Introduction
Esophageal squamous cell carcinoma (ESCC) is a
malignancy that most commonly presents with progressive dysphagia.
It has been reported to be the fourth leading cause of
cancer-related mortality and the third most common type of cancer
in China, accounting for over half of all esophageal cancer cases
worldwide (1). When diagnosed at an
early stage, when dysplasia is limited to the mucosal layer and
there is no lymph node metastasis, ESCC may be curable by
endoscopic resection (2); however,
since the majority of patients with early-stage disease are
asymptomatic, ESCC is often diagnosed at an advanced stage, when it
is most likely metastatic, and the overall 5-year survival rate
ranges from 15 to 25% (3,4). Therefore, identification of the
mechanism underlying esophageal cancer metastasis is crucial for
treatment selection.
The phosphoinositide 3-kinase (PI3K)/AKT/mammalian
target of rapamycin (mTOR) signaling pathway is one of the most
frequently activated pathways in human cancers, including ESCC
(5–9).
In mammals, PI3Ks may be classified into class I, II and III,
according to their structure and substrate specificity. Class I
PI3Ks consist of a regulatory subunit (p85, p101 or p84) and a
catalytic subunit (p110α, p110β, p110γ or p110δ). GTP-Ras interacts
with p85 to induce a p110 conformational change to phosphorylate
PI3K. PIP3, the product of class I PI3K, recruits the
serine-threonine kinase AKT, which contains a pleckstrin homology
(PH) domain, and its activating kinase 3-phosphoinositide-dependent
kinase 1 (PDK1) to cell membranes, allowing PDK1 to activate AKT
(10). AKT signals regulate multiple
oncogenic processes, including cell proliferation, differentiation,
apoptosis, epithelial-to-mesenchymal transition (EMT), migration
and invasion (11).
Ras-associated protein 1A (Rap1A) is a type of small
GTP-binding protein that belongs to the Ras subfamily. Similar to
other Ras proteins, Rap1A switches between an inactive GDP-bound
and an active GTP-bound form. Although Ras and Rap1A share almost
the same effectors near the cell surface, Rap1A is activated in the
perinuclear region, while the majority of Ras proteins are
activated at the plasma membrane (12,13).
Previous research has demonstrated that Rap1A plays an essential
regulatory role in a variety of cellular processes, including
proliferation, integrin activation, cell adhesion and migration.
However, the specific function of Rap1A varies by cell type; for
example, Rap1A mediates integrin-dependent glioblastoma cell
proliferation and tumor growth (14).
Rap1A also plays an important role in prostate cancer cell
migration and invasion (15,16) and promotes ovarian cancer cell
proliferation, migration and invasion (17). These results suggest that Rap1A may
act as an oncogene; however, the specific role and mechanism of
action of Rap1A in ESCC remains unknown. In the present study,
Rap1A was silenced in ECA109 and KYSE150 ESCC cells and its
functions were examined in vivo and in vitro. The
underlying molecular mechanism was also investigated.
Materials and methods
Cell lines and culture conditions
The human esophageal squamous cancer cell lines
ECA109 and KYSE150 were purchased from Genechem Co., Ltd. All cells
were maintained at 37°C with 5% CO2. ECA109 cells were
grown in DMEM (SH30243.01; HyClone; GE Healthcare Life Sciences)
and KYSE150 cells were maintained in RPMI-1640 medium (SH30809.01;
HyClone; GE Healthcare Life Sciences), both of which were
supplemented with 10% fetal bovine serum (FBS; 10270106; Gibco;
Thermo Fisher Scientific, Inc.) and penicillin-streptomycin
solution.
Clinical samples and
immunohistochemistry (IHC)
A total of 170 paraffin-embedded ESCC specimens and
adjacent non-tumor tissues (ANTT) acquired between 2006 and 2008
were obtained from the Fudan University-Affiliated Zhongshan
Hospital. The inclusion criteria were as follows: i) No
radiotherapy, chemotherapy or targeted therapy prior to surgery;
ii) the results of pathological examination were in accordance with
the clinical diagnosis; iii) complete clinicopathological data were
available. The TNM classification was performed according to the
eighth edition staging system published by the Journal of Thoracic
Oncology (18). The details of 95
patients who provided ESCC samples are listed in supplementary
Tables SI and SII. Rap1A antibody (rabbit polyclonal,
1:100 dilution; ab197673; Abcam) for IHC detection. Scoring was
determined by two independent pathologists by combining the
proportion of positively stained tumor or epithelial cells and the
intensity of staining. The positively stained cell proportions were
scored as follows: 0, <1% positive cells; 1, 1–25% positive
cells; 2, 25–50% positive cells; 3, 50–75% positive cells; and 4,
>75% positive cells. Staining intensity was graded as follows:
0, no staining; 1, weak staining (light yellow); 2, moderate
staining (yellowish brown); 3, strong staining (brown). The
staining index (SI) was calculated as the product of the staining
intensity score and the proportion of positive cells. Samples with
possible SI scores of 0, 1–4, 5–8 or 9–12 were considered to have
negative, low, intermediate and strong expression,
respectively.
In addition, IHC staining for Rap1A was performed on
mouse lung sections to verify whether pulmonary nodules were formed
by metastasis of ECA109 cells injected into the nude mice through
the tail vein. The detailed procedures of IHC staining were as
follows: Lung sections (4-µm) were deparaffinized in xylene and
rehydrated through an alcohol gradient; endogenous peroxidase was
inactivated by hydrogen peroxide; antigen was retrieved by
ethylenediaminetetraacetic acid (EDTA); tissue was blocked by goat
serum for 30 min, incubated with Rap1A antibody overnight at 4°C
and finally incubated with secondary antibody for 20 min at room
temperature; IHC reaction was visualized by DAB solution and the
cell nuclei were stained by hematoxylin; the sections were
dehydrated, transparentized and sealed for further examination or
archiving.
Plasmid and lentivirus construction
and cell transfection
To knock down the expression of Rap1A in ECA109 and
KYSE150 cells, four shRNAs targeting human Rap1A or SP1 were
designed, synthesized and inserted into the construct
U6-Rap1A-shRNA1/2/3/4-Ubi-Cherry-vector by Genechem. Cells were
transfected with the plasmids described above using Lipofectamine
2000 (11668–019, Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. For each Rap1A shRNA
candidate, mRNA and protein expression was analyzed and the siRNAs
targeting the sequences AAAGTCAAAGATCAATGTT and CAGCATTCCAGACTTCAAA
were determined as the two most effective for inhibiting Rap1A
expression. Lentiviruses harboring the corresponding shRNAs were
generated by transfection of 293T cells with a three-plasmid-based
lentiviral packaging system. Stable ECA109 and KYSE150 cell lines
with Rap1A silencing were generated via lentivirus infection and
used for subsequent experiments. Moreover, the SP1 coding sequence
was subcloned into the GV492 vector (Genechem) to construct a
cDNA-SP1 plasmid and investigate its role in Rap1A expression.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total cellular RNA was extracted using TRIzol
reagent (15596-026; Ambion; Thermo Fisher Scientific, Inc.) and was
reverse-transcribed to cDNA using the PrimeScript™ RT Master Mix
(RR036A; Takara) according to the manufacturer's instructions.
Subsequently, two-step RT-qPCR using TB Green (RR420A, Takara) was
performed to determine mRNA levels. PCR was run using the following
protocol: Initial activation at 95°C for 30 sec, followed by 40
cycles at 95°C for 5 sec and at 60°C for 30 sec. The PCR primers
used were as follows: Rap1A forward, 5′-TGTCTCACTGCACCTTCA-3′ and
reverse, 5′-GACTTCCCAACGCCTCCT-3′; SP1 forward,
5′-TGGCAGCAGTACCAATGGC-3′ and reverse,
5′-CCAGGTAGTCCTGTCAGAACTT-3′. All reactions were performed in
triplicate and the mRNA expression was analyzed using the
2−ΔΔCq method (16).
Growth curves and colony formation
assay
For the growth curves, cells were detached from
flasks using 0.25% Trypsin-EDTA and centrifuged at 80 × g, room
temperature for 5 min, counted, seeded at 3,000 cells per well (in
quintuplicate) in 96-well culture plates in 100 µl of
recommended medium and cultured for 1, 2, 3, 4, 5 and 6 days. Cell
growth was detected using MTT solution and DMSO, according to the
manufacturer's instructions (ST316; Beyotime Institute of
Biotechnology). The OD values at 490 nm were recorded and analyzed
to compare the proliferation among cells. For the colony formation
assay, cells were seeded (300 cells/well in triplicate) in 6-well
plates and cultured for 2–3 weeks. Fresh culture medium was
replaced every 3 days. When the colonies were detectable
macroscopically, the cells were fixed and stained using 0.1%
crystal violet solution (C0121; Beyotime Institute of
Biotechnology). The colony-forming ability was evaluated using the
colony number.
Transwell migration and invasion
assay
To evaluate cell migration and invasion ability,
24-well Transwell insert chambers with 8.0-µm polycarbonate
membranes (3422; Corning, Inc.) were used. For migration assays,
5×104 cells were plated into the upper chamber in
serum-free medium, and medium supplemented with 10% FBS was added
to the lower chamber. Following incubation for 24 h, the cells were
fixed in ethanol for 0.1% crystal violet staining (C0121; Beyotime
Institute of Biotechnology). The cells in the upper chamber were
then removed using a cotton swab. Cell migration ability was
analyzed by counting the number of stained cells under a light
microscope. For invasion assays, after starvation for 24 h,
1×105 cells were seeded in serum-free medium into the
upper chamber that was precoated with Matrigel (354480; Corning,
Inc.), and medium supplemented with 10% FBS was added to the lower
chamber to serve as a chemoattractant. Following incubation for 48
h, the cells were fixed, stained (non-invading cells were removed
with a cotton swab), and analyzed as described above.
Western blotting
The cells were lysed using RIPA lysis buffer
(P0013B; Beyotime Institute of Biotechnology), containing a
protease and phosphatase inhibitor cocktail. Lysate protein
concentrations were determined using a BCA kit (P0010S; Beyotime
Institute of Biotechnology). The denatured protein samples were
separated by 10% SDS-PAGE in electrophoresis buffer and transferred
to a PVDF membrane. The membranes were blocked in 5% milk for 2 h,
then incubated with primary antibodies overnight at 4°C. The
antibodies were directed against Rap1A (rabbit polyclonal,
ab197673, 1:1,000; Abcam), β1 integrin (rabbit polyclonal,
ab183666, 1:3,000; Abcam), matrix metalloproteinase (MMP)9 (rabbit
polyclonal, 10375-2-AP, 1:1,000; ProteinTech Group, Inc.),
E-cadherin (rabbit monoclonal, 24E10, 1:1,000; Cell Signaling
Technology, Inc.), Slug (rabbit polyclonal, arg55242, 1:1,000;
Abcam), AKT (rabbit polyclonal, arg56418, 1:600; Arigo), SP1
(rabbit polyclonal, ab13370, 1:3,000) and pAKTser473 (rabbit
monoclonal, 4060, 1:2,000; Cell Signaling Technology, Inc.). The
membranes were incubated with HRP-conjugated secondary antibodies
for 2 h, and then antigen-antibody complexes were detected using an
electrochemiluminescence kit to compare the corresponding protein
expression levels among different samples.
Chromatin immunoprecipitation (ChIP)
assay
All viable cells (>2×106) were treated
with 1% formaldehyde to cross-link proteins to DNA. The DNA
compounds were digested by a micrococcal nuclease (26156; Thermo
Fisher Scientific, Inc.) into 100-1,000-bp segments. Equal aliquots
of chromatin supernatants were incubated with anti-IgG antibody,
anti-SP1 antibody (ab13370), or anti-RNA polymerase II antibody
overnight at 4°C with rotation. Antibody-protein-DNA complexes were
precipitated by protein A agarose and reverse-cross-linked to free
the DNA. PCR and agarose gel electrophoresis were used to verify
the protein-binding site on the DNA. The primers used were as
follows: F1 forward, 5′-TTTCTGGCTGGGGTAAACCG-3′ and reverse,
5′-GCTGGCACACTTTGTCTTGGAGC-3′; F2 forward, 5′-GGCTCAGGAGTCATAGCT-3′
and reverse, 5′-GCATGTCCAAATTAACT-3′.
Luciferase reporter assay
Fragments containing the Rap1A promoter region or
mutants of the predictive SP1-binding site were inserted upstream
of the firefly luciferase coding sequences in the pGL3-basic
reporter plasmid. ECA109 and KYSE150 cells were seeded onto 96-well
plates (in triplicate), and were then transiently transfected with
pGL3-ctrl, pGL3-Rap1A promoter, pGL3-Rap1A mut1, or pGL3-Rap1A mut2
and pTK-Renilla, with or without cDNA-SP1 plasmid. Fluc and
Rluc activities were determined using the Dual-Glo Luciferase Assay
System (E2920; Promega Corporation).
Tumor growth and metastasis in
vivo
Female athymic nude mice (4-week-old, 16–18 g, n=12;
and 5-week-old, 19–21 g, n=12) were purchased from the Shanghai
Laboratory Animal Center, Chinese Academy of Sciences. The mice
were housed under specific pathogen-free conditions at the Fudan
University Laboratory Animal Division. Briefly, for xenograft
tumors, 5×106 cells (ECA109-shRap1A-NC or
ECA109-shRap1A-4) were injected into the right dorsal flank of nude
mice (n=6 per group). The length and width of the tumors were
measured twice weekly using calipers and the tumor volume was
calculated using the equation (length × width2)/2. When
the length of any tumor reached 20 mm or greater, all animals were
sacrificed and the tumors were excised. For lung metastasis,
1×106 cells were injected into the tail vein of nude
mice (n=6 per group in two groups). The animals were sacrificed by
cervical dislocation and the lungs were excised after 6 weeks or
when the animals developed significant ascites or
cachexia/emaciation, hind-limb paralysis or weakness. The nodules
formed in the lungs were counted, and the lungs were then fixed in
formalin and embedded in paraffin for further H&E and IHC
staining.
The details of IHC staining were mentioned above,
and H&E staining was performed as described below. After
hydration, the sections were stained with hematoxylin for 10 min.
The cytoplasm was visualized by incubating with eosin for ~2 min.
Finally, the sections were dehydrated, transparentized and sealed
in the same manner as for IHC staining. In the present study, the
subcutaneous and tail vein injections were performed without
anesthesia. At the end of the animal experiment, cervical
dislocation was used for euthanasia and death was verified by
cardiac arrest.
Statistical analysis
All data were statistically analyzed using SPSS 20.0
(IBM Corp.) and GraphPad Prism 5.0 software (GraphPad Software,
Inc.). For tumor volume, data are presented as the mean + 95%
confidence interval (CI), while the remaining data are shown as the
mean ± standard error of the mean. Statistical tests included the
Student's t-test, χ2 test, ANOVA and Dunnett's post hoc
test. P<0.05 was considered to indicate statistically
significant differences.
Results
Rap1A expression is upregulated in
ESCC
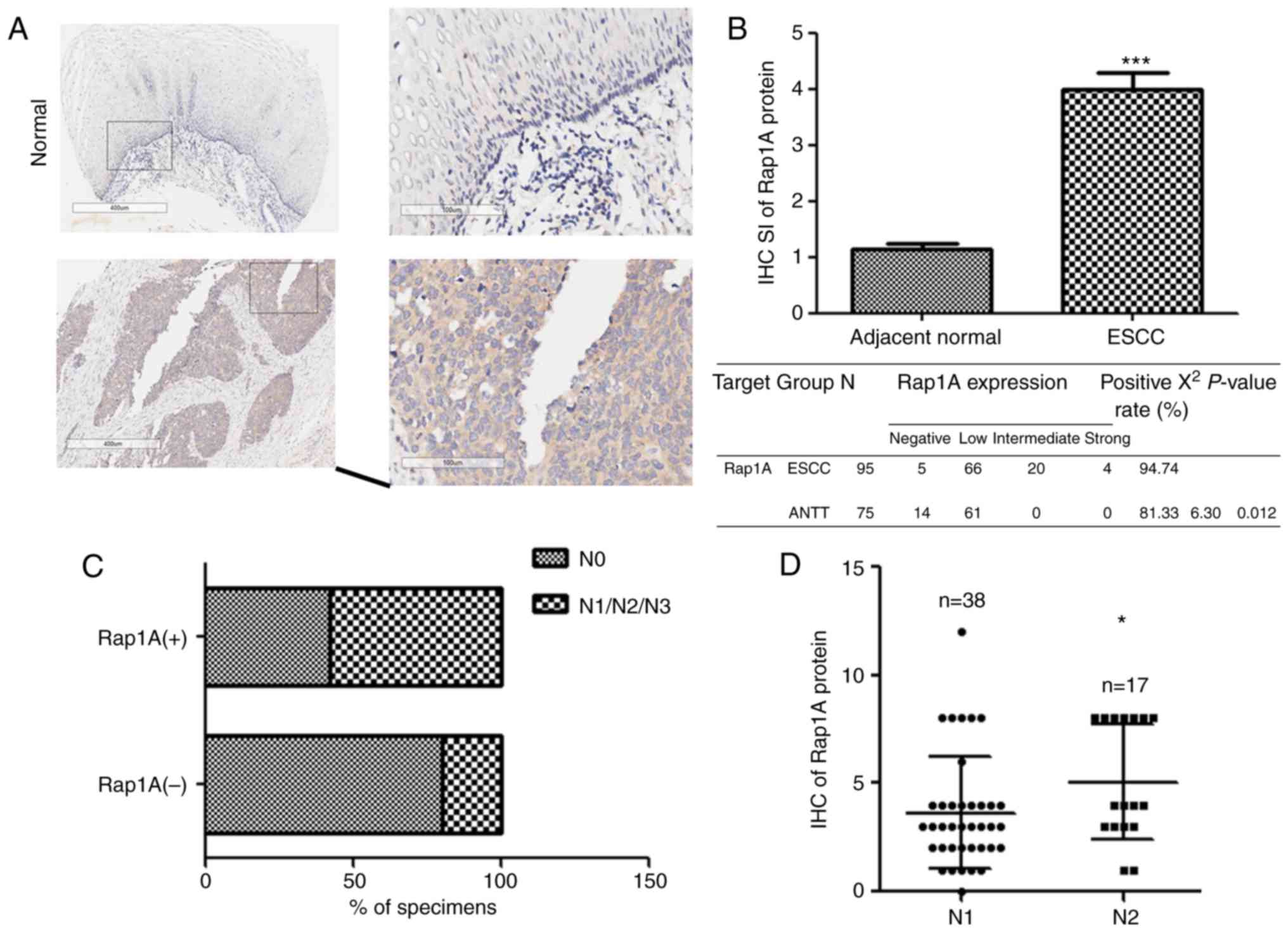
Rap1A is upregulated in cancer tissues compared with
ANTT and contributes to the progression of prostate, colorectal and
oral cavity squamous cell carcinoma (19–21). To
investigate the expression and biological role of Rap1A in human
ESCC, we examined the Rap1A expression pattern in 95 human ESCC and
75 ANTT samples. As shown in Fig. 1A and
B, Rap1A was markedly overexpressed in human ESCC at the
protein level (P<0.001). Furthermore, statistical analysis of
the ESCC samples revealed a correlation between lymph node
metastasis and Rap1A expression (Fig.
1C), i.e., Rap1A levels were markedly correlated with the N
stage of human ESCC. Moreover, Rap1A expression was higher in N2
compared with N1 stage ESCC (Fig.
1D), suggesting that Rap1A may be a potential biomarker for the
development and progression of lymph node metastasis in ESCC.
The growth-promoting effect of Rap1A
in ESCC is context-dependent
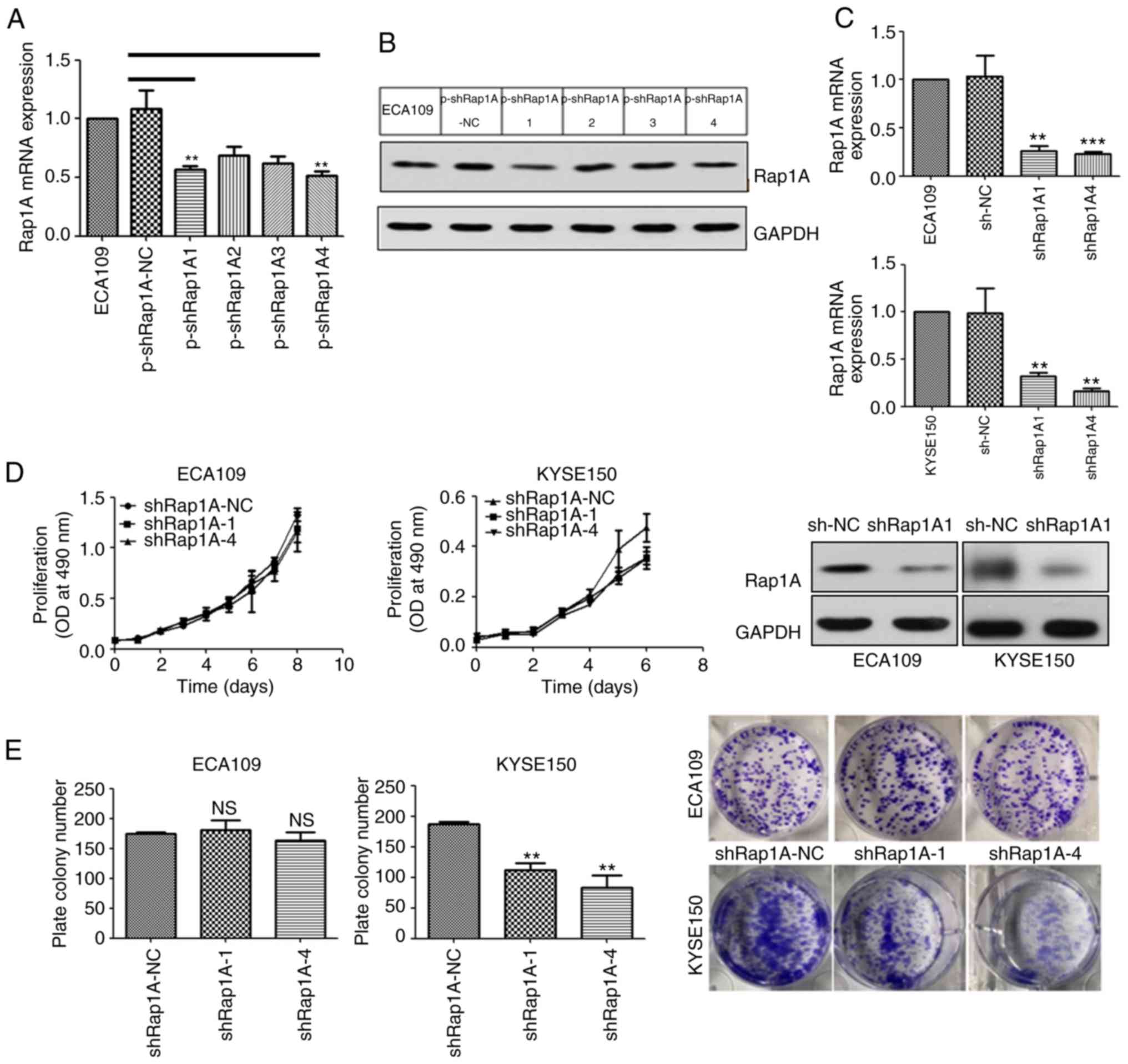
In order to elucidate the function of Rap1A in human
ESCC, four types of shRNA targeting human Rap1A mRNA were screened
using ECA109 cells (Fig. 2A and B)
and the two most efficient silencing sequences, shRap1A1 and
shRap1A4, were selected and used to establish ESCC cell lines with
stable knockdown of Rap1A (Fig.
2C).
In order to investigate the role of Rap1A in ESCC
growth, the ESCC cell lines were used to evaluate cell
proliferation. MTT analysis revealed that the downregulation of
Rap1A suppressed the proliferation of KYSE150 cells compared with
shRap1A-NC transfected cells (Fig.
2D). Consistently, colony formation assays revealed that the
number of colonies formed by KYSE150-shRap1A1 and -shRap1A4 cells
were significantly decreased compared with those formed by empty
vector-transfected cells (Fig. 2E).
However, the MTT and colony formation assays indicated that Rap1A
did not affect ECA109 cell growth (Fig.
2D and E). In summary, the growth-promoting effect of Rap1A in
ESCC is context-dependent.
Rap1A knockdown decreases the
migratory and invasive capacities of ESCC cells
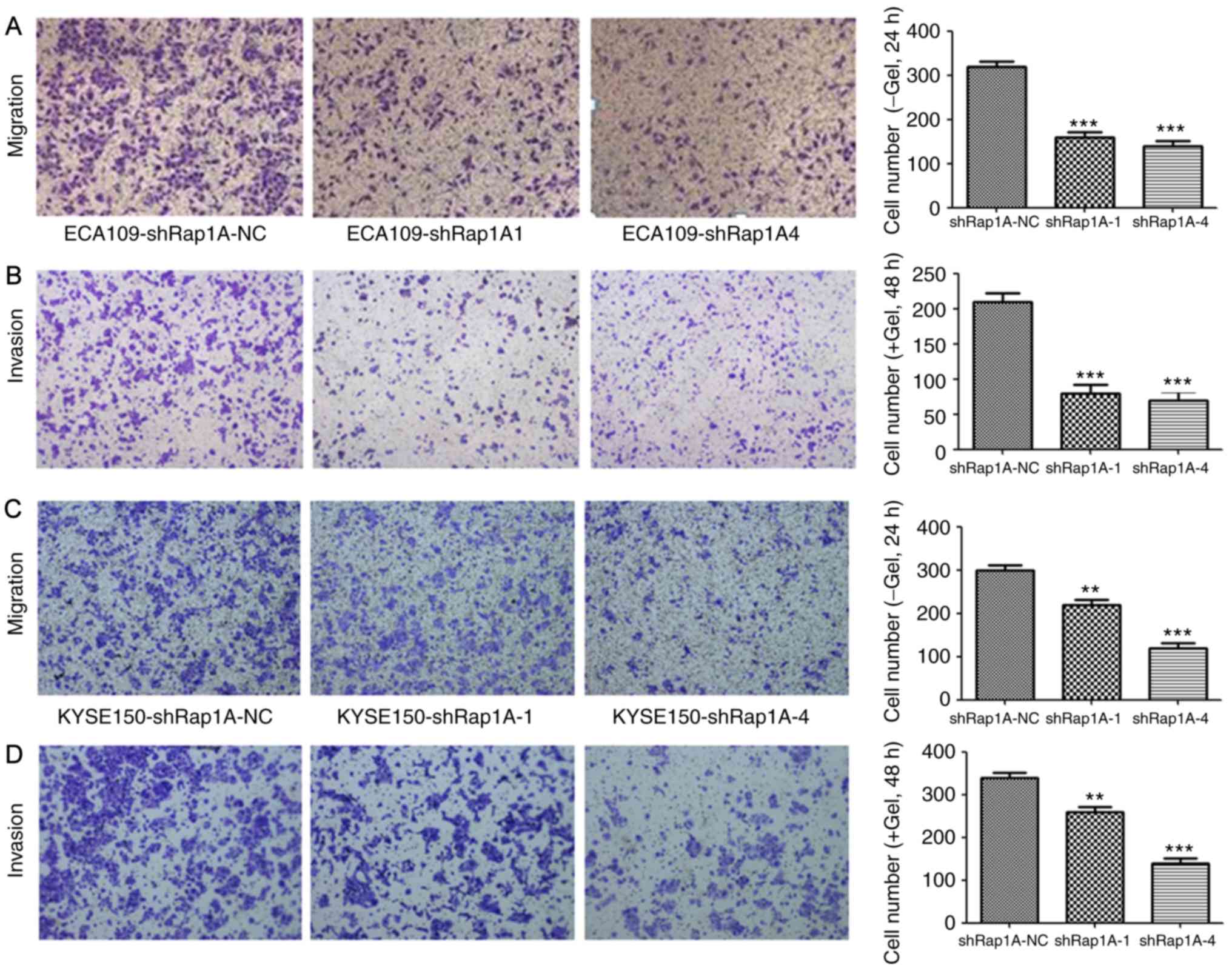
To investigate the role of Rap1A in cell migration
and invasion, a Transwell assay was performed after silencing Rap1A
expression using shRap1A to evaluate ESCC cell motility. Both cell
migration and invasion assays demonstrated that Rap1A knockdown
significantly inhibited ECA109 cell migration and invasion
(Fig. 3A and B). The same phenomenon
was also observed in KYSE150-shRap1A1 and KYSE150-shRap1A4 cells
compared with shRap1A-NC cells (Fig. 3C
and D). These in vitro results suggest that Rap1A may
promote the metastatic behavior of ESCC cells.
Rap1A may enhance EMT via AKT
signaling in ESCC cells
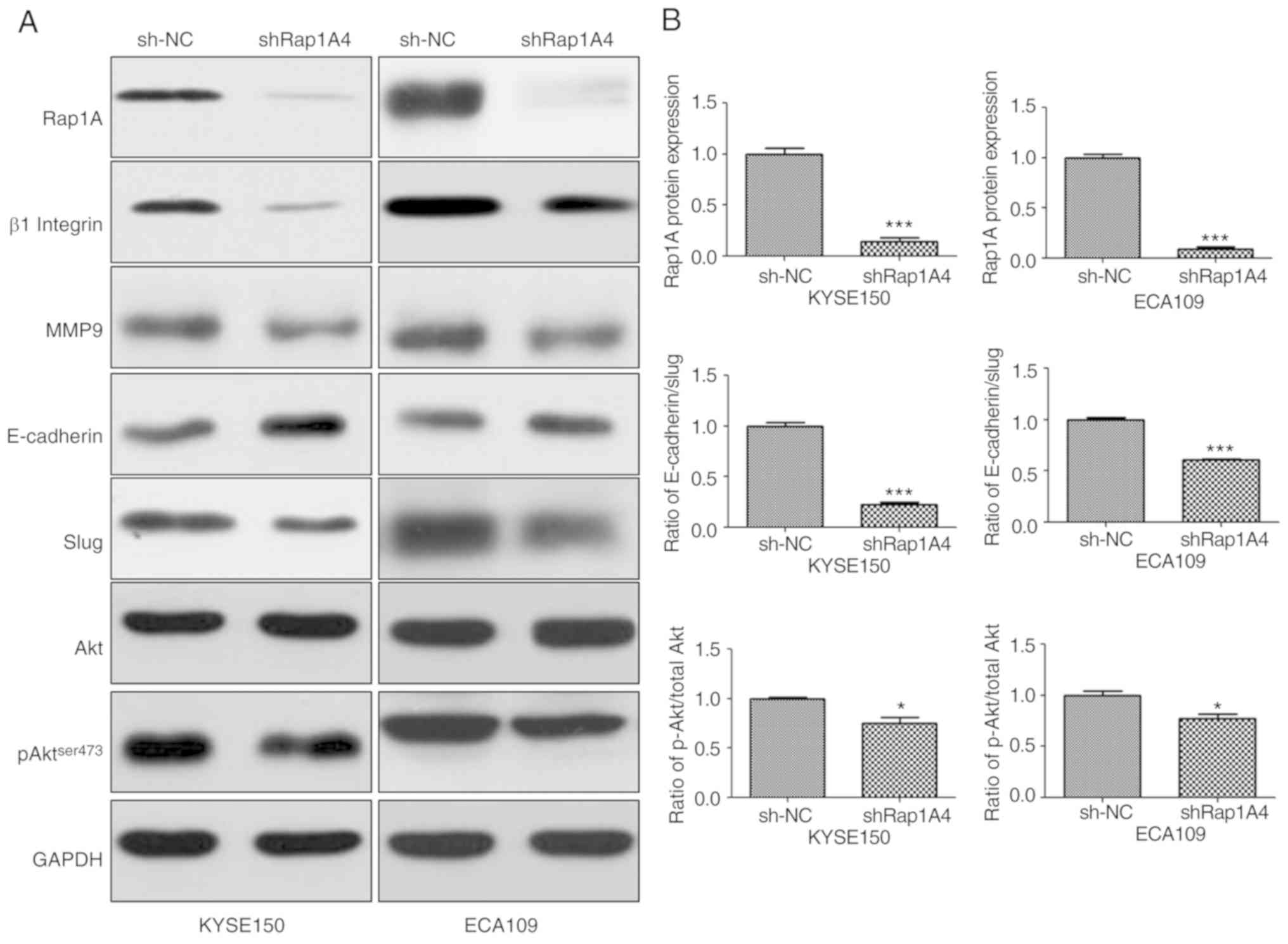
To identify the mechanism of action of Rap1A in
ESCC, the changes in AKT-, pAKTser473- and EMT-associated signaling
molecules in both ESCC cell lines were investigated following Rap1A
knockdown. β1 integrin has been reported to be stabilized by Rap1A
and enhances migration of epithelial cells; therefore, it was
included in the experiments (22). As
shown in Fig. 4, siRNA-mediated
downregulation of Rap1A resulted in the reduction of pAKT ser473
expression; furthermore, the expression of β1 integrin, EMT markers
and MMP9 were reduced compared with sh-NC. However, the reduction
of pAKT expression following Rap1A inhibition appeared to be
minimal. The difference in AKT phosphorylation between the two cell
lines was statistically significant (P<0.05). The statistical
analysis of the AKT phosphorylation level following Rap1A
inhibition was as follows: sh-NC vs. shRap1A, 1.000±0.015 vs.
0.748±0.064 in KYSE150 cells and 1.000±0.043 vs. 0.774±0.041 in
ECA109 cells, respectively; unpaired t-test was applied to estimate
the two-tailed P-values, which were 0.0188 and 0.019, respectively.
Therefore, the difference was statistically significant, although
it was minimal after Rap1A inhibition. Thus, these results indicate
that Rap1A may promote cancer cell migration and invasion via
enhancement of β1 integrin-dependent adhesion, and EMT and
MMP9-associated degradation of the extracellular matrix; AKT
signaling may participate in these processes.
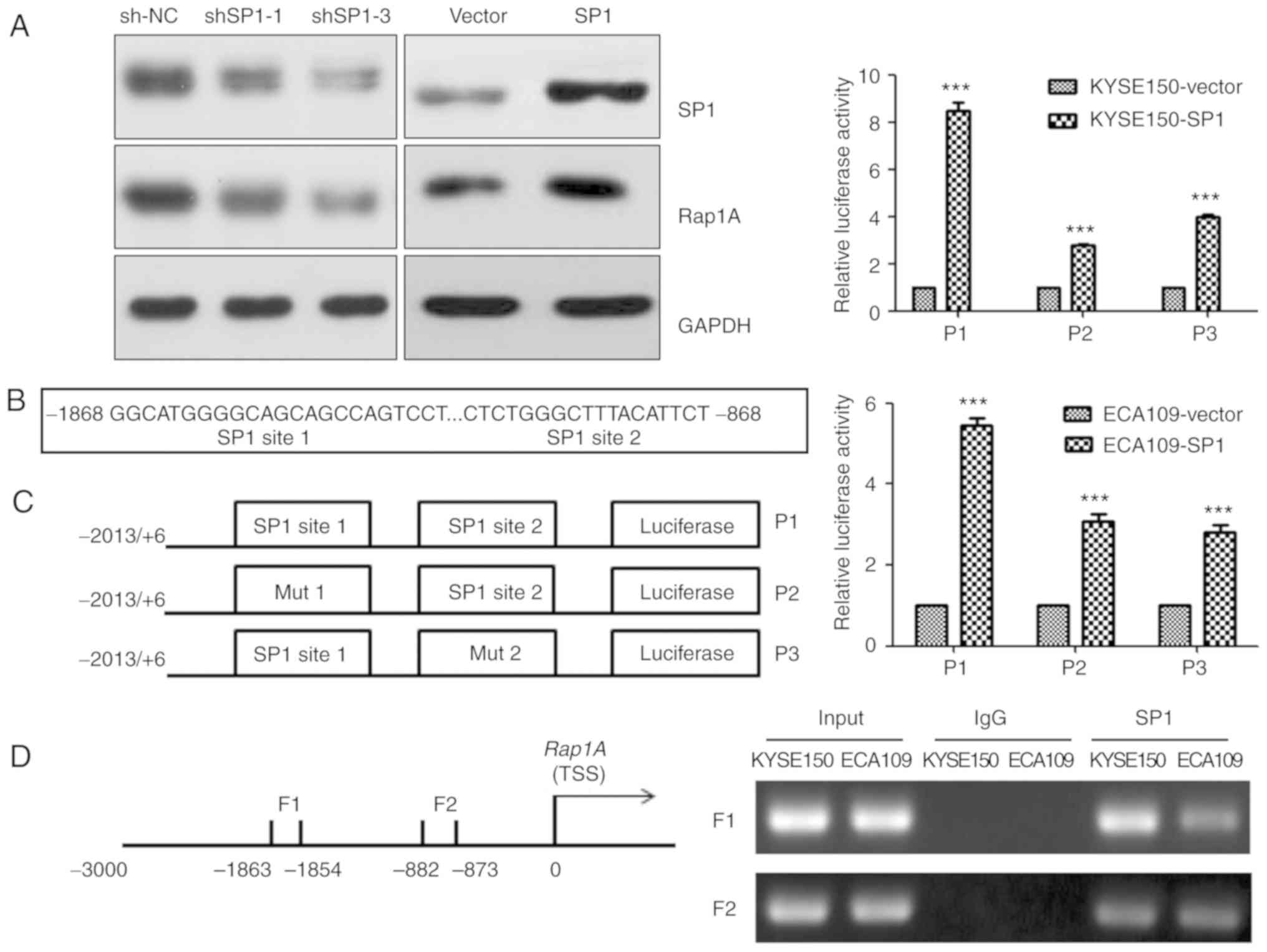
SP1 upregulates human Rap1A promoter
activity through SP1-binding sites
The transcription factor SP1 plays an important role
in ESCC (23). Consistently, we
observed that SP1 could upregulate Rap1A expression in KYSE150
cells (Fig. 5A). To explore the
molecular mechanism underlying Rap1A regulation by SP1, the CONSITE
program was used to predict the SP1-specific binding sites in the
Rap1A promoter (Fig. 5B), and then
luciferase reporter and ChIP assays were performed in both ECA109
and KYSE150 cells. Overexpressing SP1 increased the luciferase
activities of Rap1A promoter, Rap1A promoter mut1 and Rap1A
promoter mut2 (P<0.001). The Rap1A promoter exhibited the
highest luciferase activity, and SP1 overexpression in KYSE150
cells led to stronger Rap1A transactivation compared with that in
ECA109 cells (Fig. 5C). The ChIP
assay indicated that SP1 was capable of binding to the F1 (nt
−1,863 to −1,854) and F2 (nt −882 to −873) regions within the Rap1A
promoter. In addition, the ability of SP1 to bind to the F1 in
KYSE150 cells was more prominent compared with that in ECA109 cells
(Fig. 5D). These results suggest that
SP1 upregulates human Rap1A promoter activity through two
SP1-binding sites.
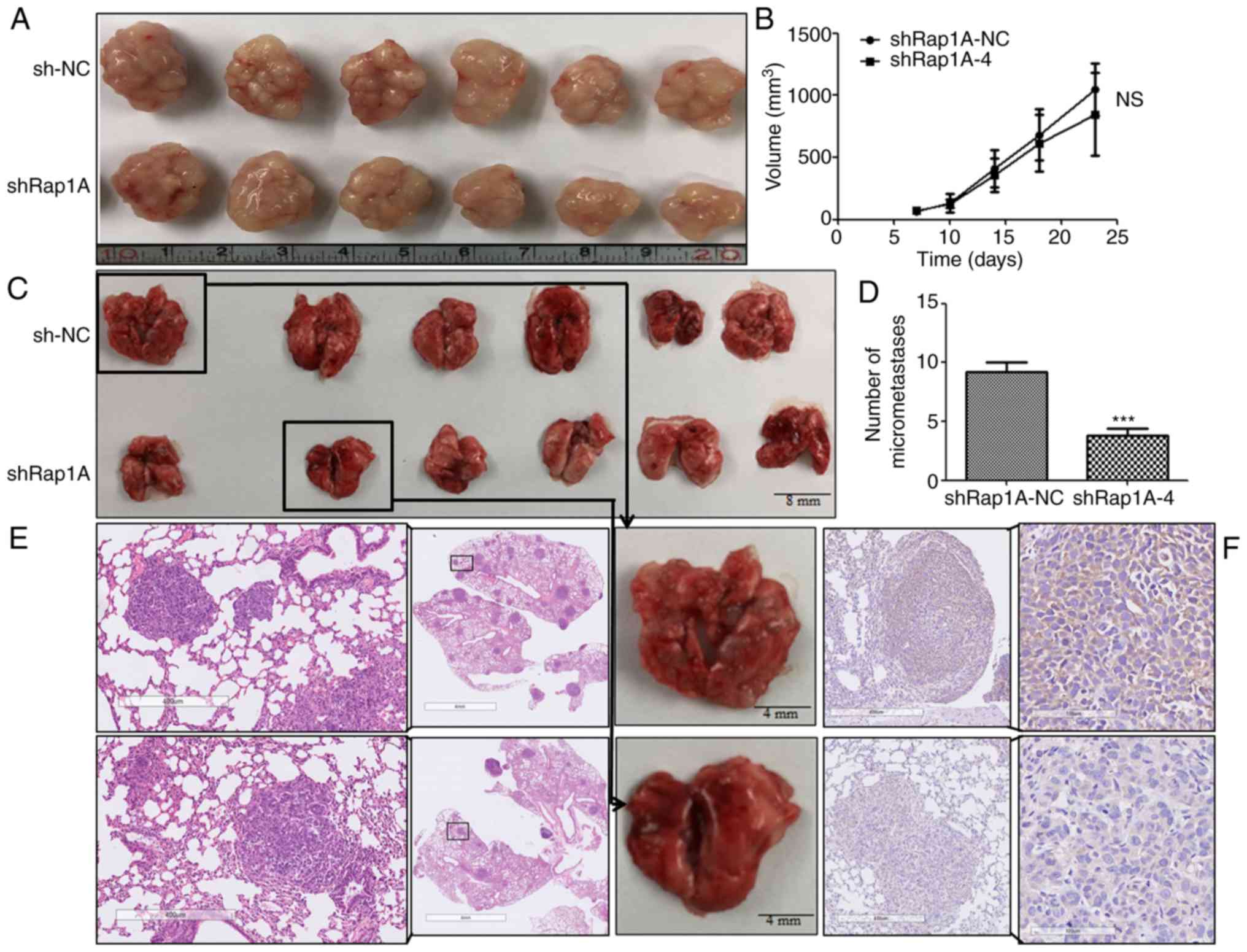
Rap1A knockdown attenuates tumor
metastasis in vivo
To determine the effect of Rap1A on ESCC
tumorigenesis and metastasis in vivo, subcutaneous tumor and
lung metastasis assays were performed in nude mice with ECA109
cells expressing shRap1A. For xenografted tumors, the results were
valid and reliable, as all mice developed a single subcutaneous
tumor (longest diameter, 16.53 mm). The results indicated that
ECA109 cells with Rap1A knockdown was associated with no
significant difference in tumor size compared with control cells
(Fig. 6A and B). In the metastasis
model (Fig. 6C-F), the results of IHC
and H&E staining indicated that downregulation of Rap1A
decreased the number of metastatic nodules in the lungs. Taken
together, these findings were consistent with the in vitro
results and indicated that Rap1A promotes ESCC metastasis in
vivo.
Discussion
The findings of the present study demonstrated the
oncogenic function of Rap1A in the progression of ESCC. Rap1A
promoted cell migration, invasiveness and metastasis, and its
growth-promoting effect was found to be context-dependent.
Moreover, Rap1A mediated EMT via AKT signaling and was regulated by
the transcription factor SP1. Thus, Rap1A may be a potential
therapeutic target for ESCC.
One of the novel findings of the present study was
the activation of AKT signaling by the Rap1A small GTPase. It is
well known that the Ras/Raf/MAPK pathway regulates a variety of
cellular functions related to tumorigenesis (24). As the closest relative of Ras, Rap1A
was previously shown to regulate MAPK/ERK signaling in solid tumors
(17,25,26). In
addition to ERK, Rap1A promotes cancer cell proliferation via AKT
signaling (27), and AKT activation
plays a crucial role in the progression of ESCC, including cancer
cell growth, migration, invasion and metastasis (11,28–30). With
both AKT2 and KRAS amplification, enhanced alterations involving
PI3K/AKT/mTOR signaling have been detected in the genomic profiling
of ESCC (31,32). Approximately 90% of tumors exhibit
some degree of EMT during tumor development (11); notably, it was herein demonstrated
that Rap1A activates AKT signaling to mediate EMT, and is regulated
by the transcription factor SP1, which is consistent with the
hypothesis that SP1 plays a key role in ESCC (23). Taken together, the findings of the
present study combined with those of previous reports further
indicated that AKT signaling may be positively regulated by Rap1A
in ESCC.
Another important finding of the present research is
that the oncogenic role of Rap1A in ESCC is context-dependent. It
was previously reported that the function of miR-196a in ESCC is
correlated with the SNPrs6573 A/C genotype in the Rap1A 3′-UTR
(33). Furthermore, ECA109 and
KYSE150 cells were found to be CC homozygotes, which interferes
with the binding of miR-196a to Rap1A 3′-UTR, resulting in a higher
constitutive expression of Rap1A (33). In the present research, silencing
Rap1A reversed the malignant phenotype of both cell lines, with the
exception of ECA109 cell proliferation. It is important to note
that both cell lines were isolated from the resected specimens of
female patients with poorly differentiated ESCC (34,35). Thus,
the impact of Rap1A on ESCC cell growth is context-dependent and
promotes tumor metastasis, which further supported previous
reports.
Moreover, the reason why Rap1A is highly expressed
in esophageal cancer was further investigated. In view of the
important role of SP1 (23), it was
hypothesized that the overexpression of Rap1A may be the result of
the transcriptional activation of SP1.
In summary, the present study demonstrated that
Rap1A promoted tumor metastasis, possibly via AKT signaling,
integrin-mediated adhesion (27,36), MMP9
and EMT; furthermore, the growth-promoting effects of Rap1A in ESCC
were associated with tumor heterogeneity, which required further
research to elucidate the underlying mechanism. Considering the
known amplification of the K-Ras gene in human esophageal cancer
(31,32,37), the
expression of Rap1A may be an ESCC marker, and inhibitors of Rap1A
may be of therapeutic value. In addition, mTOR plays a key role in
regulating several fundamental cellular processes, and its
hyperactivation is correlated with a number of human cancers
(38). ESCC without lymph node
metastasis may be effectively removed by endoscopic resection and
can have a good prognosis (39,40). Thus,
further study investigating the specific mechanism involved in
lymphatic metastasis is crucial for the treatment of ESCC.
There were certain limitations to the present study.
It was demonstrated that Rap1A was upregulated in human ESCC
tissues compared with ANTT, but there is lack of data on human
normal esophageal cells used as a negative control. The results
showing that Rap1A promoted ESCC cell metastasis and that its
growth-promoting effect was context-dependent relied on only two
cell lines; thus, experiments on more cell lines are warranted to
verify these findings. Furthermore, observation of the
morphological changes was not performed, indicating a lack of
effective verification regarding the presence of EMT. MMP2 is also
a typical enzyme that degrades extracellular matrix. Thus, the
absence of data on MMP2 is another limitation of the present study.
It is also recommended to further investigate the roles of AKT and
its associated mTOR pathway in ESCC. Despite these limitations,
however, combining previous results (33) with the present research confirmed
Rap1A as an oncogenic molecule in the progression of ESCC.
Supplementary Material
Supporting Data
Acknowledgements
The authors would like to thank Dr Jia Cao
(Department of Digestive Endoscopy, Shanghai East Hospital, Tongji
University, China), for providing valuable suggestions on revising
the manuscript.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81570595).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
The study was conceived and designed by QL, AX, TC
and MX. Study and clinical materials were provided by MX, PZ and
LY. The experiments were performed by QL, AX, YC and HL. The
datasets were analyzed by QL, AX and YC. The manuscript was written
by QL, AX and TC. All authors have read and approved the final
version of this manuscript for publication.
Ethics approval and consent to
participate
Written informed consent was obtained from all
patients prior to sample collection. The animal experiment were
approved by Medical Ethics Committee of Fudan University
(201906004Z) and all experiments on nude mice were performed
according to the biosafety and bioethics guidelines.
Patient consent for publication
Not applicable.
Competing interests
All authors declare that they have no competing
interests.
References
|
1
|
Chen W, Zheng R, Baade PD, Zhang S, Zeng
H, Bray F, Jemal A, Yu XQ and He J: Cancer statistics in China,
2015. CA Cancer J Clin. 66:115–132. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kuwano H, Nishimura Y, Oyama T, Kato H,
Kitagawa Y, Kusano M, Shimada H, Takiuchi H, Toh Y, Doki Y, et al:
Guidelines for diagnosis and treatment of carcinoma of the
esophagus April 2012 edited by the Japan esophageal society.
Esophagus. 12:1–30. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pennathur A, Gibson MK, Jobe BA and
Luketich JD: Oesophageal carcinoma. Lancet. 381:400–412. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lagergren J, Smyth E, Cunningham D and
Lagergren P: Oesophageal cancer. Lancet. 390:2383–2396. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nitulescu GM, Van De Venter M, Nitulescu
G, Ungurianu A, Juzenas P, Peng Q, Olaru OT, Grădinaru D, Tsatsakis
A, Tsoukalas D, et al: The AKT pathway in oncology therapy and
beyond (Review). Int J Oncol. 53:2319–2331. 2018.PubMed/NCBI
|
|
6
|
Guanen Q, Junjie S, Baolin W, Chaoyang W,
Yajuan Y, Jing L, Junpeng L, Gaili N, Zhongping W and Jun W:
MiR-214 promotes cell metastasis and inhibits apoptosis of
esophageal squamous cell carcinoma via PI3K/AKT/mTOR signaling
pathway. Biomed Pharmacother. 105:350–361. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao H, Chen G, Ye L, Yu H, Li S and Jiang
WG: DOK7V1 influences the malignant phenotype of lung cancer cells
through PI3K/AKT/mTOR and FAK/paxillin signaling pathways. Int J
Oncol. 54:381–389. 2019.PubMed/NCBI
|
|
8
|
Zhao J, Li Z, Chen Y, Zhang S, Guo L, Gao
B, Jiang Y, Tian W, Hao S and Zhang X: MicroRNA-766 inhibits
papillary thyroid cancer progression by directly targeting insulin
receptor substrate 2 and regulating the PI3K/AKT pathway. Int J
Oncol. 54:315–325. 2019.PubMed/NCBI
|
|
9
|
Liu SQ, Xu CY, Wu WH, Fu ZH, He SW, Qin MB
and Huang JA: Sphingosine kinase 1 promotes the metastasis of
colorectal cancer by inducing the epithelial-mesenchymal transition
mediated by the FAK/AKT/MMPs axis. Int J Oncol. 54:41–52.
2019.PubMed/NCBI
|
|
10
|
Dornan GL and Burke JE: Molecular
mechanisms of human disease mediated by oncogenic and primary
immunodeficiency mutations in Class IA phosphoinositide 3-kinases.
Front Immunol. 9:5752018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jia Y, Xiao Z, Gongsun X, Xin Z, Shang B,
Chen G, Wang Z and Jiang W: CEP55 promotes the proliferation,
migration and invasion of esophageal squamous cell carcinoma via
the PI3K/AKT pathway. Onco Targets Ther. 11:4221–4232. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Caron E: Cellular functions of the Rap1
GTP-binding protein: A pattern emerges. J Cell Sci. 116:435–440.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Khanna A, Lotfi P, Chavan AJ, Montaño NM,
Bolourani P, Weeks G, Shen Z, Briggs SP, Pots H, Van Haastert PJ,
et al: The small GTPases Ras and Rap1 bind to and control TORC2
activity. Sci Rep. 6:258232016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sayyah J, Bartakova A, Nogal N, Quilliam
LA, Stupack DG and Brown JH: The Ras-related protein, Rap1A,
mediates thrombinstimulated, integrin-dependent glioblastoma cell
proliferation and tumor growth. J Biol Chem. 289:17689–17698. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Grolez GP and Gkika D: TRPM8 puts the
chill on prostate cancer. Pharmaceuticals (Basel). 9(pii): E442016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Genova T, Grolez GP, Camillo C, Bernardini
M, Bokhobza A, Richard E, Scianna M, Lemonnier L, Valdembri D,
Munaron L, et al: TRPM8 inhibits endothelial cell migration via a
non-channel function by trapping the small GTPase Rap1. J Cell
Biol. 216:2107–2130. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lu L, Wang J, Wu Y, Wan P and Yang G:
Rap1A promotes ovarian cancer metastasis via activation of ERK/p38
and notch signaling. Cancer Med. 5:3544–3554. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rice TW, Ishwaran H, Ferguson MK,
Blackstone EH and Goldstraw P: Cancer of the esophagus and
esophagogastric junction: An eighth edition staging primer. J
Thorac Oncol. 12:36–42. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiang J, Bian C, Wang H, Huang S and Wu D:
MiR-203 down-regulates Rap1A and suppresses cell proliferation,
adhesion and invasion in prostate cancer. J Exp Clin Cancer Res.
34:82015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Liu L, Yan X, Wu D, Yang Y, Li M, Su Y,
Yang W, Shan Z, Gao Y and Jin Z: High expression of Ras-related
protein 1A promotes an aggressive phenotype in colorectal cancer
via PTEN/FOXO3/CCND1 pathway. J Exp Clin Cancer Res. 37:1782018.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen CH, Chuang HC, Huang CC, Fang FM,
Huang HY, Tsai HT, Su LJ, Shiu LY, Leu S and Chien CY:
Overexpression of Rap-1A indicates a poor prognosis for oral cavity
squamous cell carcinoma and promotes tumor cell invasion via
Aurora-A modulation. Am J Pathol. 182:516–528. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
To JY and Smrcka AV: Activated
heterotrimeric G protein αi subunits inhibit Rap-dependent cell
adhesion and promote cell migration. J Biol Chem. 293:1570–1578.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xiang J, Zang W, Che J, Chen K and Hang J:
Regulation network analysis in the esophageal squamous cell
carcinoma. Eur Rev Med Pharmacol Sci. 16:2051–2056. 2012.PubMed/NCBI
|
|
24
|
Martinelli E, Morgillo F, Troiani T and
Ciardiello F: Cancer resistance to therapies against the
EGFR-RAS-RAF pathway: The role of MEK. Cancer Treat Rev. 53:61–69.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang T, Jiang K, Zhu X, Zhao G, Wu H,
Deng G and Qiu C: MiR-433 inhibits breast cancer cell growth via
the MAPK signaling pathway by targeting Rap1A. Int J Biol Sci.
14:622–632. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rodríguez CI, Castro-Pérez E, Prabhakar K,
Block L, Longley BJ, Wisinski JA, Kimple ME and Setaluri V:
EPAC-RAP1 axis-mediated switch in the response of primary and
metastatic melanoma to cyclic AMP. Mol Cancer Res. 15:1792–1802.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Park JY and Juhnn YS: cAMP signaling
increases histone deacetylase 8 expression via the Epac2-Rap1A-AKT
pathway in H1299 lung cancer cells, histone deacetylases and
cancer. Exp Mol Med. 49:e2972017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Liu M, Hu Y, Zhang MF, Luo KJ, Xie XY, Wen
J, Fu JH and Yang H: MMP1 promotes tumor growth and metastasis in
esophageal squamous cell carcinoma. Cancer Lett. 377:97–104. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang J, Kong R and Sun L: Silencing of
Rab3D suppresses the proliferation and invasion of esophageal
squamous cell carcinoma cells. Biomed Pharmacother. 91:402–407.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kong KL, Kwong DL, Chan TH, Law SY, Chen
L, Li Y, Qin YR and Guan XY: MicroRNA-375 inhibits tumour growth
and metastasis in oesophageal squamous cell carcinoma through
repressing insulin-like growth factor 1 receptor. Gut. 61:33–42.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang JW and Choi YL: Genomic profiling of
esophageal squamous cell carcinoma (ESCC)-Basis for precision
medicine. Pathol Res Pract. 213:836–841. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang K, Johnson A, Ali SM, Klempner SJ,
Bekaii-Saab T, Vacirca JL, Khaira D, Yelensky R, Chmielecki J,
Elvin JA, et al: Comprehensive genomic profiling of advanced
esophageal squamous cell carcinomas and esophageal adenocarcinomas
reveals similarities and differences. Oncologist. 20:1132–1139.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wang K, Li J, Guo H, Xu X, Xiong G, Guan
X, Liu B, Li J, Chen X, Yang K, et al: MiR-196a binding-site SNP
regulates Rap1A expression contributing to esophageal squamous cell
carcinoma risk and metastasis. Carcinogenesis. 33:2147–2154. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Shimada Y, Imamura M, Wagata T, Yamaguchi
N and Tobe T: Characterization of 21 newly established esophageal
cancer cell lines. Cancer. 69:277–284. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xiao F, Wang X, Wang M, Guan X and Wu M:
Molecular cytogenetic study on four human esophageal cancer cell
lines. Zhonghua Yi Xue Yi Chuan Xue Za Zhi. 15:75–77. 1998.(In
Chinese). PubMed/NCBI
|
|
36
|
Tsygankova OM, Ma C, Tang W, Korch C,
Feldman MD, Lv Y, Brose MS and Meinkoth JL: Downregulation of
Rap1GAP in human tumor cells alters cell matrix and cell cell
adhesion. Mol Cell Biol. 30:3262–3274. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Salem ME, Puccini A, Xiu J, Raghavan D,
Lenz HJ, Korn WM, Shields AF, Philip PA, Marshall JL and Goldberg
RM: Comparative molecular analyses of esophageal squamous cell
carcinoma, esophageal adenocarcinoma, and gastric adenocarcinoma.
Oncologist. 23:1319–1327. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Saxton RA and Sabatini DM: mTOR signaling
in growth, metabolism, and disease. Cell. 168:960–976. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Aadam AA and Abe S: Endoscopic submucosal
dissection for superficial esophageal cancer. Dis Esophagus.
31:2018. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Zhang YQ, Chen T, Zhang C, Li QL, Chen WF,
Yao LQ, Zhou PH and Xu MD: Endoscopic submucosal dissection for
superficial proximal esophageal neoplasia is highly successful. Ann
Surg. 266:995–999. 2017. View Article : Google Scholar : PubMed/NCBI
|