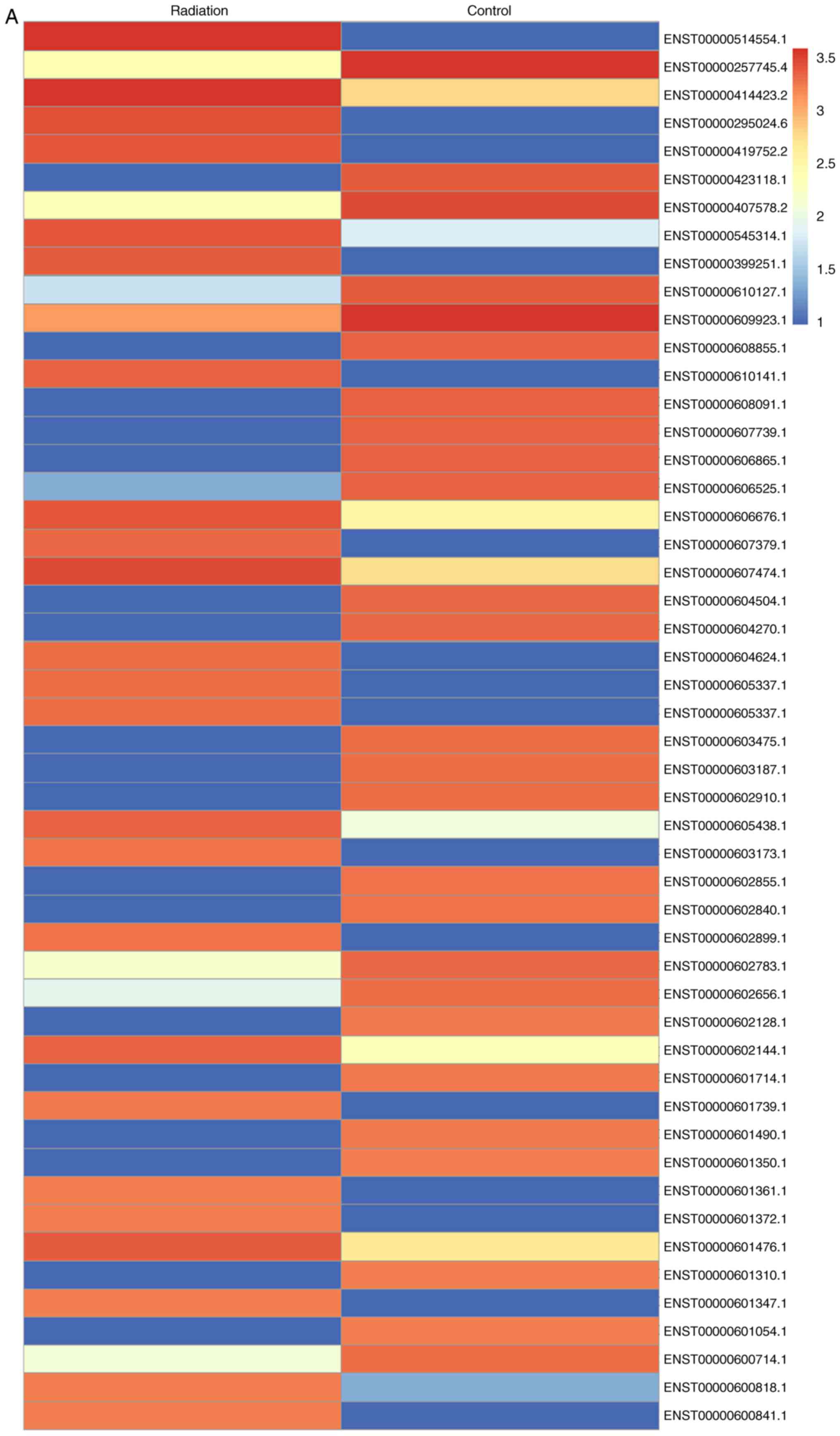 | Figure 3.Identification of TGF-β1 as a
modulator of radiation-induced fibroblast proliferation and FMT.
(A) Heat map of RNAs differentially regulated by radiation in
fibroblasts. ‘Red’ indicates upregulation, and ‘blue’ indicates
downregulation. (B) RT-qPCR validation of differentially regulated
RNAs in fibroblasts, fibroblasts treated with radiation, and
fibroblasts co-cultured with MSCsHypoxia or
MSCsHypoxia + anti-HGF antibody in the presence of
radiation. *P<0.05 vs. the Control;
▲P<0.05 vs. Radiation + MSCsHypoxia. In
fibroblasts, fibroblasts treated with radiation, and fibroblasts
co-cultured with MSCsHypoxia or MSCsHypoxia +
recombinant TGF-β1 in the presence of radiation, (C) proliferation
growth curves were determined using an MTT assay. (D and E) Col1
and α-SMA mRNA levels as analyzed by RT-qPCR. Each column
represents the mean ± SD from three independent experiments;
*P<0.05 vs. the Control; ▲P<0.05 vs.
Radiation + MSCsHypoxia. FMT,
fibroblast-to-myofibroblast transition; MSCs, mesenchymal stem
cells; HGF, hepatocyte growth factor; Col1, type I collagen; α-SMA,
α-smooth muscle actin. |
To investigate the effect of TGF-β1 on
radiation-induced fibroblast proliferation and FMT, recombinant
TGF-β1 was added to fibroblasts co-cultured with
MSCsHypoxia. The addition of recombinant TGF-β1 not only
led to a recovery of the impaired fibroblast proliferation induced
by the MSCsHypoxia (Fig.
3C), but also increased the mRNA levels of Col1 and α-SMA,
which were inhibited by co-culture with MSCsHypoxia
(Fig. 3D and E).
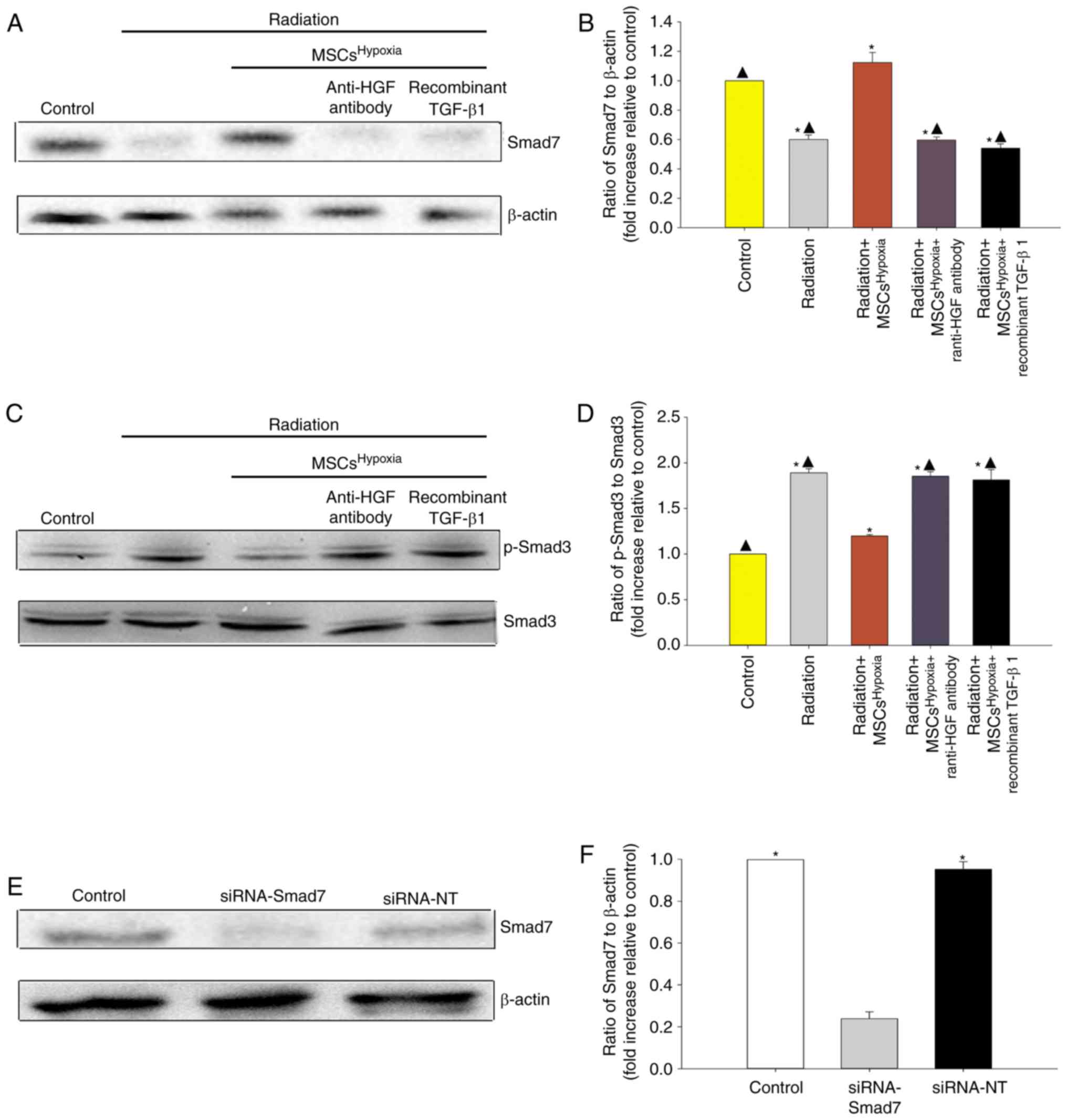
Co-culturing with
MSCsHypoxia functionally targets TGF-β1/Smad signaling
to inhibit fibroblast proliferation and FMT
TGF-β1/Smad signaling is a key pathway involved in
FMT and fibrosis (27). Therefore,
whether co-culturing with MSCsHypoxia inhibited
TGF-β1/Smad signaling to block FMT in fibroblasts, was
investigated. As revealed in Fig.
4A-D, radiation was associated with a decrease in Smad7 levels
and an increase in p-Smad3 levels in fibroblasts. Moreover,
co-culturing with MSCsHypoxia prevented the
radiation-induced decrease in Smad7 levels, and inhibited the
increase in p-Smad3 levels, in fibroblasts. The addition of the
anti-HGF antibody or recombinant TGF-β1 impaired the inhibition of
MSCsHypoxia on TGF-β-induced phospho-Smad3, and impaired
the expression of Smad7.
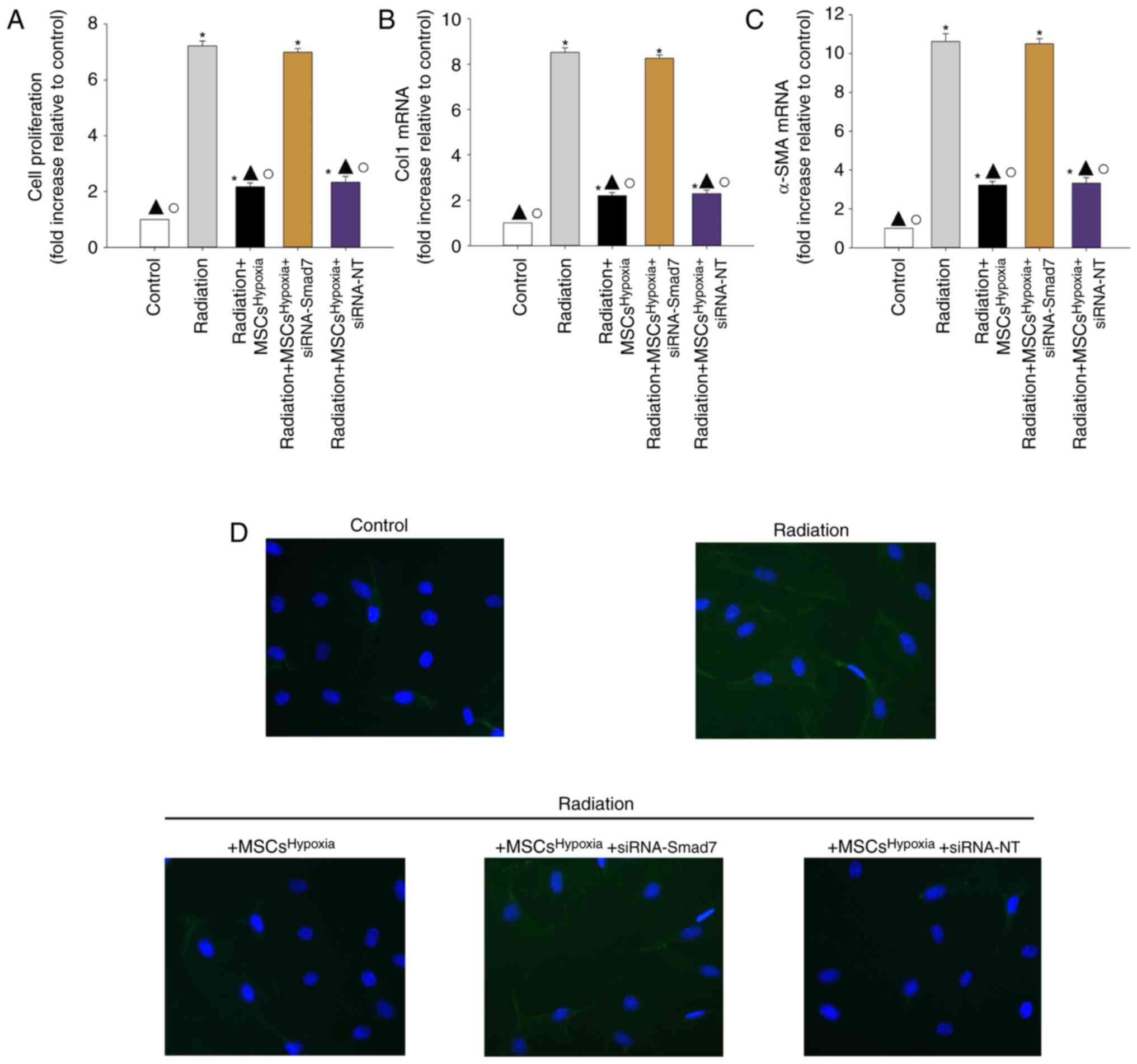
To further confirm the role of TGF-β1/Smad signaling
in FMT, fibroblasts were transfected with siRNA-Smad7 or a
non-targeting RNA as control. As revealed in Fig. 4E and F, fibroblasts that received
siRNA-Smad7 exhibited decreased expression of the Smad7 protein, as
assessed by western blotting. Functionally, silencing of Smad7
reversed the inhibitory effect of MSCsHypoxia on
fibroblast proliferation (Fig. 5A)
and on the mRNA levels of Col1 and α-SMA (Fig. 5B and C). As revealed in Fig. 5D, fibroblasts with silenced Smad7 that
were co-cultured with MSCsHypoxia still exhibited strong
expression of the α-SMA protein, as assessed by immunofluorescence,
indicating that TGF-β1/Smad signaling is a key pathway involved in
FMT and fibrosis.
MSCHypoxia co-culturing
attenuates radiation-induced oxidative stress in fibroblasts
Oxidative stress drives the progression of FMT
(28). A marked increase in the
generation of ROS (Fig. 6A and B),
reduced SOD activity (Fig. 6C), and
increased levels of lipid peroxides (MDA and 4-HNE; Fig. 6D and E) were observed in
radiation-treated fibroblasts. However, these changes were
attenuated when fibroblasts were co-cultured with
MSCsHypoxia. The attenuation of oxidative stress induced
by MSCsHypoxia was significantly suppressed when
fibroblasts were treated with the anti-HGF antibody, recombinant
TGF-β1, or siRNA-Smad7.
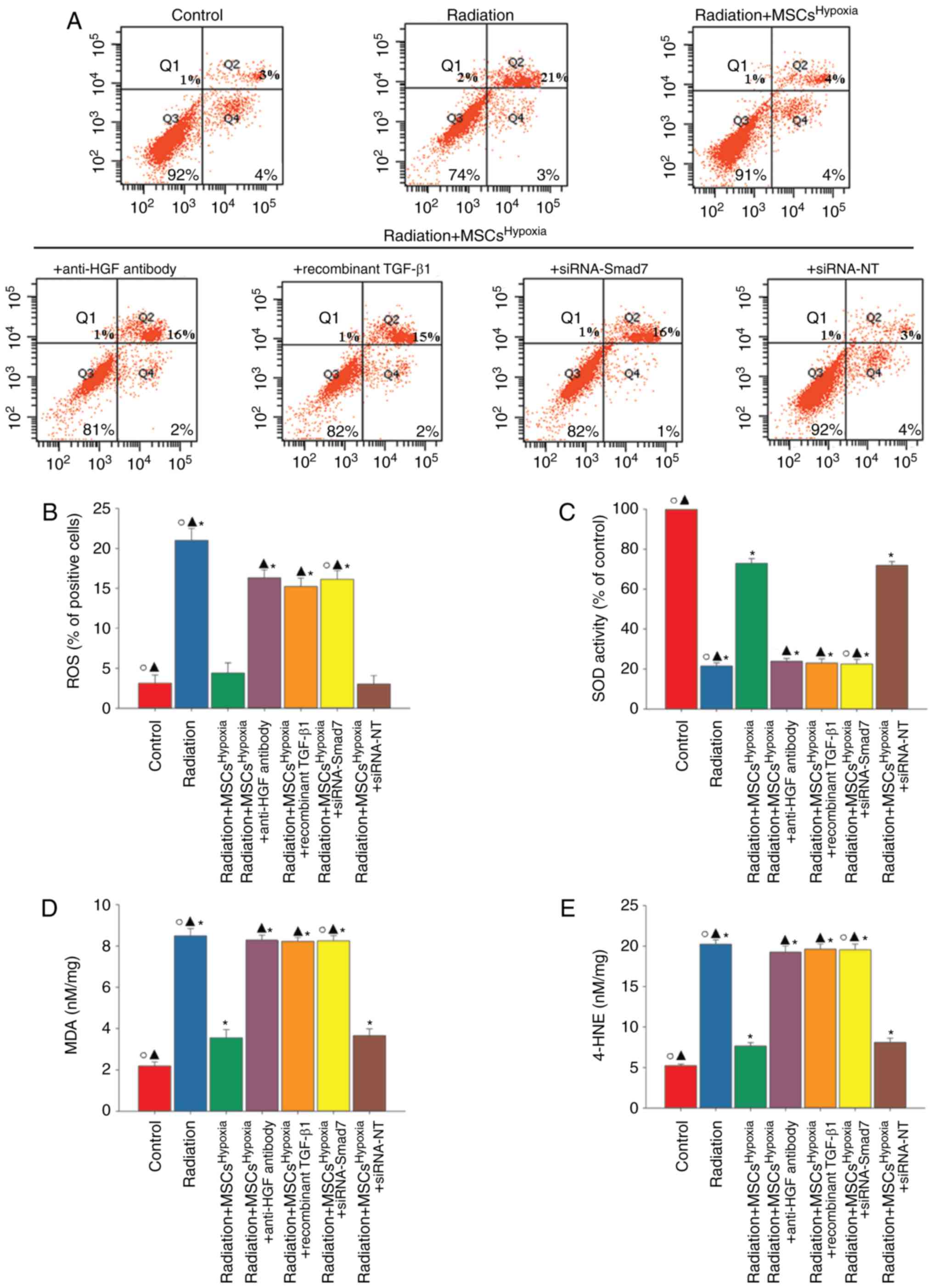 | Figure 6.MSCHypoxia co-culturing
attenuates radiation-induced oxidative stress in fibroblasts.
Fibroblasts were transfected with siRNA against Smad7, or with
siRNA-NT as a control, followed by co-culture with
MSCsHypoxia in the presence of radiation. In parallel
experiments, fibroblasts were treated with radiation, or
co-cultured with MSCsHypoxia or MSCsHypoxia +
anti-HGF antibody or MSCsHypoxia + recombinant TGF-β1 in
the presence of radiation. Fibroblasts under normal culture
conditions were used as the control. (A and B) Intracellular ROS
production was assessed using a ROS detection kit, and was analyzed
using flow cytometry. (C) SOD activity evaluated by a colorimetric
assay. (D) Lipid peroxidation as evaluated by MDA formation. (E)
Quantification of 4-HNE levels. *P<0.05 vs. the
Control; ▲P<0.05 vs. Radiation +
MSCsHypoxia; ○P<0.05 vs. Radiation +
MSCsHypoxia + siRNA-Smad7. MSCs, mesenchymal stem cells;
HGF, hepatocyte growth factor; ROS, reactive oxygen species; SOD,
superoxide dismutase; MDA, malondialdehyde; HNE,
4-hydroxynonenal. |
Discussion
Radiation therapy, used during treatment for various
types of cancer, is associated with an increased and dose-dependent
risk of delayed-onset heart disease (29). The earliest data associating radiation
and heart disease originated from studies of long-term patient
outcomes after radiotherapy treatment for malignant cancers
(3,30). In these types of treatments, the local
radiation doses to some regions of the heart can exceed 40 Gy
(31), which results in cardiac
damage and different pathologies, primarily fibrosis of the
pericardium and myocardium (32).
Radiation-related myocardial fibrosis is often asymptomatic and
diagnosed relatively late (33). It
is characterized by a proliferation of fibroblasts and FMT, which
involves the activation of fibroblasts and a transition to
α-SMA-expressing myofibroblasts, which produce an excessive amount
of ECM, leading to tissue fibrosis (34). The results of the present study
revealed that radiation-induced fibroblasts proliferate and undergo
FMT.
MSCs exhibited great potential in treating cardiac
damages, even without any pre-treatment. In the 2009, Hare et
al provided pivotal safety and provisional efficacy data for an
allogeneic bone marrow-derived stem cell without any treatment
within post-infarction patients (35). It has been theorized that MSCs reduce
cardiac fibrosis largely through paracrine signaling mechanisms
(36,37), such as via the inhibition of the
TGF-β-induced transformation of fibroblast cells into
myofibroblasts, which protects against cardiac damage (38). Hypoxia pre-conditioned MSCs have been
revealed to have an increase in paracrine signaling (39), which increases protection against
irradiation-induced damage (40). In
the present study, it was revealed that co-culturing with
MSCsHypoxia inhibited the proliferation of fibroblasts
and inhibited their radiation-induced transformation into
myofibroblasts via paracrine signaling, in an HGF-dependent manner.
It was also revealed that the addition of anti-HGF antibody
diminished the anti-fibrotic effect of MSCs Hypoxia,
confirming the important role of paracrine HGF signaling in
limiting radiation-related fibrosis.
Enhanced levels of TGF-β1 are involved in
radiation-induced cardiac fibrosis, a condition characterized by
excess fibroblast proliferation and a deposition of collagen fibers
(41). In line with these
observations, the present study revealed a significant increase in
TGF-β1 expression induced by radiation, while
MSCsHypoxia inhibited the expression of TGF-β1. The
activation of TGF-β has been predicted using both transcriptomics
and proteomic data sets (15). TGF-β
signaling is able to initiate the canonical SMAD transduction
pathway, leading to fibrosis (42).
Smad7, an inhibitory Smad, inhibits the activation of Smad2 and
Smad3 in response to TGF-β activation. Herein, it was demonstrated
that co-culturing with MSCsHypoxia induced the
expression of Smad7, which was inhibited by radiation, while
impairing the radiation-induced phosphorylation of Smad3,
indicating that the TGF-β/Smad pathway acts to modulate MSC
inhibition of radiation-induced fibrosis.
Previous studies have proposed many complex
molecular events which lead to the development of radiation-induced
cardiac damage (43). Among all these
events, radiation-induced oxidative stress and the associated
inflammatory responses are likely the key signaling cascades
leading to cardiac damage (44,45).
Radiation leads to a significant reduction in the levels of
antioxidant enzymes such as SOD, and an increase in 4-HNE adducts,
which leads to cardiac fibrosis (46). Radiation-induced ROS generation
activates MDA and promotes 4-HNE generation, which is accompanied
by an inhibition of SOD. The present results revealed that
MSCHypoxia treatment was able to attenuate oxidative
stress. In addition, treatment with anti-HGF antibody, recombinant
TGF-β1 or siRNA-Smad7 abolished the antioxidant effect of
MSCsHypoxia. While there were also some limitations, in
the following research, in future, whether hypoxia precondition
improves the treatment effect of MSCs in radiation-induced cardiac
fibrosis in vivo will be investigated.
In conclusion, in the present study, it was revealed
that radiation-induced fibroblast proliferation and FMT were
attenuated by co-culturing with MSCsHypoxia. In
addition, the results revealed that MSCsHypoxia exert
anti-fibrotic and radio-protective effects by regulating the
TGF-β/Smad signaling pathway, and inhibiting oxidative stress via
paracrine pathways. The present study provides evidence that MSCs
may be a promising candidate for the treatment of radiation-related
cardiac fibrosis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81600278 to WX and
81500261 to MH) and the Medical Science and Technology Project of
Zhejiang Province (grant no. 2018KY517 to MH).
Availability of data and materials
The data used to support the findings of this study
are included within the article.
Authors' contributions
LZ and WX made substantial contributions to the
acquisition of data, analysis and interpretation of data. MH was
involved in conception and design of the study, and drafting the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Swerdlow AJ, Higgins CD, Smith P,
Cunningham D, Hancock BW, Horwich A, Hoskin PJ, Lister A, Radford
JA, Rohatiner AZ and Linch DC: Myocardial infarction mortality risk
after treatment for Hodgkin disease: A collaborative British cohort
study. J Natl Cancer Inst. 99:206–214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Darby SC, Ewertz M, McGale P, Bennet AM,
Blom-Goldman U, Brønnum D, Correa C, Cutter D, Gagliardi G, Gigante
B, et al: Risk of ischemic heart disease in women after
radiotherapy for breast cancer. N Engl J Med. 368:987–998. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Darby S, McGale P, Peto R, Granath F, Hall
P and Ekbom A: Mortality from cardiovascular disease more than 10
years after radiotherapy for breast cancer: Nationwide cohort study
of 90000 Swedish women. BMJ. 326:256–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tukenova M, Guibout C, Oberlin O, Doyon F,
Mousannif A, Haddy N, Guérin S, Pacquement H, Aouba A, Hawkins M,
et al: Role of cancer treatment in long-term overall and
cardiovascular mortality after childhood cancer. J Clin Oncol.
28:1308–1315. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Tapio S: Pathology and biology of
radiation-induced cardiac disease. J Radiat Res. 57:439–448. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Subramanian V, Borchard S, Azimzadeh O,
Sievert W, Merl-Pham J, Mancuso M, Pasquali E, Multhoff G, Popper
B, Zischka H, et al: PPARα is necessary for radiation-induced
activation of noncanonical TGFβ signaling in the heart. J Proteome
Res. 17:1677–1689. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Curigliano G, Cardinale D, Dent S,
Criscitiello C, Aseyev O, Lenihan D and Cipolla CM: Cardiotoxicity
of anticancer treatments: Epidemiology, detection, and management.
CA Cancer J Clin. 66:309–325. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bagno L, Hatzistergos KE, Balkan W and
Hare JM: Mesenchymal stem cell-based therapy for cardiovascular
disease: Progress and challenges. Mol Ther. 26:1610–1623. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Golpanian S, Wolf A, Hatzistergos KE and
Hare JM: Rebuilding the damaged heart: Mesenchymal stem cells,
cell-based therapy, and engineered heart tissue. Physiol Rev.
96:1127–1168. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma S, Xie N, Li W, Yuan B, Shi Y and Wang
Y: Immunobiology of mesenchymal stem cells. Cell Death Differ.
21:216–225. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Le Blanc K and Mougiakakos D: Multipotent
mesenchymal stromal cells and the innate immune system. Nat Rev
Immunol. 12:383–396. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wu T, Xie Y, Huang J, Li P, Wang X, Yan Y,
Xia T, Li L, Zhu F, Li H and Wu R: The optimal intervention time of
bone marrow mesenchymal stem cells in ameliorating cardiac fibrosis
induced by viral myocarditis: A randomized controlled trial in
mice. Stem Cells Int. 2017:32580352017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang L, Ma W, Ma Y, Feng D, Chen H and
Cai B: Exosomes in mesenchymal stem cells, a new therapeutic
strategy for cardiovascular diseases? Int J Biol Sci. 11:238–245.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Meng SS, Xu XP, Chang W, Lu ZH, Huang LL,
Xu JY, Liu L, Qiu HB, Yang Y and Guo FM: LincRNA-p21 promotes
mesenchymal stem cell migration capacity and survival through
hypoxic preconditioning. Stem Cell Res Ther. 9:2802018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Dobaczewski M, Chen W and Frangogiannis
NG: Transforming growth factor (TGF)-β signaling in cardiac
remodeling. J Mol Cell Cardiol. 51:600–606. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yamamura Y, Hua X, Bergelson S and Lodish
HF: Critical role of Smads and AP-1 complex in transforming growth
factor-beta-dependent apoptosis. J Biol Chem. 275:36295–36302.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hayashi H, Abdollah S, Qiu Y, Cai J, Xu
YY, Grinnell BW, Richardson MA, Topper JN, Gimbrone MA Jr, Wrana JL
and Falb D: The MAD-related protein Smad7 associates with the TGFβ
receptor and functions as an antagonist of TGFbeta signaling. Cell.
89:1165–1173. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wei LH, Huang XR, Zhang Y, Li YQ, Chen HY,
Yan BP, Yu CM and Lan HY: Smad7 inhibits angiotensin II-induced
hypertensive cardiac remodelling. Cardiovasc Res. 99:665–673. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Berk BC, Fujiwara K and Lehoux S: ECM
remodeling in hypertensive heart disease. J Clin Invest.
117:568–575. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Fredj S, Bescond J, Louault C and Potreau
D: Interactions between cardiac cells enhance cardiomyocyte
hypertrophy and increase fibroblast proliferation. J Cell Physiol.
202:891–899. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lucas JA, Zhang Y, Li P, Gong K, Miller
AP, Hassan E, Hage F, Xing D, Wells B, Oparil S and Chen YF:
Inhibition of transforming growth factor-beta signaling induces
left ventricular dilation and dysfunction in the
pressure-overloaded heart. Am J Physiol Heart Circ Physiol.
298:H424–H432. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cheema AK, Pathak R, Zandkarimi F, Kaur P,
Alkhalil L, Singh R, Zhong X, Ghosh S, Aykin-Burns N and
Hauer-Jensen M: Liver metabolomics reveals increased oxidative
stress and fibrogenic potential in gfrp transgenic mice in response
to ionizing radiation. J Proteome Res. 13:3065–3074. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yin TC, Wu RW, Sheu JJ, Sung PH, Chen KH,
Chiang JY, Hsueh SK, Chung WJ, Lin PY, Hsu SL, et al: Combined
therapy with extracorporeal shock wave and adipose-derived
mesenchymal stem cells remarkably improved acute
ischemia-reperfusion injury of quadriceps muscle. Oxid Med Cell
Longev. 2018:60126362018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Moon J, Schwarz SC, Lee HS, Kang JM, Lee
YE, Kim B, Sung MY, Höglinger G, Wegner F, Kim JS, et al:
Preclinical analysis of fetal human mesencephalic neural progenitor
cell lines: Characterization and safety in vitro and in vivo. Stem
Cells Transl Med. 6:576–588. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Teekakirikul P, Eminaga S, Toka O, Alcalai
R, Wang L, Wakimoto H, Nayor M, Konno T, Gorham JM, Wolf CM, et al:
Cardiac fibrosis in mice with hypertrophic cardiomyopathy is
mediated by non-myocyte proliferation and requires Tgf-β. J Clin
Invest. 120:3520–3529. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang Q, Ye H, Xiang F, Song LJ, Zhou LL,
Cai PC, Zhang JC, Yu F, Shi HZ, Su Y, et al: miR-18a-5p inhibits
sub-pleural pulmonary fibrosis by targeting TGF-β receptor II. Mol
Ther. 25:728–738. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ruart M, Chavarria L, Campreciós G,
Suárez-Herrera N, Montironi C, Guixé-Muntet S, Bosch J, Friedman
SL, Garcia-Pagán JC and Hernández-Gea V: Impaired endothelial
autophagy promotes liver fibrosis by aggravating the oxidative
stress response during acute liver injury. J Hepatol. 70:458–469.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Serrano NA, Mikkelsen R, Canada J,
Mezzaroma E, Weiss E and Abbate A: Biomarkers of cardiac injury in
patients undergoing thoracic radiation therapy. Int J Cardiol.
223:507–509. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hancock SL, Donaldson SS and Hoppe RT:
Cardiac disease following treatment of Hodgkin's disease in
children and adolescents. J Clin Oncol. 11:1208–1215. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
McGale P and Darby SC: Low doses of
ionizing radiation and circulatory diseases: A systematic review of
the published epidemiological evidence. Radiat Res. 163:247–257.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Demirci S, Nam J, Hubbs JL, Nguyen T and
Marks LB: Radiation-induced cardiac toxicity after therapy for
breast cancer: Interaction between treatment era and follow-up
duration. Int J Radiat Oncol Biol Phys. 73:980–987. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Heidenreich PA, Hancock SL, Lee BK,
Mariscal CS and Schnittger I: Asymptomatic cardiac disease
following mediastinal irradiation. J Am Coll Cardiol. 42:743–749.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Gabbiani G: The myofibroblast in wound
healing and fibrocontractive diseases. J Pathol. 200:500–503. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hare JM, Traverse JH, Henry TD, Dib N,
Strumpf RK, Schulman SP, Gerstenblith G, DeMaria AN, Denktas AE,
Gammon RS, et al: A randomized, double-blind, placebo-controlled,
dose-escalation study of intravenous adult human mesenchymal stem
cells (prochymal) after acute myocardial infarction. J Am Coll
Cardiol. 54:2277–2286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Ranganath S, Levy O, Inamdar M and Karp
JM: Harnessing the mesenchymal stem cell secretome for the
treatment of cardiovascular disease. Cell Stem Cell. 10:244–258.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Molina EJ, Palma J, Gupta D, Torres D,
Gaughan JP, Houser S and Macha M: Reverse remodeling is associated
with changes in extracellular matrix proteases and tissue
inhibitors after mesenchymal stem cell (MSC) treatment of pressure
overload hypertrophy. J Tissue Eng Regen Med. 3:85–91. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shao L, Zhang Y, Lan B, Wang J, Zhang Z,
Zhang L, Xiao P, Meng Q, Geng YJ, Yu XY and Li Y: MiRNA-sequence
indicates that mesenchymal stem cells and exosomes have similar
mechanism to enhance cardiac repair. Biomed Res Int.
2017:41507052017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hu X, Xu Y, Zhong Z, Wu Y, Zhao J, Wang Y,
Cheng H, Kong M, Zhang F, Chen Q, et al: A large-scale
investigation of hypoxia-preconditioned allogeneic mesenchymal stem
cells for myocardial repair in nonhuman primates: Paracrine
activity without remuscularization. Circ Res. 118:970–983. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Shin HS, Lee S, Kim YM and Lim JY:
Hypoxia-activated adipose mesenchymal stem cells prevents
irradiation-induced salivary hypofunction by enhanced paracrine
effect through fibroblast growth factor 10. Stem Cells.
36:1020–1032. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Seemann I, Gabriels K, Visser NL, Hoving
S, te Poele JA, Pol JF, Gijbels MJ, Janssen BJ, van Leeuwen FW,
Daemen MJ, et al: Irradiation induced modest changes in murine
cardiac function despite progressive structural damage to the
myocardium and microvasculature. Radiother Oncol. 103:143–150.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Taylor C, Mcgale P, Brønnum D, Correa C,
Cutter D, Duane FK, Gigante B, Jensen MB, Lorenzen E, Rahimi K, et
al: Cardiac structure injury after radiotherapy for breast cancer:
Cross-sectional study with individual patient data. J Clin Oncol.
36:2288–2296. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Hirshfeld JW Jr, Fiorilli PN and Silvestry
FE: Important strategies to reduce occupational radiation exposure
in the cardiac catheterization laboratory: No lower limit. J Am
Coll Cardiol. 71:1255–1258. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Viczenczova C, Kura B, Egan Benova T, Yin
C, Kukreja RC, Slezak J, Tribulova N and Szeiffova Bacova B:
Irradiation-induced cardiac connexin-43 and miR-21 responses are
hampered by treatment with atorvastatin and aspirin. Int J Mol Sci.
19:2018. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Seawright JW, Samman Y, Sridharan V, Mao
XW, Cao M, Singh P, Melnyk S, Koturbash I, Nelson GA, Hauer-Jensen
M and Boerma M: Effects of low-dose rate γ-irradiation combined
with simulated microgravity on markers of oxidative stress, DNA
methylation potential, and remodeling in the mouse heart. PLoS One.
12:e01805942017. View Article : Google Scholar : PubMed/NCBI
|